


-
自然生态系统中物种之间存在较为复杂的关系,尤其是处于同一群落中的物种为了争夺有限生存空间和资源而不可避免与其他物种产生关联[1-2]。生态位能体现物种利用环境资源的能力和物种间的相互作用[3-4],也是物种共存、群落演替变化的主要动力[5]。种间联结是指不同物种在空间分布上的相互关联性,其因群落生境条件和物种特性的差异而不同[6-7],如黄土高原桥山林区的木本植物之间联结松散,但其乔木层内油松(Pinus tabuliformis Carriere)-青麸杨(Rhus potaninii Maxim.)、蒙古栎(Quercus mongolica Fischer ex Ledebour)-漆(Toxicodendron vernicifluum (Stokes) F. A. Barkl.)为显著正联结[7];广西大石围天坑群天坑森林植物群落总体呈现显著正联结,乔木层为显著正联结,而灌木层为不显著正联结[8];湖北省喜旱莲子草(Alternanthera philoxeroides (Mart.) Griseb.)植物群落表现为不显著负联结[4]。研究群落物种的联结性有助于揭示群落结构、物种分布及种间关系[9],有利于指导森林保护和经营[10-11]。森林群落中优势种不仅影响其它物种生长和分布,也具有维持群落结构和发挥生态系统功能等重要作用[12-13]。对群落主要木本植物的生态位和种间关联进行耦合研究,能够揭示群落结构、演替趋势、种间关系和空间分布[13-14]。此外,生态位和种间关联也是生态学重点研究领域之一[15]。各国学者已积极开展了不同群落类型主要物种的生态位或种间联结研究,例如人工林[16]、天然次生林[17]、珍稀保护物种群落[15,18]、浮游植物[19]等,但是前期研究均基于小面积的样地来探讨生态位,缺乏较大尺度研究,忽略物种空间分布异质性,也没有准确反映种间相互关系。因此,基于6 hm2每木定位的样地数据进行研究显得尤为必要,其能够更加全面的展现局域群落中物种,特别是主要物种的空间格局。
七姊妹山国家级自然保护区位于湖北省鄂西南武陵山区,具有独特自然环境和完整森林生态系统,被誉为亚热带常绿落叶阔叶混交林保存最为完好的区域之一[20]。亚热带常绿落叶阔叶混交林是我国特有的天然林类型,对维持生态平衡和保护生物多样性具有重要意义[20]。对七姊妹山亚热带常绿落叶阔叶混交林的研究主要涉及物种组成与多样性[21-22]、群落结构与动态[23]和少量优势物种的空间格局[13],而该区域主要木本植物的生态位和种间联结还尚未见详细报道。虽然姚良锦等[13]应用空间点格局分析该区域2个优势种的种间关系,但是没有考虑更多物种的生态位和种间关联。鉴于此,本研究以湖北七姊妹山常绿落叶阔叶混交林6 hm2 标准大样地为基础,以重要值大于1的16种主要木本植物为研究对象,探讨天然林次生林中主要木本植物的生态位和种间联结,明确主要木本植物对环境适应状况和种间关系,预测群落演替动态,为该区域生物资源保护和森林经营提供参考。
-
研究区为七姊妹山国家级自然保护区(29°39′30″ N~30°05′15″ N,109°38′30″ E~109°47′00″ E;见图1),位于湖北省恩施土家族苗族自治州宣恩县东北部,总面积34 550 hm2。海拔为651.0~2 014.5 m,地势呈现西北高西南低。属中亚热带季风湿润性气候,雨热同期、温和湿润;因地形复杂,气候随海拔梯度呈现明显垂直差异。年均气温13.7 ℃,最热月(7月)和最冷月(1月)的年平均温度分别为19.3 ℃和−2.1 ℃,年平均相对湿度84%,年平均降水量1 876.6 mm,全年日照时数1 213 h,无霜期263 d[21]。岩层以石英砂页岩、页质层岩和砂质层岩为主,土壤主要为黄棕壤和棕壤。植被类型为典型亚热带常绿落叶阔叶混交林。
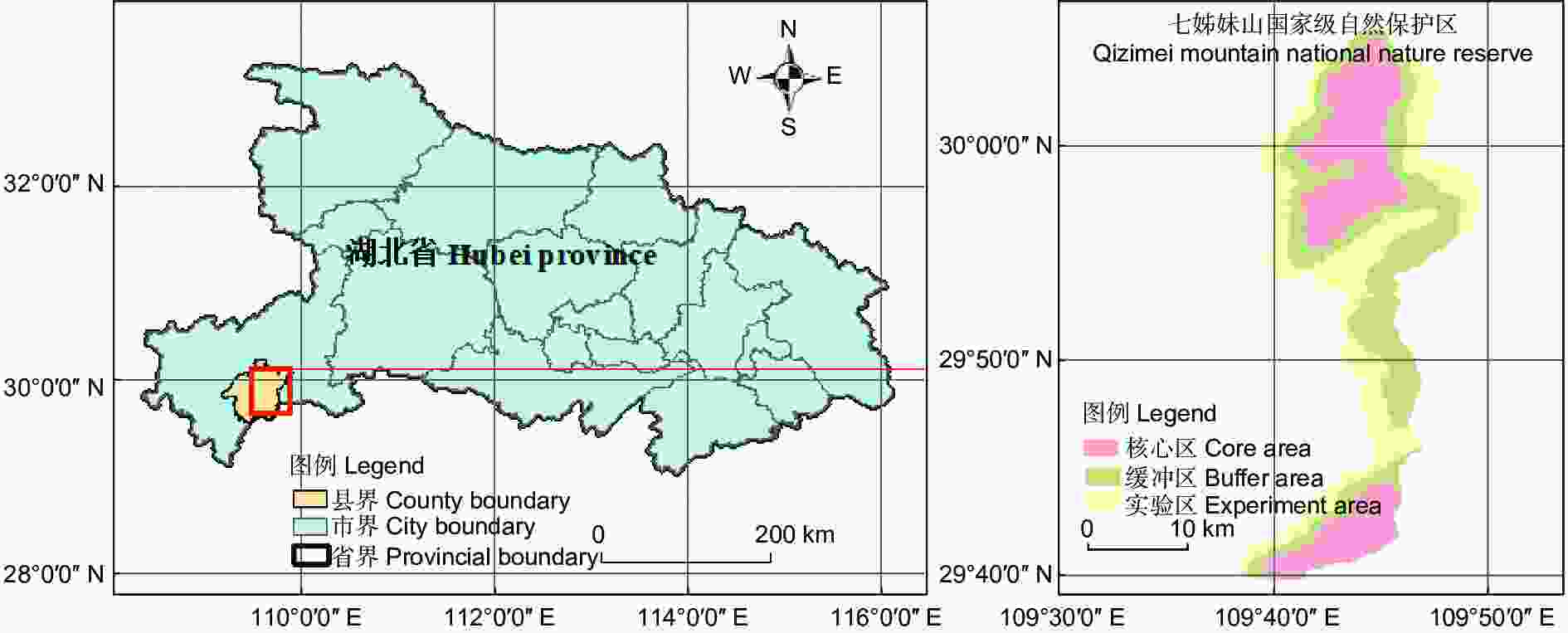
图 1 研究区域地理位置
Figure 1. Geographical location of the study area
-
2015年夏季,在湖北七姊妹山国家级自然保护区内建立6 hm2(南北长200 m、东西长300 m)固定监测标准大样地,大样地建设、调查方法和相关信息请详见文献[13,23]。大样地由150个20 m × 20 m的样地组成。2020年夏季,首轮复查该6 hm2固定监测标准大样地,根据2015年调查记录的树木个体坐标和牌号,重新测量并记录每株木本植物的胸径(DBH)、树高、冠幅及生长状况(枯立、断头、倒伏、死亡等);同时,对新增的DBH≥1 cm 的所有木本植物鉴别物种名并标记挂牌,检测并记录其胸径、树高、相对坐标和生长状况等。
-
依据物种重要值确定样地的主要物种,公式如下[17]:
$ {I_V} = \dfrac{{({R_a} + {R_f} + {R_d})}}{3} $
式中,IV为某个物种重要值,Ra为相对密度,Rf为相对频度,Rd为相对显著度。
-
(1)生态位宽度[24]:
$ {B_{{i}}} = \dfrac{1}{{r\sum\limits_{j = 1}^r {{{(\dfrac{{{n_{ij}}}}{{{N_i}}})}^2}} }} $
式中:Bi为物种i的生态位宽度,取值范围为[0, logr];r为资源等级数(用样方总数代替);nij为物种i在资源位j上的优势度(用物种重要值代替);Ni为物种i在所有资源位中重要值的总和。
(2)生态位相似性系数[25]:
$ {S_{ih}} = 1 - 0.5\sum\limits_{j = 1}^r {\left| {\dfrac{{{n_{ij}}}}{{{N_{{i}}}}}{\text{ - }}\dfrac{{{{{n}}_{{h}}}_{{j}}}}{{{{{N}}_{{h}}}}}} \right|} $
式中:Sih为物种i与物种h的生态位相似性系数;Ni和Nh分别表示物种i和物种h在所有资源位中重要值的总和;nij和nhj分别为物种i和物种h在资源位j中的重要值。该方程的值域为[0, 1],且Sih = Shi。
(3)生态位重叠指数[26]:
$ {O_{ih}} = \dfrac{{\sum\limits_{j = 1}^r {\left(\dfrac{{{n_{ij}}}}{{{N_{\text{i}}}}}\dfrac{{n_{hj}^{}}}{{{N_h}}}\right)} }}{{\sqrt {{{\left(\sum\limits_{j = 1}^r {\dfrac{{{n_{ij}}}}{{{N_{{i}}}}}} \right)}^2}{{\left(\sum\limits_{j = 1}^r {\dfrac{{{n_h}_j}}{{{N_{{h}}}}}} \right)}^2}} }} $
式中,Oih表示物种i和物种h之间的生态重叠指数,值阈为[0, 1],且Oih = Ohi;Ni、Nh、nij、nhj的含义同上述公式。
-
根据Schluter[27]提出的方差比率法(VR)计算物种总体联结性,再采用W统计量检测总体关联性是否显著,具体公式如下:
$ {V_R} = \dfrac{{\dfrac{1}{N}\sum\limits_{j = 1}^N {{{({T_j} - t)}^2}} }}{{\sum\limits_{i = 1}^S {\dfrac{{{n_i}}}{N}(1 - \dfrac{{{n_i}}}{N})} }} $
式中,VR为物种总体关联系数;N为总样地数;Tj为样地j中的物种数量;t为样地中物种平均数;S为总物种数;ni为物种i存在的样地数。在独立性零假设条件下,VR期望值为1时原假设成立,表示所有物种间无关联、相互独立;当VR >1,所有物种种间为正联结;当VR <1,所有物种种间表现为负联结[28]。
$ W = {V_R} \times N $
式中,W为W统计量,VR为物种总体关联系数,N为总样地数。若
