



-
樟树(Cinnamomum camphora(L.)Presl)又名香樟树,属樟科(Lauraceae),樟属(Cinnamomum Trew)常绿乔木,是良好的林木、绿化树、行道树和庭荫树,且具有较高的药用价值[1-2]。随着人们环保意识的增强和城市绿化工程的加快,越来越多的樟树进入城市绿化的行列,但是城市绿化移植樟树导致樟树病害的发生也日趋严重[3-4],其中,樟树溃疡病是樟树上发病最严重的病害之一,该病害主要危害主干,尤以移植后的苗木受害较重,发病率一般为15%~50%,发病严重时病株率达100%,死亡率达88%,极大地影响了樟树造林成活率和樟树的长势[5]。
樟树溃疡病于1995年由郭立中等[5]通过调查与研究湖南地区樟树病害的发生后首次报道,并且将其病原菌的有性型鉴定为囊孢壳菌(Physalospora sp.),无性型鉴定为大茎点霉菌(Macrophoma sp.)。2005年,邓先琼等[6]对该病的病原菌种类进行了再鉴定,根据樟树溃疡病菌子囊、子囊孢子、分生孢子、分生孢子梗的形态特征,将该病害的病原菌鉴定为葡萄座腔菌(Botryosphaeria dothidea Ces et de Not.)。赵桂华等[7]通过对樟树溃疡病的病原菌分离培养、致病性测定及观察菌丝特征和分生孢子的萌发状况,证实了江苏地区的樟树溃疡病由真菌耧斗大茎点霉菌(Macrophoma aquilegiae)引起。然而,前期的研究范围主要集中于湖南和江苏地区,且关于该病菌与其近缘真菌的转录间隔区(ITS)序列差异、系统进化和分类地位等方面缺乏系统研究。因此,本研究通过采自重庆地区的樟树溃疡病的发病组织进行病原菌的分离纯化,根据病原菌的形态学特征、致病性及ITS序列、β-微管蛋白基因(β-tubulin)序列和延伸因子基因(EF1-α)序列的同源性分析,对引起重庆樟树溃疡病的病原菌进行鉴定与分析,以期为系统研究樟树溃疡病及对其进行防治奠定理论基础。
-
2017年11月至2018年7月,在重庆渝中区、北碚区和涪陵区采集患有溃疡病的樟树树皮和枝条,分别装入样品袋中进行编号。参照方中达[8]有关病原真菌的组织分离方法,对采集的樟树溃疡病样本进行病原菌的分离,并采用单菌丝法[9]纯化菌株,纯化的菌株保存至-80℃的甘油中。
-
将纯化菌株活化后,接种至新鲜的PDA平板上,置于25℃培养箱中,黑暗培养。观察菌落颜色、形态及气生菌丝等特征。分生孢子的诱导采用黑光灯(365 nm)诱导法[9],在显微镜下观察孢子的形态特征。
-
将菌株接种在铺有灭菌玻璃纸的PDA平板上,培养3 d后刮取菌丝,在研钵中液氮速冻后研磨成粉,采用CTAB法[10]提取其基因组DNA,分别对ITS、β-tubulin和EF1-α基因序列进行PCR扩增。引物序列参见文献[11-13],由北京六和华大生物科技有限公司合成。ITS序列扩增程序为:94℃预变性4 min;94℃变性45 s;58℃退火45 s;72℃延伸45 s,共30个循环。β-tubulin和EF1-α基因序列扩增程序除退火温度为60℃外,其余与上述相同。将纯化的扩增产物连接到载体pMD18-T,转化至大肠杆菌(Escherichia coli)DH5α感受态细胞,之后,将经PCR验证为阳性的菌液送至北京六和华大生物科技有限公司测序。
-
将测序所得序列在NCBI上进行检索比对,并下载本属及参考属真菌分离物的序列,与测定的病原菌相关基因序列进行比较分析。利用Clustalx W(1.83)和MEGA 6软件对所测菌株和参考菌株的核苷酸序列进行多重比对和遗传进化分析,采用邻接法(Neighbor-Joining)构建系统进化树(重复次数为1 000)。
-
剪取粗细均匀的1年生樟树枝条,无菌水洗净晾干,经75%酒精表面消毒后剪成长短一致的小段,两端用石蜡封口。用打孔器在枝干中间打孔,将菌丝块接种在伤口处,以空白培养基为对照,每个菌种设置6次重复。将蘸有无菌水的灭菌棉花覆盖在菌丝块上面保湿,然后放置在25℃的恒温条件下进行培养,接种2 d去除菌丝块和棉花,接种4 d调查、记录枝条发病状况,并测量病斑长度。之后,切取发病枝条的病健交界处进行再分离,将获得的菌株和原始接种菌株进行菌落形态和ITS序列的比较。
-
