



-
马尾松( Pinus massoniana Lamb.)速生、耐干旱瘠薄、适应性强,是我国南方荒山造林的先锋树种,在一些立地条件较差的山地,马尾松甚至是唯一可造林的本土树种。由松材线虫( Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle,PWN)引起的松树萎蔫病是一种毁灭性森林病害,其中,马尾松为松材线虫主要危害对象之一。马尾松群体中存在抗病基因型,利用抗病品种防治松材线虫病为最经济、有效的途径之一,现已在安徽、浙江等地选育出一批抗性较高的马尾松家系和无性系[1],但对其抗性机理并不完全清楚。
植物抗性基因(R基因)是其对病虫害防御反应的主要基因, 其中, 富含亮氨酸重复序列(LRRs)是植物R基因编码蛋白最重要的结构特征之一, NBS-LRR 、 LRR-STK 类是R基因中的两类[2]。在植物与线虫互作过程中, NBS-LRR 类是植物抗病基因中数量最多的一类[3]。依据 NBS 在N端的不同结构,又主要分为 TIR-NBS-LRR 和 CC-NBS-LRR 两种类型。 Hs1pro-1是从甜菜( Beta vulgaris L.)中获得的第1个R基因[4]。之后,在马铃薯( Solanum tuberosum L.)和樱桃( Cerasus pseudocerasus Lindl.)中发现的R基因编码蛋白皆具有核苷酸结合位点和富亮氨酸重复序列(NBS-LRR)结构,其中, Gpa2、Mi-1.2、Mi9和Hero A编码蛋白属于N端为卷曲螺旋结构域(CC-NBS-LRR)结构类别的抗性蛋白,而Ma和Gro1-4编码蛋白属于N端为Toll/白细胞介素受体(TIR-NBS-LRR)结构类别的抗性蛋白。 CC-NBS-LRR 与 TIR-NBS-LRR 这两类抗性基因在应对病原菌侵染时发挥着不同的功能[5]。基因不受时期、部位以及环境影响均可较稳定表达即为组成型基因。在这些植物抗性基因中,基因组成型表达对抗性起重要作用。如 CC-NBS-LRR 亚家族中组成型表达基因RGC260在向日葵( Helianthus annuus L.)抗锈病过程中发挥重要作用[6]。转录因子 CBF1 的组成型表达可增强冷敏感植物的抗寒性[7]。感染柑桔黄龙病菌( Candidatus Liberibacter spp.)后,组成型表达基因 PtCDR2 和 PtCDR8 在抗黄龙病柑桔( Poncirus trifoliate L.)中表达丰度较高[8]。
目前, 在松树对松材线虫病的抗性机制研究上进展相对较慢,仅发现一些抗性相关物质或基因。如Menéndez-Gutiérrez等对不同抗性的几种松树进行研究,发现组成型氮、多酚和磷在对松材线虫病抗性较强的火炬松( P. taeda L.)和地中海松( P. halepensis L.)木质部中的含量高于易感病的赤松( P. sylvestris L.)[9],松类内酚类、单宁、黄酮和木质素含量高不利于松材线虫的繁殖生长[10]。在分子水平上发现与活性氧、木质素、萜类和乙烯等合成相关的基因与抗松材线虫病关系密切[11-14]。本研究组已利用转录组高通量测序技术发现编码扩展蛋白、萜类合酶和活性氧等基因在接种松材线虫前后表达量发生显著的变化[15],且在高抗与易感马尾松间表达量差异也达到显著水平,但这种表达量的差异为诱导型,在不接种情况下,无法利用这些基因对高抗马尾松进行早期鉴定。本试验在前期研究基础上,着重于研究和识别高抗和易感马尾松间与抗松材线虫病相关的组成型表达基因,为解析马尾松抗性机理和高抗马尾松早期选择提供理论依据。
-
试验材料来自安徽省林业科学研究院4年生马尾松无性系试验林。根据前期该院对试验林内无性系测定的抗性指数,于2017年7月中旬,选择高抗无性系‘休宁5’和易感无性系‘黄山1’各18株。供试松材线虫为高致病性株系‘广株3B’和‘全4’混合株系,于灰葡萄孢菌( Botrytis cinerea )菌落上纯培养,保存于4℃冰箱中备用。
-
将灰葡萄孢菌接种于PDA平板上,于28℃培养箱中培养5 d。待菌落长成后将松材线虫接种于该菌落中,28℃培养直至菌落完全被松材线虫取食,进一步用贝尔曼漏斗法分离松材线虫,12 h后收集于10 mL离心管中,3 000 r·min-1离心5 min, 去除上清液。将各离心管中的松材线虫汇集并混匀,最终制备成每微升含有50条松材线虫的悬浊液。
-
在高抗无性系与易感无性系中,分别选取9株接种松材线虫悬浊液,9株接种蒸馏水作为对照。参考Liu等处理方法[16]进行接种与样品采集。所取样品包括高抗马尾松和易感马尾松2个基因型,接种松材线虫和接种蒸馏水2个处理,接种后第1、15和30天3个取样时间点,3个生物学重复,共计36个样品。
-
采用植物RNA试剂盒RN38 EASYSpin plus(Aidlab Biotech,Beijing,China)分别提取接种后1、15、30 d各样本RNA。利用Ultraspec TM 2100 Pro UV检测RNA的浓度,1%琼脂糖凝胶电泳检测RNA纯度以及完整性。选择RNA浓度≥400 ng·μL-1、 OD260 / 280为1.8~2.2、 OD260 / 230>2.0、RIN≥8.0作为高质量RNA。
-
马尾松无参考基因组,为获得全面的转录组数据,首先将36份高质量RNA等质量混合,利用Illumina HiSeqTM 2000测序平台进行转录组测序。同时对36个样品进行表达谱测序,并使用Bowtie方法将36份样品中高质量读长(reads)比对到转录组上,获取各样本中所有基因的表达量。
-
转录组和表达谱数据已提交至NCBI,登录号为SRP103562。利用Benjamini-Hochberg方法进行 P 值校正,获得差异基因。采用GOseq R对差异基因进行GO富集分析, P 值≤0.05的GO term为显著富集。
-
使用Primer3.0软件对候选基因设计引物,采用ABI7300实时荧光定量PCR系统(Applied Biosystems,CA,USA)对候选基因表达情况进行qRT-PCR检测,以验证转录组测序结果。反应总体系为20 μL,包括0.4 μL ROX染料,2 μL cDNA,0.4 μL引物和10 μL SYBR Premix ExTaqTM。PCR程序为95℃预变性10 s,95℃变性5 s,58℃退火30 s,72℃延伸30 s,共40个循环。3次生物学重复,3次技术重复。利用 EF1-α 作为参考基因对候选基因表达量进行标准化,并根据2-ΔΔCT方法计算候选基因的相对表达水平。
-
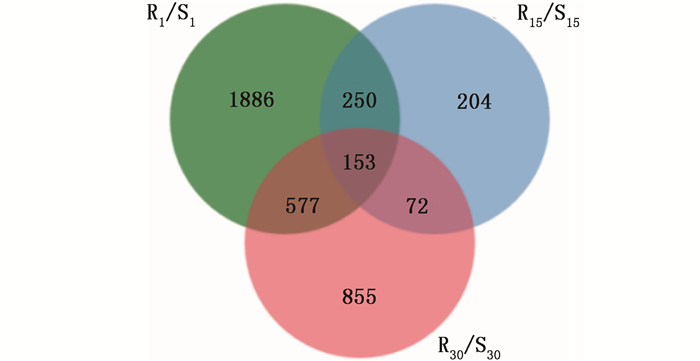
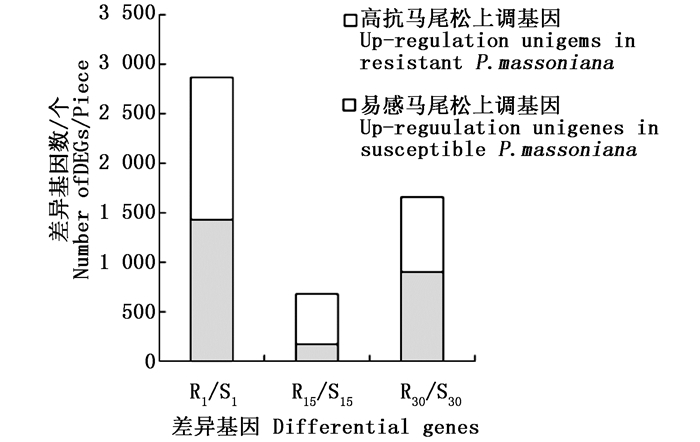
根据前期预实验结果发现,接种松材线虫后第15天马尾松顶稍略显萎蔫,接种后第30天针叶开始变黄。故本研究选取不接种松材线虫、接种松材线虫后第1、15和30天时的材料进行转录组测序。以马尾松36个样本茎段等量混合RNA为材料进行二代转录组测序,共获得80 340个unigenes,平均长度为675 bp,N50值分别为1 634 bp和1 245 bp。其中,14 994 unigenes(18.66%)大于1 kb。将高抗与易感马尾松的表达谱进行比较(将易感马尾松作为对照),在接种松材线虫后第1、15和30天分别获得的差异基因为2 866、679和1 657个,其中, 在高抗马尾松上具有较高表达量的基因数分别为1 438、508和757个(图 1)。进一步分析结果显示,这些差异基因在不同接种时间的共有基因相对较少(图 2)。如在接种松材线虫后第1天获得的差异基因中有1 886个差异基因是特有的,仅有153个差异基因出现在接种后第15天和第30天。
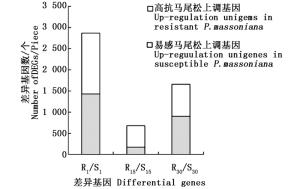
图 1 高抗与易感马尾松在不同接种时间点的差异基因数
