


-
植物物候是植物为了适应环境变化而形成的以年为周期的生长节律,如展叶和开花等,在全球生态系统中扮演着重要角色[1]。温度是影响植物物候的决定性因素[2-4]。近年来,气候急剧变暖使植物物候发生了显著的变化[5];而物候的改变会进一步影响植物的个体适应性、群落结构、生态系统功能、碳氮循环乃至全球气候[1,6-8]。因此, 准确理解植物物候对气候变化的响应具有十分重要的意义。
目前,大量研究表明,气候变暖使植物春季物候(萌芽和展叶等)显著提前[5,8]。然而,植物物候对不同季节气温的响应并不相同[9],对此研究人员关注得还较少。植物在春季物候发生之前一般需要经过生理和生态休眠2个阶段。在生理休眠阶段, 植物需要积累一定的低温才能解除休眠[10]。这意味着冬季气温上升可能会导致低温累积不足, 从而使春季物候延迟[11]。在生态休眠阶段, 植物则需要满足一定的热量累积来促进其分生组织的发育[10],这个阶段气温上升会使春季物候提前[12]。对于萌芽期来说,其影响因子跟其他春季物候又不相同[13]。夏秋季节,植物生长停止后,需要经过芽形成、休眠建立、休眠解除和积温促进几个阶段后才开始萌芽,而在每个时段芽的发育对气象因子的响应都不同[14]。
相较于春季物候,秋季物候(叶变色和落叶等)对气候变暖的响应更加多样化[15-16]。随着气温上升,秋季物候有推迟趋势[3,17],也有提前趋势[18],有的则没有变化[12]。较多观点认为,秋季气温和光周期是影响秋季物候的主要因子,并认为秋季气温上升使秋季物候推迟[8,12]。然而,秋季物候的变化可能跟夏末[17]、初秋[18]和春末[19]等季节的气温都有关系。尤其是近年来有研究发现,相对于秋季气温的推迟作用,春夏季气温升高会加速叶片衰老从而使秋季物候提前[15,20],且秋季气温的推迟作用可能会被春夏季气温的提前作用所抵消[21]。可见,不同季节气温的上升对秋季物候也具有不同的作用[18]。因此,衡量植物物候对各季节气温的响应方式及差异对于深入理解物候与气温的关系十分重要。
此外,过去的研究多集中在1个或2个物候期[8,22],而不同物候期对同一阶段气温的响应也有差异[8]。如研究发现,展叶期和始花期对气候变暖的响应机制有很大不同[9,12],二者对温度的敏感度表现出了显著差异[22-23];还有研究显示,亚高山多年生草本植物始花期对气候变暖的响应比末花期更强烈[24];而木本植物芽的形成和叶片衰老对夏秋季气温上升的响应程度也不相同[16]。不同植物物候期对气候变化响应的差异会造成生物间的物候期不匹配[8],改变资源配置和种间竞争[1],从而影响植物与传粉者、寄主植物与病原物等之间的相互作用[25-26],但目前这方面的研究还没有得到广泛的关注。
过去几十年里,北半球,尤其是30°~90° N地区,冬春季气温的上升速率显著高于其他季节 [27],在一些地区其差异甚至达2倍以上[16]。这使物候对气候变化的响应及其未来的变化趋势更加复杂,也使得相关研究更加迫切;但目前,物候对非对称性季节气温变化响应的研究还较少。因此,本研究将利用2003—2019年北京市东灵山暖温带森林24种木本植物的萌芽期、展叶始期、始花期和盛花期4种春季物候期及叶变色期和落叶期2种秋季物候期数据,采用偏最小二乘回归和相关系数法分析各物候期对气温响应的最优相关阶段,对比物候期对各相关时段的气温响应敏感度的差异,以期揭示北京市东灵山森林木本植物物候对气温响应的季节差异性,为应对气候变化对生态系统带来的影响提供理论基础。
-
研究点位于中国科学院北京森林生态系统定位研究站(简称北京站),地处北京市门头沟区东灵山(115˚26΄E 和39˚58΄N) , 海拔1 260 m。该地区为暖温带半湿润大陆性季风气候, 年平均、最低和最高气温分别为5.4、0.6和12.1 ℃,年降水量490 mm。植被类型为暖温带落叶阔叶林(优势种为辽东栋(Quercus mongolica Mary))和人工常绿针叶林(优势种为油松(Pinus tabuliformis Carr.))。
-
物候资料为2003—2019年的萌芽期、展叶始期、始花期、盛花期、叶变色期和落叶期数据, 来源于国家生态科学数据中心(
http://www.cnern.org.cn )。为保证足够的样本量, 筛选具有10 a以上记录的植物作为研究对象, 最终选取24种木本植物(乔木和灌木各12种),观测年数为11~16 a (表1),平均萌芽期和展叶始期分别为4月14日和27日,平均始花期和盛花期分别为5月7日和13日,平均叶变色期和落叶期分别为9月12日和23日。气象数据采用2002—2019年北京站和1970—2020年河北省怀来县气象站的日平均气温数据,前者来自国家生态科学数据中心,后者来自国家气象科学数据中心。怀来县气象站位于研究点以北55 km处,因北京站的气温数据有一定缺失, 采用怀来县气象站的气温数据通过线性回归方法对缺失数据进行插补。表 1 研究所选物种及相关信息
Table 1. Summary of the species used in the study and related information
乔木物种
Tree species观测年数
Number of
observed
years/a灌木物种
Shrub species观测年数
Number of
observed
years/a白桦 Betula platyphylla Sukaczev 14~15 大花溲疏 Deutzia grandiflora Bunge 15 黑桦 Betula dahurica Pall. 11 蒿柳 Salix schwerinii E. L. Wolf 15 胡桃楸 Juglans mandshurica Maxim. 11 金花忍冬 Lonicera chrysantha Turcz. 13~14 花曲柳 Fraxinus chinensis subsp.