樟树溃疡病可危害不同树龄樟树的主干、主枝和树梢,树干受害后皮层坏死,后期皮层纵横开裂并翘起(图 1A)。樟树枝梢被病菌侵染后易形成黑褐色病斑,随后病斑以上枝条变黑褐色坏死,形成枝枯(图 1B、D)。在幼龄树上,大部分病斑位于主干的中部和下部,在主干与侧枝的分蘖处也容易产生病斑,被为害的主干部分形成凹陷的U形溃疡病灶(图 1C),将发病部分的主干横切,发现患病部分已经深入到了树干的木质部(图 1E),在病患交接的地方为红棕色的病斑,患病严重的部位则出现黑色腐烂的病斑(图 1C、E、F)。

图 1 樟树溃疡病的症状
Figure 1. Typical canker symptoms of camphor tree
-
从重庆渝中区、北碚区和涪陵区共采集到樟树溃疡病样品62份,分离得到48株具有葡萄座腔菌属真菌特征的菌株,单菌丝纯化后获得68株纯化菌株。根据纯化菌株在PDA培养基上的培养特性及分生孢子特征,可分为3种类型,分别称为Ⅰ型、Ⅱ型和Ⅲ型,其中,Ⅰ型菌株有43份,Ⅱ型菌株有16份,Ⅲ型菌株有9份。从3种类型的菌株中选取14株代表菌株进行形态学分析(表 1),包括分离自渝中区的3份菌株(YZS7-6a、YZS8-1a和YZS8-1b)、北碚区的5份菌株(YZS9-1a、YZS9-1b、YZS9-1c、YZS9-1d和YZS11-1a)及涪陵区的6份菌株(YZS1-1a、YZS1-2a、YZS1-6a、YZS3-2a、YZS3-3a和YZS5-1a)。
表 1 樟树溃疡病菌不同类型菌株的分生孢子大小
Table 1. Conidial dimensions of camphor tree canker strains
菌株Strains 分生孢子大小Conidial dimensions 长×宽Length×width/μm (长×宽)平均值±标准误Mean of Length×width±SD/μm 长/宽L/W Ⅰ型 YZS1-2a (18.36~32.23)×(5.55~12.35) (25.64±2.44)×(8.53±1.51) 3.08±0.54 YZS11-1a (19.21~26.21)×(5.42~10.40) (22.36±2.14)×(9.16±0.97) 3.04±0.51 YZS5-1a (21.36~32.24)×(8.55~14.56) (28.16±2.44)×(11.22±1.44) 2.54±0.35 Ⅱ型 YZS9-1a (21.82~32.23)×(9.03~17.96) (26.33±2.35)×(11.79±1.62) 2.48±0.49 YZS9-1b (18.23~29.66)×(7.50~13.58) (24.01±2.17)×(10.40±1.31) 2.34±0.29 YZS1-1a - - - YZS3-2a - - - YZS9-1c - - - YZS9-1d - - - Ⅲ型 YZS1-6a (19.12~26.51)×(10.54~14.86) (22.43±1.45)×(12.62±1.13) 1.79±0.15 YZS3-3a (19.36~26.23)×(9.85~15.48) (22.96±1.53)×(13.27±1.13) 1.74±0.21 YZS8-1a (21.86~29.58)×(12.69~18.19) (26.50±1.66)×(15.32±1.19) 1.74±0.15 YZS8-1b (22.23~33.61)×(12.65~19.97) (28.45±2.29)×(16.41±1.54) 1.74±0.14 YZS7-6a - - - 注:每菌株分别统计50个分生孢子;“-”观察期内未产生分生孢子(30 d)。
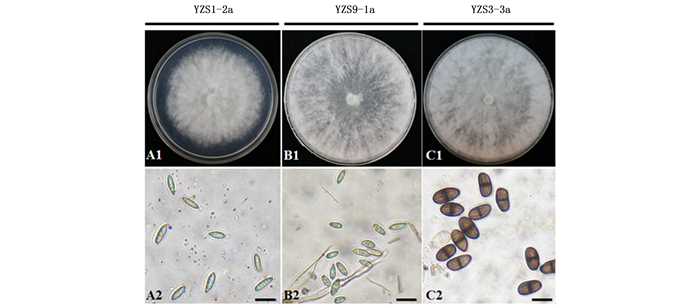
Note: The length and width of 50 conidia were measured from each selected strains. “-” No conidia were observed during the observation period (30 days).Ⅰ型代表菌株(3株)在PDA平板培养2 d,菌落为白色,边缘不整齐,生长缓慢,气生菌丝较短且很密集,呈棉絮状(图 2 A1);4 d后菌落长满整个培养皿,气生菌丝逐渐变成灰白色,菌落背面为灰绿色,后期变成黑色。分生孢子器黑色,多为单生,菌株YZS1-2a和YZS11-1a能够泌出乳白色的分生孢子角。显微镜下可观察到分生孢子为梭形、透明的单细胞(图 2 A2),分生孢子的长度为18.36~32.24 μm,宽度为5.42~14.56 μm,平均长宽比为2.80(表 1)。这类型菌株的菌落特征和分生孢子形态与B. dothidea的代表菌株相似[14-15]。