Figure 1. The differential genes at different time of inoculation in resistant and susceptible P. massoniana
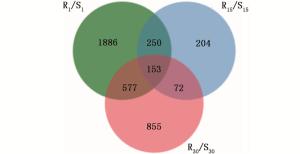
图 2 马尾松在不同接种时间点的共有差异基因
Figure 2. Common differential genes of P. massoniana at different inoculation time points
-
对高抗与易感马尾松接种松材线虫后3个时间点的差异基因进行GO富集分析,其中, 将在生物过程(Biological process,BP)、细胞组成(Cellular component,CC)和分子功能(Molecular function,MF)中最显著的前10位GO项列于表 1~3。结果显示:在BP类别中(表 1),接种后1、15和30 d最显著富集的GO项皆为氧化-还原过程,其次为跨膜转运。另外GO项刺激反应、转录调节以及香叶基二磷酸代谢过程也较显著。在CC类别中,接种后1、15和30 d显著富集的前10位GO项,尽管排序略有不同,但类目相同(表 2),其中, 最显著的GO项为膜整体构成(GO:0016021),其次为质膜(GO:0005886)。在MF类别中,接种后1、15和30 d最显著富集的前3位GO项相同,分别为氧化还原酶活性(GO:0016491)、蛋白结合(GO:0005515)和DNA特异序列结合(GO:0000981)。
表 1 高抗与易感马尾松接种松材线虫后在不同时间点差异基因的GO富集分析(BP)
Table 1. GO enrichment analysis of different genes at different time of inoculation in resistant and susceptible P. massoniana (BP)
基因本体编号
GO ID类别 Term 差异基因 GO 富集 Different genes GO enrichment 接种后1 d
1d After inoculation接种后15 d
15 d After inoculation接种后30 d
30 d After inoculationGO:0055114 氧化-还原过程 Oxidation-reduction process 2.80E-11 4.80E-11 3.50E-10 GO:0055085 跨膜转运 Transmembrane transport 2.30E-05 4.80E-05 1.10E-05 GO:0008152 代谢过程 Metabolic process 8.90E-05 - - GO:0042026 蛋白质重折叠 Protein refolding 9.70E-05 - - GO:0006886 胞内蛋白转运 Intracellular protein transport 0.000 32 0.000 35 - GO:0009725 激素应答 Response to hormone 0.000 46 0.000 49 - GO:0050896 刺激反应 Response to stimulus 0.000 50 0.000 30 0.000 49 GO:0006950 胁迫应答 Response to stress 0.000 97 0.001 11 0.000 93 GO:0006355 转录调节 Regulation of transcription, DNA-templated 0.001 25 0.001 85 0.001 25 GO:0033383 香叶基二磷酸代谢过程 Geranyl diphosphate metabolic process 0.001 57 0.001 46 0.000 56 GO:0006468 蛋白质磷酸化 Protein phosphorylation - 0.000 18 0.000 17 GO:0006749 谷胱甘肽代谢过程 Glutathione metabolic process - 0.000 42 - GO:0006097 乙醛酸循环 Glyoxylate cycle - - GO:0030435 孢子形成 Sporulation resulting in formation of a cellular spore - - GO:0006886 胞内蛋白转运 Intracellular protein transport - - 0.000 31 GO:0009266 温度刺激应答 Response to temperature stimulus - - 0.002 32 GO:0006464 细胞蛋白质修饰过程 Cellular protein modification process - - 0.002 79 表 2 高抗与易感马尾松接种松材线虫后在不同时间点差异基因的GO富集分析(CC)
Table 2. GO enrichment analysis of different genes at different time of inoculation in resistant and susceptible P. massoniana (CC)
基因本体编号
GO ID类别 Term 差异基因 GO 富集 Different genes GO enrichment 接种后1 d
1d After inoculation接种后15 d
15 d After inoculation接种后30 d
30 d After inoculationGO:0016021 膜整体构成 Integral component of membrane 3.60E-24 1.20E-21 3.50E-17 GO:0005886 质膜 Plasma membrane 5.90E-11 2.90E-11 3.60E-11 GO:0009507 叶绿体 Chloroplast 1.10E-09 3.60E-10 6.00E-10 GO:0005829 细胞质基质 Cytosol 3.30E-07 3.10E-07 2.90E-07 GO:0009570 叶绿体基质 Chloroplast stroma 2.20E-05 1.70E-05 1.80E-05 GO:0044464 细胞组分 Cell part 4.80E-05 0.000 12 9.90E-05 GO:0005773 液泡 Vacuole 6.40E-05 5.40E-05 5.80E-05 GO:0005768 核内体 Endosome 9.60E-05 6.10E-05 7.30E-05 GO:0009941 叶绿体被膜 Chloroplast envelope 0.000 16 3.10E-05 0.000 14 GO:0005840 核糖体 Ribosome 0.000 21 1.50E-06 2.00E-05 表 3 高抗与易感马尾松接种松材线虫后在不同时间点差异基因的GO富集分析(MF)
Table 3. GO enrichment analysis of different genes at different time of inoculation in resistant and susceptible P. massoniana (MF)
基因本体编号
GO ID类别 Term 差异基因 GO 富集 Different genes GO enrichment 接种后1 d
1d After inoculation接种后15 d
15 d After inoculation接种后30 d
