



-
中华紫胶虫(Kerria chinensis Mahdihassan)隶属于半翅目(Hemiptera)、胶蚧科(Tachardiidae)、胶蚧属(Kerria),是一种具有重要经济价值的资源昆虫,主要寄生于交趾黄檀(Dalbergia cocrinchinensis Pierre ex Laness)、光叶合欢(Aibizia lucida (Steud.) Nielsen)等植物上,通过吸取植物韧皮部的汁液生长繁殖。紫胶是雌虫通过腺体分泌出的一种纯天然的树脂,主要成分由紫胶树脂、紫胶蜡、紫胶色酸等组成。紫胶是重要天然林产化工原料,加工后被广泛应用于军工、日用化工、电子等行业,因而具有重要的经济价值[1-2]。前人在紫胶虫研究方面已做了大量的工作,但主要是围绕紫胶虫生物学特征、生态适应性、寄主植物、遗传多样性等方面展开的,在紫胶合成的分子机理、泌胶相关基因的验证等方面的研究尚未见报道[3-6]。
紫胶树脂是由羟基脂肪酸和倍半萜烯酸酯相互形成的内酯及交酯混合而成,结构比较复杂[2]。羟基脂肪酸是紫胶树脂的主要组成部分,与之相关的脂肪酸合成代谢途径,其主要由脂肪酸合成酶(FAS)、超长链脂肪酸延伸酶(ELO)、脂肪酸去饱和酶(FAD)等参与完成,其中,脂肪酸去饱和酶(FAD)在脂肪酸代谢过程中发挥着重要作用。FAD在卷叶蛾、烟草蛾、小菜蛾等蛾类以及意大利蜜蜂、家蚕等昆虫中已有广泛的研究[7-10],研究发现,脂肪酸去饱和酶Δ6(FAD 6)是合成长链多不饱和脂肪酸(LC-PUFAs)的限速酶,在近十几年的研究中引起了广泛的关注[11],而在果蝇中对FAD研究的比较系统,其中果蝇的fad1为第一个报道的昆虫FAD[12],fad1具有Δ9功能特性,最优底物为棕榈酸,能通过激素来调控果蝇的脂质代谢[13]。在紫胶树脂中紫胶桐酸(9, 10, 16-三羟基棕榈酸)是其主要成分[2],因此,推测FAD基因在紫胶合成通路中起着重要的调控作用。
RNA干扰(RNAi)是指在真核生物中,由双链RNA(dsRNA)诱发同源mRNA降解,使靶基因表达沉默的现象[14]。RNAi转染有注射、喂食、浸泡、病毒感染和转基因等方法,较常用的是注射法和喂食法。在RNA干扰研究的初期,注射最初主要应用于线虫[14],并随着在果蝇中运用的开展[15],目前已在小菜蛾(Plutella xyllostella Linnaeus)、褐飞虱(Nilaparvata lugens Stal)、赤拟谷盗(Tribolium castaneum Herbst)等研究对象中建立了注射dsRNA诱导RNAi的体系[16-20];但当研究对象个体过小时,注射难度增大且成活率降低这一问题仍无法得到有效解决。喂食法诱导RNAi因其操作简单方便,对研究对象危害性小等特点已在马铃薯甲虫(Leptinotarsa decemlineata Say)、麦长管蚜(Sitobion avenae Fabricius)等昆虫中得到应用[21-22],但喂食法存在作用较慢、效率较低的缺点[23-24]。中华紫胶虫体型较小,幼虫在寄主植物上寻找适宜的枝条将口针刺入树皮下固定取食后将终生不动。由于注射法和喂食法各自存在缺点,在中华紫胶虫个体过小且不能喂食时,使用哪种转染方法将dsRNA转染到其体内显得至关重要。涂干法一般用于蛀干害虫的防治,具体方法为将药剂通过毛刷等工具直接涂抹于树干上。选择性的给药方法使药剂更有效的接触虫体,并避免了大量无选择性给药给环境带来的影响。这样的给药方法恰好适用于本实验寄生于树干上的紫胶虫。前人研究发现,dsRNA在体外有高稳定特性,将其喷洒在植物叶片上,几天之后仍然可以保持相对较高的稳定性[25]。鉴于上述原因,本实验拟采用虫体涂抹法将dsRNA转染到紫胶虫体内,对其FAD基因的功能进行验证。