图 2 樟树溃疡病菌菌落特征及分生孢子形态
Figure 2. Colonial and conidial morphologies of camphor tree canker strains
Ⅱ型代表菌株(6株)的菌落初期为白色,气生菌丝呈束状直立生长,气生性较强但生长稀疏,培养至2 d菌落已长满整个培养皿,菌丝尖端可接触至培养皿盖(图 2 B1);4 d后菌落逐渐变成灰白色,背面呈灰棕色,最后变成黑色。分生孢子器黑色,多分布在培养皿的边缘,菌株YZS9-1a和YZS9-1b可泌出乳白色的分生孢子角,显微镜下可观察到分生孢子为椭圆形、无隔膜、透明的单细胞(图 2 B2),分生孢子长度为18.23~32.23 μm,宽度为7.50~17.96 μm,平均长宽比为2.41(表 1)。其他4株代表菌株在培养30 d时能够观察到黑色的孢子器,但镜检时均未观察到分生孢子。这类型菌株的菌落特征和分生孢子形态与B. parva代表菌株相似[15]。
Ⅲ型代表菌株(5株)的菌落初期为白色,气生菌丝呈放射状生长,气生性较上述2种类型菌株强,且很浓密,菌落生长最快,培养2 d菌落已长满整个培养皿(图 2 C1);培养4 d后,菌落正面灰白色,背面呈灰黑色,气生菌丝呈棉絮状。分生孢子器位于培养基表面,且为聚生,其中,4株代表菌株能够泌出乳白色的分生孢子角。显微镜下发现有2种类型的分生孢子,初期无色透明、无隔膜,成熟后则变成深褐色,且孢子中间形成一条横隔(图 2 C2),分生孢子长度为19.12~33.61 μm,宽度为9.85~19.97 μm,平均长宽比为1.75(表 1)。代表菌株YZS7-6a在培养30 d时镜检时均未观察到成熟的分生孢子。该类型菌株的菌落特征和分生孢子形态与B. rhodina代表菌株相似[15]。
-
对选取的14株代表菌株进行ITS、β-tubulin和EF1-α序列扩增,3个基因分别扩增到长度约550、400、200 bp的特异性片段,与预期目标片段大小一致。将获得的ITS序列在NCBI上Blastn比对,结果显示:菌株YZS1-2a、YZS5-1a、YZS11-1a与B. dothidea(无性型:七叶树壳梭孢Fusicoccum aesculi)的代表菌株CMW8000相似性为100%;菌株YZS1-1a、YZS3-2a、YZS9-1a、YZS9-1b、YZS9-1c、YZS9-1d与B. parva(无性型:小新壳梭孢Neofusicoccum parvum)的代表菌株CMW9081相似性为99%;菌株YZS1-6a、YZS3-3a、YZS7-6a、YZS8-1a、YZS8-1b与B. rhodina(无性型:可可毛色二孢Lasiodiplodia theobromae)的代表菌株CMW9074相似性为99%。
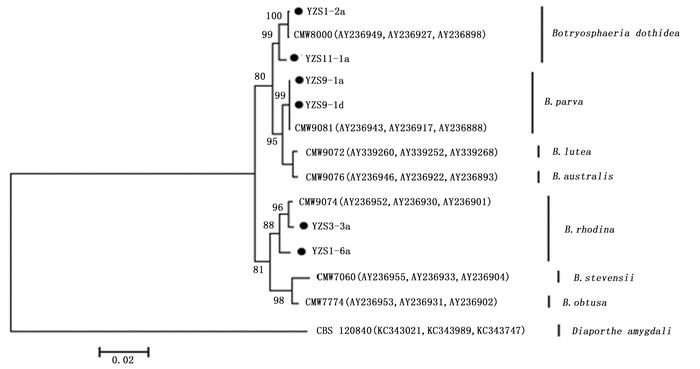
将来源于重庆市不同地区的14份樟树溃疡病菌株以及GenBank上登录的不同国家地区及不同寄主的葡萄座腔菌属菌株的ITS核苷酸序列进行系统进化树分析,结果表明:14份樟树溃疡病菌株聚集为3个不同的分支(图 3),其中,3份形态特征为Ⅰ型的菌株和来源于瑞士李树的菌株CMW8000(登录号AY236949)聚在同一组群;6份形态特征为Ⅱ型的菌株和来源于新西兰杨树的菌株CMW9081(登录号AY236943)聚在同一组群;5份形态特征为Ⅲ型的菌株和来源于墨西哥松树的菌株CMW9074(登录号AY236952)聚在同一组群。此外,将部分代表菌株的ITS、β-tubulin和EF1-α序列组合构建进化树,获得的进化树与根据ITS序列所构建的进化树结果一致(图 4)。因此,本研究中获得的樟树溃疡病菌为3种不同类型的葡萄座腔菌属真菌,分别是B. dothidea、B. parva和B. rhodina。
图 3 基于不同葡萄座腔菌属真菌的ITS序列构建的系统进化树
Figure 3. Phylogenetic tree of Botryosphaeria species associated with camphor canker disease in Chongqing Municipality based on the Neighbour-Joining analysis of ITS nucleotide sequences