Rhynchophylla (Hance) E. Murray11 六道木 Zabelia biflora (Turcz.) Makino 15~16 华北落叶松 Larix gmelinii var. principis
-rupprechtii (Mayr) Pilg.11 毛榛 Corylus mandshurica Maxim. & Rupr. 13~14 辽东栎 Quercus mongolica Mary 15~16 巧玲花 Syringa pubescens Turcz. 11 裂叶榆 Ulmus laciniate (Trautv.) Mayr 15~16 土庄绣线菊 Spiraea pubescens Turcz. 11 青杨 Populus cathayana Rehder 15 卫矛 Euonymus alatus (Thunb.) Siebold 13~14 山桃 Prunus davidiana (Carrière) Franch. 15~16 小花溲疏 Deutzia parviflora Bunge 15~16 山杏 Prunus sibirica L. 15~16 小叶鼠李 Rhamnus parvifolia Bunge 15 五角枫 Acer pictum Thunb. 13~14 迎红杜鹃 Rhododendron mucronulatum Turcz. 15 油松 Pinus tabuliformis Carrière 15 照山白 Rhododendron micranthum Turcz. 15 -
由于各物候期对气温的响应存在季节性差异,而如何识别各响应阶段起止时间,目前还没有非常准确的办法。文中采用偏最小二乘(PLS)回归来初步判断气温对每个物候期影响显著的时段。PLS回归可通过建立潜在因子来解决自变量高度相关和模型过度拟合问题,在自变量超出样本量和相关度较高的情况下具有较强优势[28]。为了使识别效果更显著, 采用日平均气温的11 d(该日及其前后各5 d)滑动平均值来代替日值[29]。将24种植物的各物候期分别与物候期前一年日平均气温的滑动平均值建立PLS回归,结果输出标准化模型系数和变量投影重要性(VIP)2个重要参数。标准化模型系数表示气温对物候影响的方向及强度,系数为负表明气温升高使物候期提前,系数为正则表明气温升高使物候期推迟。VIP 值反映了日平均气温对物候期影响的显著性,当VIP≥0.8时,判断模型系数显著[30]。因此, 把PLS回归结果中模型系数显著且连续为正或为负的时段看作气温对物候期影响的相关时段。
由于PLS初步判断的相关时段可能不是最优的,有时会出现模型系数和显著性短暂不连续的现象,因此,文中再通过相关系数法计算温度对物候期影响最大的相关时段。其计算方法为:以相关时段的结束日期为结束日,以1 d为步长,分别计算其前1、2、3……n d的平均气温与物候期的Pearson相关系数,并选择相关系数绝对值最大的时段作为最优相关时段[31],n一般设定在通过PLS判断的相关时段之内。
将每种植物的6个物候期分别作为因变量,将最优相关时段内平均气温的滑动平均值分别作为自变量,进行线性回归,将所得的回归系数作为衡量物候期对各气温要素响应敏感度的指标。此外,应用线性回归分析北京站物候期及平均气温在2003—2019年的变化趋势。最后,利用非参数检验中的Wilcoxon 符号秩检验分别检测各物候期变化趋势及各物候期对最优相关时段内气温响应敏感度的差异。
-
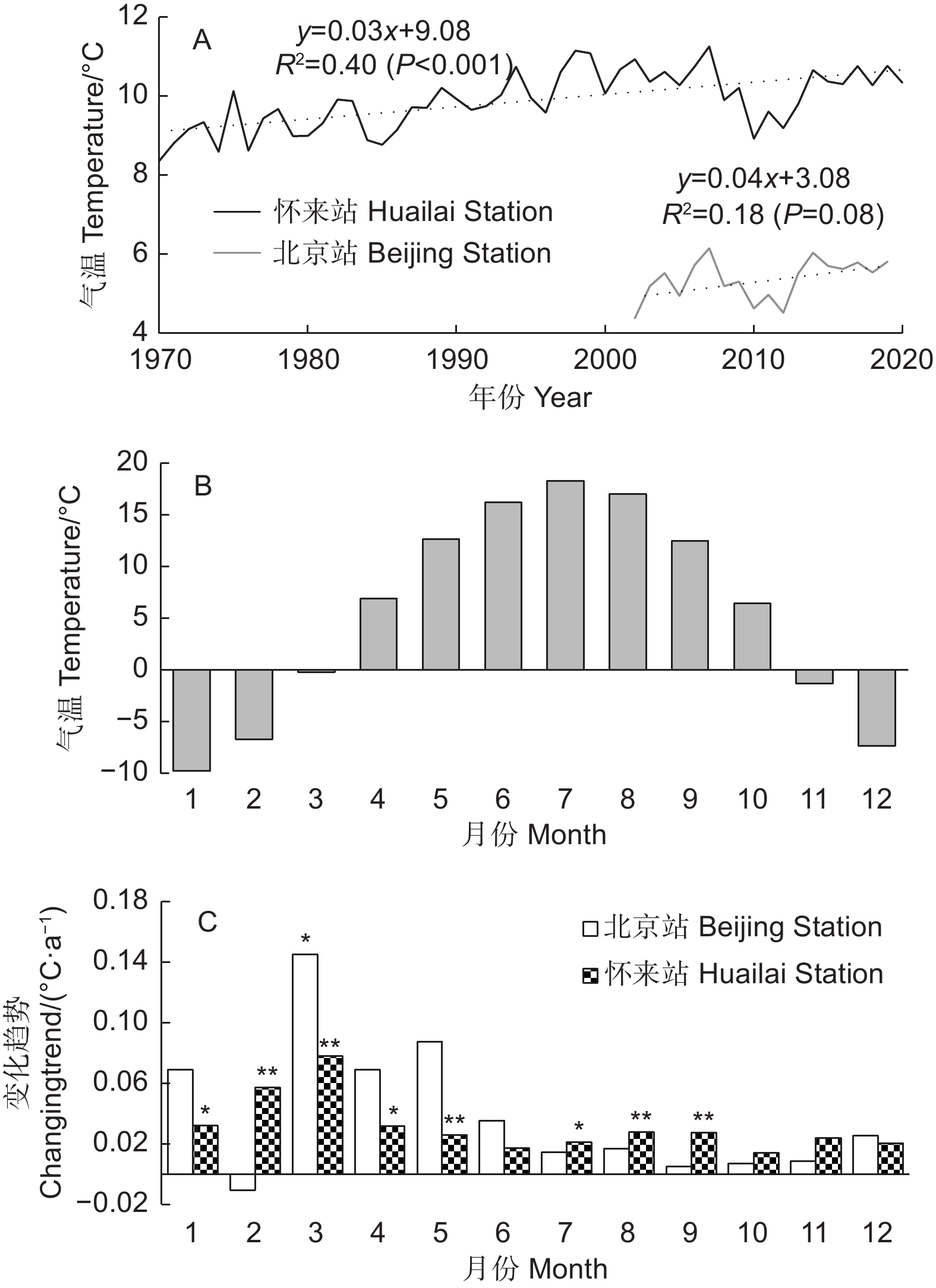
2002—2019年,北京站年平均气温未表现出显著的上升趋势(P=0.08),可能因为2008—2013年气温出现了明显的下降,不过长期看,1970—2020年,怀来气象站年平均气温以0.03 ℃·a−1的速度显著上升,说明研究区气候总体呈变暖趋势(图1A)。从月平均气温的变化趋势看,在研究时段内,北京站3月气温显著升高(图1C);而怀来县在过去50 a里冬末春初(2月和3月)气温上升最快(速率分别为0.06、0.08 ℃·a−1,P<0.01),是其他月份的2倍以上(图1C),其次是1月和4月及5、7—9月的气温变化速率。
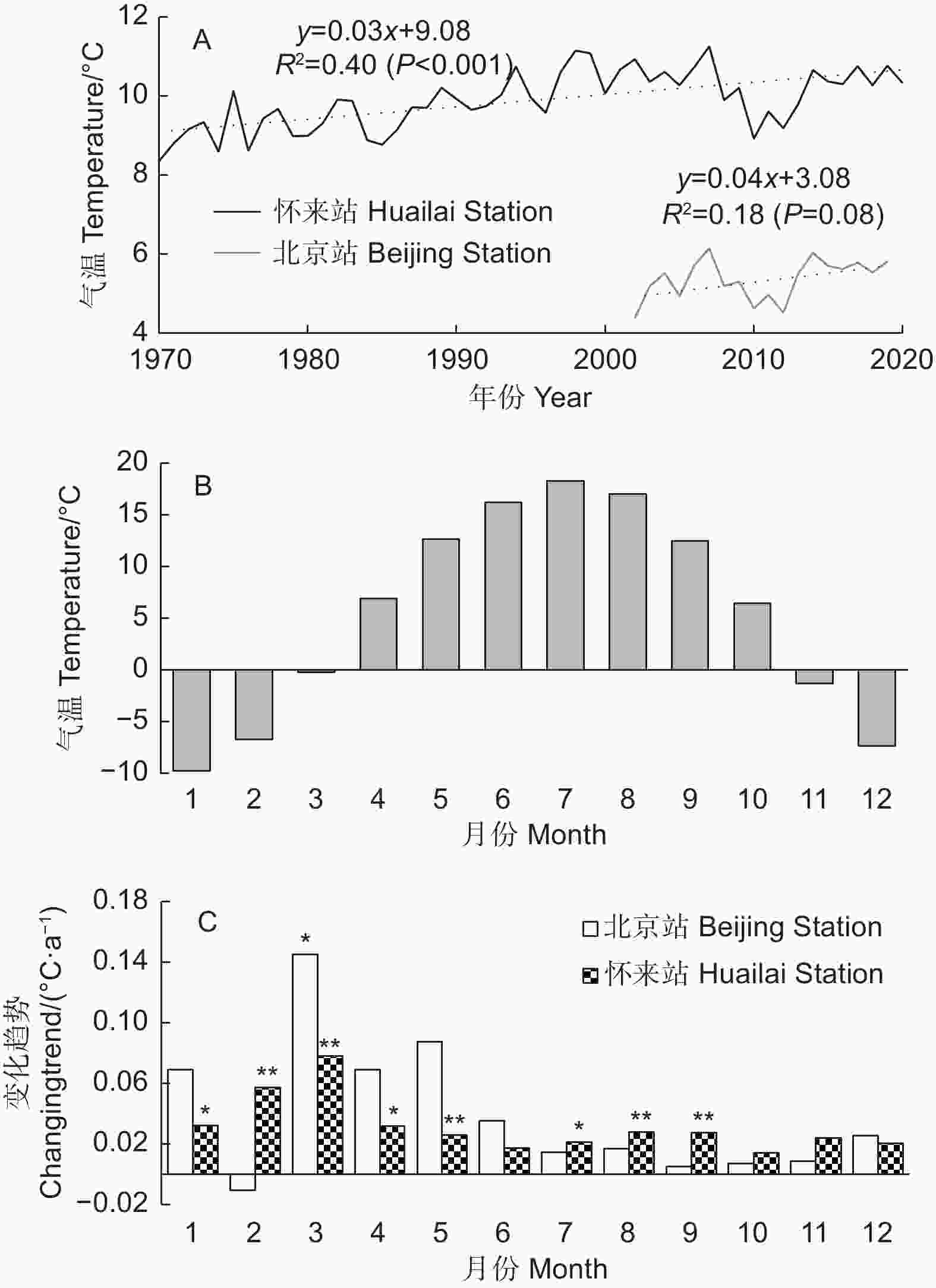
图 1 2002—2019年北京站和1970—2020年怀来站年(A)和逐月(C)平均气温变化趋势及北京站各月平均气温(B)
Figure 1. Interannual changing trends of annual(A) and monthly (C) average temperature of Beijing Station during 2002-2019 and Huailai Station during 1970-2020 as well as average monthly temperature of Beijing Station (B)
与气温一致,2003—2019年北京站大部分物种的物候期变化趋势不显著(图2A~F)。在萌芽期,有18个物种推迟(5个显著),6个物种提前(1个显著)。在其他物候期,大部分物种呈提前趋势,其中,展叶始期提前的物种最多达23个(9个显著),然后是盛花期和落叶期(19个)、始花期(16个),最后是叶变色期15个。萌芽期总体呈推迟趋势(0.41 d·a−1),与其他物候期均显著不同(图2G)。此外,展叶始期平均提前速率最快为−0.95 d·a−1,显著高于始花期(−0.40 d·a−1)和叶变色期(−0.38 d·a−1),但与盛花期(−0.65 d·a−1)和落叶期(−0.71 d·a−1)无显著差异。
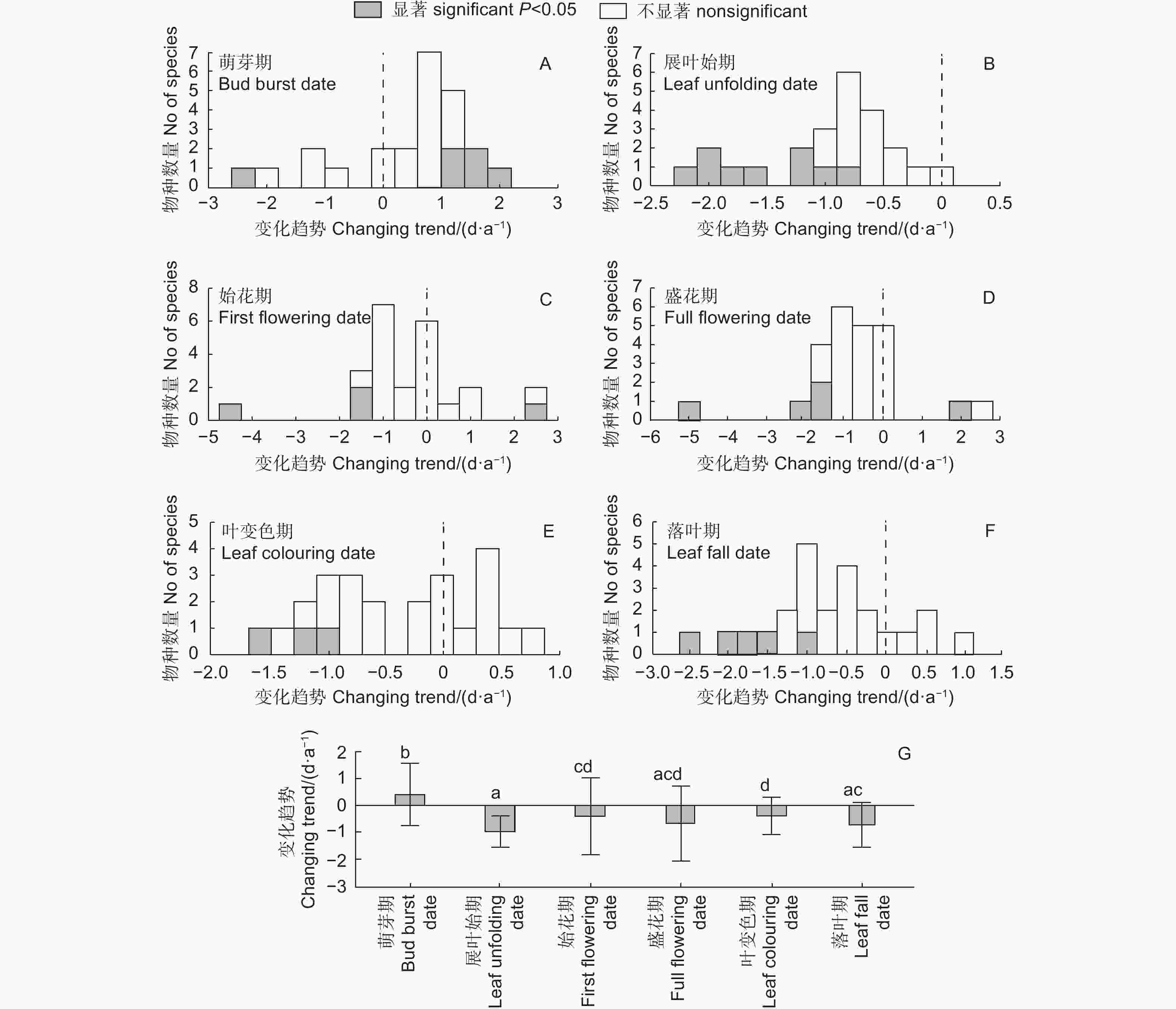
图 2 北京站2003—2019年各物候期的年际变化趋势及差异(平均值 ± 标准差,n=24)
Figure 2. Theinterannual changing trends of phenophases and their differences (mean ± SD, n=24) at Beijing Station during 2002 and 2019