30 d After inoculationGO:0016491 氧化还原酶活性 Oxidoreductase activity 2.10E-10 8.80E-09 5.50E-08 GO:0005515 蛋白结合 Protein binding 1.90E-07 5.00E-08 2.00E-07 GO:0000981 DNA特异序列结合 Sequence-specific DNA binding 1.80E-05 4.10E-06 1.40E-05 GO:0003824 催化活性 Catalytic activity 4.60E-05 0.000 14 0.000 19 GO:0005488 结合 Binding 5.00E-05 - - GO:0004721 磷蛋白质磷酸蛋白活性 Phosphoprotein phosphatase activity 0.000 14 - 7.60E-05 GO:0010181 FMN结合 FMN binding 0.000 26 5.60E-05 0.000 17 GO:0008874 葡萄糖酸5-脱氢酶活性 Gluconate 5-dehydrogenase activity 0.000 49 - - GO:0016832 醛裂解酶活性 Aldehyde-lyase activity 0.000 53 - - GO:0004591 酮戊二酸脱氢酶 Oxoglutarate dehydrogenase 0.000 62 0.000 18 0.000 3 GO:0004738 丙酮酸脱氢酶活性 Pyruvate dehydrogenase activity - 0.000 14 0.000 29 GO:0004375 甘胺酸脱氢酶 Glycine dehydrogenase - 0.000 23 0.000 37 GO:0051287 NAD结合 NAD binding - 0.000 39 GO:0016758 转移酶活性 Transferase activity - 0.000 55 0.000 58 -
为更好地了解GO项刺激反应参与马尾松抗松材线虫病的方式,对相关差异基因进行进一步分析。结果显示:在该GO项中包含26个R基因。在接种后第1天,与刺激反应GO项相关的差异基因共71个,其中,R基因25个;接种后第15天,与刺激反应GO项相关的差异基因共17个,其中,R基因4个;接种后第30天,与刺激反应GO项相关的差异基因共47个,其中,R基因14个。在这些R基因中,除1个 NBS-LRR 基因(c76976.graph_c0)和1个编码受体丝氨酸/苏氨酸蛋白激酶基因(c71845.graph_c0)在易感马尾松接种松材线虫后表达量显著发生变化外(以接种蒸馏水作为对照),其它基因不论在高抗还是易感马尾松中(表 4),接种松材线虫后表达量变化皆未达显著水平( P <0.05),意味着这些基因的表达差异为组成型,即不论接种与否对这些基因的表达影响不大。
表 4 R基因在高抗和易感马尾松间的差异表达
Table 4. Differential expression of differential R genes in resistant and susceptible P. massoniana
类别 Term 基因编号 Gene_ID 基因表达量 Relative level of gene expression/RPKM RI1 SI1 RI15 SI15 RI30 SI30 受体丝氨酸/苏氨酸蛋白激酶
LRR receptor-like serine/ threonine-protein kinasec63612.graph_c0 11.14 0.78 10.80 0.67 15.65 0.38 c75632.graph_c0 25.21 75.21 - - 8.99 43.55 c74934.graph_c1 9.49 31.04 - - 11.00 43.81 c78638.graph_c0 1.18 11.57 - - 0.30 9.71 c72330.graph_c0 2.55 10.92 - - 1.34 5.37 c52089.graph_c0 0.61 5.38 - - - - c77321.graph_c0 15.01 34.64 - - - - c77212.graph_c0 5.26 13.39 - - - - c76446.graph_c0 7.21 22.56 - - - - c64844.graph_c0 1.10 5.51 - - - - c71845.graph_c0 5.21 40.05 - - - - c72922.graph_c0 - - - - 17.85 60.44 NBS-LRR蛋白
NBS-LRR proteinc79162.graph_c0 0.00 5.56 0.65 5.82 - - c49318.graph_c0 0.40 5.25 0.90 4.83 0.19 5.22 c66133.graph_c0 10.50 30.33 - - 12.53 30.92 c73896.graph_c0 2.48 8.31 - - 3.32 9.25 c76976.graph_c0 11.13 40.39 - - 9.57 29.29 c76662.graph_c0 2.63 7.31 - - - - c62473.graph_c0 1.72 7.55 - - - - TIR-NBS-LRR蛋白
TIR-NBS-LRR proteinc65785.graph_c0 2.68 0.00 - - 2.71 0.00 c73334.graph_c0 0.60 51.29 4.19 16.98 0.60 22.60 c53610.graph_c0 2.44 7.51 - - 2.20 7.67 c76602.graph_c0 1.95 4.46 - - 1.93 3.21 c65671.graph_c0 0.00 3.34 - - 0.00 4.84 c75804.graph_c0 2.88 8.01 - - - - c66096.graph_c0 4.10 23.63 - - - - 注:RI1,高抗马尾松接种松材线虫后第1天的基因表达;S1,易感马尾松接种松材线虫后第1天的基因表达;RI15,高抗马尾松接种松材线虫后第15天的基因表达;SI15,易感马尾松接种松材线虫后第15天的基因表达;RI30,高抗马尾松接种松材线虫后第30天的基因表达;SI30,易感马尾松接种松材线虫后第30天的基因表达。
Notes: RI1, Genes expression of resistant P. massoniana at 1day after PWN inoculation; SI1, Genes expression of susceptible P. massoniana at 1day after PWN inoculation; RI15, Genes expression of resistant P. massoniana at 15day after PWN inoculation; SI15, Genes expression of susceptible P. massoniana at 15 days after PWN inoculation; RI30, Genes expression of resistant P. massoniana at 30 day after PWN inoculation; SI30, Genes expression of susceptible P. massoniana at 30 days after PWN inoculation.这些R基因中, NBS-LRR 基因在易感马尾松上皆有高的表达量,而受体丝氨酸/苏氨酸蛋白激酶和 TIR-NBS-LRR 基因表达量变化趋势不完全一致,除1个受体丝氨酸/苏氨酸蛋白激酶基因(c63612.graph_c0)和1个 TIR-NBS-LRR 基因(c65785.graph_c0)外,其余基因在易感马尾松中表达量仍高于高抗马尾松中的表达量。值得注意的是, TIR-NBS-LRR 基因(c65785.graph_c0)仅在高抗马尾松中表达,而在易感马尾松中不论接种与否,在接种后任一时间表达量皆为0,表明该基因可能具有开发为鉴定高抗马尾松分子标记的潜力。