在野外对中华紫胶虫进行RNAi实验时,dsRNA的需求量很大,所以需要选择一种可大量较低成本获得的dsRNA的方法。目前,dsRNA的制备主要方法有试剂盒法、大肠杆菌诱导法和转基因植物法等,其中,用试剂盒法实验操作简单、耗时少等特点,但其成本高且产量低,不易长时间保存,不适用野外大用量实验;转基因植物法操作周期长,而且得到目的植株概率低;利用原核表达系统制备dsRNA,有较低的成本并能够获得大量dsRNA。基于大肠杆菌诱导法的优势,其制备dsRNA的方法已在家蚕(Bombyx mori Linnaeus)、棉铃虫(Helicoverpa armigera Hubner)、线虫等物种中广泛应用[26-28]。为了能够较低成本获得dsRNA用于中华紫胶虫RNAi实验,本研究通过采用大肠杆菌诱导法构建中华紫胶虫FAD基因RNAi载体,采用菌液涂抹的方式将dsRNA转染到紫胶虫体内,检测FAD基因对紫胶泌胶量的影响,从而为在分子水平上验证紫胶合成相关基因功能提供科学依据,也为进一步研究紫胶虫泌胶机理奠定基础。
-
中华紫胶虫采自中国林业科学研究院资源昆虫研究所元江试验站(102°00′46" E,23°36′11" N)。DH5α感受态细胞购买于天根生化科技(北京)有限公司,载体L4440购自Addgene公司,菌株HT115由BioVector质粒载体菌种细胞基因保藏中心提供。
-
利用RNA提取试剂盒(生工生物工程(上海)股份有限公司)提取中华紫胶虫总RNA,用PrimeScriptTM RT reagent Kit试剂盒(Takara公司)反转录得到cDNA第一链。根据转录组测序得到的FAD基因片段,通过siDirect version 2.0分析FAD碱基序列,找到干扰位点较多的片段,从而尽可能的避免脱靶效应。使用Primer-Premier 6.0设计引物,并通过对引物的特异性检测等预实验筛选,最终确定一条266 bp片段作为RNA干扰片段。在合成引物的正反向引物中添加限制性内切酶(Sal I和Sac I,下划线标记)位点(表 1)。所有的引物由生工生物工程(上海)股份有限公司合成。
表 1 dsRNA和RT-qPCR的引物
Table 1. Primers used for dsRNA synthesis and RT-qPCR
基因
Gene引物
Primers引物序列
Sequence of primer(5’→3’)长度
Product/bpFAD ds-f ATGAGCTCGCAATGATACAACGAACCA 266 ds-R ATGTCGACGAACGATGTGACCATAAGC RT-F CATCGTTCTTACAAGGCTAA 142 RT-R TATGTGGATCGGCATTCG β-actin RT-F ATCGTGCTGAGTGAGGAA 143 RT-R CGCTTCGCTGATTATCGTA -
以制备好的cDNA为模板,使用引物对FAD基因进行扩增,反应条件为95℃,2 min预变性后,95℃,30 s变性,50℃,30 s退火,72℃,1 min延伸,共34个循环,72℃终延伸10 min后,4℃保存。获得的PCR产物用1.0%的琼脂糖凝胶进行电泳并割胶回收。
-
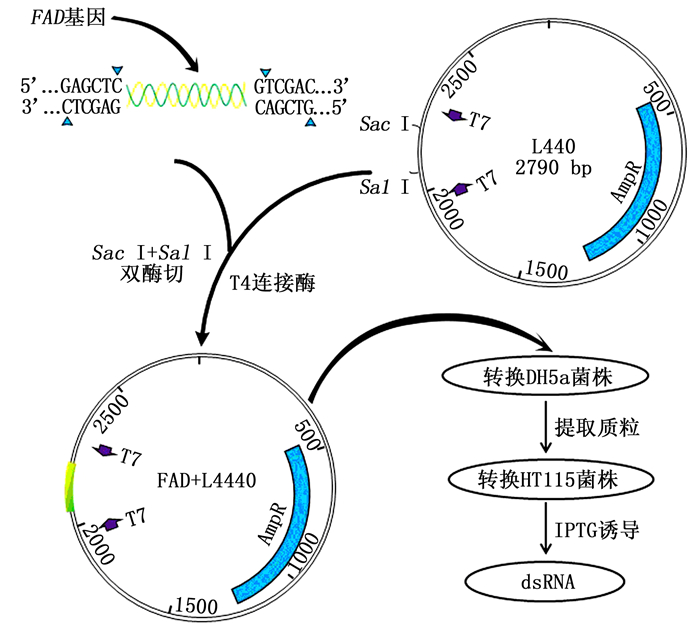