-
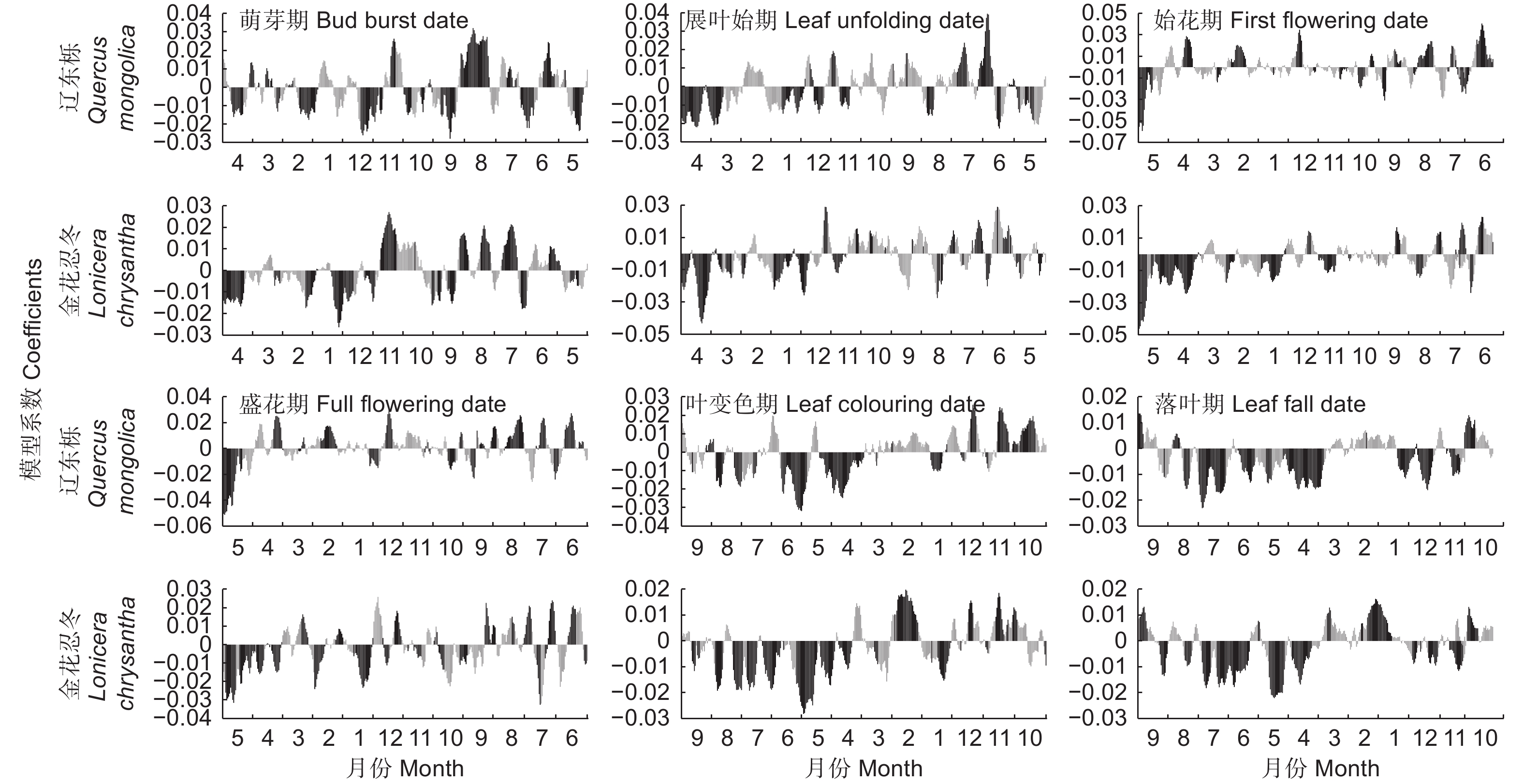
以具有代表性的乔木辽东栎和灌木金花忍冬为例,各物候期对前一年11 d滑动平均气温响应的PLS分析结果见图3。总体来说,展叶始期、始花期和盛花期对气温响应的最敏感阶段为物候期前几个月。因此,对这3种物候期从物候期前一天开始计算其最优相关时段;萌芽期、叶变色期和落叶期对气温的响应则呈现出不同的方式。萌芽期对其前3个月左右的气温响应敏感度相对较低,而对3个月(约为1月中旬)之前的气温呈现出较强负相关,持续至9月份,一般在11—12月份达到最高值。同时,萌芽期也对夏季7、8月份的气温表现出了较高的敏感度,相关系数为正值。因此,对于萌芽期,将分别从萌芽期前1 d、前90 d和前230 d分别计算其最优相关时段。叶变色期和落叶期一般对物候期前2个月(约为8、9月)气温敏感度不高,且出现正值,而对4—7月左右的气温表现出强负相关。因此,叶变色期和落叶期分别从物候期前1 d和前60 d开始计算其相关时段。
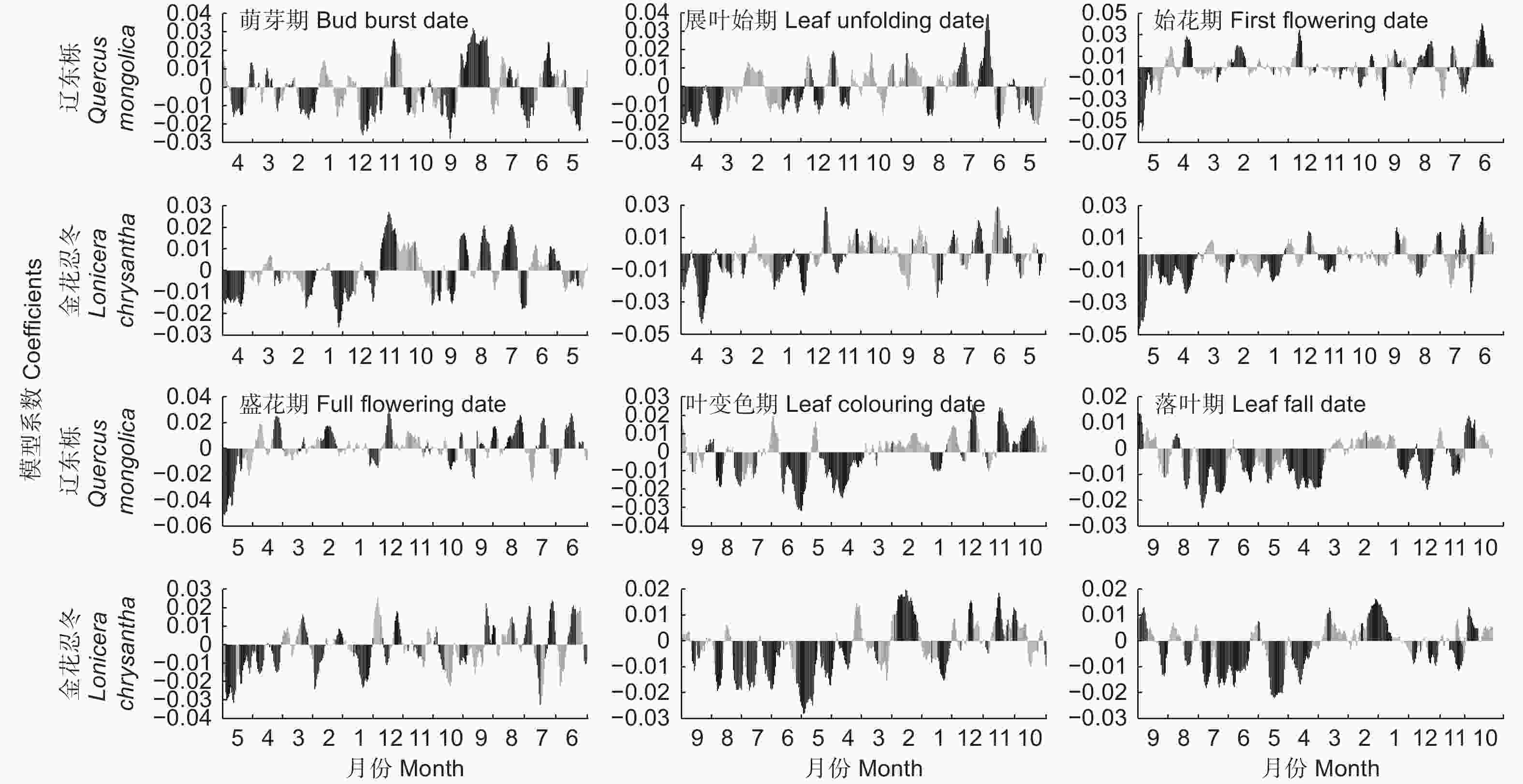
图 3 辽东栎和金花忍冬各物候期与物候期前一年11 d滑动平均气温的PLS分析结果
Figure 3. Results of Partial Least Squares regression between 11-day moving average temperature and phenophases of Quercus mongolica Mary and Lonicera chrysantha Turcz
展叶始期、始花期和盛花期对春季气温响应的最优相关时段分别为物候期前48、64、55 d(3月上中旬至4月底、5月上中旬)(表2)。萌芽期对冬春(物候期前90 d内)气温响应的最优相关时段为物候期前34 d(3月11日至4月14日),对前一年的秋冬季(物候期前90 d后)气温响应的最优相关时段平均为10月28日至下一年1月14日,而对夏季(物候期前230 d后)气温响应的最优相关时段平均为7月14日至8月27日。叶变色期和落叶期对夏秋季(物候期前60 d内)气温响应的平均最优相关时段均在8月下旬至9月中下旬之间,而对春夏季(物候期前60 d后)气温响应的最优相关时段均在4月初至7月中下旬之间。
表 2 各物候期对气温响应的最优相关时段的开始、结束日期及时长
Table 2. Temperature relevant periods of different phenophases
相关时段
Temperature relevant periods开始日期
Start dates结束日期
End dates时长/d
Length萌芽期-冬春 Bud burst date-winter and spring 3月11日 ± 23.91 d 4月14日 ± 5.77 d 34 ± 23.63 萌芽期-秋冬 Bud burst date-autumn and winter 10月28日 ± 38.93 d 1月14日 ± 5.77 d 78 ± 36.10 萌芽期-夏季 Bud burst date-summer 7月14日 ± 13.07 d 8月27日 ± 5.77 d 44 ± 12.79 展叶始期 Leaf unfolding date 3月10日 ± 20.38 d 4月27日 ± 4.82 d 48 ± 17.78 始花期 First flowering date 3月4日 ± 42.90 d 5月7日 ± 13.51 d 64 ± 39.36 盛花期 Full flowering date 3月19日 ± 27.84 d 5月13日 ± 12.86 d 55 ± 26.94 叶变色期-夏秋 Leaf coloring date-summer and autumn 8月24日 ± 12.65 d 9月12日 ± 5.71 d 19 ± 12.28 叶变色期-春夏 Leaf coloring date-spring and summer 4月4日 ± 31.90 d 7月14日 ± 5.71 d 101 ± 32.83 落叶期-夏秋 Leaf fall date-summer and autumn 8月31日 ± 14.67 d 9月23日 ± 6.31 d 23 ± 14.39 落叶期-春夏 Leaf fall date-spring and summer 4月7日 ± 32.06 d 7月25日 ± 6.31 d 110 ± 32.52 注:数值为均值 ± 标准差(n=24)
Note: Values are presented as means ± SD (n=24). -
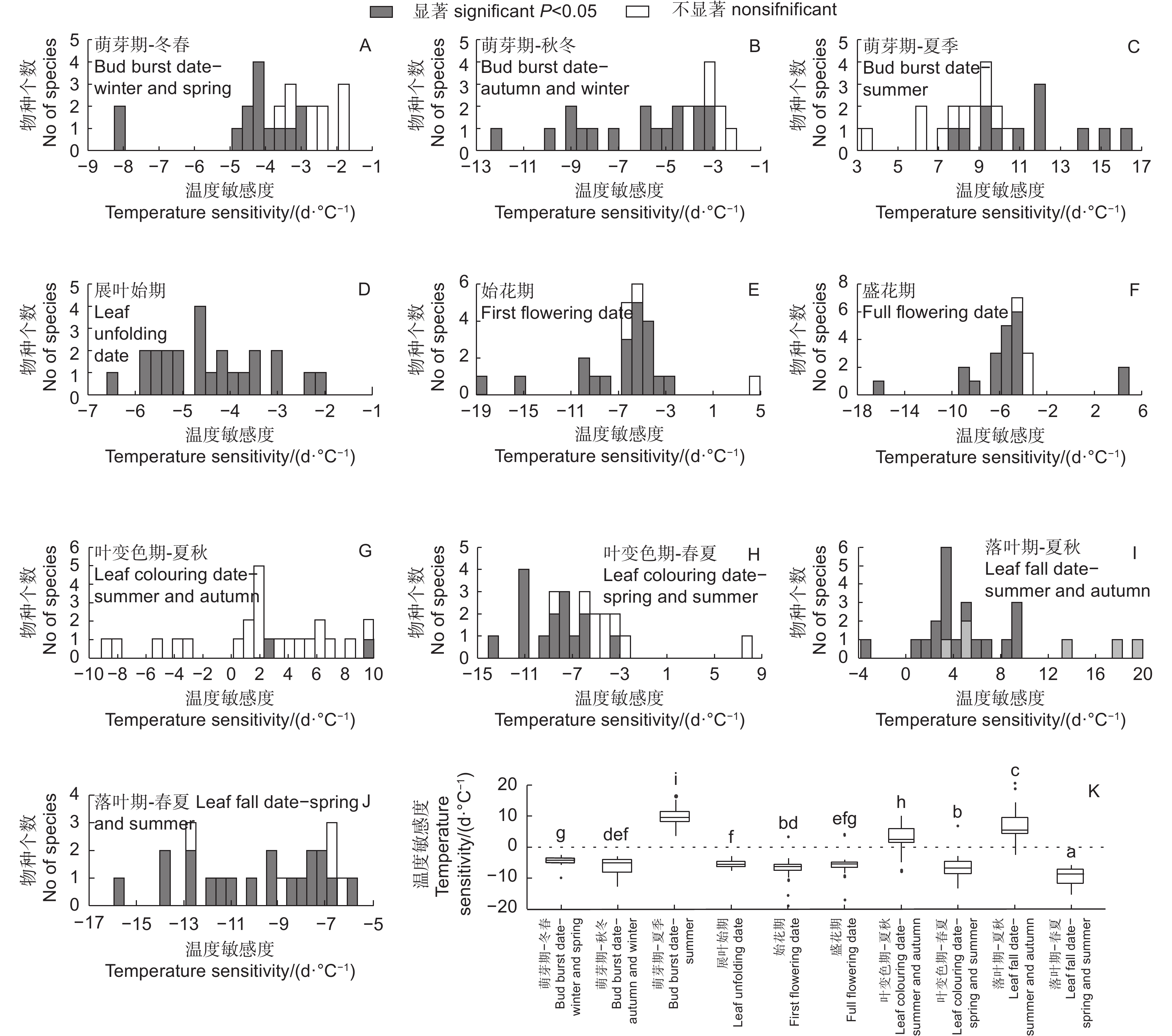
从冬春季最优相关时段气温对24种木本植物春季物候影响的显著性看,展叶始期全部物种都显著,始花期和盛花期显著的物种也较多,各有20种,而萌芽期只有14个物种显著,但秋冬季气温对萌芽期影响显著的物种较多,有17种(图4)。萌芽期和展叶始期对冬春或秋冬气温响应的敏感度均为负值,而始花期和盛花期的敏感度则绝大部分为负值,1~2种为正值,意味着气温上升使全部物种的展叶始期提前,大部分物种的花期提前。此外,萌芽期与夏季气温呈正相关(12个物种显著),意味着夏季气温升高会使下一年萌芽期推迟(图4C)。图4K表明:从春季物候对气温响应敏感度的差异看,萌芽期对冬春气温响应的敏感度(−3.71 d·℃−1)显著低于对秋冬气温的敏感度(−5.48 d·℃−1),也显著低于展叶始期和始花期对冬春气温的敏感度(分别为−4.52、−6.38 d·℃−1),但与盛花期(−5.05 d·℃−1)则无显著差异(盛花期的温度敏感度标准差较大);但萌芽期对秋冬气温的响应与展叶始期、始花期和盛花期无显著差异。萌芽期对夏季气温的响应敏感度(9.74 d·℃−1)与其他均显著不同。此外,始花期也表现出了显著高于展叶始期和盛花期的温度敏感度。
