

-
传统森林经营多以单一功能为目标,如追求木材生产的用材林经营就较少关注其它多种功能。由于在森林多种功能之间常存在矛盾,追求某种功能时可能会降低其它功能[1-2],这就难以满足社会日益快速增长的森林多种功能需求。因此,以多功能利用为特征的森林多功能经营应运而生,但需解决的技术瓶颈首先是尚不清楚森林各垂直层之间的关系,因而难以通过适当经营来调控乔木层结构并形成和维持林分的合理垂直结构,进而实现森林多功能利用[3-4]。
辽东栎(Quercus wutaishanica Mayr)次生林是黄土高原及宁夏六盘山区的典型天然林,研究其林分垂直结构特征变化规律可为其多功能经营提供科学依据。以前有关辽东栎林的研究多关注其分布范围、自然更新[5]、物种多样性和区系组成[6]、林分结构和空间分布[7]、细根生物量[8]等,有关林分生物量和碳储量层次组成的研究多是仅关注某个方面[9],而对林分结构特征(如林分密度和郁闭度等)如何影响各植物层生物量以及如何在此基础上开展多功能管理的系统研究还很少[10],尤其在有大量辽东栎林自然分布的六盘山区,此类研究还未见报道,这严重限制着辽东栎林的多功能经营技术发展。
本研究通过调查研究辽东栎林的垂直分层(乔木层、林下植物层、枯落物层)生物量组成及其随林分结构的变化规律,并权衡确定能使各层生物量均保持在较高水平的最佳林分结构,以便为辽东栎林多功能管理提供科学依据。
-
辽东栎林是六盘山区的地带性植被类型之一,在20世纪50年代曾进行过大规模采伐,在20世纪80年代建立自然保护区后停止采伐,现存几乎都是采伐后萌蘖更新形成的次生林,占自然保护区天然次生林面积的31.5%。伴生乔木树种主要有白桦(Betula platyphylla Suk.)、山杨(Populus davidiana Dode.)、少脉椴(Tilia paucicostata Maxim.)。林下灌木主要有陕西荚蒾(Viburnum schensianum Maxim.)、水荀子(Cotoneaster multiflora Bunge.)、绣球绣线菊(Spiraea blumei G.Don)、山楂(Crataegus pinnatifida Bunge.)、北京忍冬(Lonicera elisae Franch.)、榛(Corylus heterophylla Fisch.)、桦叶荚蒾(Viburnum betulifolium Batal.)、鞘柄菝葜(Smilax stans Maxim.)等;林下草本主要有淫羊藿(Epimedium brevicornu Maxim.)、点叶薹草(Carex hancockiana Maxim.)、糙苏(Phlomis umbrosa Turcz.)等。
宁夏六盘山自然保护区(35º27′22.5″~35º33′29.7″ N,106º12′10.6″~106º16′30.5″ E)面积904 km2,是六盘山森林分布的核心区,在自然地理区划上处于暖温带半湿润区向半干旱区的过渡带。保护区海拔变化在1 700~2 931 m,年均气温5.8 ℃,年最高气温30. 0 ℃,年极端最低气温−26. 0 ℃,无霜期90~130 d;土壤类型主要为石灰性灰褐土,总体特征是土层薄、质地粗。年均降水量557 mm,由南向北逐渐递减,降水集中于6—9月[11]。
-
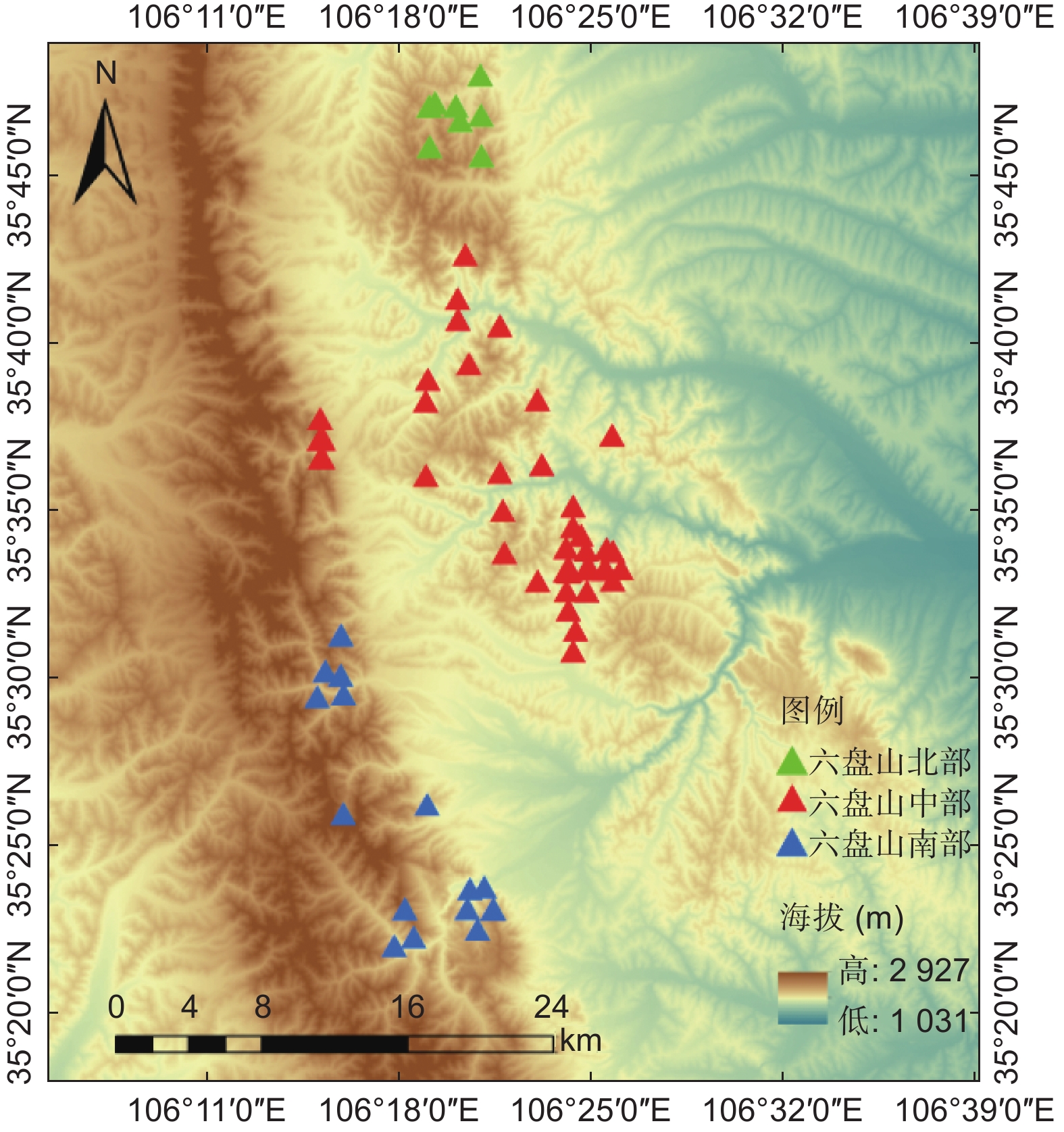
本研究所用样地共70个,包括2015年的24个25.82 m × 25.82 m的国家森林资源清查固定样地、2021年作者调查的15个30 m × 30 m的森林生态水文研究固定样地、2022年作者面上调查的31个20 m × 20 m的临时样地。为尽可能真实反映六盘山区辽东栎林生长情况,将六盘山辽东栎林分布区按年均降水量分为500~600(北部)、600~700(中部)、>700 mm(南部),按坡向分为阴坡、半阴坡、半阳坡,按海拔划分为1 800~1 950、1 950~2 100、2 100~2 250、>2 250 m等区间,样地尽可能均匀分布在这些环境条件区间的组合类型中(见图1)。调查样地基本都为辽东栎纯林,仅有少量白桦、山杨和椴树混生,其株数占比不足8%。
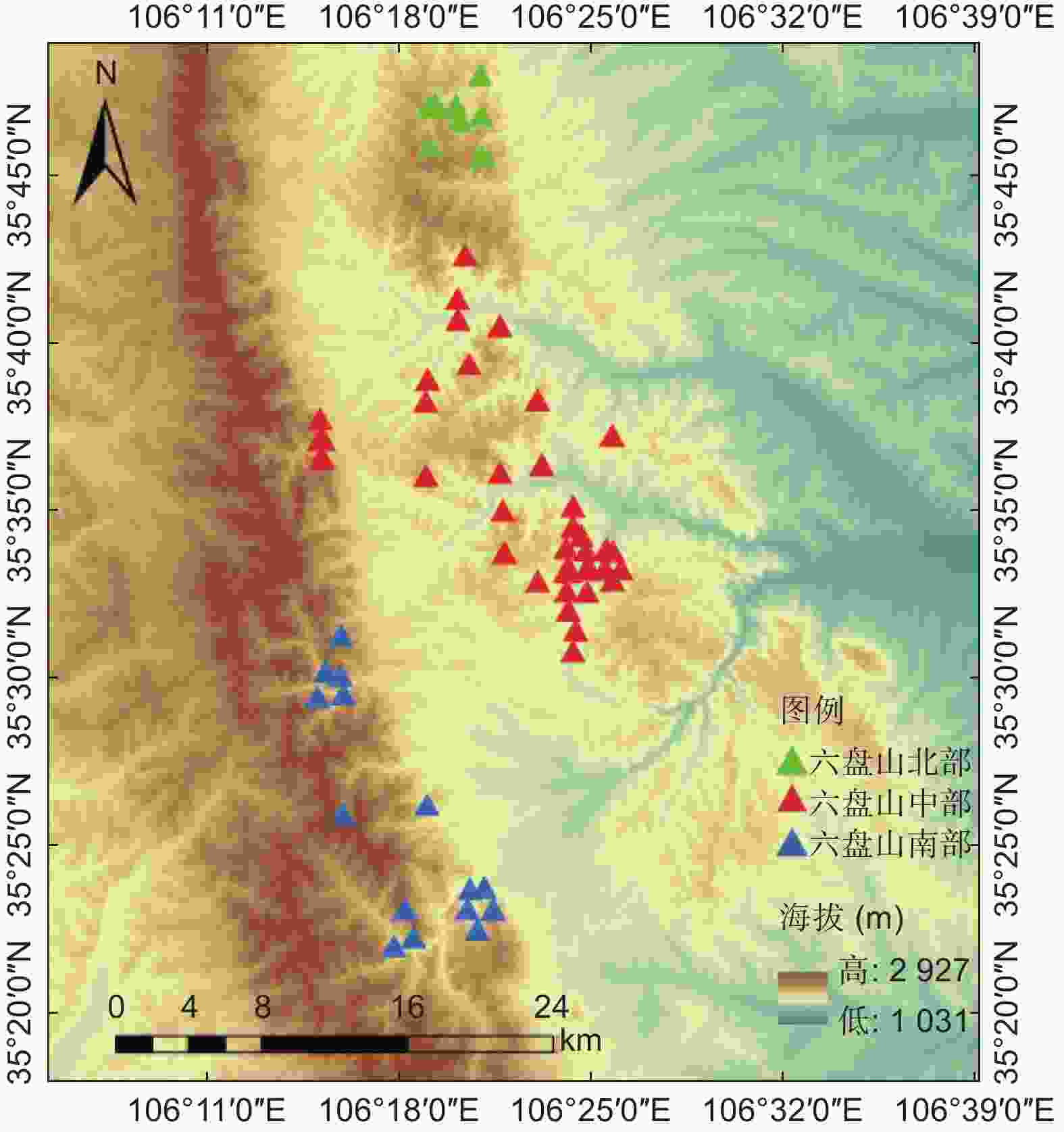
图 1 宁夏六盘山区辽东栎林调查样地位置布设
Figure 1. Sample plots layout of Quercus wutaishanica forest stands in Liupan Mountains Area of Ningxia
对样地内所有乔木进行每木检尺,并记录样地基本信息;同时沿样地对角线布设3个面积5 m × 5 m的灌木典型样方,调查样方内胸径<5 cm的乔木幼苗和灌木的种类、数量、高度、地径(距地面10 cm处的直径)、冠幅、覆盖度等;并在样地四角及中间设置5个面积1 m × 1 m的草本典型样方,调查草的种类、数量、平均高度、多度等,同时在每个草本样方内布设面积30 cm × 30 cm的枯落物典型样方,即每个样地内有5个枯落物样方,调查枯落物层厚度和生物量等。
-
对每个样地,分乔木层、林下植物层(灌木层、草本层)、枯落物层计算其生物量,用于后续分析。
-
在计算辽东栎及少量白桦、椴树的单株生物量时,应用在六盘山区基于解析木调查建立的单株生物量方程[12];对出现很少的山杨,其单株生物量计算采用在七老图岭山建立的方程[13](表1)。将样地内所有单株乔木生物量求和,并依样地面积求得乔木层生物量(式1)。
表 1 辽东栎林乔木单株生物量计算模型