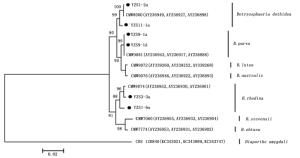
图 4 基于不同葡萄座腔菌属真菌的ITS、β-tubulin和EF1-α序列构建的系统进化树
Figure 4. Phylogenetic tree of Botryosphaeria species associated with camphor canker disease in Chongqing Municipality based on the Neighbour-Joining analysis of combined ITS、β-tubulin and EF1-α nucleotide sequences
-
将选取的代表菌株接种樟树1年生离体枝条,2 d开始发病,4 d时枝条上形成棕色或黑色的溃疡状病斑。病斑初期棕色,后期变成深褐色,最终变成黑色,溃疡部位软化腐烂并凹陷(图 5A),接种无菌PDA的枝条对照均未发病。对发病枝条进行再分离,所得病原菌与原接种病原菌形态特征一致,且ITS序列相同,符合科赫氏法则,说明分离得到的菌株均为樟树溃疡病的病原菌。采用SPSS软件对不同类型樟树溃疡病菌株接种樟树1年生离体枝条后4 d的病斑长度进行显著性差异分析,结果显示:B. parva菌株的致病性最强,B. rhodina菌株的致病性次之,B. dothidea菌株的致病性最弱,相同种的菌株之间致病性差异不显著(图 5B)。

图 5 不同菌株接种樟树离体枝条症状及致病性测定结果
Figure 5. Disease symptoms and lengths of necrosis on excised shoots inoculated with strains of Botryosphaeria species
-
随着重庆市绿化工程的加快,越来越多的樟树被用作道旁树。本研究调查发现,新移植的樟树上溃疡病的发生较重,极大地影响了樟树成活率和樟树的长势。目前,关于樟树溃疡病病原菌的报道较少,且多根据生物学特征进行鉴定。1995年郭立中等[5]首次报道了湖南地区的樟树溃疡病,2005年邓先琼等[6]根据病原菌的形态特征和培养特性,将该地区的樟树溃疡病菌鉴定为B. dothidea,本研究与该报道的结果一致。之后,赵桂华[7]于2009年对江苏省句容市的樟树溃疡病菌进行了分离,所得菌株初期为白色,后期变为深灰褐色,分生孢子为无色的近梭形单细胞,本研究中的Ⅰ型菌株与该特征相近,但是赵桂华等[7]将其鉴定为耧斗大茎点霉菌。其原因可能是因为葡萄座腔菌属真菌的培养特性容易受到培养条件、人员操作等因素的影响,且不同种间形态特征差异不明显,仅根据形态学特征对病原菌进行鉴定存在一定的偏差[15-18]。ITS序列能够实质性地反映出属间、种间碱基对的差异,近年来被广泛应用于真菌不同种间或近似属间的系统发育研究中[9, 19-20]。因此,本研究通过观察菌株形态学特征,并结合真菌ITS、β-tubulin和EF1-α序列的进化关系,对来源于重庆地区的樟树溃疡病菌进行鉴定,研究发现,引起樟树溃疡病的优势菌为B. dothidea;此外,新鉴定的B. parva和B. rhodina 2个种为引起樟树溃疡病的首次报道。该结果可为系统研究樟树溃疡病的病原菌提供理论依据,同时对樟树溃疡病的防治也具有重要指导意义。
樟树溃疡病菌在1年生离体枝条上的致病力测定结果显示:来源于重庆不同地区的樟树溃疡病菌间致病力的差异与其地域来源无明显的相关性,但与菌株种类存在明显相关性,B. parva菌株的致病性最强,B. rhodina菌株的致病性次之,B. dothidea菌株的致病性最弱。尽管本研究分离得到的优势种为B. dothidea,但其它2个高致病力的种可对我国其它地区的樟树存在潜在威胁。此外,有关报道表明,葡萄座腔菌属的许多种是我国落叶果树和林木的重要病原菌[16, 18],其中,B. dothidea、B. parva和B. rhodina在我国普遍存在,且在梨、桃、蓝莓和葡萄上均有其危害的报道[9, 21-23]。虽然目前重庆地区仅在梨树上发现有B. dothidea造成的危害[9],但重庆是我国柑橘、梨和枇杷等果树的主产区之一,因此,本研究从重庆不同地区樟树上分离得到的B. dothidea、B. parva和B. rhodina致病菌是否也能够引起柑橘、梨、葡萄和枇杷等果树的病害,有待进一步深入研究。
-