$ {x}_{0.95(N)}^{2} < W < {x}_{0.05(N)}^{2} $ ,表示物种间总体关联不显著(P>0.05),反之为显著联结(P<0.05)[29]。 -
(1)χ2检验值:
基于重要值前16种主要木本植物在150个样方中是否出现,将原始数据转换成(0,1)二元数据矩阵(0为未出现,1为出现),构建种对间2 × 2联列表;求每一种对的a、b、c、d值,其中a为两物种同时出现的样地数,b和c分别是两个物种单独出现的样地数,d为两物种都未出现的样地数。考虑取样非连续性,因此χ2检验值需用Yates连续校正公式计算[29]。
$ {\chi ^2} = \dfrac{{N{{[\left| {ad - bc} \right| - 0.5N]}^2}}}{{(a + b)(b + d)(c + d)(a + c)}} $
式中,χ2为χ2检验值;N为总样地数。ad>bc,两物种间为正联结;ad<bc,两物种间呈现负联结;ad = bc,两物种相互独立。若χ2 > 6.635(P<0.01),表示两个物种呈现极显著联结;若3.841 ≤ χ2 ≤ 6.635(0.01≤ P ≤0.05),两物种间显著联结;若 χ2 < 3.841(P>0.05),表示种对间无联结性,相互独立。
(2)联结系数:联结系数可补充 χ2检验值,并反映两物种间的联结程度,公式如下[18]:
$ 若{ab}\ge {bc},则{A}_{C}=\dfrac{ad-bc}{(a + b)(b + d)} $
$ 若{bc}\ge {ad}且d\ge a,则{A}_{C}=\dfrac{ad-bc}{(a + b)(a + c)} $
$ 若{bc} > {ad}且d < a,则{A}_{C}=\dfrac{ad-bc}{(c + d)(b + d)} $
式中,AC为联结系数,取值范围为[−1, 1];AC越趋近1,表示两物种间正联结越强;AC越趋近-1,表示两物种间负相关越强;AC = 0,两物种之间无关联。
(3)Jaccard指数:Jaccard指数能较准确判断物种间正联结强度,避免d值和a值偏高或偏低造成的偏差[11],公式如下[30]:
$ {J_A} = \dfrac{a}{{(a + b + c)}} $
式中,JA表示为Jaccard指数,0≤ JA ≤1;JA越趋向于1,两物种间关联程度越大;JA为0时,两物种之间相互独立。
(4)Pearson相关系数与Spearman秩相关系数:基于定量数据,Pearson相关系数与Spearman秩相关系数能效弥 χ2检验值的结果,更灵敏地反映物种间关系[31],公式如下[29]:
$ {r_p}(i,k) = \dfrac{{\sum\limits_{j = 1}^N {({x_{ij}} - \overline {{x_i}} )({x_k}_j - \overline {{x_k}} )} }}{{\sqrt {\sum\limits_{j = 1}^N {{{({x_{ij}} - \overline {{x_i}} )}^2}\sum\limits_{j = 1}^N {{{({x_k}_j - \overline {{x_k}} )}^2}} } } }} $
$ {r_s}(i,{\text{k}}) = 1 - \dfrac{{6\sum\limits_{j = 1}^N {{{({x_{ij}} - {x_{kj}})}^2}} }}{{{N^3} - N}} $
式中,rp(i, k)和rs(i, k)分别表示为物种i和物种k在样地j中的Pearson相关系数和Spearman秩相关系数,取值范围为[-1, 1],正值表示正相关,负值为负相关;N为总样地数;xij与xkj分别为物种i和物种k在样地j中重要值;
$ \overline {{x_i}} $ 与$ \overline {{x_k}} $ 分别为物种i和物种k在所有样地中的重要值平均值。统计分析在R4.1.3中完成重要值计算(vegan包)、生态位特征(spaa包)、总体联结性检验(spaa包)、种间联结性分析(spaa包)和种间相关性测定(psych包)。
-
2020年调查发现七姊妹山国家级自然保护区亚热带常绿落叶阔叶混交林群落共有266种木本植物(40 871 株),隶属64科140属,以桦木科(Betulaceae)、壳斗科(Fagaceae)、金缕梅科(Hamamelidaceae)、山茱萸科(Cornaceae)等为主。该群落中共有16个物种的重要值高于1%(见表1),这16个物种视为常绿落叶阔叶混交林的主要物种,重要值之和达43.45%。其中,13个物种属于乔木生活型、2个物种为灌木生活型,仅有1个物种为木质藤本生活型。另外,这些主要木本植物的株数也排在前27位,仅川陕鹅耳枥(Carpinus fargesiana H. Winkl.)、多脉青冈(Cyclobalanopsis multinervis W.C.Cheng & T.Hong)和交让木(Daphniphyllum macropodum Miq.)3个物种的重要值高于5%。综上,选择重要值超过1%的16个物种进行后续生态位和联结性分析。
表 1 主要木本植物组成及重要值
Table 1. Composition and important values of main woody plants
种名
Species种代码
Species
code株数
Nubmer of
plants相对密度
Relative
density/%相对频度
Relative
frequency/%相对显著度
Relative
dominance/%重要值
Importance
value/%川陕鹅耳枥 (Carpinus fargesiana) Cf 3 128 7.65 0.01 14.76 7.48 多脉青冈 (Cyclobalanopsis multinervisg) Cm 3 707 9.07 0.06 8.27 5.80 交让木 (Daphniphyllum macropodum) Dm 3 422 8.37 0.02 7.36 5.25 缺萼枫香树 (Liquidambar acalycina) La 448 1.10 0.16 9.16 3.47 灯台树 (Bothrocaryum controversum) Bc 538 1.32 0.08 8.27 3.22 四照花 (Dendrobenthamia japonica var. chinensis) Dj 2 138 5.67 0.02 2.60 2.76 亮叶桦 (Betula luminifera) Bl 493 1.21 0.14 6.19 2.51 翅柃 (Eurya alata) Ea 2 019 4.94 0.12 1.18 2.08 杉木 (Cunninghamia lanceolata) Cl 1 125 2.75 0.23 2.87 1.95 尖叶四照花 (Dendrobenthamia angustata) Da 1 147 2.81 0.07 1.61 1.50 黄丹木姜子 (Litsea elongata) Le 1 413 3.46 0.03 0.98 1.49 君迁子 (Diospyros lotus) Dl 391 0.96 0.21 2.86 1.34 梾木 (Swida macrophylla) Sm 388 0.95 0.15 2.85 1.32 漆 (Toxicodendron vernicifluum) Tv 266 0.65 0.15 2.90 1.23 油茶 (Camellia oleifera) Co 886 2.17 0.34 0.61 1.04 中华猕猴桃 (Actinidia chinensis) Ac 958 2.34 0.08 0.58 1.00 -