Table 1. The relations of single tree biomass of Quercus wutaishanica secondary forest
树种
Tree species拟合方程
Fitting equations方程来源
Equation source辽东栎 Quercus wutaishanica B=0.732 0(D) 1.968 1 文献[12] 白桦 Betula platyphylla B=0.668 4(D) 1.965 6 少脉椴 Tilia paucicostata B=0.291 6(D) 2.146 8 山杨Populus tremula B=0.262 4(D) 2.215 0 文献[13] 注:B为乔木单株整体生物量(kg·株−1),D为乔木单株胸径(cm)。
Note: B is the overall biomass of a single tree (kg·tree−1), and D is the diameter at breath height (cm) of single tree.$ {W}_{\mathrm{乔}}=\frac{\sum _{i=1}^{n}{W}_{\mathrm{乔}i}}{{S}_{\mathrm{乔}}} $
(1) 式中:
$ {W}_{\mathrm{乔}} $ 为样地乔木层生物量(t∙hm−2),$ {W}_{\mathrm{乔}i} $ 为样地内第i株乔木生物量(t∙hm−2),$ {S}_{\mathrm{乔}} $ 为样地面积(hm2),n为样地内乔木株数。 -
不同灌木种的生物量方程差异较大。因此,作者构建了本区域辽东栎林下6种主要灌木的单株生物量方程。在2021年8月进行灌木调查时,将全部样地中的植株根据高(H)和地径(D),分为3级,每级标准木选3~6株,最后共选择绣球绣线菊16株、山楂8株、北京忍冬8株、水荀子6株、榛15株、桦叶荚蒾14株。采用收获法测定单株生物量,将样株在离地面5 cm左右处锯断,将干、枝、叶(含花、果)分离,再将树根尽可能全部挖出,全部带回室内置于70 ℃烘箱中烘干48 h达到恒质量后,对每种灌木样株的各器官分别称取干质量。利用这些数据,建立了6种灌木的单株生物量与地径的回归方程(表2),用于计算林下灌木生物量。对样地内其它灌木种类,其单株生物量计算采用已发表的六盘山区混合灌木生物量模型[14]。将各样地的3个灌木样方内的所有灌木单株生物量求和并依据样方总面积求得该样地的灌木层生物量(式2)。
表 2 辽东栎林下主要灌木种类的单株生物量计算模型
Table 2. The relations of single plant biomass of main shrub species in Quercus wutaishanica
灌木种类
Shrub species拟合方程
Fitted equations决定系数
R2方程来源
Equation source绣球绣线菊(Spiraea blumei) B=0.498(D)1.653 2 0.925 8 依据本研究的灌木生物量调查数据建立 山楂(Crataegus pinnatifida) B=0.104 6D2−0.045 1D + 0.030 9 0.981 5 北京忍冬(Lonicera elisae) B=0.075 6(D)1.257 0.832 0 水荀子(Cotoneaster multiflora) B=0.062 2(D)1.693 9 0.982 8 榛(Corylus heterophylla) B=0.045 2(D)1.743 0.907 7 桦叶荚蒾(Viburnum betulifolium) B=−0.000 6D2 + 0.176 3D−0.116 1 0.908 7 其它灌木Other shrub species B地上=1.890 7ln D−2.496 2 0.862 1 文献[14] B根=0.896 6 lnD−2.263 3 0.568 5 注:B为灌木单株生物量(kg·株−1);D为灌木地径(cm)。
Note: B is the overall biomass of a single shrub (kg·plant−1), and D is the ground diameter (cm) of single shrub plant.$ {W}_{\mathrm{灌}}={\sum }_{1}^{3}\frac{\sum _{i=1}^{n}{W}_{\mathrm{灌}-\mathrm{单}}}{3\times {S}_{\mathrm{灌}}} $
(2) 式中:
$ {W}_{\mathrm{灌}} $ 为样地灌木层生物量(t∙hm−2),$ {W}_{\mathrm{灌}-\mathrm{单}} $ 为灌木样方内第i 株生物量(t∙hm−2),$ {S}_{\mathrm{灌}} $ 为灌木样方面积(hm2),n为各样方内灌木株数。 -
由于辽东栎林下草本种类繁多,测定草本生物量时未分种类,而是采用全部收获法。在2021年8月,对15个辽东栎林样地内的75个1 m × 1 m草本样方,将草本植株全部收获,包括地上和地下部分,实测鲜质量后带回实验室在70 ℃烘箱中烘24 h后称取干质量,选用幂函数建立了草本总干质量(包括地上和地下生物量)与地上鲜质量的回归关系(式3),用于计算2022年31个样地内各样方草本总干质量。将各样地内5个草本样方的生物量求和并根据样方总面积计算各样地的草本层生物量(式4)。基于2021和2022年草本的调查数据,选用指数函数建立了草本层生物量与各样方草本平均高度和盖度的回归关系(式5),用于计算2015年24个样地内草本层生物量。