本研究对采自重庆不同地区的樟树溃疡病样品进行了病原菌的分离与纯化,根据获得的病原菌的形态学特征、ITS序列、β-tubulin序列和EF1-α序列及致病性,将重庆樟树溃疡病菌鉴定为B. dothidea、B. parva和B. rhodina,且不同种间致病力存在差异,B. parva对樟树致病性最强,B. rhodina次之,而B. dothidea致病性最弱,其中,B. rhodina和B. parva为引起樟树溃疡病的首次报道。该研究结果可为深入研究樟树溃疡病的发生流行规律以及制定防治策略奠定基础。
重庆樟树溃疡病病原菌的鉴定及序列分析
Identification and Sequence Analysis of Canker Pathogen of Camphor Tree in Chongqing
-
摘要:
目的 为了明确重庆樟树溃疡病的病原菌种类。 方法 采集重庆市渝中区、北碚区和涪陵区的樟树溃疡病样品进行病原菌的分离与纯化,观察病原菌的形态学特征,对不同菌落形态代表菌株的rDNA-ITS、β-tubulin和EF1-α序列进行扩增和测序,构建系统进化树,并测定其致病性。 结果 从重庆不同地区采集的樟树溃疡病样品中共分离得到48份菌株,单菌丝纯化后得到68株纯化菌株,根据纯化菌株在PDA培养基上的培养特性及分生孢子特征,将其分为Ⅰ型、Ⅱ型和Ⅲ型3种类型。选取不同类型的代表菌株构建系统进化树,结果表明:这3类菌株分别为葡萄座腔菌属中的Botryosphaeria dothidea、B.parva和B.rhodina。致病性测定结果发现:所有菌株均能使樟树枝条产生黑色坏死病斑,且B.parva菌株的致病性最强,B.rhodina菌株的致病性次之,B.dothidea菌株的致病性最弱。 结论 重庆樟树溃疡病的病原菌为B.dothidea、B.parva和B.rhodina,其中B.rhodina和B.parva为引起樟树溃疡病的首次报道。 Abstract:Objective To identify the pathogen of camphor tree (Cinnamomum camphora) canker disease, the diseased branches of camphor tree were collected from the Fuling, Beibei and Yuzhong district of Chongqing. Method Pathogens were isolated from the diseased samples and purified by single-mycelium tips culture. The species of pathogens were primarily identified by the morphological characterizations of purified strains. To analyze the phylogenetic of the selected strains, the sequences of rDNA-ITS, β-tubulin and EF1-α genes were amplified and sequenced. The selected strains showing different morphological and molecular characteristics were tested for their pathogenicity on detached shoots of camphor. Result A total of 48 isolates were isolated from the diseased camphor tree canker samples in Chongqing, and 68 purified strains were obtained by single-mycelium tip technique. Based on the characterization of fungal colony and conidia, the 68 purified strains were grouped into three types. Fourteen strains from the three types were selected to construct phylogenetic tree. The phylogenetic analysis