-
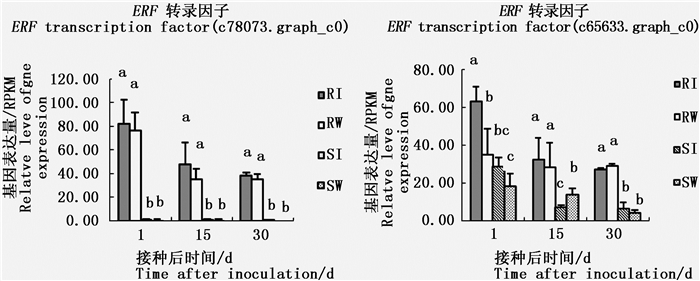
差异基因GO富集分析发现:GO项转录调节生物过程在3个时间点皆达到显著水平。GO项转录调节共包括41个基因,经nr注释有39个为未知基因,剩余的2个为 ERF (Ethylene-responsive factor)转录因子,其中, ERF 转录因子(c78073.graph_c0)在高抗马尾松中的表达量始终显著高于易感马尾松中的表达量(图 3),但不论高抗还是易感马尾松,在接种松材线虫与接种蒸馏水间差异不显著,因此可认为该基因的表达也为组成型。由于该基因在易感马尾松中的表达量极低,约为0,因此该基因也具有开发为检测高抗马尾松分子标记的潜力。而另一个 ERF 转录因子(c65633.graph_c0),在接种松材线虫后第1天和第15天,在高抗马尾松或易感马尾松上基因表达量受松材线虫的影响。如接种松材线虫第1天,该基因在高抗马尾松上的表达量较对照(接种蒸馏水)显著升高;但在接种松材线虫后第30天,高抗马尾松和易感马尾松中该基因在接松材线虫与接种蒸馏水间表达量差异皆不显著。同时结果还显示:3个时间点不论接种松材线虫与否, ERF 转录因子(c65633.graph_c0)在高抗马尾松上的表达量皆高于易感马尾松。由此推测, ERF 转录因子(c78073.graph_c0、c65633.graph_c0)在马尾松抗松材线虫病中可能起正调控作用。

图 3 两个转录因子在高抗和易感马尾松上的表达
Figure 3. Expression of two transcription factors in resistant and susceptible P. massoniana
-
GO项香叶基二磷酸代谢过程在3个时间点也达到显著水平,其中,接种松材线虫后,在高抗与易感马尾松间共发现13个萜类合成相关基因。这13个差异基因中编码(-)-β-蒎烯合酶、(+)-α-蒎烯合酶、(-)-柠檬烯合酶、长叶烯合酶和δ-芹子烯合酶的基因表达量在接种松材线虫和接种蒸馏水处理间差异显著,意味着松材线虫会诱导这些基因的表达量发生显著变化;而编码 GGPPS 的3个基因(c33360.graph_c0、c33943.graph_c0和c58549.graph_c0),不论接种松材线虫与否,3个时间点在高抗与易感马尾松间的表达量差异皆达极显著水平(图 4),表明这种表达差异也为组成型,推测 GGPPS 在抵御松材线虫病的过程中起重要作用。
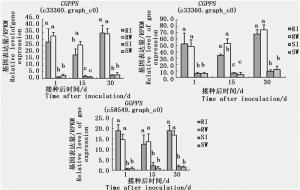
图 4 3个 GGPPS 基因在高抗和易感马尾松上的表达差异
Figure 4. Differential expression of three GGPPS genes in resistant and susceptible P. massoniana
-
为了验证转录组测序结果的可靠性,对接种松材线虫后高抗与易感马尾松间的41个差异基因(包括26个 LRR 抗性基因、2个转录因子和13个萜类相关基因)进行qRT-PCR验证。结果显示:所有基因qRT-PCR结果表达趋势与转录组测序结果一致,相关系数为0.935 8,达极显著水平,表明转录组测序结果较可靠。

图 5 41个差异基因在转录组测序结果与实时定量PCR结果的相关性
Figure 5. Correlation of 41 unigene expression results obtained from qPCR and RNA-seq
-
本研究发现,编码NBS-LRR蛋白及TIR-NBS-LRR蛋白的组成型基因表达量在高抗马尾松与易感马尾松间差异显著。Yamaguchi等[17]分离出一个 NBS-LRR 抗性功能基因 TYNBS1,在西红柿( Lycopersicon esculentum Mill.)防御黄叶卷叶病中至关重要。Grand等[18]研究表明,组成型 HSF 23和 CaMBP 基因在水稻( Oryza sativa L.)抗稻瘟病中起正向调节作用。Wen等[19]在拟南芥( Arabidopsis thaliana (L.) Heynh.)和烟草( Nicotiana tabacum L.)中构建野生华东葡萄( Vitis pseudoreticulata W. T. Wang) VIR-NBS-ARC-LRR 基因的组成型异源过表达增强了对植物病原性真菌和细菌的抗性。王猛[20]利用抑制消减杂交技术分析了松材线虫诱导后马尾松针叶中基因的表达情况,也提出NBS-LRR蛋白等组成型基因在抗松材线虫病过程中发挥了作用。这些结果表明, NBS-LRR 组成型基因参与了马尾松对松材线虫病的防御过程;但本研究发现,接种后1、15和30 d,大部分 NBS-LRR 及 TIR-NBS-LRR 组成型基因在易感马尾松上表达量更高。Sagi等[21]在研究鹰嘴豆( Cicer arietinum Linn.)抵御叶枯病过程中也发现,在早期大多数 NBS-LRR 基因在易感鹰嘴豆中的表达量高于高抗鹰嘴豆。关于LRR-RLKs的作用机制,目前普遍认为植物利用LRR-RLKs蛋白激活MAPK信号途径,从而启动下游的防御反应,如诱发PR、GST等蛋白表达[22]。本研究发现,LRR-RLKs蛋白激酶基因与 NBS-LRR 组成型基因表达量的调节方向一致,皆表现出在易感马尾松上表达量更高。 NBS-LRR 及 TIR-NBS-LRR 组成型基因在易感马尾松上高表达反而使马尾松感病,具体原因有待进一步研究。
乙烯响应因子( ERF )是一个很大的转录因子家族,可参与植物对生物或非生物的胁迫响应。如 ERF 109的过量表达可增强植物对氧化胁迫的抗性,也可减缓细胞程序性死亡的速度和提高植物对盐的耐受力[23]。Berrocal-Lobo等[24]发现,拟南芥中 ERF1的组成型表达即可增强其对尖孢镰刀菌( Fusarium oxysporum Schl.)的抗性,且接种尖孢镰刀菌后 ERF1表达量将上调。Shin等报道,接种线虫后,日本红松( Pinus koraiensis L.)中001_06-B09( AP2/ ERF 转录因子)组成型表达量上调[25]。本研究发现, ERF 在高抗马尾松上的表达量高于易感马尾松上的表达量, ERF 转录因子可能在马尾松抵御松材线虫病中起重要调节作用。
已有研究认为,针叶树种中二萜是主要的防御化合物,松脂对病虫害的防御主要由松节油的化学毒害和二萜的物理阻碍形成,即二萜通过氧化聚合封闭伤口,以防御虫和病菌继续侵入[26-29],其中,二萜由甲基赤藓醇磷酸途径(MEP pathway)合成,GGPPS为异戊基转移酶[30],而 GGPPS 表达量与松脂产量相关, GGPPS 表达量高的马尾松松脂产量也较高[15]。松脂产量则与针叶树种对病虫害的抗性密切相关[31], GGPPS 可能参与了对病虫害胁迫的抗性反应。本研究也发现, GGPPS 在高抗马尾松上的表达量始终远高于易感马尾松上的表达量,由此推测, GGPPS 可能在马尾松抵御松材线虫病的进程中起正调节作用。本研究组后续将对 LRR 基因、乙烯响应因子( ERF )和 GGPPS 等基因进行深入研究。
-
本研究通过表达谱分析揭示,接种松材线虫后第1天高抗与易感马尾松间差异基因最多,这些差异基因主要富集在生物学进程中的氧化-还原过程、跨膜转运、刺激反应、转录调节以及香叶基二磷酸代谢过程等GO项上,其中, LRR 基因、 ERF 转录因子和 GGPPS 可能与马尾松的抗性相关, TIR-NBS-LRR 基因(c65785.graph_c0)、乙烯响应转录因子(c78073.graph_c0)、 GGPPS 基因(c33360.graph_c0、c33943.graph_c0和c58549.graph_c0)为组成型表达基因,具有被用于开发鉴定高抗马尾松分子标记的潜力。