$ W_{{\text {草 }}-{\text {样方}}}=1.032\;2 \times W_{{\text {地上 }}-{\text {鲜}}}^{1.012\;5}\quad R^2=0.91$
(3) $ {W}_{\mathrm{草}}=\sum _{1}^{5}\frac{{W}_{\mathrm{草}-\mathrm{样}\mathrm{方}}}{1\;000\times 5\times {S}_{\mathrm{草}}} $
(4) $ {W}_{\mathrm{草}}=0.030 \;9\times {\mathrm{e}}^{0.001\;9\times {C}_{\mathrm{草}}\times {H}_{\mathrm{草}}}\quad R^2=0.81 $
(5) 式中:
$ {W}_{\mathrm{草}-\mathrm{样}\mathrm{方}} $ 、$ {W}_{\mathrm{地}\mathrm{上}-\mathrm{鲜}} $ 和$ {W}_{\mathrm{草}} $ 分别表示样方内的草本总干质量(kg)、地上鲜质量(kg)和样地的草本层生物量(t∙hm−2),$ {S}_{\mathrm{草}} $ 为草本样方面积(hm2),$ {C}_{\mathrm{草}} $ 为样方草本盖度(%),$ {H}_{\mathrm{草}} $ 为样方内草本平均高度(cm),1 000为生物量单位转换系数。因调查样地内草本生物量较低,将其和灌木层合并,称为林下植物层生物量(
$ {W}_{\mathrm{林}\mathrm{下}} $ ,t∙hm−2)(式6)。$ {W}_{\mathrm{林}\mathrm{下}}={W}_{\mathrm{灌}} + {W}_{\mathrm{草}} $
(6) -
将30 cm × 30 cm样方内枯落物全部收集,带回实验室在75 ℃烘箱内烘24 h后称干质量,计算各样地内5个枯落物样方的生物量平均值,并依样方总面积计算各样地的枯落物层生物量(式7),然后建立其与枯落物层厚度的统计关系,用于估算枯落物层生物量(式8)。
$ {W}_{\mathrm{枯}}=\frac{{\sum }_{1}^{5}{W}_{\mathrm{枯}-\mathrm{样}\mathrm{方}}}{1\;000\times 5\times {S}_{\mathrm{枯}}} $
(7) $ {W}_{\mathrm{枯}}=2.940 \;1\times T + 4.997\; 6 \quad R^2=0.75$
(8) 式中:
$ {W}_{\mathrm{枯}} $ 为样地枯落物层生物量(t∙hm−2),$ {W}_{\mathrm{枯}-\mathrm{样}\mathrm{方}} $ 为某样方内枯落物干质量(kg),$ {S}_{\mathrm{枯}} $ 为枯落物样方面积(hm2),$ T $ 为枯落物层厚度(cm),1 000为生物量单位转换系数。 -
由于林分生长指标在野外同时受多因子的复杂影响,很难区分出各单一因子的影响。因此,使用可在一定范围内排除其它因子影响的上外包线法,分析单因子作用[15]。具体做法是:将各因变量Y对应各单一自变量因子X的数据绘成散点图,根据自变量变化范围将其分成若干区段,在每段内选取高于全体数值平均值一倍偏差的数据点[16],或在一些数据密度较低的区段中直接选择最高值的数据,用这些被选取的数据点拟合因变量对单因子的响应关系式,确定响应函数类型,并能在一定程度上定量表征响应关系。
利用Microsoft Excel 2016对六盘山各分区的辽东栎林样地的生长及生境指标求取算术平均值;对各样地生物量总和统计后利用Origin 2019进行非线性回归分析及制图。
-
六盘山区辽东栎林主要生长在海拔1 789~2 420 m的阴坡和半阴坡,少量在半阳坡,阳坡几乎没有。从南到北的样地立地特征存在差异(表3),如年降水量减少,土层厚度降低;平均林龄为38~45 a,各分区差异不大。样地的平均胸径、树高、郁闭度都是从南向北表现为降低,林分密度表现为增加。
表 3 六盘山各分区内辽东栎林样地的平均立地和结构特征
Table 3. Site and structure characteristics of the Quercus wutaishanica stand plots in different sub-regions
项目
Item六盘山南部
South of Liupan
Mountains六盘山中部
Middle of Liupan
Mountains六盘山北部
North of Liupan
Mountains海拔 Altitude/m 1 960~2 280 1 789~2 420 2 030~2 260 土层厚度 Soil thickness/cm 78 ± 24 64 ± 20 58 ± 12 坡度 Slope gradient/(°) 34 ± 7 30 ± 8 37 ± 2 林分密度 Tree density/(株∙hm−2) 1 037 ± 494 1 055 ± 337 1 314 ± 388 郁闭度 Canopy density 0.71 ± 0.12 0.68 ± 0.15 0.57 ± 0.08 林龄 Stand age/a 38 ± 17 45 ± 10 38 ± 4 平均树高 Average tree height/m 12.3 ± 3.8 8.2 ± 2.7 6.6 ± 0.8 平均胸径 Average DBH/cm 17.3 ± 4.9 13.3 ± 3.8 11.4 ± 1.7 -