results showed that the three types belong to Botryosphaeria dothidea (type Ⅰ), B. parva (type Ⅱ) and B. rhodina (type Ⅲ). Pathogenicity tests on the camphor tree shoots showed that all the tested strains could induce canker lesions. Remarkably, the strains of B. parva exhibited the strongest virulence than that of the B. rhodina and B. dothidea strains. The B. dothidea strains showed the weakest virulence. Conclusion B. parva, and B. rhodina are isolated from camphor canker disease for the first time. It is suggested that B. dothidea, B. parva, and B. rhodina are the common causal agents for camphor tree canker diseases in Chongqing. -
Key words:
- camphor tree canker
- / Botryosphaeria
- / sequence analysis
- / pathogenicity
-
图 2 樟树溃疡病菌菌落特征及分生孢子形态
Figure 2. Colonial and conidial morphologies of camphor tree canker strains
图 3 基于不同葡萄座腔菌属真菌的ITS序列构建的系统进化树
Figure 3. Phylogenetic tree of Botryosphaeria species associated with camphor canker disease in Chongqing Municipality based on the Neighbour-Joining analysis of ITS nucleotide sequences
图 4 基于不同葡萄座腔菌属真菌的ITS、β-tubulin和EF1-α序列构建的系统进化树
Figure 4. Phylogenetic tree of Botryosphaeria species associated with camphor canker disease in Chongqing Municipality based on the Neighbour-Joining analysis of combined ITS、β-tubulin and EF1-α nucleotide sequences
图 5 不同菌株接种樟树离体枝条症状及致病性测定结果
Figure 5. Disease symptoms and lengths of necrosis on excised shoots inoculated with strains of Botryosphaeria species
表 1 樟树溃疡病菌不同类型菌株的分生孢子大小
Table 1. Conidial dimensions of camphor tree canker strains
菌株Strains 分生孢子大小Conidial dimensions 长×宽Length×width/μm (长×宽)平均值±标准误Mean of Length×width±SD/μm 长/宽L/W Ⅰ型 YZS1-2a (18.36~32.23)×(5.55~12.35) (25.64±2.44)×(8.53±1.51) 3.08±0.54 YZS11-1a (19.21~26.21)×(5.42~10.40) (22.36±2.14)×(9.16±0.97) 3.04±0.51 YZS5-1a (21.36~32.24)×(8.55~14.56) (28.16±2.44)×(11.22±1.44) 2.54±0.35 Ⅱ型 YZS9-1a (21.82~32.23)×(9.03~17.96) (26.33±2.35)×(11.79±1.62) 2.48±0.49 YZS9-1b (18.23~29.66)×(7.50~13.58) (24.01±2.17)×(10.40±1.31) 2.34±0.29 YZS1-1a - - - YZS3-2a - - - YZS9-1c - - - YZS9-1d - - - Ⅲ型 YZS1-6a (19.12~26.51)×(10.54~14.86) (22.43±1.45)×(12.62±1.13) 1.79±0.15 YZS3-3a (19.36~26.23)×(9.85~15.48) (22.96±1.53)×(13.27±1.13) 1.74±0.21 YZS8-1a (21.86~29.58)×(12.69~18.19) (26.50±1.66)×(15.32±1.19) 1.74±0.15 YZS8-1b (22.23~33.61)×(12.65~19.97) (28.45±2.29)×(16.41±1.54) 1.74±0.14 YZS7-6a - - - 注:每菌株分别统计50个分生孢子;“-”观察期内未产生分生孢子(30 d)。
Note: The length and width of 50 conidia were measured from each selected strains. “-” No conidia were observed during the observation period (30 days). 下载: 导出CSV
下载: 导出CSV
-
[1] 李振华, 温强, 戴小英, 等.樟树资源利用现状与展望[J].南方林业科学, 2007 (6) : 30-33. [2] 曾祥谓, 谢锦忠, 朱春玲, 等.上海佘山国家森林公园主要森林群落的结构特征和植物多样性[J].林业科学研究, 2010, 23(3) : 375-381. [3] 陈正春, 胡耀民, 程平.樟树常见的几种虫害及防治[J].江西林业科技, 2012, 40(6) : 22-23. doi: 10.3969/j.issn.1006-2505.2012.06.010 [4] 张岳德, 章其江, 董涛海.香樟树食叶类害虫的发生种类及防治技术[J].现代农业科技, 2012, 8 (8): 180-182. doi: 10.3969/j.issn.1007-5739.2012.08.115 [5] 郭立中, 邓先琼, 韦石泉.樟树的一种新病害——樟树溃疡病病原菌鉴定[J].植物病理学报, 1995, 25 (1) : 28. [6] 邓先琼, 郭立中.樟树溃疡病病原菌再鉴定[J].湖南林业科技, 2005, 32 (1) : 39-43. doi: 10.3969/j.issn.1003-5710.2005.01.011 [7] 赵桂华.耧斗大茎点霉菌引起的樟树溃疡病病原菌特性研究[J].西部林业科学, 2009, 38 (2) : 2-5. [8] 方中达.植病研究方法[M].第3版.北京:中国农业出版社, 1998 : 46-50. [9] Zhai L F, Zhang M X, Lv G, et al. Biological and molecular characterization of four Botryosphaeria species isolated from pear plants showing stem wart and stem canker in China[J]. Plant Disease, 2014, 98 (6) : 716-726. doi: 10.1094/PDIS-10-13-1060-RE [10] Sambrook J, and Russell D W. Molecular Cloning: A Laboratory Manual[M]. New York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 2001 : 44-46. [11] Glass N L, and Donldson G C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes[J]. Applied and Environmental Microbiology, 1995, 61 (4) : 1323-1330. [12] Ignazio