由图2可知,川陕鹅耳枥的生态宽度最高(90.90),其次是四照花(Dendrobenthamia japonica (DC.) Fang var. chinensis (Osborn.) Fang)(81.42),共有8个物种的生态位宽度均高于60。油茶(Camellia oleifera Abel.)的生态位宽度最小(11.89)。生态位宽度与物种重要值的排序并不完全相同,多脉青冈、交让木和缺萼枫香树(Liquidambar acalycina Chang)的重要值排在前4名,但这3个物种的生态位宽度分别排名5、9、10;中华猕猴桃的生态位宽度为66.45,排名第7,但是其重要值排名最后(第16名)。
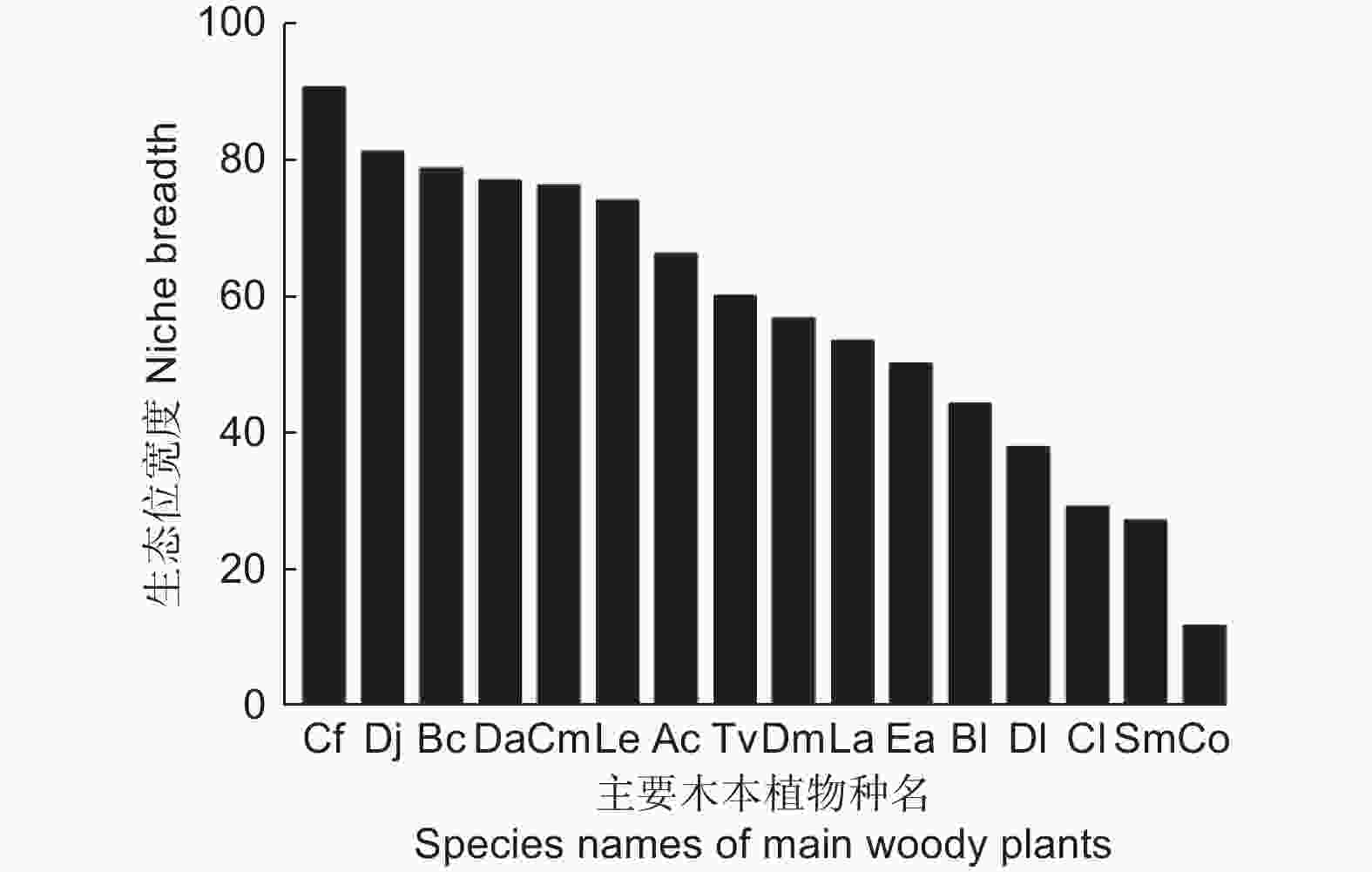
图 2 不同主要木本植物的生态位宽度
Figure 2. Niche width of different main woody plants
如表2所示,有21种对的生态位相似性系数高于0.5,占120组种对的17.50%。四照花-黄丹木姜子(Litsea elongata (Wall. ex Nees) Benth. et Hook. f.)的生态位相似性系数最高,为0.67。川陕鹅耳枥与其他15个物种之间的生态位相似性系数均较高,平均值为0.47。生态位相似性系数在0.2~0.5之间共有87对,占72.5%。12组种对的生态位相似性系数指数小于0.2,以油茶-君迁子(Diospyros lotus L.)最小(0.04)。生态位重叠指数的平均值为0.38,四照花-黄丹木姜子的生态位重叠指数最高(0.72),其次是川陕鹅耳枥-多脉青冈(0.70)、川陕鹅耳枥-四照花(0.70)。32组种对的生态位重叠指数大于0.5,其占总种对数的26.67%;19组种对的生态位重叠指数小于0.2,占15.83%。当0.2≤生态位重叠指数≤0.5时,共有69对,占57.50%,说明大部分物种对生境需求具有一定相似性、互补性。川陕鹅耳枥与其余物种的生态位重叠指数均较高,均值为0.49,油茶与其余物种的生态位重叠较少,油茶-君迁子最小(0.04)。
表 2 主要木本植物生态位相似性系数(对角线左)和生态位重叠指数(对角线右)
Table 2. Niche similarity coefficient (left diagonal) and niche overlap index (right diagonal) of main woody plants
Cf Cm Dm La Bc Dj Bl Ea Cl Da Le Dl Sm Tv Co Ac Cf 0.70 0.56 0.58 0.45 0.70 0.49 0.59 0.45 0.68 0.53 0.19 0.27 0.44 0.30 0.48 Cm 0.61 0.31 0.55 0.40 0.53 0.24 0.51 0.34 0.63 0.55 0.18 0.25 0.37 0.26 0.45 Dm 0.53 0.38 0.29 0.52 0.58 0.57 0.26 0.20 0.38 0.47 0.23 0.23 0.44 0.05 0.42 La 0.50 0.47 0.33 0.36 0.47 0.33 0.48 0.38 0.57 0.51 0.15 0.17 0.30 0.34 0.33 Bc 0.44 0.42 0.50 0.38 0.58 0.37 0.27 0.33 0.51 0.57 0.39 0.28 0.48 0.15 0.61 Dj 0.65 0.51 0.58 0.44 0.53 0.33 0.51 0.43 0.65 0.72 0.22 0.28 0.44 0.17 0.47 Bl 0.49 0.33 0.50 0.33 0.41 0.39 0.21 0.16 0.24 0.26 0.41 0.21 0.38 0.09 0.37 Ea 0.54 0.49 0.34 0.44 0.32 0.48 0.30 0.34 0.60 0.48 0.07 0.15 0.20 0.35 0.41 Cl 0.36 0.31 0.24 0.35 0.31 0.35 0.19 0.32 0.59 0.22 0.08 0.10 0.17 0.61 0.37 Da 0.61 0.59 0.45 0.51 0.48 0.59 0.32 0.53 0.46 0.57 0.13 0.22 0.37 0.46 0.55 Le 0.52 0.53 0.51 0.43 0.51 0.67 0.36 0.47 0.27 0.56 0.21 0.29 0.40 0.14 0.52 Dl 0.24 0.23 0.27 0.22 0.39 0.27 0.35 0.13 0.11 0.21 0.27 0.60 0.56 0.04 0.37 Sm 0.38 0.38 0.36 0.28 0.42 0.42 0.32 0.29 0.19 0.38 0.45 0.44 0.50 0.06 0.33 Tv 0.44 0.40 0.44 0.34 0.46 0.45 0.40 0.26 0.22 0.39 0.43 0.45 0.46 0.08 0.46 Co 0.21 0.20 0.09 0.24 0.11 0.16 0.09 0.29 0.39 0.30 0.15 0.04 0.11 0.08 0.38 Ac 0.49 0.43 0.46 0.32 0.55 0.49 0.41 0.37 0.33 0.52 0.52 0.35 0.45 0.44 0.20 -
根据16个主要木本植物的物种总体关联系数为1.67,其大于1,说明该群落整体呈现正联结。本研究中的W统计量为247.73,W统计量大于 χ20.05(150),说明物种间的总体联结性表现为显著正关联。
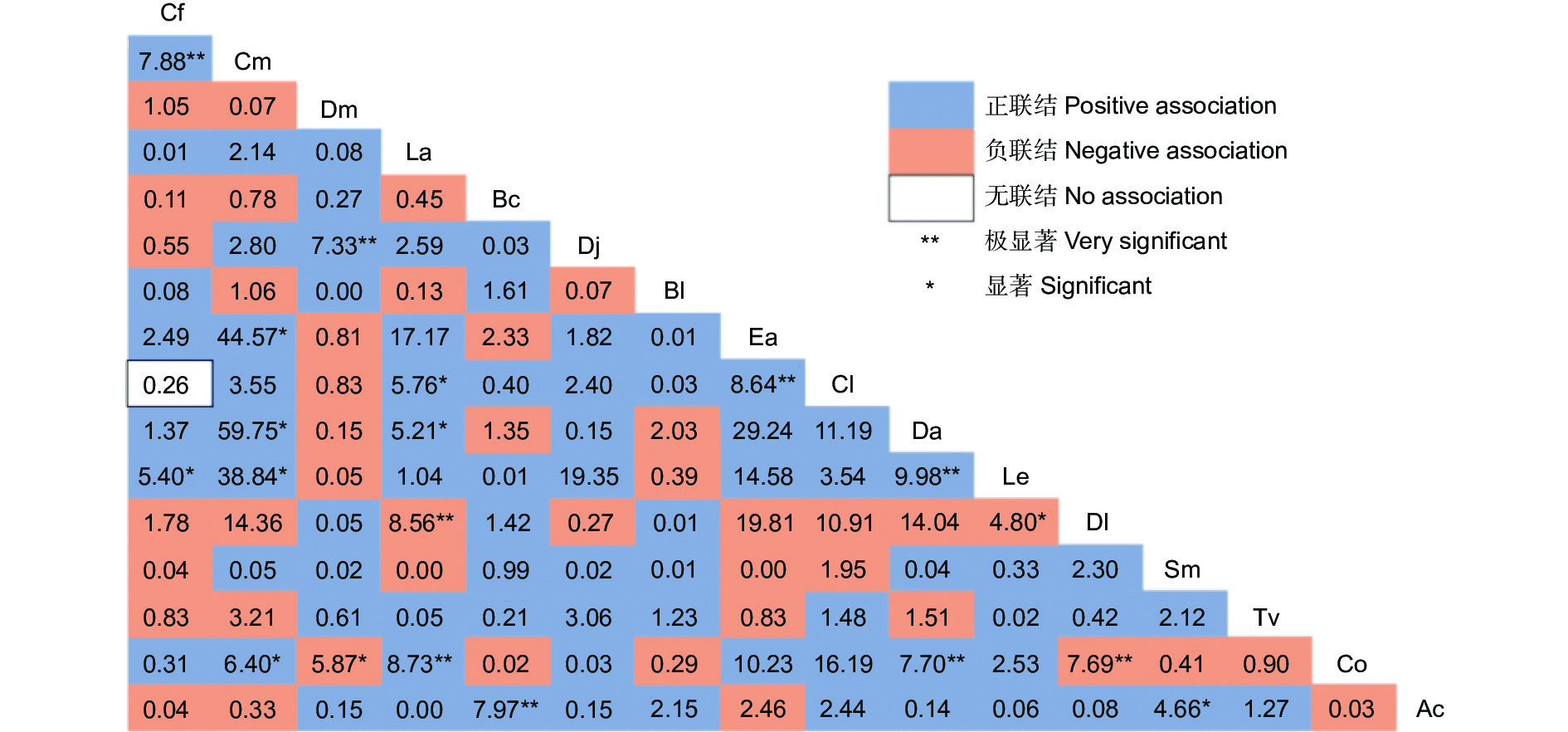
从 χ2检验结果可以看出(图3),正负联结比值为1.77,正联结有76对,占总种对数的63.33%。有7对属于极显著正联结,分别为川陕鹅耳枥-多脉青岗、四照花-交让木、缺萼枫香树-油茶、灯台树(Bothrocaryum controversum Hemsley)-中华猕猴桃(Actinidia chinensis Planch.)、杉木(Cunninghamia lanceolata (Lamb.) Hook.)-翅柃(Eurya alata Kobuski)、尖叶四照花(Dendrobenthamia angustata (Chun) Fang)-黄丹木姜子、尖叶四照花-油茶;显著正联结有8对,主要为多脉青冈与其他物种(翅柃、尖叶四照花、黄丹木姜子、油茶等)。所有种对间,只有杉木-川陕鹅耳枥是无联结。负联结数占总种对数的35.83%,君迁子-油茶、缺萼枫香树-君迁子呈现极显著负关联,黄丹木姜子-君迁子、交让木-油茶表现为显著负关联。未达到显著水平的种对数占总种对数的84.17%。中华猕猴桃-灯台树是极显著正联结,中华猕猴桃-梾木(Swida macrophylla (Wall.) Soják)呈现显著正联结。
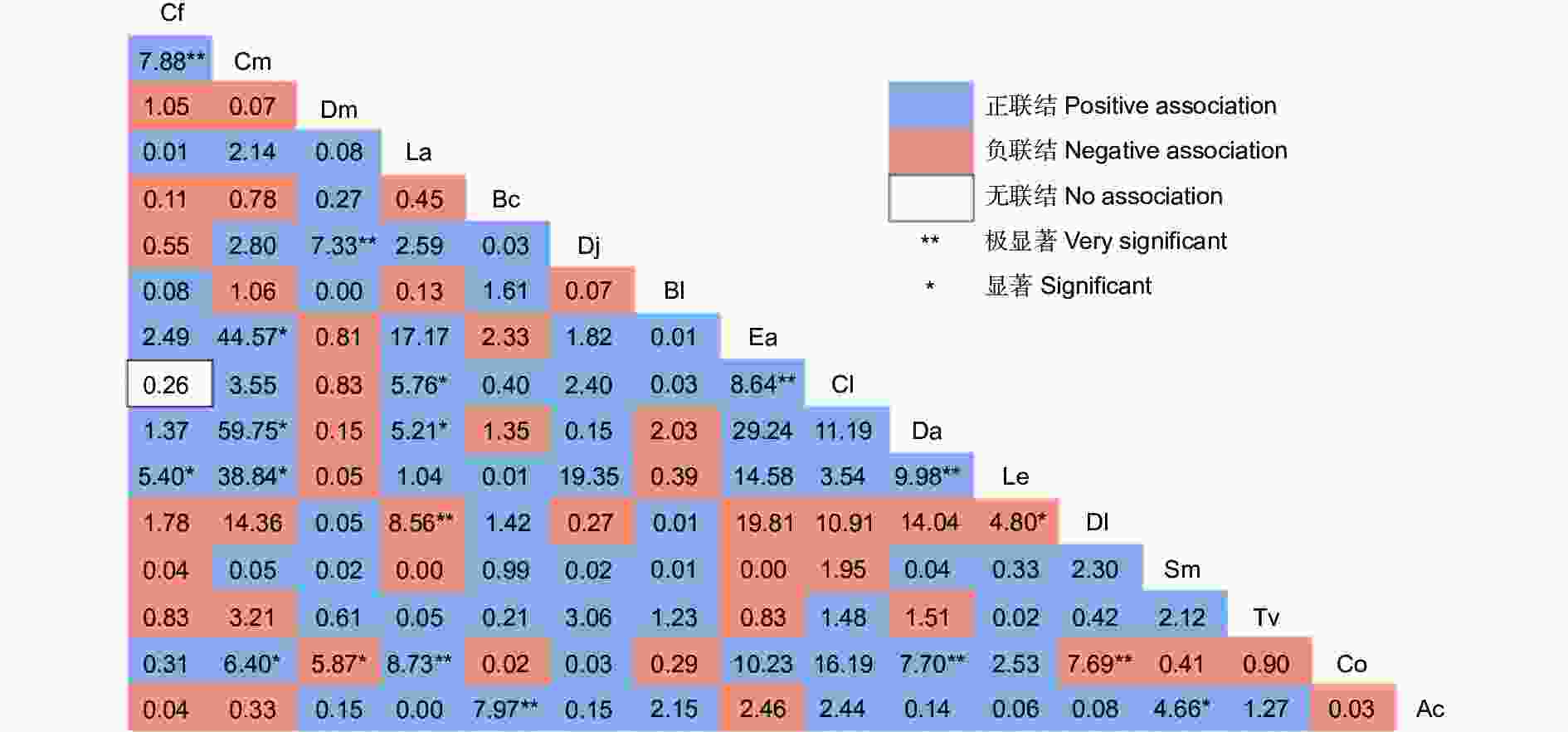
图 3 主要木本植物的 χ2检验半矩阵
Figure 3. Semi-matrix diagram of χ2 test of main woody plants
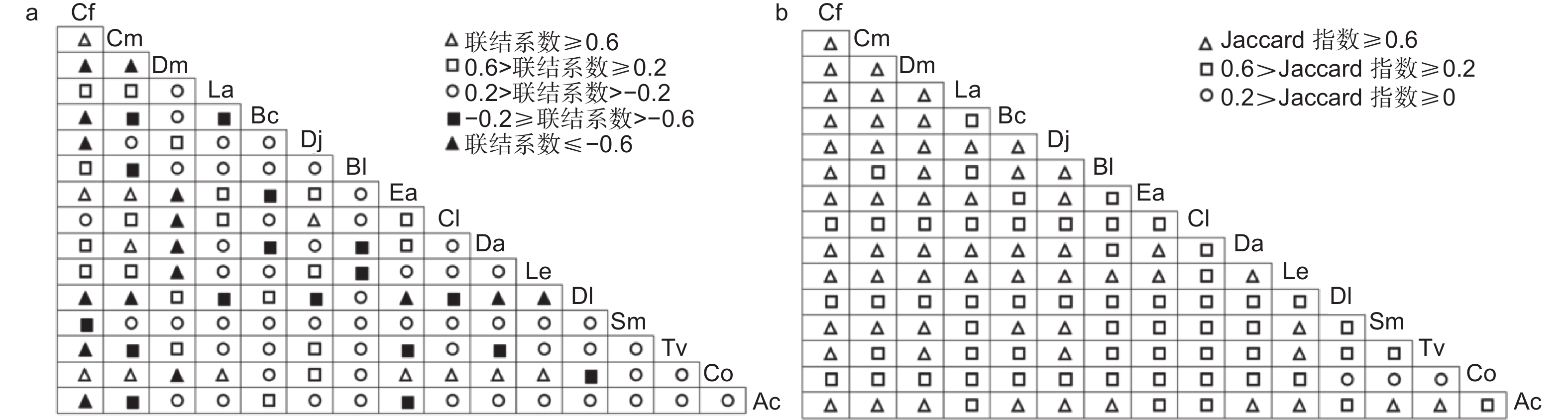
如图4a所示,当联结系数≥0.6时,有12对种间关系表现为很强正联结,川陕鹅耳枥-油茶、多脉青冈-油茶、尖叶四照花-油茶、黄丹木姜子-油茶的联结系数达到1。当0.2≤联结系数<0.6时,有20对呈现正联结、但联结较弱,占总种对数的16.67%。45.83%种对的联结系数在(-0.2, 0.2)之间,表现为联系极弱、相对独立,川陕鹅耳枥-杉木的联结系数为0。16对联结系数小于等于-0.6,其中10对的联结系数为-1,川陕鹅耳枥与其他物种(交让木、灯台树、四照花、君迁子、漆(Toxicodendron vernicifluum (Stokes) F. A. Barkl.)、中华猕猴桃)的竞争最激烈。联结系数在(-0.6, -0.2]之间有17对,呈现较弱负联结,占总种对数的14.17%。由图4b可知,Jaccard指数≥0.6时,有59对表现为较强的联结性,占总种对数的49.17%。中华猕猴桃与川陕鹅耳枥、多脉青冈、交让木、灯台树、四照花、尖叶四照花、黄丹木姜子、梾木、漆10个物种的Jaccard指数高于等于0.6。有58对的Jaccard指数在[0.2, 0.6)之间,呈现较弱的联结性。油茶-梾木、漆-油茶、油茶-君迁子这3种对间的Jaccard指数小于0.2。