六盘山区辽东栎林的生物量为乔木层>枯落物层>灌木层>草本层(表4),所有样地的各层生物量占比分别为86.92%、10.85%、1.77%、0.46%。生物量存在空间差异,总生物量为南部最高(234.89 ± 94.16 t∙hm−2),分别是中部和北部的1.6倍和2.1倍;乔木层和枯落物层生物量也是南部>中部>北部;但林下植物层生物量为中部最高(4.15 ± 3.00)t∙hm−2,分别是北部的1.14倍和南部的1.51倍。
表 4 六盘山各分区内辽东栎林样地的各植物层生物量
Table 4. The biomass of different layers of Quercus wutaishanica forests in different sub-regions t∙hm−2
植物层
Plant layers六盘山南部
South of Liupan
Mountains六盘山中部
Middle of Liupan
Mountains六盘山北部
North of Liupan
Mountains平均值
Average乔木层 Overstory tree layer 209.28 ± 92.21 124.50 ± 61.39 94.59 ± 64.73 146.39 ± 79.67 林下植物层 Understory plant layer 2.75 ± 1.35 4.15 ± 3.00 3.65 ± 2.00 3.76 ± 2.67 灌木层 Shrub layer 2.44 ± 1.36 3.11 ± 2.32 3.28 ± 1.95 2.98 ± 2.11 草本层 Herb layer 0.31 ± 0.31 1.04 ± 2.22 0.37 ± 0.32 0.78 ± 1.84 枯落物层 Humus layer 22.86 ± 6.45 16.70 ± 7.74 15.70 ± 3.52 18.27 ± 7.67 总生物量 Total biomass 234.89 ± 94.16 145.35 ± 64.05 113.94 ± 68.06 168.42 ± 82.79 -
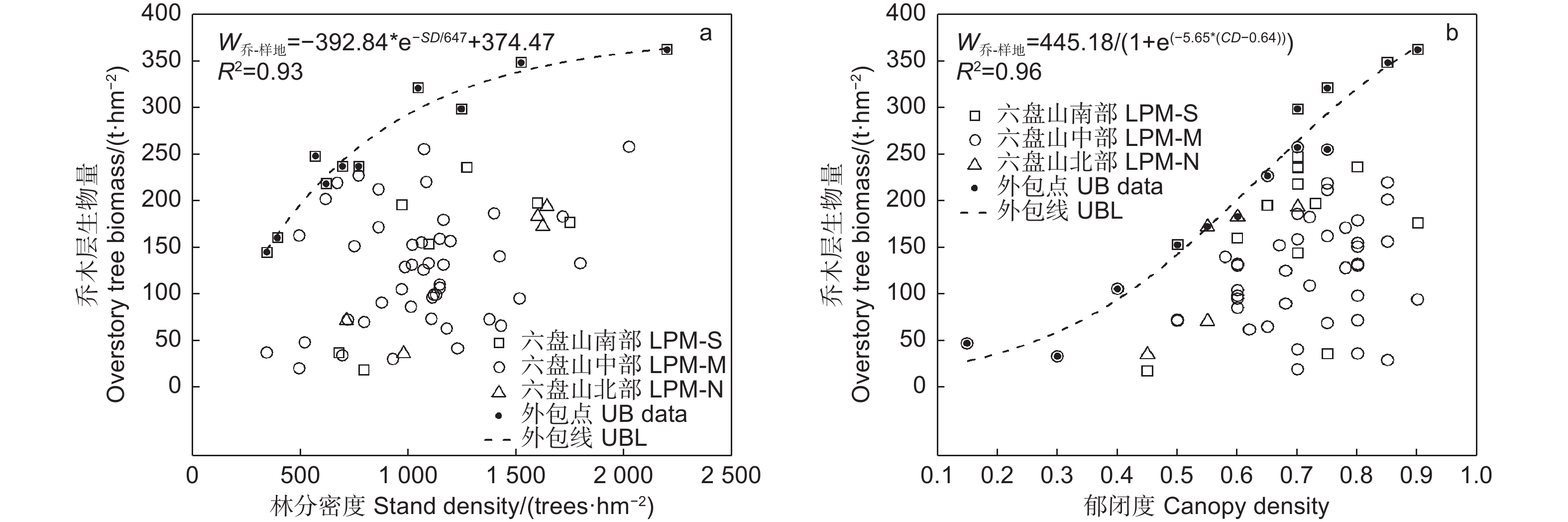
由图2中的外包线可知,乔木层生物量随林分密度增加先快速增加,在密度达1 050 株∙hm−2(对应上外包线乔木层生物量296.9 t∙hm−2)后增加逐渐变缓并趋平稳(图2a);随郁闭度增加呈“S”型增长模式,郁闭度为0.4以下时缓慢增长,0.4~0.75(对应上外包线乔木层生物量292.3 t∙hm−2)时快速增加,0.75之后增加趋缓(图2b)。
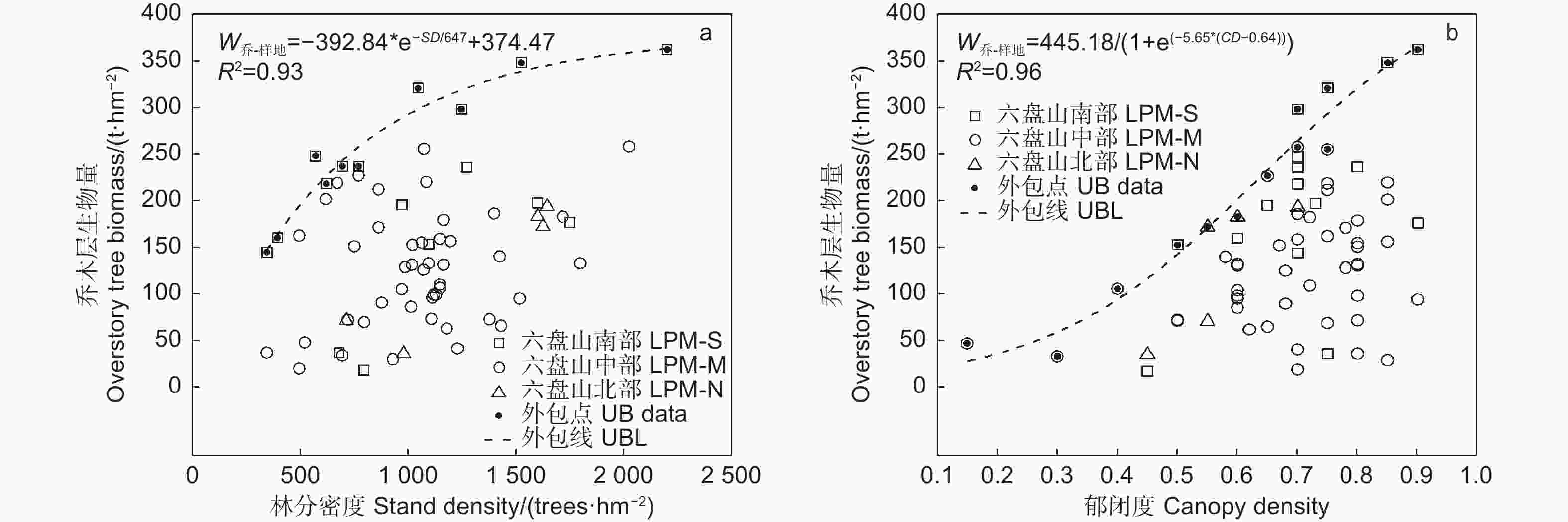
图 2 辽东栎林乔木层生物量随林分结构的变化
Figure 2. Variation of overstory tree biomass of Quercus wutaishanica forests with stand structure
-
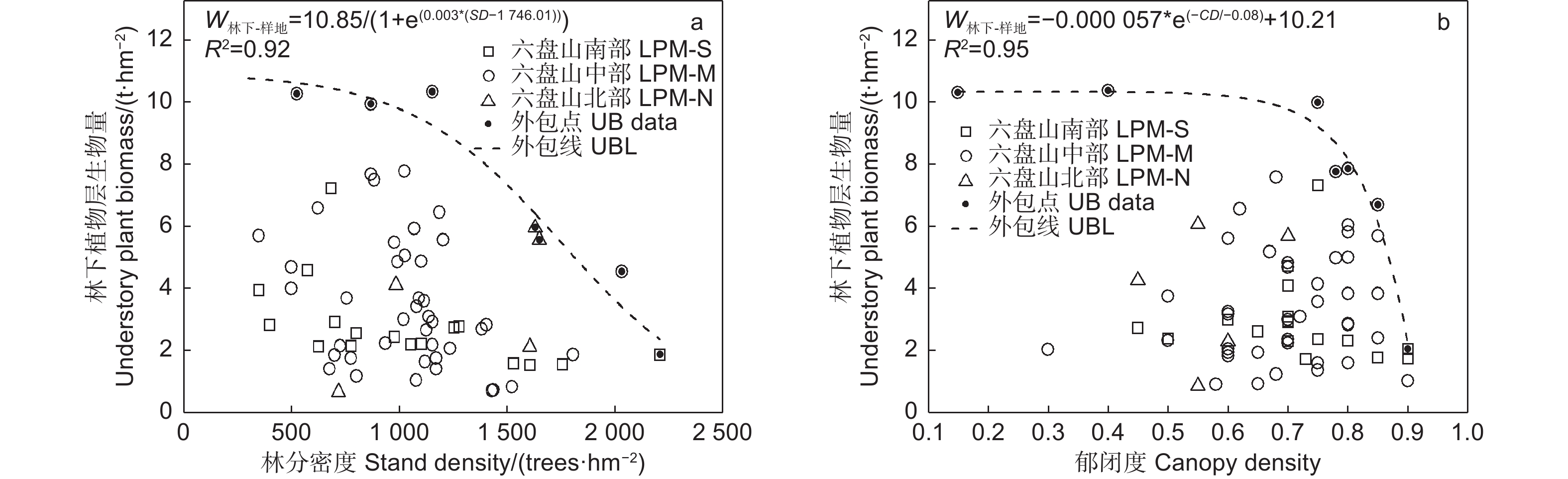
由图3中的外包线可知,林下植物层生物量随林分密度和郁闭度增加均呈先平稳后加速下降的变化,但响应密度的降低起点更早、降低速度更缓。林下植物层生物量在密度达到1 150株∙hm−2和郁闭度达到0.75(分别对应上外包线的林下植物层生物量9.17、9.08 t∙hm−2)之前保持平稳,之后快速下降并最终趋于零。

图 3 辽东栎林下植物层生物量随林分结构的变化
Figure 3. Variation of understory plant biomass of Quercus wutaishanica forests with stand structure
-
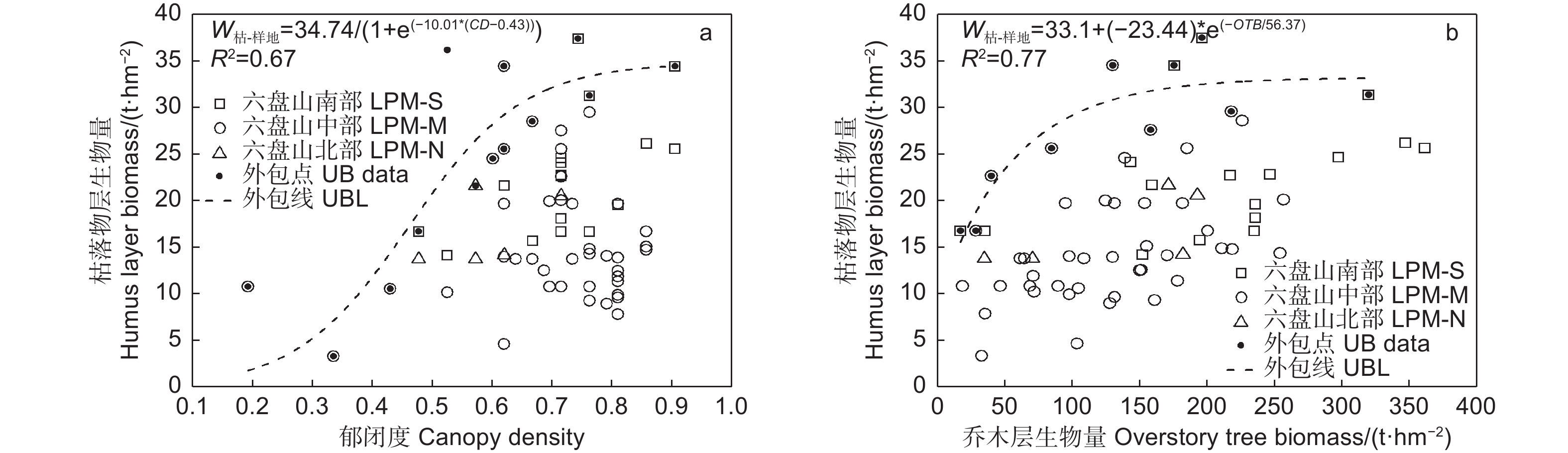
图4中的上外包线表明,枯落物层生物量随郁闭度增加呈“S”型增长,在郁闭度为0.3(对应上外包线枯落物层生物量7.23 t∙hm−2)以前增长缓慢,在0.3~0.7之间快速增长,之后增速变缓并趋平稳,在郁闭度0.75和0.90时分别对应33.34和34.41 t∙hm−2(图4a);枯落物层生物量随乔木层生物量增加表现为先快速增加,在乔木层生物量为130 t∙hm−2后增加变缓并趋平稳(图4b)。
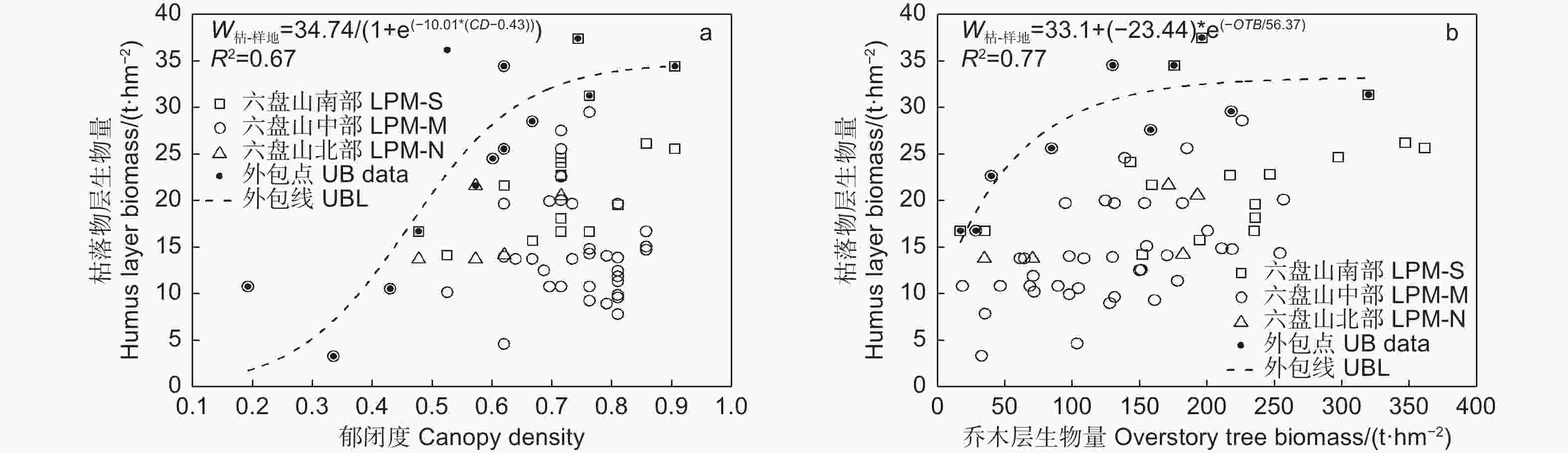
图 4 辽东栎林下枯落物层生物量随林分结构的变化
Figure 4. Variation of little layer biomass of Quercus wutaishanica forests with stand structure
-
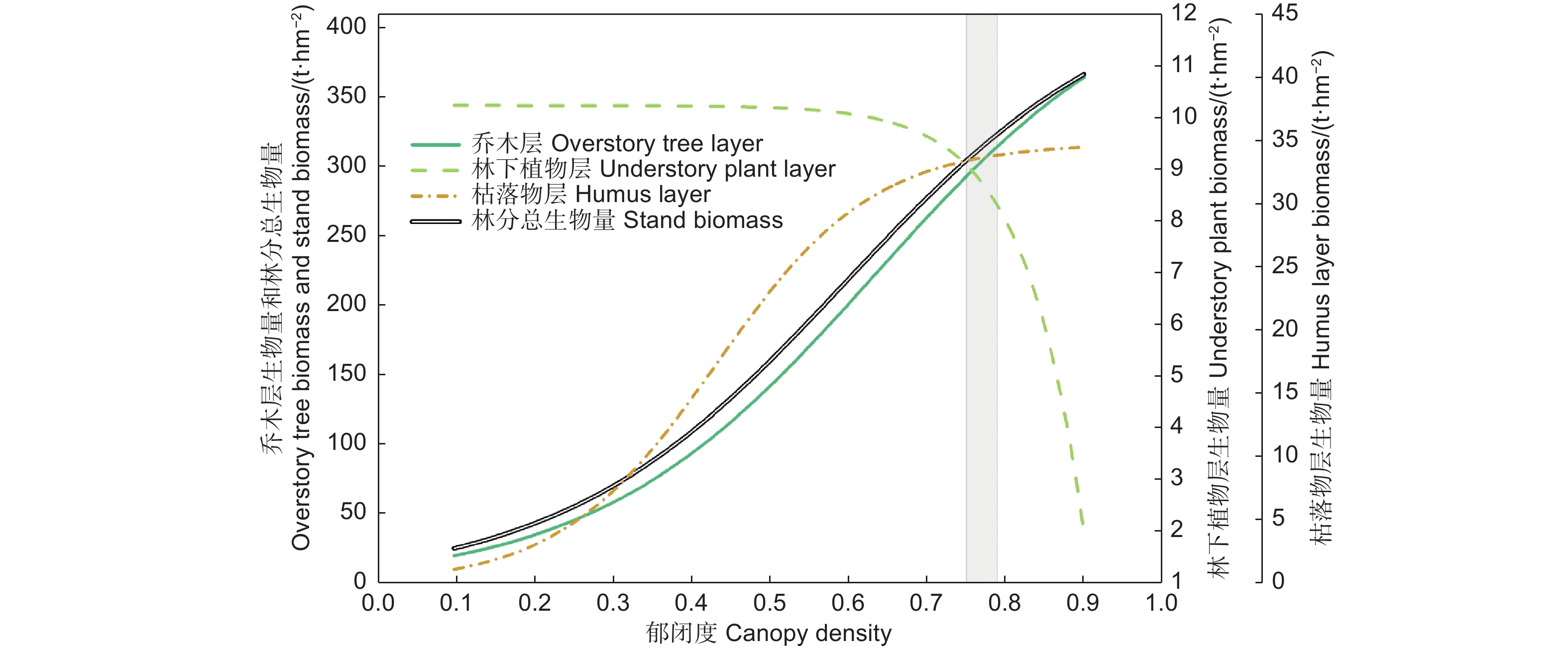
因为林冠郁闭度比其他林分结构对光照的影响更直接,而且生产应用上也容易估计和获取。因此,在图5中,汇总和对比了林分各层生物量随郁闭度的变化,以理解它们的权衡或协同关系。
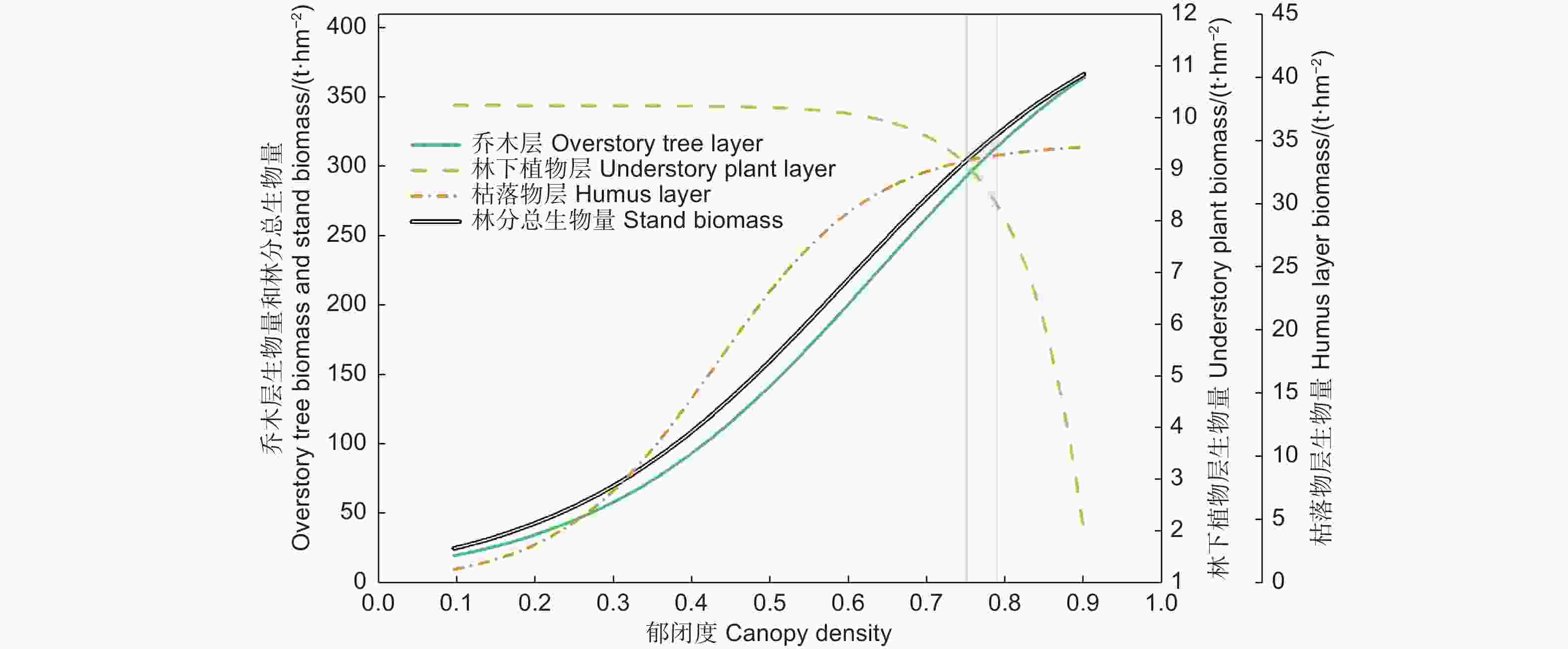
图 5 辽东栎林乔木层、林下植物层、枯落物层及总生物量随郁闭度的变化(基于各层生物量响应郁闭度的外包线)
Figure 5. Variation of the biomass components of overstory tree layer, understory plant layer, litter layer and stand of Quercus wutaishanica forests with rising canopy density (based on the upper boundary line response of each component to canopy density)