基于高通量转录组测序筛选马尾松抗松材线虫病相关基因
Identification of Candidate Constitutive Expressed Resistant Genes of Pine Wilt Disease in Pinus massoniana Based on High-throughput Transcriptome Sequencing
-
摘要:
目的 研究马尾松抗松材线虫病相关基因,为解析马尾松抗性机理和高抗马尾松早期选择提供理论依据。 方法 对高抗和易感2种马尾松基因型接种松材线虫,在接种后第1、15和30天取样进行转录组高通量测序,通过对高抗和易感马尾松差异基因识别以及富集分析,以筛选与抗松材线虫病相关的组成型差异基因。 结果 对高抗和易感马尾松接种松材线虫后的转录组进行比较,在接种后第1、15和30天分别获得2 866、679和1 657个差异基因,且在接种松材线虫后不同时间点间,共有差异基因相对较少。对差异基因进行GO富集分析,结果显示:在接种后第1、15和30天,最显著的生物学进程是氧化-还原过程,而GO项刺激反应、转录调节以及香叶基二磷酸代谢过程也显著富集。对这几类GO项相关基因进一步分析,发现接种后3个时间点GO项刺激反应中共包括26个R基因,除了2个R基因外,其它R基因表达皆为组成型,在接种松材线虫和对照间差异不显著,而GO项转录调节中仅2个差异基因被nr注释为 ERF 转录因子,其余为未知基因。接种松材线虫后, ERF 转录因子在高抗马尾松上的表达量始终高于易感马尾松上的表达量,且在接种松材线虫和对照间表达量变化不显著,也为组成型基因表达。GO项香叶基二磷酸代谢过程中, GGPPS 3个基因也在高抗马尾松上具有更高表达量,为组成型基因表达。这些基因中 TIR-NBS-LRR 基因(c65785.graph_c0)、 ERF 转录因子(c78073.graph_c0)和3个 GGPPS 基因在高抗马尾松中表达量较高,而在易感马尾松中表达量较低,甚至为0,可作为开发分子标记的候选基因用于鉴定抗性马尾松。 结论 高抗和易感马尾松在接种松材线虫后第1天其表达量差异达到显著水平的基因最多,其中, LRR 基因、 ERF 转录因子和 GGPPS 与马尾松的抗性相关,部分 TIR-NBS-LRR 基因、 ERF 转录因子和 GGPPS 有望被开发为鉴定高抗马尾松的分子标记。 Abstract:Objective To study the genes related to the resistance of Masson pine ( Pinus massoniana ) to pine wood nematode ( Bursaphelenchus xylophilus , PWN). Method Based on two genotypes of resistant and susceptible P. massoniana for inoculating pine wood nematode, and sampling at the 1 dpi, 15 dpi and 30 dpi after inoculation, the authors performed a transcriptome analysis to identify differentially expressed constitutive genes associated with resistance to PWN infected. Result Comparing resistant and susceptible transcriptomes of P. massoniana inoculated with B. xylophilus , 2 866, 679 and 1 657 differentially expressed genes were obtained at 1, 15, or 30 days post-inoculation (dpi), and there were relatively few common differentially expressed genes at different time points. The GO enrichment analysis was performed on the differentially expressed genes, which indicated that the most significant biological process at the 1 dpi, 15 dpi and 30 dpi was oxidation-reduction process, while the stimulus response, transcriptional regulation and the geranyl diphosphate metabolic process in GO items were also significantly enriched. A further analysis of these related genes in the stimulus response GO term, 26 R genes were found at three time points after inoculation. The expression of the other R genes was constitutive except for two R genes, and no significant difference was observed between PWN and water inoculation. In the transcriptional regulation GO term, only two differentially expressed genes of ERF transcription factors were annotated by nr database, and the rest differentially expressed genes were unknown. After inoculation with PWN, the expression level of ERF transcription factors in resistant P. massoniana was always higher than that in susceptible ones, and there was insignificantly difference between trees inoculating PWN and the control. The result means that expression of the two ERF transcription factors is constitutive. In geranyl diphosphate metabolic process GO term, three GGPPS genes having higher expression level in high resistant P. massoniana , are constitutive expression genes. Here, the authors also found that the TIR-NBS-LRR gene (c65785.graph_c0), ERF transcription factor (c78073.graph_c0), and the three GGPPS genes, having higher