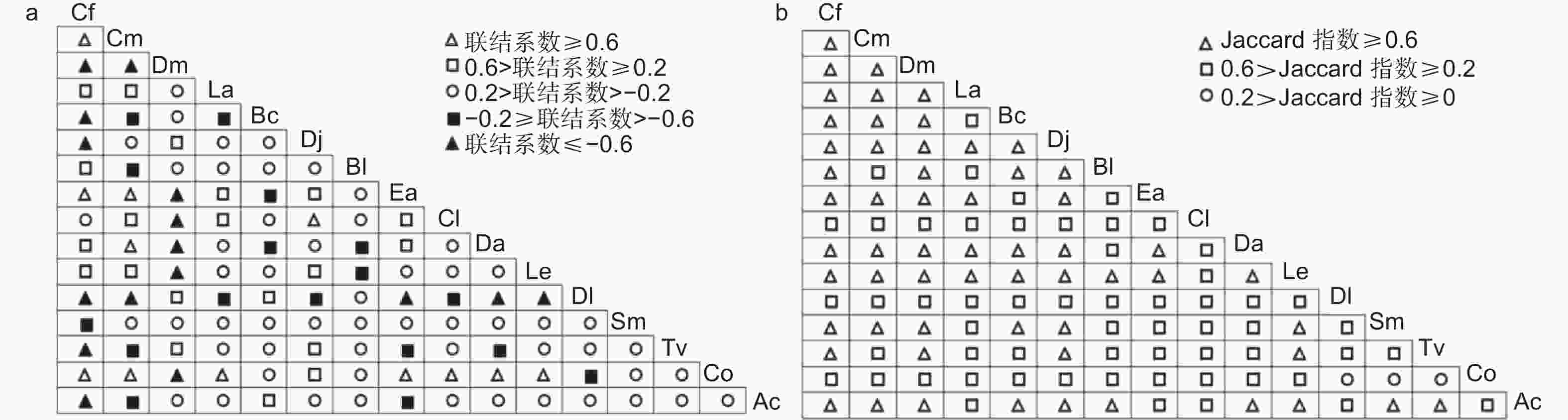
图 4 主要木本植物的联结系数和Jaccard指数半矩阵
Figure 4. Semi-matrices diagram about association coefficient and Jaccard index of main woody plants
-
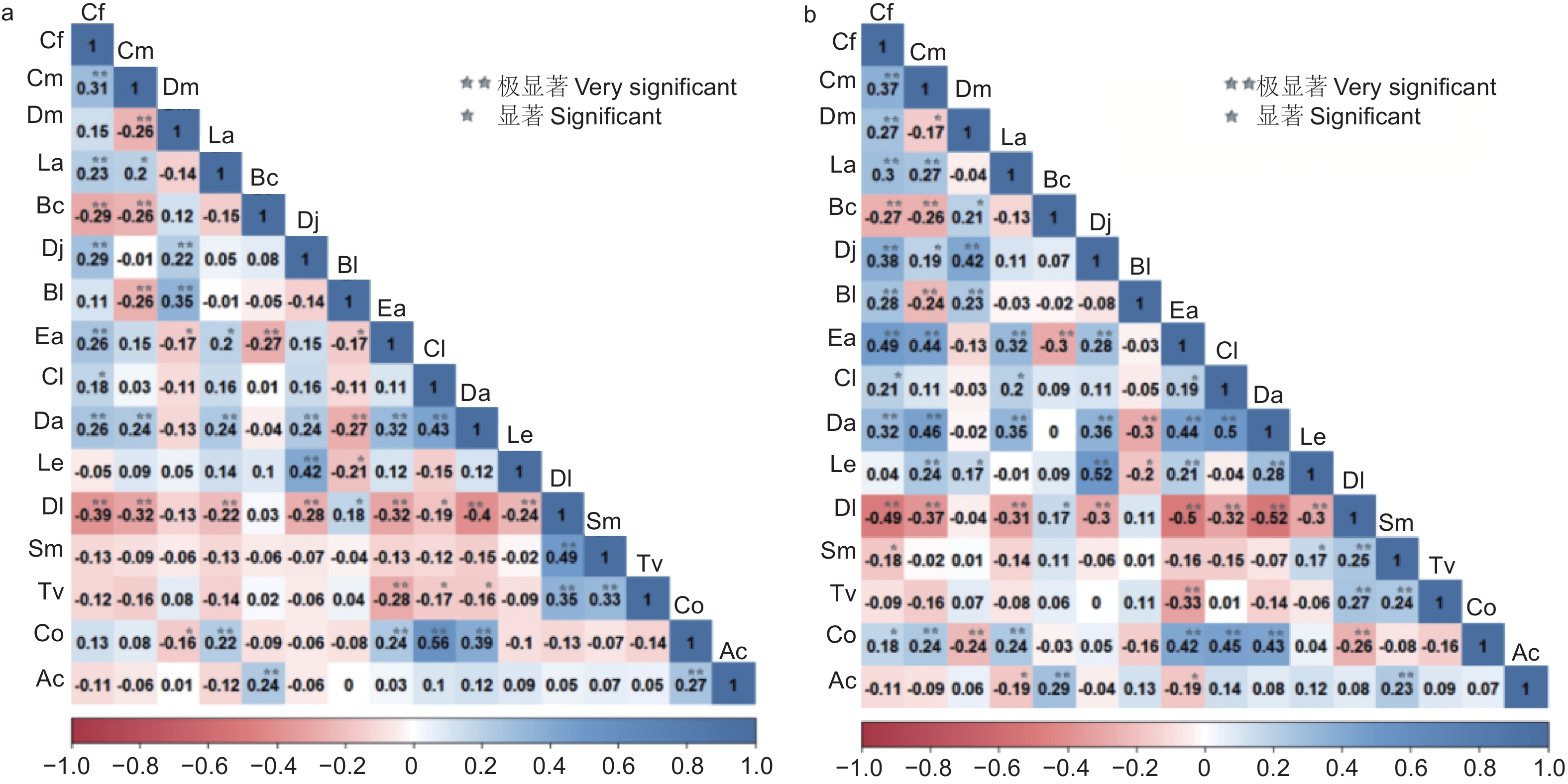
Pearson相关检验结果显示(见图5a),正负关联比为1:1.03,正相关的种对达59对,占总种对数的49.17%。22种对呈极显著正相关,4种对呈显著正相关,以杉木-油茶的正相关系数最高(0.56)、君迁子-梾木次之(0.49)。呈现负相关的种对占总种对数的50.83%,其中极显著负相关、显著负相关和不显著负相关的种对分别有14、7、40,分别占总种对数的11.67%、5.83%、33.33%。君迁子-尖叶四照花的负相关系数最高(−0.40),其次是川陕鹅耳枥-君迁子(−0.39)。未达显著相关的种对占总种对数60.83%,33对表现为不显著正相关。除中华猕猴桃-灯台树(0.24)、中华猕猴桃-油茶(0.27)呈现极显著正相关外,中华猕猴桃与其它物种的相关性均不显著。通过Spearman秩相关分析可以看出(图5b),正相关种对有68对,32对呈现极显著正相关,9对呈现显著正相关。四照花-黄丹木姜子的正相关系数最高(0.52),杉木-尖叶四照花(0.50)、翅柃-川陕鹅耳枥(0.49)次之。正负相关比为1.31。负相关的种对数比正相关的种对数减少23.53%,其中16对极显著负相关、5对显著负相关。负相关系数最高的是尖叶四照花-君迁子(−0.52),其次是翅柃-君迁子(−0.50)、君迁子-川陕鹅耳枥(−0.49)。未显著相关的种对数为58,其中不显著负相关比不显著正相关高14.81%。中华猕猴桃与灯台树、梾木的关系为极显著正相关,与缺萼枫香树、翅柃的种间关系表现为显著负相关。
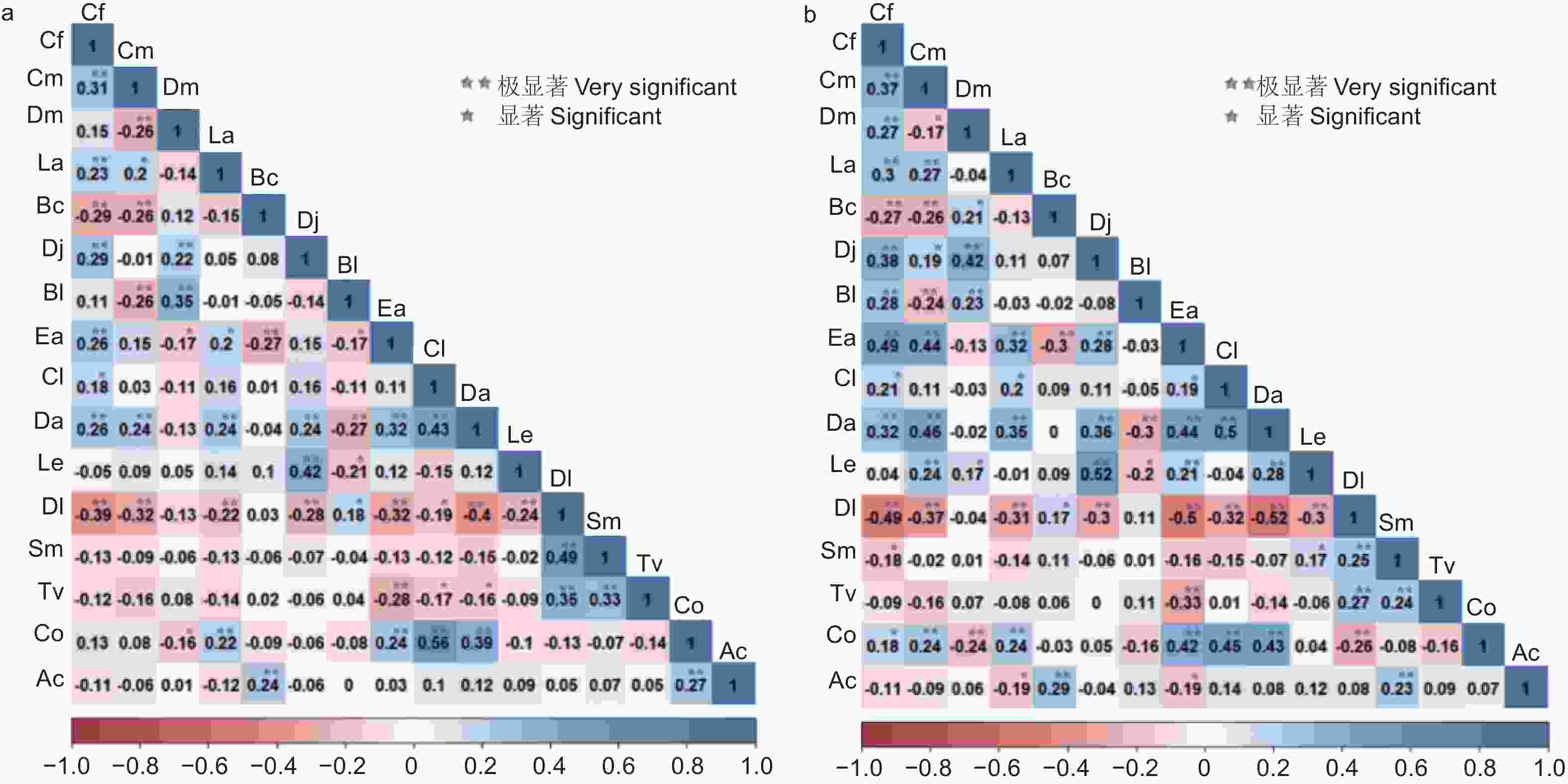
图 5 主要木本植物的Pearson和Spearman的相关系数及显著性
Figure 5. Correlation coefficients and significance of Pearson and Spearman for main woody plants
-
物种重要值和生态位宽度能体现物种在群落中的地位和作用[14],但两者侧重点不一样。在湖北七姊妹山亚热带常绿落叶阔叶混交林中,川陕鹅耳枥占据最高重要值,且生态位宽度最大,但其相对频度最小(排名16),说明川陕鹅耳枥为集群分布物种,且集群分布地还有其他优势物种存在。川陕鹅耳枥依旧是鄂西南亚热带常绿落叶阔叶混交林中典型优势树种和演替先锋种[13,21],占据绝对优势地位,具有很强的竞争力和资源利用能力,可以在适宜更新地块成片发展而形成集群分布格局的种群。四照花、灯台树和尖叶四照花的生态位宽度相对较高,可能因为这3种植物的适应性强、抵抗力强,且耐寒、耐旱、耐阴,可以充分利用环境资源。这与其他学者结果一致,生态位宽度越高,越有利于占据优势地位、充分利用资源[15,32]。油茶的相对频度排位第一,属于该群落中分布最均匀的物种,其相对显著度排名倒数第二,侧面反映了油茶个体较小;油茶的物种重要值和生态位宽度都较小,可能随着群落逐步发展完善、林内日益郁闭,导致喜光性小乔木油茶在林冠下的生长和更新减慢,竞争资源能力趋于衰弱。同时,杉木在该群落中也呈现均匀分布,其相对频度排名第二,但生态位宽度和物种重要值的排名较靠后,可能因为杉木喜光、不耐严寒及湿热,怕风,怕旱,在逐渐复杂化的群落结构中竞争力和生存力有所衰退。虽然中亚热带的长江流域曾广泛种植杉木和油茶,但是本研究中所有物种均来源于天然萌生,没有人为栽培种。加之该群落位于国家级自然保护区中,很少受外界干扰,致力于保护森林生态系统。今后构建亚热带常绿落叶混交林时优先选择川陕鹅耳枥、四照花、灯台树和尖叶四照花等适应性强的乡土树种。此外,本研究中主要木本植物的重要值排序与生态位宽度排序并不完全相同,这与闫东锋等[17]的研究结果一致。多脉青冈、交让木、缺萼枫香树的物种重要值分别排名第2、第3、第4,但其生态位宽度却排名靠后。因为物种生物学特性不同,对资源利用能力不一致,应对环境变化的方式也存在差异[33]。这也在一定程度上表明湖北七姊妹山亚热带常绿落叶阔叶混交林的资源还未被完全充分利用。
生态位重叠指数和生态位相似性系数作为量化生态位的重要指标,反映种间资源利用程度与性质[34-35]。生态位相似性系数和生态位重叠指数以0.20~0.50之间为主,说明该群落大部分物种对生境需求具有一定相似性。生态位宽度最大的川陕鹅耳枥与其他物种之间的生态位重叠和相似程度较高;生态位宽度最小的油茶与其余物种的生态重叠度和相似性都很低。这进一步证实了生态位宽度最大的物种与其他物种的生态位重叠和相似程度越高[33,36],川陕鹅耳枥、多脉青冈、四照花和尖叶四照花这些适应力强的物种具有相似的资源需求、生活习性,可能共享资源或呈现竞争关系[37]。四照花-黄丹木姜子的生态位重叠指数和生态位相似性系数均最高,这说明在有限资源条件下,四照花与黄丹木姜子之间的竞争最为激烈,可能因为四照花属于落叶树种,黄丹木姜子属于常绿树种,两者均为中小型乔木,且喜欢较阴湿环境。中华猕猴桃为本研究主要木本植物中唯一的木质藤本植物,它能攀爬到不同高度的树木、与其他15个主要木本植物的生态位重叠指数与生态位相似性指数均较高,说明藤本植物对攀援对象要求不严格,若不影响被攀爬物种的生存,则是一种互利共生关系,但若影响被攀爬物种的生存则可能是竞争或互斥关系。
-
种群间的总体关联性是反映群落所处演替阶段及群落稳定性的重要标准[8,14],湖北七姊妹山亚热带常绿落叶阔叶混交林总体呈现显著正关联,这与江常春等[38]、黄林娟等[8]和王乃江等[39]的研究结果一致。可能因为研究区处于国家级自然保护区内,很少受到人为活动的破坏与干扰,物种对资源利用相似性较强[40]。
综合应用 χ2检验、联结系数和Jaccard指数可以准确判定群落种对的联结性[39]。亚热带常绿落叶落叶混交林的16个主要木本植物的 χ2检验、联结系数和Jaccard指数基本一致呈现为:正关联种对数大于负关联种对数,极显著或显著正关联种对数比极显著或显著负关联多,不显著关联种对数也占有重要比例。这表明亚热带常绿落叶阔叶混交林处于自然演替的早期至中期阶段,种间联结不紧密[38]。四照花-交让木、多脉青冈-黄丹木姜子、油茶-多脉青冈均表现为很强正联结,说明两物种间具有相似的生态适应性,互补性强[7];油茶-君迁子则呈现很强负联结,因为两者均为喜阳、寿命较长的速生树种,生境竞争激烈。杉木-川陕鹅耳枥的 χ2检验和联结系数均为无联结,说明它们对环境资源的需求不一致,各自占据一定的生态位。此外,川陕鹅耳枥与交让木、灯台树、四照花、君迁子、漆、中华猕猴桃之间都表现为很强负联结,与多脉青岗、交让木、四照花、黄丹木姜子、尖叶四照花则为很强正联结,进一步说明了植物种间关联性与自身生物学特性密切相关,也与生存环境的地形、光照、水分、温度等紧密相关[11]。结合5个种间联结性分析指标,研究发现中华猕猴桃可能与灯台树、梾木等是互利共生关系,与其他不显著正联结的物种为无利无害关系,与负联结物种(川陕鹅耳枥、缺萼枫香树、翅柃)也许是竞争或互斥关系。