辽东栎林各层生物量及其相互关系存在最佳郁闭度范围。在郁闭度0.75以前,林下植物层生物量基本稳定不变,受乔木层和枯落物层生物量的增加影响很小。在郁闭度高于0.75以后,林下植物层生物量的减少与乔木层、枯落物层生物量的继续增加呈越来越明显的竞争关系。若将郁闭度维持在0.75~0.79的范围,可使辽东栎林各层生物量均维持在较高水平。此时乔木层、林下植物层、枯落物层生物量在上外包线上分别为292.3~314.1、8.29~9.08、33.34~33.78 t∙hm−2,都在其上外包线的最大值的80%以上,分别为80%~86%、81%~89%、97%~98%,这时林分总生物量的上外包线数值为334.72~356.17 t∙hm−2,为最大预测值的84%~89%。若郁闭度进一步增加,虽然会使乔木层和枯落物层生物量继续缓慢增加,但林下植物层生物量迅速下降,郁闭度0.9时降到1.98 t∙hm−2,仅为上外包线最大值的20%。因此,可确定郁闭度最优范围是0.75~0.79,这能使辽东栎林各层生物量均维持在较高水平。
-
本研究中辽东栎林的乔木层生物量随林分密度和郁闭度增加而升高,但达到一定阈值后升高趋缓或趋于平稳,这是因为在低于密度和郁闭度阈值时树木个体遭受的光照、水分、养分等竞争较小,树木密度增加带来了乔木层生物量增加;但超过阈值后对单株树木生长的竞争影响越来越大,使降低树木个体生长和生物量的作用越来越接近增加林木密度提高乔木层生物量的作用,从而乔木层生物量逐渐趋于稳定,这与李玲燕[17]的研究结果一致。
以往相关研究都是利用林冠郁闭度预测植物尤其林下植物生物量[18-19];郁闭度通常与树木大小和密度共同导致的冠层生物量密切相关,有必要降低过密林分的密度,以增加林下光照促进林下植物生长[20]。本研究中,林分密度和郁闭度低于某阈值时对林下植物的限制作用相对降低,这是因为林下灌草可通过植物种类变化(增加耐荫性)来适应林下环境(尤其光照)变化[21],从而使林下植物生物量维持相对稳定;当超过阈值后,过高的林分密度和郁闭度使林下光照降低,使耐荫植物生长也受限制,从而使林下植物生物量加速降低[22],甚至限制林下植物生长发育以及灌草的丰富度[23]。
本研究中,辽东栎林枯落物层生物量随乔木层生物量和郁闭度增加而增大。这是因枯落物输入大部分源于乔木层枯枝落叶,密度增大导致的乔木层生物量增加必然导致枯落物输入量增加;只是枯落物层生物量随乔木层生物量增大的升高速度逐渐变缓,这是因一小部分枯落物输入来源于死亡的林下植物,而且枯落物层生物量还受枯落物分解速率影响,当乔木层生物量很大导致郁闭度过高时,源于林下植物的枯落物输入量会变小[24],而且高度郁闭的林下阴湿环境利于枯落物分解[25],从而使枯落物层生物量增速变缓,这与郭耆等[26]的研究结果一致。
-
郁闭度是个重要的林分和林冠结构指标,它与林龄和林木密度的相关性较弱,且年内变化可忽略,容易估测得到,所以有重要的生态学和林学意义[27]。郁闭度影响着林内的光照、水分、温度等条件,对林下植物生长发育、林下树木更新、枯落物层生物量动态等都有重要影响[28],是指导森林结构调控和多功能管理的实用林分结构指标。
一般来说,具有乔灌草复层结构的森林的稳定性和多功能性更高,因此需确定最佳郁闭度,以便同时维持乔木层、林下植物层和枯落物层的较高生物量[29-30]。本研究表明,六盘山区辽东栎林能维持各层生物量均衡的最优郁闭度范围是0.75~0.79,这时各层生物量均处于较高水平。这个郁闭度范围稍高于对陕西黄龙山林区建议的适宜郁闭度(0.7)[31],原因可能是黄龙山的年降水量(611.8 mm)以及海拔较低(962.6~1 783.5 m)和气温(8.6 ℃)较高导致蒸散较强使得树木可利用水分低于六盘山区。类似研究结果在一些其它树种森林的研究中也得到证实[32],如对同属六盘山区的华北落叶松人工林,若维持最佳郁闭度0.7(对应中龄林的林分密度1 100~1 500 株∙hm−2),可同时保持较高的乔木层、林下植物层、枯落物层和土壤层碳密度[33-34]。
-
在本研究中,权衡确定最佳郁闭度时,把乔木层、林下植物层、枯落物层的生物量视为同等重要。但是,如需给予各层次生物量不同权重时,就需确定各层生物量要求的最佳郁闭度的加权平均值,这时的森林多功能管理会更适合一些特殊要求。本研究的另一个不足是,有关研究基于观测数据的外包线,没有考虑林龄的影响,也没给出依据最佳郁闭度计算不同林龄的对应密度的经验公式,需在未来继续积累数据后深入开展这些研究,从而能计算确定不同林龄时的多功能管理最佳密度。再则,本研究在确定最佳林分结构时,并未直接与森林的多种服务功能(水文调节、木材生产、生物多样性保护、固碳等)及林分稳定性和自然更新的要求联系起来,未来需在积累更多数据后首先建立包括郁闭度和密度等在内的林分结构指标与各种服务功能的数量关系,然后计算确定各自要求的林分结构指标并综合权衡后确定能满足多功能管理要求的辽东栎林分结构指标,实现更精细的辽东栎林多功能管理。
-
研究了宁夏六盘山区辽东栎次生林不同植物层生物量受林分结构特征的影响,得到如下结论:(1)随林分密度和郁闭度增大,乔木层生物量先快速上升后缓慢上升,林下植物层生物量先保持平缓后加速下降,枯落物层生物量则表现为增速逐渐变缓。(2)基于辽东栎林各植物层生物量对郁闭度的响应,认为能使各植物层生物量均处于较高水平的最优郁闭度范围为0.75~0.79。
六盘山区辽东栎林分结构对各植物层生物量的影响