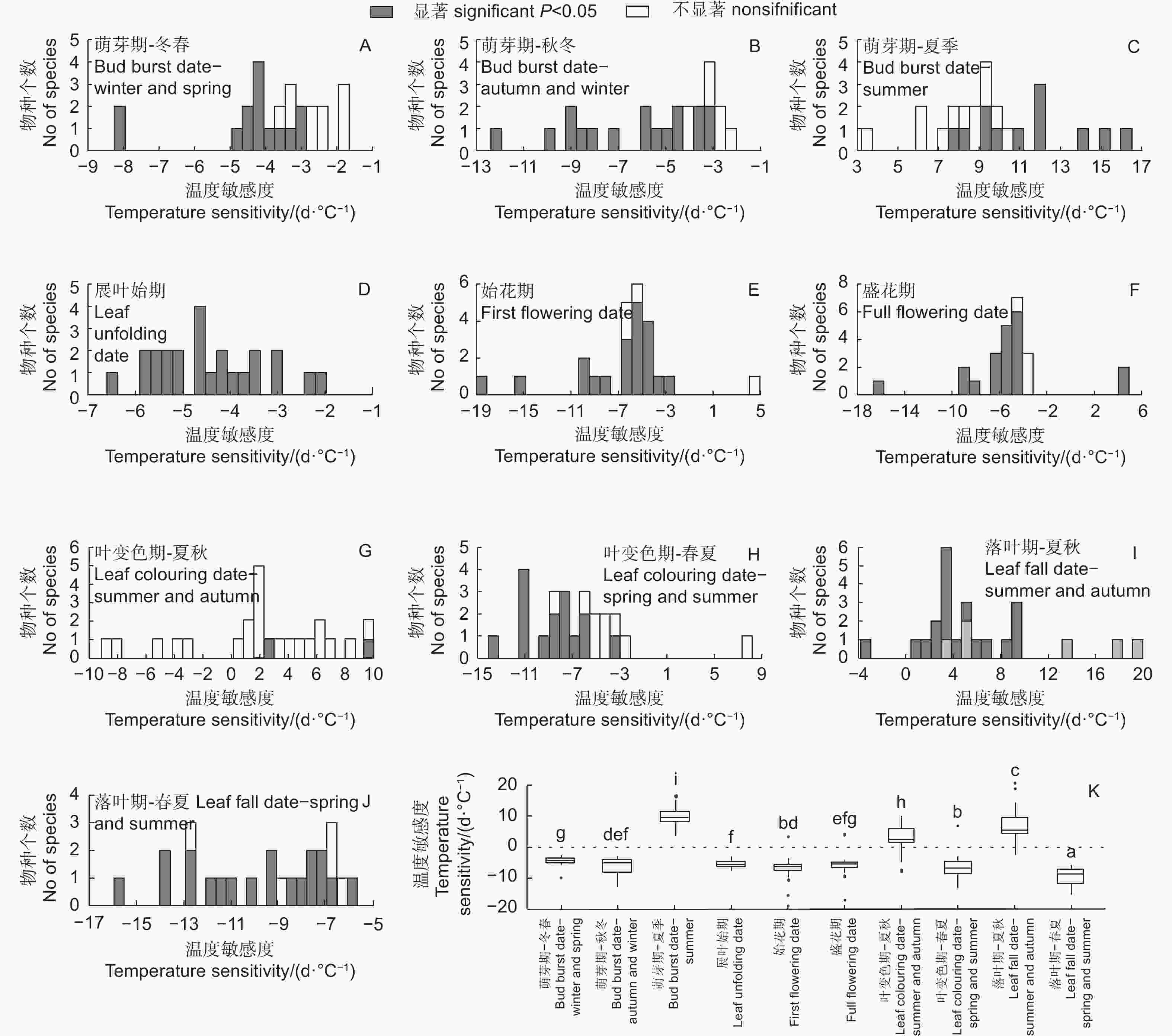
图 4 北京站24种植物各物候期对气温的响应敏感度及差异对比
Figure 4. Temperature sensitivities of different plants phenophases at Beijing Forestry Ecosystem Research Station and comparisons of their differences
对于秋季物候,夏秋季最优相关时段气温对叶变色期和落叶期的影响大部分(19和23)为正值,少数为负值,意味着总体上夏秋季气温升高使秋季物候推迟,但显著的物种较少(图4G、I);而春夏季气温对叶变色期和落叶期的影响几乎都为负值,意味着春夏季气温升高使秋季物候提前,显著的物种也较多,分别有15和19种(图4H、J)。从秋季物候对气温响应敏感度的差异看,叶变色期和落叶期对春夏季气温的敏感度分别为−7.12、−9.55 d· ℃−1,基本上均显著高于萌芽期、展叶始期、始花期和盛花期对气温的响应(除了叶变色期的敏感度与始花期无显著差异,同时也不考虑萌芽期对夏季气温的敏感度)(图4K),而这2个物候期对夏秋季气温的响应敏感度分别为2.27、5.96 d· ℃−1,与其他物候期对气温的响应均显著不同。
-
展叶始期、始花期和盛花期对物候期前几个月(冬春季)最优相关时段内的气温均表现出了最强的响应敏感度,这与前人的研究结果一致[5],说明该时段的积温累积仍是影响这3个物候期的最主要因素;而冬季气温并没有表现出对这3种物候期的显著推迟作用,说明虽然冬季气温上升会使低温累积量减少, 但目前低温累积仍能满足植物解除休眠的需要, 不会成为春季物候的限制因子,这与在自然环境下大部分温带地区的研究结论一致[32]。此外,始花期对气温的响应敏感度比展叶始期强,这与前人的研究结论相同[22-23]。可能是由于光周期对展叶始期的影响比始花期大,从而限制了展叶始期随气温上升而提前的速度,或是展叶始期所需热量累积比始花期大,因此,需要更多时间来累积热量,从而使其温度敏感度相对始花期较低[22]。但也有研究发现,展叶始期对气温的敏感度比始花期高[16],针对这一现象还需要更多深入的研究。盛花期的温度敏感性也显著低于始花期,这与Flynn等[23]对美国落基山亚高山草甸群落的研究结论相同,该群落中植物的盛花期和末花期对气温的响应敏感度均比始花期低,各花期的变化相对独立,这可能会使每个物种的物候期发生不同的变化,从而使生态系统发生较大的改变。
-
本研究发现,萌芽期对气温的响应方式与其他春季物候期不同。Basler等[13]也指出,影响芽膨大和萌芽期的因子与之后的物候期不同。植物芽在芽形成、休眠建立、休眠解除和积温促进等各阶段中对气候的响应方式不同[14],这可能都会对萌芽期产生影响。本研究中对萌芽期影响最大的是前一年的秋冬季(10月28日至下一年1月14日)气温,这段时间气温上升1 ℃,萌芽期会提前5.48 d,显著高于对冬春季(3月11日至4月14日)气温的响应(3.71 d)。对这一现象暂时还未发现相关报道。首先,秋冬季一般属于低温累积期,该时段内气温上升使萌芽期提前的原因可能是气温升高使低温累积提前得到满足,从而使萌芽期提前。对北京板栗(Castanea mollissima Blume)和冬枣(Ziziphus jujuba Mill. cv. Dongzao)始花期的研究也发现,低温累积期内气温升高使低温累积量上升[33]。该时期研究点气温较低(11月和12月平均气温分别为−1.32和−7.36 ℃,图1B),气温上升可能反而使落入有效低温的时数增多,从而促进低温的累积[34]。其次,在气温均上升1 ℃的情况下,秋冬季气温使萌芽期提前的天数比冬春季气温更多,可能是因为冷激量的满足比积温累积对萌芽期的影响更大。有研究显示,相对于积温和光周期来说,冷激是影响萌芽期最重要的因子[23]。
此外,笔者也发现有一半植物的萌芽期随前一年夏季(7、8月)气温的上升而显著推迟,但目前相关研究非常少。有研究发现,树木(尤其是寒带树木)的抗寒力会随夏季气温的升高而增加,从而使其春季抗寒力的下降过程减缓[35],这在一定程度上可能会推迟植物的春季萌芽期。很多研究表明,前一年秋季气温升高会使下一年树木萌芽期推迟,其原因可能是秋季气温升高使休眠建立的时间延迟进而推迟了下一年的春季物候[36-37]。本研究通过PLS分析也发现,秋季部分时间气温上升使春季物候推迟,但时间较短且物种较少,没有形成对萌芽期的显著影响。其原因可能是上述研究多为实验研究,实验研究多采用幼苗,幼苗和成熟树木的物候期具有很大差异[38],幼苗的生长一般到9月才停止,而成熟树木可能在7月初就停止生长了[39]。这可能会使成熟树种休眠建立的时间提前到7、8月份,从而使夏季气温的变化影响休眠的建立。萌芽期的影响因素较复杂,将来还需从多方面进行深入的探索。
-
秋季物候对气温响应最敏感的时段在春夏季(4月初至7月中下旬左右),该时段气温升高使叶变色期和落叶期提前,而夏秋季(8月下旬至9月中下旬)气温升高使2个物候期推迟。这与普遍的认知不同,普遍观点认为秋季气温和光周期是秋季物候的主要因素,从而认为气温上升会推迟秋季物候[40]。不过本研究的结论与最近的部分研究一致[15,20]。Liu等[20]认为,生长季气温升高可能会加速叶片衰老,从而使叶变色期提前,而秋季气温升高可能会影响绿叶的光合作用速率和叶绿素的转化,从而延长叶片寿命,并推迟叶变色期[41]。Zani等[15]进一步指出,春夏气温升高使春夏植物光合作用增强,加速碳汇饱和,使碳汇能力受限,从而间接导致叶片提前衰老。春夏季气温对秋季物候的提前效应抵消了部分秋季气温对秋季物候的延迟效应,秋季物候最终的变化趋势可能取决于春夏季和秋季气温的综合影响[21]。
-