将FAD回收的产物和L4440载体用Sac I和Sal I(NEB(北京)有限公司)进行双酶切,酶切产物通过琼脂糖凝胶电泳检测之后利用T4连接酶连接过夜。将其连接产物转化到DH5α感受态细胞后,涂布于含有氨苄青霉素的SOC固体培养基上,培养14 h左右,挑取单菌落于SOC液体培养基中培养。对其构建的FAD-L4440重组质粒做菌液PCR验证并提取质粒进一步通过测序进行验证。
-
将构建好的FAD-L4440重组质粒转化到HT115感受态菌株中,涂布于含有氨苄青霉素和四环素的SOC固体培养基上,培养14 h左右,挑取单菌落转入液体培养基37℃过夜培养,然后将过夜培养液加入2×YT液体培养基中培养至OD600=0.4,加入IPTG进行诱导,继续摇床培养4 h后收集菌体。
-
用无菌水稀释菌体,配制成高(620 ng·mL-1)、中(62 ng·mL-1)、低(6.2 ng·mL-1)不同浓度的测试菌液,并以不做任何处理的自然组(ck)和L4440-HT115菌液作为实验对照组。转染时期选择紫胶虫幼虫期,在幼虫期其少量泌胶,虫体表面没有紫胶包被,菌液可以直接作用于虫体表面,通过紫胶虫表面的蜡腺和气孔进入体内,每天将不同浓度的测试液和对照液涂抹在中华紫胶虫幼虫体表面,每天涂抹2次,连续处理3 d后收集12、24、48、72 h的实验样品,1个月后收集实验组和对照组的样品对其个体泌胶量进行测定。
-
使用Primer-Premier 6.0设计荧光定量引物(表 1),利用RNA提取试剂盒(生工生物工程(上海)股份有限公司、上海)提取实验组和对照组的总RNA,使用PrimeScriptTM RT reagent Kit试剂盒(Takara公司)进行反转录;之后以相对定量的方法进行基因表达量检测。选择β-actin基因作为内参,并对每个样进行3次独立的生物学重复,采用2-ΔΔCT法计算相对表达量[29]。
-
提取中华紫胶虫中总RNA并反转录为cDNA后进行PCR扩增,根据FAD基因序列,预期扩增片段大小为266 bp,扩增片段的电泳检测结果显示介于250~500 bp之间(图 2-A),与预期相符。
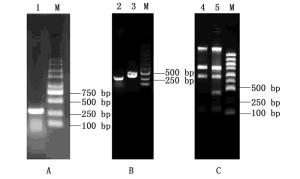
图 2 干扰载体构建和dsRNA诱导表达
Figure 2. The construction of RNAi plasmid and the induction dsRNA
-
根据图 1载体构建的流程,FAD基因胶回收产物和L4440载体经Sac I和Sal I双酶切,分别回收酶切产物,利用T4连接酶连接过夜。将FAD-L4440连接好后转入到50 μL DH5α感受态细胞中,涂平板培养12 h,挑取单菌落在液体培养基中培养。经T7单引物菌液PCR扩增后,获得预期的目的条带(图 2-B),并送公司测序,结果显示与目的片段吻合,验证FAD-L4440重组质粒已成功构建。
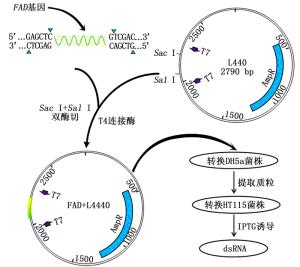
图 1 干扰载体构建及dsRNA诱导表达流程
Figure 1. Schematic presentation of RNAi plasmid construction and dsRNA producing process
-
将FAD-L4440重组质粒转入到HT115感受态细胞中,扩大培养至菌液OD600为0.4,加入IPTG诱导4 h。诱导后的菌液提总RNA检测,结果见图 2-C。凝胶电泳结果显示:诱导后FAD-L4440-HT115菌液提取的RNA含有1条目的基因的RNA条带,而诱导前FAD-L4440-HT115菌液提取的RNA不含此条带,据此可以判定诱导后的FAD-L4440-HT115特异条带为FAD的dsRNA条带,证明FAD基因dsRNA的诱导表达成功。