The Effects of Stand Structure of Quercus wutaishanica Forests on the Biomass of Different Plant Layers in the Liupan Mountains of Northwest China
-
摘要:
目的 量化宁夏六盘山区辽东栎次生林各植物层(乔木层、林下植物层(草本和灌丛)、枯落物层)生物量随郁闭度及林分密度的变化,确定能使各层生物量均维持在较高水平的最佳林分结构,以充分发挥森林多种功能。 方法 基于野外调查样地数据,利用上外包线分析法确定辽东栎林各层生物量对林分密度和郁闭度的响应规律,并经综合权衡后确定最佳郁闭度范围。 结果 随郁闭度和林分密度增加,乔木层生物量先快速增加,在郁闭度0.75和密度1 050株∙hm−2时分别达到292.3和296.9 t∙hm−2,分别为其外包线最大值的80%和82%,之后增速变缓;林下植物层生物量先缓慢下降,在郁闭度0.75和密度1 150株∙hm−2时分别达到9.08和9.17 t∙hm−2,分别为其外包线最大值的89%和86%,之后快速减小;枯落物层生物量先逐渐增加,在郁闭度0.75时达到33.34 t∙hm−2,为外包线最大值的97%。综合分析各层生物量随郁闭度的变化表明,需维持林冠郁闭度在0.75~0.79左右,才能使林分各层生物量都保持在各自上外包线最大值的80%以上。 结论 在宁夏六盘山区,要使辽东栎林的乔木层、林下植物层、枯落物层的生物量均维持在较高水平,需维持林冠郁闭度在最佳范围0.75~0.79左右,这可用于指导该地区及类似地区的辽东栎林多功能经营。 Abstract:Objective To quantify the effects of stand density and canopy density on the biomass of different plant layers (overstory tree layer, understory plant layer, and litter layer) of Quercus wutaishanica secondary forests in the Liupan Mountains of Ningxia in northwest China, and to determine the optimal stand structure that can maintain the biomass of each layer at a relatively high level for fully using the multiple forest ecosystem services. Method Using sample plot data obtained from field survey, the response of biomass in each layer of Quercus wutaishanica forests to stand density and canopy density were determined by upper boundary lines (UBL) analysis. Then these UBLs were superimposed to find an optimal canopy density range. Result With the increase of canopy density and stand density, the overstory tree layer biomass increased rapidly at first, reaching 292.3 and 296.9 t·hm−2 when the canopy density was 0.75 and the density was 1 050 trees∙hm−2, respectively (80% and 82% of the UBL predicted maximum), and then the growth rate slowed down; the understory plant biomass decreased slowly first, and reached 9.08 and 9.17 t∙hm−2 when the canopy density was 0.75 and the density was 1 150 plants∙hm−2, which were 89% and 86% of the maximum value of UBL, respectively. Then it decreased rapidly; the litter layer biomass increased rapidly at first, and reached 33.34 t∙hm−2 when the canopy density was 0.75, which was 97% of the maximum value of UBL. Therefore, the canopy density should be maintained at about 0.75-0.79 in order to keep the biomass of each layer of the stand above 80% of the maximum value of UBL. Conclusion In order to maintain the biomass at relatively high level for all different plant layers of Quercus wutaishanica forests in Liupan Mountains, the canopy density should be maintained at about 0.75-0.79, which will be helpful for guiding the multifunctional management of Quercus wutaishanica forests in the study region and similar areas. -