expression in resistant P. massoniana , the expression levels were extremely low or even zero in susceptible P. massoniana , which suggested that these genes can be chosen as candidate genes for the development of molecular markers to identify resistant P. massoniana . Conclusion Most genes changed the expression level at 1 dpi between resistant and susceptible P. massoniana . The R genes, ERF transcription factors and GGPPS are related to the resistance of P. massoniana , some of which are expected to be developed as molecular markers for identifying PWN-resistance P. massoniana . -
图 1 高抗与易感马尾松在不同接种时间点的差异基因数
Figure 1. The differential genes at different time of inoculation in resistant and susceptible P. massoniana
图 2 马尾松在不同接种时间点的共有差异基因
Figure 2. Common differential genes of P. massoniana at different inoculation time points
图 3 两个转录因子在高抗和易感马尾松上的表达
Figure 3. Expression of two transcription factors in resistant and susceptible P. massoniana
图 4 3个 GGPPS 基因在高抗和易感马尾松上的表达差异
Figure 4. Differential expression of three GGPPS genes in resistant and susceptible P. massoniana
图 5 41个差异基因在转录组测序结果与实时定量PCR结果的相关性
Figure 5. Correlation of 41 unigene expression results obtained from qPCR and RNA-seq
表 1 高抗与易感马尾松接种松材线虫后在不同时间点差异基因的GO富集分析(BP)
Table 1. GO enrichment analysis of different genes at different time of inoculation in resistant and susceptible P. massoniana (BP)
基因本体编号
GO ID类别 Term 差异基因 GO 富集 Different genes GO enrichment 接种后1 d
1d After inoculation接种后15 d
15 d After inoculation接种后30 d
30 d After inoculationGO:0055114 氧化-还原过程 Oxidation-reduction process 2.80E-11 4.80E-11 3.50E-10 GO:0055085 跨膜转运 Transmembrane transport 2.30E-05 4.80E-05 1.10E-05 GO:0008152 代谢过程 Metabolic process 8.90E-05 - - GO:0042026 蛋白质重折叠 Protein refolding 9.70E-05 - - GO:0006886 胞内蛋白转运 Intracellular protein transport 0.000 32 0.000 35 - GO:0009725 激素应答 Response to hormone 0.000 46 0.000 49 - GO:0050896 刺激反应 Response to stimulus 0.000 50 0.000 30 0.000 49 GO:0006950 胁迫应答 Response to stress 0.000 97 0.001 11 0.000 93 GO:0006355 转录调节 Regulation of transcription, DNA-templated 0.001 25 0.001 85 0.001 25 GO:0033383 香叶基二磷酸代谢过程 Geranyl diphosphate metabolic process 0.001 57 0.001 46 0.000 56 GO:0006468 蛋白质磷酸化 Protein phosphorylation - 0.000 18 0.000 17 GO:0006749 谷胱甘肽代谢过程 Glutathione metabolic process - 0.000 42 - GO:0006097 乙醛酸循环 Glyoxylate cycle - - GO:0030435 孢子形成 Sporulation resulting in formation of a cellular spore - - GO:0006886 胞内蛋白转运 Intracellular protein transport - - 0.000 31 GO:0009266 温度刺激应答 Response to temperature stimulus - - 0.002 32 GO:0006464 细胞蛋白质修饰过程 Cellular protein modification process - - 0.002 79  下载: 导出CSV
下载: 导出CSV
表 2 高抗与易感马尾松接种松材线虫后在不同时间点差异基因的GO富集分析(CC)
Table 2. GO enrichment analysis of different genes at different time of inoculation in resistant and susceptible P. massoniana (CC)
基因本体编号
GO ID类别 Term 差异基因 GO 富集 Different genes GO enrichment 接种后1 d
1d After inoculation接种后15 d
15 d After inoculation接种后30 d
30 d After inoculationGO:0016021 膜整体构成 Integral component of membrane 3.60E-24 1.20E-21 3.50E-17 GO:0005886 质膜 Plasma membrane 5.90E-11 2.90E-11 3.60E-11 GO:0009507 叶绿体 Chloroplast 1.10E-09 3.60E-10 6.00E-10 GO:0005829 细胞质基质 Cytosol 3.30E-07 3.10E-07 2.90E-07 GO:0009570 叶绿体基质 Chloroplast stroma 2.20E-05 1.70E-05 1.80E-05 GO:0044464 细胞组分 Cell part 4.80E-05 0.000 12 9.90E-05 GO:0005773 液泡 Vacuole 6.40E-05 5.40E-05 5.80E-05 GO:0005768 核内体 Endosome 9.60E-05 6.10E-05 7.30E-05 GO:0009941 叶绿体被膜 Chloroplast envelope 0.000 16 3.10E-05 0.000 14 GO:0005840 核糖体 Ribosome 0.000 21 1.50E-06 2.00E-05