χ2检验、联结系数、Pearson相关系数和Spearman秩相关系数一致显示:君迁子-尖叶四照花、川陕鹅耳枥-君迁子呈现很强负相关。一方面,可能因为君迁子属于阳性树种,具有耐阴、耐寒、耐湿、耐干旱瘠薄和抗二氧化硫污染等特点,而尖叶四照花也是喜光树种,对温度、土壤和水分等环境的适应范围广。因此,君迁子和尖叶四照花对生境资源需求相同,利用资源方式相似,两者竞争激烈。另一方面,君迁子和川陕鹅耳枥均为喜光速生落叶乔木,都具有耐寒、耐干旱瘠薄等生物学特征,且川陕鹅耳枥作为典型优势种,竞争力强,所以君迁子和川陕鹅耳枥之间表现为很强竞争关系。这体现了物种之间竞争有限资源而相互排斥,也反映了物种间的生态位分离[37]。另外,Spearman结果的正关联对数、极显著或显著对数及正负相关比均高于Pearson结果,这与Gu等[41]江常春等[38]的研究结果一致。
研究尺度对种间联结性的精度起着决定性作用[42]。一般而言,环境异质性在大尺度研究中起到主要作用,正联结现象的几率增加;种间竞争在小尺度内中占主导作用,负联结概率增加[43]。大部分已有研究对象的尺度较小且非完整天然林群落,比如中亚热带常绿阔叶林的研究多集中于100 m2左右[14,43],因此本研究基于完整的6 hm2天然林次生林数据开展亚热带常绿落叶阔叶混交林的生态位与种间联结研究,以补充完善现有研究。
-
湖北七姊妹山国家级自然保护区亚热带常绿落叶阔叶混交林物种丰富,大部分物种对生境需求具有一定相似性;该群落物种总体关联性呈现显著正关联,但种间联系不紧密,群落处于演替早期至中期阶段。
该群落中的树种及树种搭配可以为亚热带地区人工营造混交林提供一定参考依据。此外,本研究仅客观描述现有物种生态位和种间关系,并不能揭示其背后的生态学过程与形成机制,需要进一步综合应用植物生理生化、植物功能性状、遗传基因等多方面深入揭示种间关联形成机制,今后也需要结合长期固定动态研究跟踪群落演替过程中的变化规律。另外,各物种的空间分布格局(集群分布、随机分布、均匀分布)与植物群落的生态位和种间联结性密切相关,应该进一步研究不同群落演替阶段下,各个主要物种的空间分布格局对植物群落生态位与种间联结的影响。
湖北七姊妹山亚热带常绿落叶阔叶混交林主要木本植物的生态位与种间联结
Niche and Interspecific Association of Main Woody Plants in the Subtropical Evergreen-Deciduous Broadleaved Mixed Forest in Qizimei Mountain, Hubei Province
-
摘要:
目的 探究亚热带常绿落叶阔叶混交林的生态位特征和种间联结,阐明主要木本植物种间关系及对环境适应状况,为亚热带地区生物资源保护和森林管理提供理论依据。 方法 本研究基于湖北七姊妹山国家级自然保护区6 hm2(300 m × 200 m)标准大样地数据,从物种组成、生态位宽度、生态位相似性系数、生态位重叠指数、方差比率法、χ2检验、联结系数、Jaccard指数等方面研究了16种主要木本植物(重要值大于1%)的生态位和种间联结。 结果 (1)2020年湖北七姊妹山亚热带常绿落叶阔叶混交林共有64科140属266种木本植物,主要有桦木科、壳斗科和金缕梅科等,物种组成逐渐复杂。(2)川陕鹅耳枥的物种重要值和生态位宽度最高,占据绝对优势地位,与其他主要木本植物之间具有较高的生态位重叠和生态位相似性。四照花、灯台树和尖叶四照花的优势地位次之。生态位重叠指数和生态位相似系数均以0.2~0.5之间为主,大部分物种对生境需求具有一定相似性、互补性。物种重要值排序与生态位宽度排序并不完全相同。(3)湖北七姊妹山亚热带常绿落叶阔叶混交林内物种间总体关联性呈现显著正联结,且正负联结比值大于1。16种主要木本植物的χ2检验、联结系数、Jaccard指数、Pearson相关系数和Spearman秩相关系数的结果基本一致:正关联种对数大于负关联种对数,极显著或显著正关联种对数比极显著或显著负关联种对数多,不显著关联种对数也占有重要比例。 结论 亚热带常绿落叶阔叶混交林物种丰富,群落处于演替早期至中期阶段,种间关系松散、不紧密。 -
关键词:
- 亚热带常绿落叶阔叶混交林
- / 物种重要值
- / 生态位
- / 种间联结
- / 七姊妹山国家级自然保护区
Abstract:Objective The species niches characteristics and interspecific associations of the subtropical evergreen-deciduous broadleaved mixed forest were explored to clarify the interspecific relationship and environmental adaptation of main woody plants, and ultimately provide a theoretical basis for biological resource protection and forest management in subtropical regions. Method Based on 6 hm2 (300 m × 200 m) standard large sample plot data of Qizimei Mountain National Nature Reserve in Hubei Province, the niches and interspecific associations of 16 main woody plants (important value greater than 1%) were studied from species composition, niche breadth, niche similarity coefficient, niche overlap index, variance ratio method, χ2 test, association coefficient, and Jaccard index. Result (1) In 2020, there were 266 species of woody plants belonging to 140 genera and 64 families in the subtropical evergreen-deciduous broadleaved mixed forest in Qizimei Mountain, Hubei Province, mainly including Betulaceae, Fagaceae and Hamamelidaceae. The species composition tended to be more complex. (2) The species importance value and niche breadth of Carpinus fargesiana were the highest, occupying an absolute dominant position. There was a high niche overlap and niche similarity between C. fargesiana and other main woody plants. The dominant position of Carpinus fargesiana, Dendrobenthamia japonica var. chinensis and Dendrobenthamia angustata was second. Both the niche overlap index and the niche similarity coefficient were mainly between 0.2 and 0.5, and most species had certain similarity and complementarity to habitat