C, and Linda M K. A method for designing primer sets for speciation studies in filamentous ascomycetes[J]. Mycologia, 1999, 91 (3) : 553-556. doi: 10.1080/00275514.1999.12061051 [13] White T J, Bruns T, Lee S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[M]. New York: Academic Press, 1990 : 315-322. [14] Marsberg A, Kemler M, Jami F, et al. Botryosphaeria dothidea: a latent pathogen of global importance to woody plant health[J]. Moleculal Plant Pathology, 2017, 18 (4) : 477-488. [15] Dissanayake A J, Phillips A J, Li X, et al. Botryosphaeriaceae: Current status of genera and species[J]. Mycosphere, 2016, 7 (7) : 1001-1073. doi: 10.5943/mycosphere [16] 陈海燕, 田呈明, 梁军, 等.引起树木溃疡病病原菌Botryosphaeria属及相关无性态分类研究[J].西北林学院学报, 2006, 21 (6) : 145-150. doi: 10.3969/j.issn.1001-7461.2006.06.037 [17] Slippers B, Wingfield M J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: diversity, ecology and impact[J]. Fungal Biology Reviews, 2007, 21 (2-3) : 90-106. doi: 10.1016/j.fbr.2007.06.002 [18] 李文英, 李夏, 解开治, 等.葡萄座腔菌科真菌的系统学和多样性探讨[J].生物多样性, 2017, 25 (8) : 874-885. [19] 杨莉, 杨双昱, 麻文建, 等.核桃褐斑病病原菌的分离鉴定和发病规律的调查[J].林业科学研究, 2017, 30 (6) : 1004-1008. [20] 耿显胜, 张威, 仲建平, 等.早竹丛枝病的调查及病原菌的分子鉴定[J].林业科学研究, 2017, 30(5) : 805-811. [21] Wang F, Zhao L N, Li G H, et al. Identification and characterization of Botryosphaeria spp. causing gummosis of peach trees in Hubei Province, central China[J]. Plant Disease, 2011, 95 (11) : 1378-1384. doi: 10.1094/PDIS-12-10-0893 [22] Xu C, Zhang H, Zhou Z, et al. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China[J]. European Journal of Plant Pathology, 2015, 143 (4) : 737-752. [23] Yan J Y, Xie Y, Zhang W, et al. Species of Botryosphaeriaceae involved in grapevine dieback in China[J]. Fungal Diversity, 2013, 61 (1) : 221-236. doi: 10.1007/s13225-013-0251-8 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 5534
- HTML全文浏览量: 2035
- PDF下载量: 48
- 被引次数: 0