2002—2019年,研究点春初(3月份)的气温上升速率至少是其他月份的2倍,4月气温的上升速率相对夏秋季也较高,这与其他地方(如欧洲)的气温变化趋势相近[16]。这使受该时段气温影响最大的展叶始期、始花期和盛花期等春季物候在研究时段内产生提前趋势。而萌芽期受秋冬(10月至下一年1月)气温的增温影响最大,然后才是冬春(3、4月)气温,二者均使萌芽期提前,但在2003—2019年秋冬季增温较小,同时夏季(7、8月)气温上升又使萌芽期以较高的速率显著推迟,最终使萌芽期呈现一定的推迟趋势。秋季物候受春夏季(4—7月)气温影响较大,其次是夏秋季(8、9月)气温。由于秋季物候对春夏季气温响应敏感度更高,且在研究时段内4—7月的气温上升幅度比8—9月份高,使夏秋季气温对秋季物候的提前作用大于秋季气温的推迟作用,最终使秋季物候呈现出一定的提前趋势。可见,植物物候对不同季节气温的响应方式和强度以及各季节气温变化存在很大差异,这将对植物物候的未来变化趋势产生影响,决定了未来的生态系统功能和森林可持续性[16,42]。
气候变化对植物物候的影响过程非常复杂,本研究采用实地观测数据,仅从气温的角度分析了植物不同物候期对气温响应的季节性差异,未来的研究还应结合实验研究、实地观测、遥感监测及模型模拟等方法,从冷激、积温、光周期和水分等多因子协同作用的角度进行深入探索(如春季物候在一定程度上受季前降水的影响[43]),以进一步加深对物候对气候变化响应的理解。
-
本研究分析了北京东灵山24种木本植物的6种物候期对季节性气温响应的最优相关时段、响应敏感度及差异。主要结论如下:展叶始期、始花期和盛花期对气温响应的最优相关时段在3月上中旬至4月底、5月上中旬。始花期对相关时段内气温的响应敏感度显著高于展叶始期和盛花期。萌芽期对气温的响应方式与上述3种物候期不同。气温对萌芽期影响最大的时段为秋冬季10月28日至下一年1月14日,其次是冬春季3月11日至4月14日和夏季7月14日至8月27日。萌芽期对秋冬季最优相关时段气温响应的敏感度显著高于其对冬春气温的敏感度,并且与展叶始期、始花期和盛花期无显著差异,而萌芽期对夏季气温的响应敏感度与其他均显著不同。与普遍的观点不同,叶变色期和落叶期对气温响应的最优相关时段为春夏季4月初至7月中下旬,其响应敏感度为负值,而2个物候期对夏秋季(8月下旬至9月中下旬)气温的响应敏感度多为正值。近50年来,研究点冬末春初(2和3月)气温上升最快,是其他月份的2倍以上,而秋冬季(10—12月)气温无显著变化。植物物候对不同季节气温的响应方式和强度以及各季节气温变化存在很大差异,这可能会对植物物候乃至整个生态系统未来的变化趋势产生重大影响。
致谢:感谢国家生态系统观测研究网络/中国生态系统研究网络科技资源服务平台,特别感谢中国科学院北京森林生态系统定位研究站提供数据支撑。
北京东灵山森林木本植物物候对气温响应的季节差异
Divergent Responses of Woody Plant Phenology to Seasonal Temperature in Dongling Mountain of Beijing
-
摘要:
目的 明确北京东灵山暖温带森林木本植物不同物候期对气候变暖响应的季节性差异。 方法 本文利用2003-2019年北京市暖温带森林24种木本植物的6种物候期数据,采用偏最小二乘回归和相关分析计算各物候期对季节气温响应的最优相关时段,并应用Wilcoxon符号秩检验对比各物候期对相关时段内气温响应敏感度的差异。 结果 展叶始期、始花期和盛花期对气温响应的最优相关时段分别为物候期前48、64、55 d(3月上中旬至4月底、5月上中旬),该时段内气温每升高1 ℃,始花期平均提前6.38 d,显著高于展叶始期(提前4.52 d)和盛花期(提前5.05 d)。萌芽期对气温的响应方式与上述3种物候期不同,其最优相关时段为秋冬季10月28日至下一年1月14日,其次是冬春季3月11日至4月14日(萌芽期前34 d)和夏季7月14日至8月27日。秋冬季相关时段气温每上升1 ℃,萌芽期提前5.48 d,显著高于冬春气温使其提前的天数(3.71 d),而夏季气温上升1 ℃使萌芽期推迟9.74 d,与其他均显著不同。与普遍的观点不同,叶变色期和落叶期对气温响应的最优相关时段为春夏季4月初至7月中下旬,而对夏秋季(8月下旬至9月中下旬)气温响应显著的物种很少。在气温上升1 ℃的情况下,春夏季相关时段气温使叶变色期和落叶期分别提前7.12和9.55 d,而夏秋季气温使其分别推迟2.27 、5.96 d。近50年,研究点冬末春初(2、3月)气温的上升速度是其他月份的2倍以上,而秋冬季(10—12月)气温无显著变化。 结论 北京东灵山森林木本植物各物候期对不同季节气温的响应方式和强度具有显著差异,且各季节气温的变化呈现不一致性,这可能会对从植物个体到生态系统的各个层次都产生深刻的影响。 Abstract:Objective To investigate the divergent responses of plant phenology to seasonal temperature in warm temperate forests in Doling Mountain, Beijing. Method By using data of 6 phenophases of 24 woody plants in warm temperate forests in Beijing during 2003 and 2019, we calculated temperature relevant periods of different phenophases to seasonal temperature by using Partial Least Square regression and correlation analysis and compared the corresponding temperature sensitivities in these periods by Wilcoxon signed-rank test. Result Temperature relevant