图 1 宁夏六盘山区辽东栎林调查样地位置布设
Figure 1. Sample plots layout of Quercus wutaishanica forest stands in Liupan Mountains Area of Ningxia
图 2 辽东栎林乔木层生物量随林分结构的变化
Figure 2. Variation of overstory tree biomass of Quercus wutaishanica forests with stand structure
图 3 辽东栎林下植物层生物量随林分结构的变化
Figure 3. Variation of understory plant biomass of Quercus wutaishanica forests with stand structure
图 4 辽东栎林下枯落物层生物量随林分结构的变化
Figure 4. Variation of little layer biomass of Quercus wutaishanica forests with stand structure
图 5 辽东栎林乔木层、林下植物层、枯落物层及总生物量随郁闭度的变化(基于各层生物量响应郁闭度的外包线)
Figure 5. Variation of the biomass components of overstory tree layer, understory plant layer, litter layer and stand of Quercus wutaishanica forests with rising canopy density (based on the upper boundary line response of each component to canopy density)
表 1 辽东栎林乔木单株生物量计算模型
Table 1. The relations of single tree biomass of Quercus wutaishanica secondary forest
树种
Tree species拟合方程
Fitting equations方程来源
Equation source辽东栎 Quercus wutaishanica B=0.732 0(D) 1.968 1 文献[12] 白桦 Betula platyphylla B=0.668 4(D) 1.965 6 少脉椴 Tilia paucicostata B=0.291 6(D) 2.146 8 山杨Populus tremula B=0.262 4(D) 2.215 0 文献[13] 注:B为乔木单株整体生物量(kg·株−1),D为乔木单株胸径(cm)。
Note: B is the overall biomass of a single tree (kg·tree−1), and D is the diameter at breath height (cm) of single tree. 下载: 导出CSV
下载: 导出CSV
表 2 辽东栎林下主要灌木种类的单株生物量计算模型
Table 2. The relations of single plant biomass of main shrub species in Quercus wutaishanica
灌木种类
Shrub species拟合方程
Fitted equations决定系数
R2方程来源
Equation source绣球绣线菊(Spiraea blumei) B=0.498(D)1.653 2 0.925 8 依据本研究的灌木生物量调查数据建立 山楂(Crataegus pinnatifida) B=0.104 6D2−0.045 1D + 0.030 9 0.981 5 北京忍冬(Lonicera elisae) B=0.075 6(D)1.257 0.832 0 水荀子(Cotoneaster multiflora) B=0.062 2(D)1.693 9 0.982 8 榛(Corylus heterophylla) B=0.045 2(D)1.743 0.907 7 桦叶荚蒾(Viburnum betulifolium) B=−0.000 6D2 + 0.176 3D−0.116 1 0.908 7 其它灌木Other shrub species B地上=1.890 7ln D−2.496 2 0.862 1 文献[14] B根=0.896 6 lnD−2.263 3 0.568 5 注:B为灌木单株生物量(kg·株−1);D为灌木地径(cm)。
Note: B is the overall biomass of a single shrub (kg·plant−1), and D is the ground diameter (cm) of single shrub plant.