requirements. In addition, species importance ranking was not exactly the same as niche breadth ranking. (3) The overall correlation between species in the subtropical evergreen-deciduous broadleaved mixed forest of the Qizimei Mountain, Hubei Province showed a significantly positive association, and the ratio of positive to negative association was greater than one. The results of χ2 test, association coefficient, Jaccard index, Pearson and Spearman correlation coefficient of 16 main woody plants were generally consistent. The number of positive associated species pairs was greater than that of negative associated species pairs, and the number of extremely significant or significant positive associated species pairs was more than that of extremely significant or significant negative associated species pairs. The number of non-significantly associated species pairs also accounted for an important proportion. Conclusion The subtropical evergreen-deciduous broadleaved mixed forest is rich in species. The community is in the early to middle stages of succession, and the interspecific association is loose and not close. -
图 4 主要木本植物的联结系数和Jaccard指数半矩阵
Figure 4. Semi-matrices diagram about association coefficient and Jaccard index of main woody plants
图 5 主要木本植物的Pearson和Spearman的相关系数及显著性
Figure 5. Correlation coefficients and significance of Pearson and Spearman for main woody plants
表 1 主要木本植物组成及重要值
Table 1. Composition and important values of main woody plants
种名
Species种代码
Species
code株数
Nubmer of
plants相对密度
Relative
density/%相对频度
Relative
frequency/%相对显著度
Relative
dominance/%重要值
Importance
value/%川陕鹅耳枥 (Carpinus fargesiana) Cf 3 128 7.65 0.01 14.76 7.48 多脉青冈 (Cyclobalanopsis multinervisg) Cm 3 707 9.07 0.06 8.27 5.80 交让木 (Daphniphyllum macropodum) Dm 3 422 8.37 0.02 7.36 5.25 缺萼枫香树 (Liquidambar acalycina) La 448 1.10 0.16 9.16 3.47 灯台树 (Bothrocaryum controversum) Bc 538 1.32 0.08 8.27 3.22 四照花 (Dendrobenthamia japonica var. chinensis) Dj 2 138 5.67 0.02 2.60 2.76 亮叶桦 (Betula luminifera) Bl 493 1.21 0.14 6.19 2.51 翅柃 (Eurya alata) Ea 2 019 4.94 0.12 1.18 2.08 杉木 (Cunninghamia lanceolata) Cl 1 125 2.75 0.23 2.87 1.95 尖叶四照花 (Dendrobenthamia angustata) Da 1 147 2.81 0.07 1.61 1.50 黄丹木姜子 (Litsea elongata) Le 1 413 3.46 0.03 0.98 1.49 君迁子 (Diospyros lotus) Dl 391 0.96 0.21 2.86 1.34 梾木 (Swida macrophylla) Sm 388 0.95 0.15 2.85 1.32 漆 (Toxicodendron vernicifluum) Tv 266 0.65 0.15 2.90 1.23 油茶 (Camellia oleifera) Co 886 2.17 0.34 0.61 1.04 中华猕猴桃 (Actinidia chinensis) Ac 958 2.34 0.08 0.58 1.00  下载: 导出CSV
下载: 导出CSV
表 2 主要木本植物生态位相似性系数(对角线左)和生态位重叠指数(对角线右)
Table 2. Niche similarity coefficient (left diagonal) and niche overlap index (right diagonal) of main woody plants
Cf Cm Dm La Bc Dj Bl Ea Cl Da Le Dl Sm Tv Co Ac Cf 0.70 0.56 0.58 0.45 0.70 0.49 0.59 0.45 0.68 0.53 0.19 0.27 0.44 0.30 0.48 Cm 0.61 0.31 0.55 0.40 0.53 0.24 0.51 0.34 0.63 0.55 0.18 0.25 0.37 0.26 0.45 Dm 0.53 0.38 0.29 0.52 0.58 0.57 0.26 0.20 0.38 0.47 0.23 0.23 0.44 0.05 0.42 La 0.50 0.47 0.33 0.36 0.47 0.33 0.48 0.38 0.57 0.51 0.15 0.17 0.30 0.34 0.33 Bc 0.44 0.42 0.50 0.38 0.58 0.37 0.27 0.33 0.51 0.57 0.39 0.28 0.48 0.15 0.61 Dj 0.65 0.51 0.58 0.44 0.53 0.33 0.51 0.43 0.65 0.72 0.22 0.28 0.44 0.17 0.47 Bl 0.49 0.33 0.50 0.33 0.41 0.39 0.21 0.16 0.24 0.26 0.41 0.21 0.38 0.09 0.37 Ea 0.54 0.49 0.34 0.44 0.32 0.48 0.30 0.34 0.60 0.48 0.07 0.15 0.20 0.35 0.41 Cl 0.36 0.31 0.24 0.35 0.31 0.35 0.19 0.32 0.59 0.22 