periods of leaf unfolding dates, first flowering dates and full flowering dates were 48, 64 and 55 days before the phenological dates (early-mid March to late April or early-mid May), respectively. For every increase of 1 °C, the flowering period was advanced by an average of 6.38 days, which was significantly higher than that of the leaf development period (4.52 days earlier) and full flowering period (5.05 days earlier). The response mode of germination period to temperature was different from the above three phenophases, and its optimal correlation period was from October 28th to January 14th in autumn and winter, followed by March 11th to April 14th in winter and spring (34 days before germination) and summer from July 14 to August 27. For every temperature increase of 1 °C in autumn and winter, the date of budburst advanced by 5.48 days, which was significantly higher than the number of days (3.71 days) that the temperature in winter and spring advanced (3.71 days), while the temperature increase of 1 °C in summer delayed the budburst by 9.74 days, which was significantly different from other. The optimal correlation period of the leaf coloring and defoliation stages to the temperature response was from early April to mid-to-late July in spring and summer, and the species that responded significantly to temperature in summer and autumn (late August to mid-to-late September) rare. When the temperature increased by 1 °C, the temperature in the spring and summer related periods brought forward the leaf coloring and leaf defoliation periods by 7.12 and 9.55 days, respectively, while the summer and autumn temperatures delayed them by 2.27 and 5.96 days, respectively. In the past 50 years, the temperature rise rate in late winter and early spring (February and March) at the research site was more than twice that of other months, while the temperature in autumn and winter (October-December) had no significant change. Conclusion The divergent responses of plant phenology to seasonal temperature and seasonally asymmetric warming may exert profound impacts on plants and the entire ecosystem. -
图 1 2002—2019年北京站和1970—2020年怀来站年(A)和逐月(C)平均气温变化趋势及北京站各月平均气温(B)
Figure 1. Interannual changing trends of annual(A) and monthly (C) average temperature of Beijing Station during 2002-2019 and Huailai Station during 1970-2020 as well as average monthly temperature of Beijing Station (B)
图 2 北京站2003—2019年各物候期的年际变化趋势及差异(平均值 ± 标准差,n=24)
Figure 2. Theinterannual changing trends of phenophases and their differences (mean ± SD, n=24) at Beijing Station during 2002 and 2019