下载: 导出CSV
表 3 高抗与易感马尾松接种松材线虫后在不同时间点差异基因的GO富集分析(MF)
Table 3. GO enrichment analysis of different genes at different time of inoculation in resistant and susceptible P. massoniana (MF)
基因本体编号
GO ID类别 Term 差异基因 GO 富集 Different genes GO enrichment 接种后1 d
1d After inoculation接种后15 d
15 d After inoculation接种后30 d
30 d After inoculationGO:0016491 氧化还原酶活性 Oxidoreductase activity 2.10E-10 8.80E-09 5.50E-08 GO:0005515 蛋白结合 Protein binding 1.90E-07 5.00E-08 2.00E-07 GO:0000981 DNA特异序列结合 Sequence-specific DNA binding 1.80E-05 4.10E-06 1.40E-05 GO:0003824 催化活性 Catalytic activity 4.60E-05 0.000 14 0.000 19 GO:0005488 结合 Binding 5.00E-05 - - GO:0004721 磷蛋白质磷酸蛋白活性 Phosphoprotein phosphatase activity 0.000 14 - 7.60E-05 GO:0010181 FMN结合 FMN binding 0.000 26 5.60E-05 0.000 17 GO:0008874 葡萄糖酸5-脱氢酶活性 Gluconate 5-dehydrogenase activity 0.000 49 - - GO:0016832 醛裂解酶活性 Aldehyde-lyase activity 0.000 53 - - GO:0004591 酮戊二酸脱氢酶 Oxoglutarate dehydrogenase 0.000 62 0.000 18 0.000 3 GO:0004738 丙酮酸脱氢酶活性 Pyruvate dehydrogenase activity - 0.000 14 0.000 29 GO:0004375 甘胺酸脱氢酶 Glycine dehydrogenase - 0.000 23 0.000 37 GO:0051287 NAD结合 NAD binding - 0.000 39 GO:0016758 转移酶活性 Transferase activity - 0.000 55 0.000 58
下载: 导出CSV
表 4 R基因在高抗和易感马尾松间的差异表达
Table 4. Differential expression of differential R genes in resistant and susceptible P. massoniana
类别 Term 基因编号 Gene_ID 基因表达量 Relative level of gene expression/RPKM RI1 SI1 RI15 SI15 RI30 SI30 受体丝氨酸/苏氨酸蛋白激酶
LRR receptor-like serine/ threonine-protein kinasec63612.graph_c0 11.14 0.78 10.80 0.67 15.65 0.38 c75632.graph_c0 25.21 75.21 - - 8.99 43.55 c74934.graph_c1 9.49 31.04 - - 11.00 43.81 c78638.graph_c0 1.18 11.57 - - 0.30 9.71 c72330.graph_c0 2.55 10.92 - - 1.34 5.37 c52089.graph_c0 0.61 5.38 - - - - c77321.graph_c0 15.01 34.64 - - - - c77212.graph_c0 5.26 13.39 - - - - c76446.graph_c0 7.21 22.56 - - - - c64844.graph_c0 1.10 5.51 - - - - c71845.graph_c0 5.21 40.05 - - - - c72922.graph_c0 - - - - 17.85 60.44 NBS-LRR蛋白
NBS-LRR proteinc79162.graph_c0 0.00 5.56 0.65 5.82 - - c49318.graph_c0 0.40 5.25 0.90 4.83 0.19 5.22 c66133.graph_c0 10.50 30.33 - - 12.53 30.92 c73896.graph_c0 2.48 8.31 - - 3.32 9.25 c76976.graph_c0 11.13 40.39 - - 9.57 29.29 c76662.graph_c0 2.63 7.31 - - - - c62473.graph_c0 1.72 7.55 - - - - TIR-NBS-LRR蛋白
TIR-NBS-LRR proteinc65785.graph_c0 2.68 0.00 - - 2.71 0.00 c73334.graph_c0 0.60 51.29 4.19 16.98 0.60 22.60 c53610.graph_c0 2.44 7.51 - - 2.20 7.67 c76602.graph_c0 1.95 4.46 - - 1.93 3.21 c65671.graph_c0 0.00 3.34 - - 0.00 4.84 c75804.graph_c0 2.88 8.01 - - - - c66096.graph_c0 4.10 23.63 - - - - 注:RI1,高抗马尾松接种松材线虫后第1天的基因表达;S1,易感马尾松接种松材线虫后第1天的基因表达;RI15,高抗马尾松接种松材线虫后第15天的基因表达;SI15,易感马尾松接种松材线虫后第15天的基因表达;RI30,高抗马尾松接种松材线虫后第30天的基因表达;SI30,易感马尾松接种松材线虫后第30天的基因表达。
Notes: RI1, Genes expression of resistant P. massoniana at 1day after PWN inoculation; SI1, Genes expression of susceptible P. massoniana at 1day after PWN inoculation; RI15, Genes expression of resistant P. massoniana at 15day after PWN inoculation; SI15, Genes expression of susceptible P. massoniana at 15 days after PWN inoculation; RI30, Genes expression of resistant P. massoniana at 30 day after PWN inoculation; SI30, Genes expression of susceptible P. massoniana at 30 days after PWN inoculation.