0.08 0.10 0.17 0.61 0.37 Da 0.61 0.59 0.45 0.51 0.48 0.59 0.32 0.53 0.46 0.57 0.13 0.22 0.37 0.46 0.55 Le 0.52 0.53 0.51 0.43 0.51 0.67 0.36 0.47 0.27 0.56 0.21 0.29 0.40 0.14 0.52 Dl 0.24 0.23 0.27 0.22 0.39 0.27 0.35 0.13 0.11 0.21 0.27 0.60 0.56 0.04 0.37 Sm 0.38 0.38 0.36 0.28 0.42 0.42 0.32 0.29 0.19 0.38 0.45 0.44 0.50 0.06 0.33 Tv 0.44 0.40 0.44 0.34 0.46 0.45 0.40 0.26 0.22 0.39 0.43 0.45 0.46 0.08 0.46 Co 0.21 0.20 0.09 0.24 0.11 0.16 0.09 0.29 0.39 0.30 0.15 0.04 0.11 0.08 0.38 Ac 0.49 0.43 0.46 0.32 0.55 0.49 0.41 0.37 0.33 0.52 0.52 0.35 0.45 0.44 0.20
下载: 导出CSV
-
[1] BROOKER R W, MAESTRE F T, CALLAWAY R M, et al. Facilitation in plant communities: the past, the present, and the future[J]. Journal of Ecology, 2008, 96(1): 18-34. [2] 张东梅, 赵文智, 罗维成. 荒漠草原带盐碱地优势植物生态位与种间联结[J]. 生态学杂志, 2018, 37(5):1307-1315. doi: 10.13292/j.1000-4890.201805.027 [3] PERALTA G, PERRY G L W, VÁZQUEZ D P, et al. Strength of niche processes for species interactions is lower for generalists and exotic species[J]. Journal of Animal Ecology, 2020, 89(9): 1-28. [4] 郭佳琦, 陈俊辰, 黄 旬, 等. 喜旱莲子草入侵群落主要物种生态位和种间联结研究[J]. 生态环境学报, 2021, 30(8):1607-1616. doi: 10.16258/j.cnki.1674-5906.2021.08.007 [5] MA Y X, WANG Y L, LI G T, et al. HunShanDaKe sand east edge of vegetation ecological niche and interspecific association studies[J]. IOP Conference Series: Earth and Environmental Science, 2019, 351(1): 012045. [6] GREIG-SMITH P. Quantitative plant ecology[M]. Oxford: Blackwell Scientific Publication, 1983. [7] 许金石, 陈 煜, 王国勋, 等. 陕北桥山林区主要木本植物群落种间联结性[J]. 西北植物学报, 2014, 34(7):1467-1475. doi: 10.7606/j.issn.1000-4025.2014.07.1467 [8] 黄林娟, 于燕妹, 安小菲, 等. 广西大石围天坑群天坑森林主要木本植物种间关联性研究[J]. 广西植物, 2021, 41(5):695-706. doi: 10.11931/guihaia.gxzw202008028 [9] 舒 琪, 徐瑞晶, 胡 璇, 等. 海南岛甘什岭青梅群落主要乔木树种生态位与种间联结[J]. 生态学杂志, 2021, 40(9):2689-2697. doi: 10.13292/j.1000-4890.202109.040 [10] SETA T, DEMISSEW S, WOLDU Z. Floristic diversity and composition of the Biteyu forest in the Gurage mountain chain (Ethiopia): implications for forest conservation[J]. Journal of Forestry Research, 2019, 30(1): 319-335. doi: 10.1007/s11676-018-0623-8 [11] 赵 安, 田 青, 周晓雷, 等. 迭山北坡云冷杉林火烧迹地灌木树种种间关联性[J]. 林业科学研究, 2022, 35(2):163-170. [12] LOHBECK M, POORTER L, MARTÍNEZ-RAMOS M, et al. Changing drivers of species dominance during tropical forest succession[J]. Functional Ecology, 2014, 28(4): 1052-1058. doi: 10.1111/1365-2435.12240 [13] 姚良锦, 姚 兰, 易咏梅, 等. 亚热带常绿落叶阔叶混交林优势种川陕鹅耳枥和多脉青冈的空间格局[J]. 林业科学, 2018, 54(12):1-11. doi: 10.11707/j.1001-7488.20181201 [14] 张 盟, 郑俊鸣, 万佳艺, 等. 福建省东门屿优势木本植物的生态位与种间联结[J]. 森林与环境学报, 2022, 42(1):11-19. [15] ZHANG L, SU Z Y, CHEN B G. Interspecific relationships in the forest community dominated by Pinus kwangtungensis, an endangered species endemic to China[J]. Frontiers of Forestry in China, 2007, 2(2): 128-135. doi: 10.1007/s11461-007-0021-4 [16] 薛 鸥, 魏天兴. 鹫峰国家森林公园低效人工林林下植物种间关联[J]. 植物研究, 2016, 36(1):34-42. doi: 10.7525/j.issn.1673-5102.2016.01.005 [17] 闫东锋, 张妍妍, 吕康婷, 等. 太行山南麓不同海拔梯度天然林优势树种生态位特征[J]. 生态环境学报, 2021, 30(8):1571-1580. doi: 10.16258/j.cnki.1674-5906.2021.08.003 [18] 温鑫鸿, 王其炳, 潘 辉, 等. 天宝岩柳杉群落主要乔木种群的种间联结性[J]. 森林与环境学报, 2022, 42(1):1-10. [19] MA Y M, LI Q H, PAN S P, et al. Niche and interspecific associations of Pseudoanabaena limnetica-Exploring the influencing factors of its succession stage[J]. Ecological Indicators, 2022, 138: 108806. [20] 黄永涛, 姚 兰, 艾训儒, 等. 鄂西南两个自然保护区亚热带常绿落叶阔叶混交林类型及其常绿和落叶物种组成结构分析[J]. 