图 3 辽东栎和金花忍冬各物候期与物候期前一年11 d滑动平均气温的PLS分析结果
Figure 3. Results of Partial Least Squares regression between 11-day moving average temperature and phenophases of Quercus mongolica Mary and Lonicera chrysantha Turcz
图 4 北京站24种植物各物候期对气温的响应敏感度及差异对比
Figure 4. Temperature sensitivities of different plants phenophases at Beijing Forestry Ecosystem Research Station and comparisons of their differences
表 1 研究所选物种及相关信息
Table 1. Summary of the species used in the study and related information
乔木物种
Tree species观测年数
Number of
observed
years/a灌木物种
Shrub species观测年数
Number of
observed
years/a白桦 Betula platyphylla Sukaczev 14~15 大花溲疏 Deutzia grandiflora Bunge 15 黑桦 Betula dahurica Pall. 11 蒿柳 Salix schwerinii E. L. Wolf 15 胡桃楸 Juglans mandshurica Maxim. 11 金花忍冬 Lonicera chrysantha Turcz. 13~14 花曲柳 Fraxinus chinensis subsp.
Rhynchophylla (Hance) E. Murray11 六道木 Zabelia biflora (Turcz.) Makino 15~16 华北落叶松 Larix gmelinii var. principis
-rupprechtii (Mayr) Pilg.11 毛榛 Corylus mandshurica Maxim. & Rupr. 13~14 辽东栎 Quercus mongolica Mary 15~16 巧玲花 Syringa pubescens Turcz. 11 裂叶榆 Ulmus laciniate (Trautv.) Mayr 15~16 土庄绣线菊 Spiraea pubescens Turcz. 11 青杨 Populus cathayana Rehder 15 卫矛 Euonymus alatus (Thunb.) Siebold 13~14 山桃 Prunus davidiana (Carrière) Franch. 15~16 小花溲疏 Deutzia parviflora Bunge 15~16 山杏 Prunus sibirica L. 15~16 小叶鼠李 Rhamnus parvifolia Bunge 15 五角枫 Acer pictum Thunb. 13~14 迎红杜鹃 Rhododendron mucronulatum Turcz. 15 油松 Pinus tabuliformis Carrière 15 照山白 Rhododendron micranthum Turcz. 15  下载: 导出CSV
下载: 导出CSV
表 2 各物候期对气温响应的最优相关时段的开始、结束日期及时长
Table 2. Temperature relevant periods of different phenophases
相关时段
Temperature relevant periods开始日期
Start dates结束日期
End dates时长/d
Length萌芽期-冬春 Bud burst date-winter and spring 3月11日 ± 23.91 d 4月14日 ± 5.77 d 34 ± 23.63 萌芽期-秋冬 Bud burst date-autumn and winter 10月28日 ± 38.93 d 1月14日 ± 5.77 d 78 ± 36.10 萌芽期-夏季 Bud burst date-summer 7月14日 ± 13.07 d 8月27日 ± 5.77 d 44 ± 12.79 展叶始期 Leaf unfolding date 3月10日 ± 20.38 d 4月27日 ± 4.82 d 48 ± 17.78 始花期 First flowering date 3月4日 ± 42.90 d 5月7日 ± 13.51 d 64 ± 39.36 盛花期 Full flowering date 3月19日 ± 27.84 d 5月13日 ± 12.86 d 55 ± 26.94 叶变色期-夏秋 Leaf coloring date-summer and autumn 8月24日 ± 12.65 d 9月12日 ± 5.71 d 19 ± 12.28 叶变色期-春夏 Leaf coloring date-spring and summer 4月4日 ± 31.90 d 7月14日 ± 5.71 d 101 ± 32.83 落叶期-夏秋 Leaf fall date-summer and autumn 8月31日 ± 14.67 d 9月23日 ± 6.31 d 23 ± 14.39 落叶期-春夏 Leaf fall date-spring and summer 4月7日 ± 32.06 d 7月25日 ± 6.31 d 110 ± 32.52 注:数值为均值 ± 标准差(n=24)
Note: Values are presented as means ± SD (n=24).