下载: 导出CSV
表 3 六盘山各分区内辽东栎林样地的平均立地和结构特征
Table 3. Site and structure characteristics of the Quercus wutaishanica stand plots in different sub-regions
项目
Item六盘山南部
South of Liupan
Mountains六盘山中部
Middle of Liupan
Mountains六盘山北部
North of Liupan
Mountains海拔 Altitude/m 1 960~2 280 1 789~2 420 2 030~2 260 土层厚度 Soil thickness/cm 78 ± 24 64 ± 20 58 ± 12 坡度 Slope gradient/(°) 34 ± 7 30 ± 8 37 ± 2 林分密度 Tree density/(株∙hm−2) 1 037 ± 494 1 055 ± 337 1 314 ± 388 郁闭度 Canopy density 0.71 ± 0.12 0.68 ± 0.15 0.57 ± 0.08 林龄 Stand age/a 38 ± 17 45 ± 10 38 ± 4 平均树高 Average tree height/m 12.3 ± 3.8 8.2 ± 2.7 6.6 ± 0.8 平均胸径 Average DBH/cm 17.3 ± 4.9 13.3 ± 3.8 11.4 ± 1.7
下载: 导出CSV
表 4 六盘山各分区内辽东栎林样地的各植物层生物量
Table 4. The biomass of different layers of Quercus wutaishanica forests in different sub-regions t∙hm−2
植物层
Plant layers六盘山南部
South of Liupan
Mountains六盘山中部
Middle of Liupan
Mountains六盘山北部
North of Liupan
Mountains平均值
Average乔木层 Overstory tree layer 209.28 ± 92.21 124.50 ± 61.39 94.59 ± 64.73 146.39 ± 79.67 林下植物层 Understory plant layer 2.75 ± 1.35 4.15 ± 3.00 3.65 ± 2.00 3.76 ± 2.67 灌木层 Shrub layer 2.44 ± 1.36 3.11 ± 2.32 3.28 ± 1.95 2.98 ± 2.11 草本层 Herb layer 0.31 ± 0.31 1.04 ± 2.22 0.37 ± 0.32 0.78 ± 1.84 枯落物层 Humus layer 22.86 ± 6.45 16.70 ± 7.74 15.70 ± 3.52 18.27 ± 7.67 总生物量 Total biomass 234.89 ± 94.16 145.35 ± 64.05 113.94 ± 68.06 168.42 ± 82.79
下载: 导出CSV
-
[1] 中国林业科学研究院“多功能林业”编写组. 中国多功能林业发展道路探索[M]. 北京: 中国林业出版社, 2010. [2] 樊宝敏, 吴水荣, 王彦辉. 多功能林业发展模式[M]. 北京: 科学出版社, 2013. [3] Chen S, Shahi C, Chen H, et al. Trade-offs and synergies between economic gains and plant diversity across a range of management alternatives in boreal forests[J]. Ecological Economics, 2018, 151: 162-172. doi: 10.1016/j.ecolecon.2018.05.014 [4] Carpentier S, Filotas E, Handa I T, et al. Trade-offs between timber production, carbon stocking and habitat quality when managing woodlots for multiple ecosystem services[J]. Environmental Conservation, 2016, 44(1): 1-10. [5] 李英武, 张炳乾, 赵树俭. 宁夏六盘山辽东栎天然更新调查分析[J]. 陕西林业科技, 2003(3):13-15. doi: 10.3969/j.issn.1001-2117.2003.03.004 [6] 徐秀梅. 宁夏六盘山辽东栎林群落特征分析[J]. 宁夏农林科技, 1997(2):17-20. [7] 王占印, 于澎涛, 王双贵, 等. 宁夏六盘山区辽东栎林的空间分布及林分特征[J]. 林业科学研究, 2011, 24(1):97-102. [8] 邓 磊, 关晋宏, 高万里, 等. 黄土区辽东栎群落细根生物量对物种多样性及气候的响应[J]. 西北林学院学报, 2018, 33(5):16-22. doi: 10.3969/j.issn.1001-7461.2018.05.03 [9] 张希彪, 上官周平. 黄土丘陵区主要林分生物量及营养元素生物循环特征[J]. 生态学报, 2005(3):527-537. doi: 10.3321/j.issn:1000-0933.2005.03.021 [10] Ahmad B, Wang Y H, Hao J, et al. Optimizing stand structure for trade-offs between overstory timber production and understory plant diversity: A case-study of a larch plantation in northwest China[J]. Land Degradation & Development, 2018, 29(9): 2998-3008. [11] 宁夏林业厅自然保护区办公室. 六盘山自然保护区科学考察[M]. 银川: 宁夏人民出版社, 1989. [12] 程积民, 金晶炜, 田 瑛, 等. 宁夏森林植被及土壤碳密度分布特征[J]. 农业工程学报, 2016, 32(13):109-117. doi: 10.11975/j.issn.1002-6819.2016.13.016 [13] 姜 鹏, 任佳佳, 任 仙, 等. 典型针阔混交林中山杨生物量和碳储量研究[J]. 西北林学院学报, 2014, 29(1):15-20 + 83. doi: 10.3969/j.issn.1001-7461.2014.01.03 [14] 刘延惠, 王彦辉, 于澎涛, 等. 六盘山主要植被类型的生物量及其分配[J]. 林业科学研究, 2011, 24(4):443-452. [15] Matsushita M, Takata K, Hitsuma G, et al. A novel growth model evaluating age-size effect on long-term trends in tree growth[J]. Functional Ecology, 2015, 29(10): 1250-1259. doi: 10.1111/1365-2435.12416 [16] Schmidt U, Thni H, Kaupenjohann M. Using a boundary line approach to analyze N2O flux data from agricultural soils[J]. Nutrient Cycling in Agroecosystems, 2000, 57(2): 119-129. doi: 10.1023/A:1009854220769 [17] 李玲燕, 代林利, 刘 丽, 等. 不同密度12年生杉木林地上部分生物量的垂直分布[J]. 亚热带农业研究, 2020, 16(4):229-236. [18] Peek J M, Korol J J, Gay D, et al. Overstory–understory biomass changes over a 35-year period in southcentral Oregon[J]. Forest Ecology & Management, 2001, 150(3): 267-277. [19] 李明泽, 毛学刚, 范文义. 基于郁闭度联立方程组模型的森林生物量遥感估测[J]. 林业科学, 2014, 50(2):85-91. [20] Liu G H, Hui C, Chen M, et al. Variation in individual biomass decreases faster than mean biomass with increasing density of bamboo stands[J]. Journal of Forestry Research, 2020, 31(3): 981-987. doi: 10.1007/s11676-018-0796-1 [21] 黄团冲, 贺康宁, 王先棒, 等. 北川河流域森林冠层结构对林下植被多样性的影响[J]. 中国水土保持科学, 2018, 16(4):106-114. [22] 温 晶, 张秋良, 李嘉悦, 等. 间伐强度对兴安落叶松林林下植被多样性及生物量的影响[J]. 中南林业科技大学学报, 2019, 39(5):95-100 + 118. [23] Wang Y H, Xiong W, Gampe S, et al. A Water yield-oriented practical approach for multifunctional forest management and its application in dryland regions of China[J]. Jawra Journal of the American Water Resources Association, 2015, 51(3): 689-703. doi: 10.1111/1752-1688.12314 [24] Helman D, Leu S, Mussery A. Contrasting effects of two Acacia species on understorey growth in a drylands environment: Interplay of canopy shading and litter interference[J]. Journal of Vegetation Science, 2017, 28(6): 1140-1150. doi: 10.1111/jvs.12576 [25] Mlambo D, Mwenje E. Influence of Colophospermum mopane canopy cover on litter decomposition and nutrient dynamics in a semi-arid African savannah[J]. African Journal of Ecology, 2010, 48(4): 1021-1029. doi: 10.1111/j.1365-2028.2010.01208.x [26] 郭 耆, 赵厚本, 周光益, 等. 南亚热带4个树种人工林生物量及其分配格局[J]. 林业科学研究, 2022, 35(1):182-189. [27] 李永宁, 张宾兰, 秦淑英, 等. 郁闭度及其测定方法研究与应用[J]. 世界林业研究, 2008, 21(1):40-46. [28] 韩海荣, 姜玉龙. 栓皮栎人工林光环境特征的研究[J]. 北京林业大学学报, 2000, 22(4):92-96. doi: 10.3321/j.issn:1000-1522.2000.04.016 [29] 季 蕾, 亢新刚, 郭韦韦, 等. 金沟岭林场3种林型不同郁闭度林下灌草生物量[J]. 东北林业大学学报, 2016, 44(9):29-33 + 39. doi: 10.3969/j.issn.1000-5382.2016.09.007 [30] Nadkarni N M, Schaefer D, Matelson T J, et al. Biomass and nutrient pools of canopy and terrestrial components in a primary and a secondary montane cloud forest, Costa Rica[J]. Forest Ecology and Management, 2004, 198(1-3): 223-236. doi: 10.1016/j.foreco.2004.04.011 [31] 于世川, 张建国, 叶权平, 等. 抚育间伐对黄龙山辽东栎林分质量的影响[J]. 西北林学院学报, 2018, 33(3):52-60. doi: 10.3969/j.issn.1001-7461.2018.03.09 [32] Wang Q W, Robson T M, Pieriste M, et al. Canopy structure and phenology modulate the impacts of solar radiation on C and N dynamics during litter decomposition in a temperate forest [J]. The Science of the Total Environment, 2022, 820. [33] Ahmad B, Wang Y H, Hao J, et al. Variation of carbon density components with overstory structure of larch plantations in northwest China and its implication for optimal forest management [J]. Forest Ecology and Management, 2021, 496. [34] 黄小男. 六盘山华北落叶松人工林生物量对立地条件和林分结构的响应与模拟[D]. 北京: 北京林业大学, 2020. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5147
- HTML全文浏览量: 1134
- PDF下载量: 84
- 被引次数: 0

