植物生态学报, 2015, 39(10):990-1002. doi: 10.17521/cjpe.2015.0096 [21] 朱强. 鄂西南七姊妹山常绿落叶阔叶混交林物种多样性格局及维持机制[D]. 恩施: 湖北民族大学, 2020. [22] 郭秋菊, 陈礼波, 艾训儒, 等. 七姊妹山常绿落叶阔叶混交林中典型群落的结构与多样性分析[J]. 林业资源管理, 2017(1):91-97. doi: 10.13466/j.cnki.lyzygl.2017.01.016 [23] 姚良锦, 姚 兰, 易咏梅, 等. 湖北七姊妹山亚热带常绿落叶阔叶混交林的物种组成和群落结构[J]. 生物多样性, 2017, 25(3):275-284. doi: 10.17520/biods.2016180 [24] LEVINS R. Evolution in changing environments: some theoretical explorations[M]. Princeton: Princeton University Press, 1968, 12-25. [25] SCHOENER T W. Resource partitioning in ecological communities: research on how similar species divide resources helps reveal the natural regulation of species diversity[J]. Science, 1974, 185(4145): 27-39. doi: 10.1126/science.185.4145.27 [26] PIANKA E R. The structure of lizard communities[J]. Annual Review of Ecology and Systematics, 1973, 4(1): 53-74. doi: 10.1146/annurev.es.04.110173.000413 [27] SCHLUTER D. A variance test for detecting species associations with some example applications[J]. Ecology, 1984, 65(3): 998-1005. doi: 10.2307/1938071 [28] ROUSSET O, LEPART J. Positive and negative interactions at different life stages of a colonizing species (Quercus humilis)[J]. Journal of Ecology, 2000, 88(3): 401-412. doi: 10.1046/j.1365-2745.2000.00457.x [29] 张金屯. 数量生态学 (第三版)[M]. 北京: 科学出版社, 2018. [30] KADMON R, PULLIAM H R. Island biogeography: effect of geographical isolation on species composition[J]. Ecology, 1993, 74(4): 977-981. doi: 10.2307/1940467 [31] BISHARA A J, HITTNER J B. Testing the significance of a correlation with nonnormal data: comparison of Pearson, Spearman, transformation, and resampling approaches[J]. Psychological methods, 2012, 17(3): 399-417. doi: 10.1037/a0028087 [32] 孙杰杰, 江 波, 吴初平, 等. 浙江省檫木林生境与生态位研究[J]. 生态学报, 2019, 39(3):884-894. [33] 李一凡, 刘梦芸, 甘先华, 等. 深圳市坝光湿地园银叶树群落优势种生态位特征[J]. 生态环境学报, 2020, 29(11):2171-2178. doi: 10.16258/j.cnki.1674-5906.2020.11.004 [34] MILNE G R, MASON C H. An ecological niche theory approach to the measurement of brand competition[J]. Marketing Letters, 1990, 1(3): 267-281. doi: 10.1007/BF00640803 [35] 杨新东, 陈流保, 曾献兴, 等. 广东翁源青云山自然保护区森林群落优势树种的生态位特征[J]. 热带亚热带植物学报, 2020, 28(6):565-573. doi: 10.11926/jtsb.4214 [36] WALKE B. Conserving biological diversity through ecosystem resilience[J]. Conservation Biology, 1995, 9(4): 747-752. doi: 10.1046/j.1523-1739.1995.09040747.x [37] 何友均, 崔国发, 邹大林, 等. 三江源自然保护区玛珂河林区寒温性针叶林优势灌木种间联结研究[J]. 林业科学, 2006, 42(12):126-129. doi: 10.3321/j.issn:1001-7488.2006.12.022 [38] 江常春, 梁希妮, 田 磊, 等. 浙江省常绿阔叶林优势树种种间联结分析[J]. 浙江师范大学学报(自然科学版), 2022, 45(4):437-445. [39] 王乃江, 张文辉, 陆元昌, 等. 陕西子午岭森林植物群落种间联结性[J]. 生态学报, 2010, 30(1):67-78. [40] JOHNSON D J, BEAULIEU W T, BEVER J D, et al. Conspecific negative density dependence and forest diversity[J]. Science, 2012, 336(6083): 904-907. doi: 10.1126/science.1220269 [41] GU L, O'HARA K L, LI W, et al. Spatial patterns and interspecific associations among trees at different stand development stages in the natural secondary forests on the Loess Plateau, China[J]. Ecology and Evolution, 2019, 9(11): 6410-6421. doi: 10.1002/ece3.5216 [42] JONSSON, B G, MOEN J. Patterns in species associations in plant communities: the importance of scale[J]. Journal of Vegetation Science, 1998, 9(3): 327-332. doi: 10.2307/3237097 [43] 宋育红, 邢建宏, 邓贤兰. 格氏栲自然保护区常绿阔叶林群落优势树种种间联结性分析[J]. 井冈山大学学报(自然科学版), 2021, 42(4):64-70. -

 点击查看大图
点击查看大图

图(5) / 表(2)
计量
- 文章访问数: 2556
- HTML全文浏览量: 1470
- PDF下载量: 66
- 被引次数: 0
