下载: 导出CSV
-
[1] CLELAND E E, CHUINE I, MENZEL A, et al. Shifting plant phenology in response to global change[J]. Trends in Ecology and Evolution, 2007, 22(7): 357-365. doi: 10.1016/j.tree.2007.04.003 [2] CHUINE I, COUR P. Climatic determinants of budburst seasonality in four temperate‐zone tree species[J]. New Phytologist, 1999, 143(2): 339-349. doi: 10.1046/j.1469-8137.1999.00445.x [3] MENZEL A, FABIAN P. Growing season extended in Europe[J]. Nature, 1999, 397(6721): 659-659. doi: 10.1038/17709 [4] PIAO S L, LIU Q, CHEN A P, et al. Plant phenology and global climate change: Current progresses and challenges[J]. Global Change Biology, 2019, 25(6): 1922-1940. doi: 10.1111/gcb.14619 [5] MENZEL A, YUAN Y, MATIU M, et al. Climate change fingerprints in recent European plant phenology[J]. Global Change Biology, 2020, 26(4): 2599-2612. doi: 10.1111/gcb.15000 [6] CRAINE J M, WOLKOVICH E M, TOWNE E G, et al. Flowering phenology as a functional trait in a tallgrass prairie[J]. New Phytologist, 2012, 193(3): 673-682. doi: 10.1111/j.1469-8137.2011.03953.x [7] RICHARDSON A D, KEENAN T F, MIGLIAVACCA M, et al. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system[J]. Agricultural and Forest Meteorology, 2013, 169: 156-173. doi: 10.1016/j.agrformet.2012.09.012 [8] ETTINGER A K, GEE S, WOLKOVICH E M. Phenological sequences: How early-season events define those that follow[J]. American Journal of Botany, 2018, 105(10): 1771-1780. doi: 10.1002/ajb2.1174 [9] COOK B I, WOLKOVICH E M, PARMESAN C. Divergent responses to spring and winter warming drive community level flowering trends[J]. Proceedings of the National Academy of Sciences, 2012, 109(23): 9000-9005. doi: 10.1073/pnas.1118364109 [10] HÄNNINEN H. Boreal and Temperate Trees in a Changing Climate[M]. Dordrecht, Netherlands: Springer Business Media, 2016. [11] CHEN X, WANG L, INOUYE D. Delayed response of spring phenology to global warming in subtropics and tropics[J]. Agricultural and Forest Meteorology, 2017, 234: 222-235. [12] MENZEL A, SPARKS T H, ESTRELLA N, et al. European phenological response to climate change matches the warming pattern[J]. Global Change Biology, 2006, 12(10): 1969-1976. doi: 10.1111/j.1365-2486.2006.01193.x [13] BASLER D, KORNER C. Photoperiod and temperature responses of bud swelling and bud burst in four temperate forest tree species[J]. Tree Physiology, 2014, 34(4): 377-388. doi: 10.1093/treephys/tpu021 [14] AZEEZ A, ZHAO Y C, SINGH R K, et al. EARLY BUD-BREAK 1 And EARLY BUD-BREAK 3 control resumption of poplar growth after winter dormancy[J]. Nature Communications, 2021, 12(1): 1-12. doi: 10.1038/s41467-020-20314-w [15] ZANI D, CROWTHER T W, MO L, et al. Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees[J]. Science, 2020, 370(6520): 1066-1071. doi: 10.1126/science.abd8911 [16] ZOHNER C M, RENNER S S. Ongoing seasonally uneven climate warming leads to earlier autumn growth cessation in deciduous trees[J]. Oecologia, 2019, 189: 549-561. doi: 10.1007/s00442-019-04339-7 [17] FU Y S, PIAO S L, DELPIERRE N, et al. Larger temperature response of autumn leaf senescence than spring leaf-out phenology[J]. Global Change Biology, 2018, 24(5): 2159-2168. doi: 10.1111/gcb.14021 [18] ESTRELLA N, MENZEL A. Responses of leaf colouring in four deciduous tree species to climate and weather in Germany[J]. Climate Research, 2006, 32(3): 253-267. [19] MENZEL A, ESTRELLA N, HEITLAND W, et al. Bayesian analysis of the species-specific lengthening of the growing season in two European countries and the influence[J]. International Journal of Biometeorology, 2008, 52: 209-218. doi: 10.1007/s00484-007-0113-8 [20] LIU G, CHEN X, ZHANG Q, et al. Antagonistic effects of growing season and autumn temperatures on the timing of leaf coloration in winter deciduous trees[J]. Global Change Biology, 2018, 24(8): 3537-3545. doi: 10.1111/gcb.14095 [21] CHEN L, HÄNNINEN H, ROSSI S, et al. Leaf senescence exhibits stronger climatic responses during warm than during cold autumns[J]. Nature Climate Change, 2020, 10(8): 777-780. doi: 10.1038/s41558-020-0820-2 [22] GENG X, FU Y H, PIAO S, et al. Higher temperature sensitivity of flowering than leaf‐out alters the time between phenophases across temperate tree species[J]. Global Ecology and Biogeography, 2022, 31(5): 901-911. doi: 10.1111/geb.13463 [23] FLYNN D F B, WOLKOVICH E M. Temperature and photoperiod drive spring phenology across all species in a temperate forest community[J]. New Phytologist, 2018, 219(4): 1353-1362. doi: 10.1111/nph.15232 [24] CARADONNA P J, ILER A M, INOUYE D W. Shifts in flowering phenology reshape a subalpine plant community[J]. Proceedings of the National Academy of Sciences, 2014, 111(13): 4916-4921. doi: 10.1073/pnas.1323073111 [25] DUCHENNE F, THÉBAULT E, MICHEZ D, et al. Phenological shifts alter the seasonal structure of pollinator assemblages in Europe[J]. Nature Ecology and Evolution, 2020, 4(1): 115-121. [26] KUDO G, IDA T Y. Early onset of spring increases the phenological mismatch between plants and pollinators[J]. Ecology, 2013, 94(10): 2311-2320. doi: 10.1890/12-2003.1 [27] XIA J Y, CHEN J Q, PIAO S L, et al. Terrestrial carbon cycle affected by non-uniform climate warming[J]. Nature Geoscience, 2014, 7(3): 173-180. doi: 10.1038/ngeo2093 [28] GUO L, DAI J, WANG M, et al. Responses of spring phenology in temperate zone trees to climate warming: a case study of apricot flowering in China[J]. Agricultural and Forest Meteorology, 2015, 201: 1-7. [29] LUEDELING E, GASSNER A. Partial least squares regression for analyzing walnut phenology in California[J]. Agricultural and Forest Meteorology, 2012, 158: 43-52. [30] WOLD S, SJOSTROM M, ERIKSSON L. PLS-regression: a basic tool of chemometrics[J]. Chemometrics and Intelligent Laboratory Systems, 2001, 58(2): 109-130. doi: 10.1016/S0169-7439(01)00155-1 [31] 陶泽兴, 葛全胜, 戴君虎, 等. 1980—2018年中国东部主要木本植物展叶始期的温度相关时段变化[J]. 生态学报, 2020, 40(21):7777-7789. [32] CHEN X, AN S, INOUYE D W, et al. Temperature and snowfall trigger alpine vegetation green‐up on the world's roof[J]. Global Change Biology, 2015, 21(10): 3635-3646. doi: 10.1111/gcb.12954 [33] GUO L, DAI J H, RANJITKAR S, et al. Chilling and heat requirements for flowering in temperate fruit trees[J]. International Journal of Biometeorology, 2014, 58(6): 1195-1206. doi: 10.1007/s00484-013-0714-3 [34] LUEDELING E, BROWN P H. A global analysis of the comparability of winter chill models for fruit and nut trees[J]. International Journal of Biometeorology, 2011, 55: 411-421. doi: 10.1007/s00484-010-0352-y [35] BALDUMAN L M, AITKEN S N, HARMON M, et al. Genetic variation in cold hardiness of Douglas-fir in relation to parent tree environment[J]. Canadian Journal of Forest Research, 1999, 29(1): 62-72. doi: 10.1139/x98-180 [36] BEIL I, KREYLING J, MEYER C, et al. Late to bed, late to rise—Warmer autumn temperatures delay spring phenology by delaying dormancy[J]. Global Change Biology, 2021, 27(22): 5806-5817. doi: 10.1111/gcb.15858 [37] WANG F, ZHANG R, LIN J, et al. High autumn temperatures increase the depth of bud dormancy in the subtropical Torreya grandis and Carya illinoinensis and delay leaf senescence in the deciduous Carya[J]. Trees, 2022, 36: 1-13. doi: 10.1007/s00468-021-02164-1 [38] VITASSE Y. Ontogenic changes rather than difference in temperature cause understory trees to leaf out earlier[J]. New Phytologist, 2013, 198(1): 149-155. doi: 10.1111/nph.12130 [39] PERRY T O. Dormancy of Trees in Winter: Photoperiod is only one of the variables which interact to control leaf fall and other dormancy phenomena[J]. Science, 1971, 171(3966): 29-36. doi: 10.1126/science.171.3966.29 [40] DELPIERRE N, DUFRÊNE E, SOUDANI K, et al. Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France[J]. Agricultural and Forest Meteorology, 2009, 149(6-7): 938-948. doi: 10.1016/j.agrformet.2008.11.014 [41] ARCHETTI M, RICHARDSON A D, O'KEEFE J, et al. Predicting climate change impacts on the amount and duration of autumn colors in a New England Forest[J]. PLoS One, 2013, 8(3): e57373. doi: 10.1371/journal.pone.0057373 [42] YAN T, SONG H H, ZENG H. Spring phenophases of larch are more sensitive to spring warming than to year-round warming: Results of a seasonally asymmetric warming experiment[J]. Forest Ecology and Management, 2020, 474: 118368. doi: 10.1016/j.foreco.2020.118368 [43] 李耀斌, 张远东, 顾峰雪等. 中国温带草原和荒漠区域春季物候的变化及其敏感性分析[J]. 林业科学研究, 2019, 32(4):1-10. doi: 10.13275/j.cnki.lykxyj.2019.04.001 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2571
- HTML全文浏览量: 1206
- PDF下载量: 76
- 被引次数: 0