下载: 导出CSV
-
[1] 徐六一, 章健, 高景斌, 等.马尾松抗松材线虫病候选无性系的抗病性测定[J].林业工程学报, 2012, 26(4):27-30. [2] 张盈玉, 马荣才.参与植物防御反应的lrr型蛋白结构与功能[J].中国农业科技导报, 2009, 11(3):12-18. doi: 10.3969/j.issn.1008-0864.2009.03.003 [3] Kandoth P K, Mitchum M G. War of the worms:how plants fight underground attacks[J]. Current Opinion in Plant Biology, 2013, 16(4):457-463. doi: 10.1016/j.pbi.2013.07.001 [4] Cai D, Kleine M, Kifle S, et al . Positional cloning of a gene for nematode resistance in sugar beet[J]. Science, 1997, 275(5301):832-834. doi: 10.1126/science.275.5301.832 [5] 孙岩, 刘宝玲, 苑丽霞, 等.亚麻荠NBS类抗病基因家族的鉴定与分析[J].分子植物育种, 2017, 15(4):1417-1425. [6] Osman R. Isolation and expression of an NBS-LRR protein-encoding resistance gene candidate that segregates with a rust resistance gene in sunflower[J]. Journal of Phytopathology, 2010, 158(6):433-443. [7] 韦善君, 孙振元, 巨关升, 等.冷诱导基因转录因子 CBF1 的组成型表达对植物的抗寒性及生长发育的影响[J].核农学报, 2005, 19(6):465-468. doi: 10.3969/j.issn.1000-8551.2005.06.013 [8] Rawat N, Kumar B, Albrecht U, et al .Genome resequencing and transcriptome profiling reveal structural diversity and expression patterns of constitutive disease resistance genes in Huanglongbing-tolerant Poncirus trifoliata and its hybrids[J]. Horticulture Research, 2017, 4:17064. doi:10.1038/hortres.2017.64. [9] Menéndez-Gutiérrez M, Alonso M, Jiménez E, et al . Interspecific variation of constitutive chemical compounds in Pinus , spp. xylem and susceptibility to pinewood nematode ( Bursaphelenchus xylophilus )[J]. European Journal of Plant Pathology, 2018, 150(6):939-953. [10] Pimentel C S, Firmino P N, Calv~o T, et al . Pinewood nematode population growth in relation to pine phloem chemical composition[J]. Plant Pathology, 2017, 66(5):856-864. doi: 10.1111/ppa.12638 [11] Css D S, de Vasconcelos M W. Identification of genes differentially expressed in Pinus pinaster and Pinus pinea after infection with the pine wood nematode[J]. European Journal of Plant Pathology, 2012, 132(3):407-418. doi: 10.1007/s10658-011-9886-z [12] Santos C S, Pinheiro M, Silva A I, et al . Searching for resistance genes to Bursaphelenchus xylophilus using high throughput screening[J]. BMC Genomics, 2012, 13:599. doi:10.1186/1471-2164-13-599. [13] Hirao T, Fukatsu E, Watanabe A. Characterization of resistance to pine wood nematode infection in Pinus thunbergii using suppression subtractive hybridization[J]. BMC Plant Biology, 2012, 12(1):1-13. [14] Gaspar D, Trindade C, Usié A, et al . Expression profiling in Pinus pinaster in response to infection with the pine wood nematode Bursaphelenchus xylophilus [J]. Forests, 2017, 8(8):279-300. doi: 10.3390/f8080279 [15] Liu Q H, Zhou Z C, Fan H H, et al . Genome-wide identification of differentially expressed genes associated with the high yielding of oleoresin in secondary xylem of masson pine ( Pinus massoniana Lamb.) by transcriptomic analysis[J]. PloS One, 2015, 10(7):e0132624. doi:10.1371/journal.pone.0132624. [16] Liu Q H, Wei Y C, Xu L Y, et al . Transcriptomic profiling reveals differentially expressed genes associated with pine wood nematode resistance in masson pine ( Pinus massoniana Lamb.)[J]. Scientific Reports, 2017, 7:4693. DOI:10.1038/s41598-017-04944-7. [17] Yamaguchi H, Ohnishi J, Saito A, et al . An NB-LRR gene, TYNBS1, is responsible for resistance mediated by the Ty-2 Begomovirus resistance locus of tomato[J]. Theoretical and Applied Genetics, 2018, 131(6):1345-1362. doi: 10.1007/s00122-018-3082-x [18] Grand X, Espinoza R, Michel C, et al . Identification of positive and negative regulators of disease resistance to rice blast fungus using constitutive gene expression patterns[J]. Plant Biotechnology Journal, 2012, 10(7):840-850. doi: 10.1111/j.1467-7652.2012.00703.x [19] Wen Z F, Yao L P, Stacy D S, et al . Constitutive heterologous overexpression of a TIR-NB-ARC-LRR gene encoding a putative disease resistance protein from wild Chinese Vitis pseudoreticulata in Arabidopsis and tobacco enhances resistance to phytopathogenic fungi and bacteria[J]. Plant Physiology and Biochemistry, 2017, 112:346-361 doi: 10.1016/j.plaphy.2017.01.017 [20] 王猛.马尾松抑制消减文库的构建及抗病性相关基因的克隆[D].长沙: 中南林业科技大学, 2010. [21] Sagi M S, Deokar A A, Tar'An B. Genetic analysis of NBS-LRR gene family in chickpea and their expression profiles in response to ascochyta blight infection[J]. Frontiers in Plant Science, 2017, 8:838-852. doi: 10.3389/fpls.2017.00838 [22] Dievart A, Clark S E. LRR-containing receptors regulating plant development and defense[J]. Development, 2004, 131(2):251-261. [23] Bahieldin A, Atef A, Edris S, et al . Ethylene responsive transcription factor ERF109 retards PCD and improves salt tolerance in plant[J]. BMC Plant Biology, 2016, 16(1):216-225. doi: 10.1186/s12870-016-0908-z [24] Berrocal-Lobo M, Molina A. Ethylene response factor 1 mediates Arabidopsis resistance to the soilborne fungus Fusarium oxysporum [J]. Mol Plant Microbe Interact, 2004, 17(7):763-770. doi: 10.1094/MPMI.2004.17.7.763 [25] Shin H, Lee H, Woo K S, et al . Identification of genes upregulated by pinewood nematode inoculation in Japanese red pine[J]. Tree Physiology, 2009, 29(3):411-421. doi: 10.1093/treephys/tpn034 [26] Martin D M, Bohlmann J. Molecular biochemistry and genomics of terpenoid defenses in conifers[J]. Rec Adv Phytochem, 2005, 39(5):29-56. [27] Huber D P, Ralph S, Bohlmann J. Genomic hardwiring and phenotypic plasticity of terpenoid-based defenses in conifers[J]. J Chem Ecol, 2004, 30(12):2399-2418. doi: 10.1007/s10886-004-7942-2 [28] Miller B, Madilao L L, Ralph S, et al . Insect-induced conifer defense:White pine weevil and methyl jasmonate induce traumatic resinosis, de novo formed volatile emissions, and accumulation of terpenoid synthase and putative octadecanoid pathway transcript in Sitka spruce[J]. Plant Physiol, 2005, 137(1):369-382. doi: 10.1104/pp.104.050187 [29] Keeling C J, Bohlmann J. Genes, enzymes and chemicals of terpenoid diversity in the constitutive and induced defense of conifers against insects and pathogens[J]. New Phytol, 2006, 170(4):657-675. doi: 10.1111/j.1469-8137.2006.01716.x [30] Thabet I, Guirimand G, Guihur A, et al . Characterization and subcellular localization of geranylgeranyl diphosphate synthase from Catharanthus roseus [J]. Mol Biol Rep, 2012, 39(3):3235-3243. doi: 10.1007/s11033-011-1091-9 [31] Strom B L, Goyer R A, Ingram Jr L L. Oleoresin characteristics of progeny of loblolly pines that escaped attack by the southern pine beetle[J]. For Ecol Manage, 2002, 158(1):169-178. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4447
- HTML全文浏览量: 2518
- PDF下载量: 118
- 被引次数: 0