-
采用虫体涂抹法将不同浓度的测试液和对照液分别涂抹在虫体上,每天2次,连续3 d后,收集不同时间段的样品,利用RT-qPCR检测干扰效果。以12 h的L4440-HT115菌液组的mRNA表达水平为对照,以中华紫胶虫细胞质肌动蛋白(β-actin)的mRNA表达水平作为内参。结果表明:使用涂抹FAD-L4440-HT115菌液处理组的FAD基因表达量明显降低,其中,低浓度在12 h和中浓度在24 h时相对于对照组有显著差异,分别降低了90.90%和86.52%,中浓度在72 h时干扰效率高于其他两个组(图 3)。

图 3 不同浓度菌液涂抹对中华紫胶FAD基因的影响
Figure 3. The effects of smering stem different E· coli on the FAD gene of K. chinensis
-
采集不同处理的紫胶,剥取新鲜胶块称质量,用95%酒精浸泡胶块,待胶完全溶解后,过滤取虫体统计个体数并称质量,然后计算其个体泌胶量。结果表明,虫体涂抹法的低浓度和中浓度处理组相比ck组个体泌胶量有显著性差异(P<0.05),分别降低了14.89%和12.73%(图 4)。
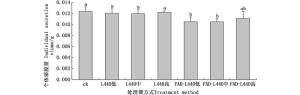
图 4 RNAi后个体泌胶量测定
Figure 4. The average amount of lac secret after RNAi teratment
-
中华紫胶虫一年两代,雌虫通过腺体分泌紫胶,其幼虫期泌胶量较少,成虫期大量分泌紫胶逐渐形成胶壳将虫体和寄主植物枝条覆盖起来,中华紫胶虫成虫中后期个体泌胶速率最大,整个生活周期个体泌胶量在17~30 mg[2]。为了能够让dsRNA菌液直接作用于虫体表面从而通过其体表的紫胶腺体、蜡腺和气孔进入体内,选择合适的虫期进行干扰试验是整个实验设计中关键的一步。由于紫胶虫大量泌胶时期虫体已完全被紫胶包被,dsRNA菌液很难被虫体吸收,所以本实验选择在紫胶虫幼虫期对其进行转染。
在昆虫RNAi研究中,选择一种合适的转染方法将其导入到昆虫体内是实验成功的前提,目前主要的转染方法有注射法、喂食法、浸泡法和转基因等方法。在紫胶虫的RNAi实验中,笔者发现常规的转染方法对紫胶虫并不适用,主要原因是中华紫胶虫雌成虫虫体长只有6 mm左右,体宽4 mm左右,个体较小[1]。紫胶虫寄生于寄主植物树干,幼虫以口针刺入寄主植物韧皮部,吸食寄主植物汁液,口针一旦刺入寄主植物,便不再拔出,终生不动,如果将紫胶虫从树干上取下,造成的机械伤害会使紫胶虫很快死亡;且随着紫胶虫的生长发育,紫胶虫可分泌少量蜡丝及大量紫胶,蜡丝及大量的紫胶将紫胶虫虫体完全覆盖,且紫胶致密坚硬,对紫胶虫有很好保护作用[1]。因此,选择适合的转染方式对紫胶虫进行RNAi至关重要。本课题组前期已使用涂干法将外源保幼激素类似物(JHA)涂抹在白蜡虫体表,结果表明,外源激素可通过白蜡虫体表的腺体进入体内,提高白蜡虫2龄雄幼虫的个体泌蜡量和羽化率[30]。根据涂干法在外源激素导入白蜡虫中的成功运用及dsRNA在体外有较高的稳定性的特点,结合紫胶虫体小且不能喂食不适用注射和喂食等方法,本实验采用虫体涂抹法将dsRNA以不同浓度配比施于虫体表面,成功的将目的基因导入虫体,为RNAi转染过程中不适用注射法、喂食等转染方法的昆虫提供技术参考。
适合的菌液浓度是RNAi实验得以顺利进行的重要保障,不同的昆虫发生显著的RNAi干扰效应时,所需要的dsRNA的剂量不同。如在给烟草天蛾(Manduca sexta Linnaeus)注射少量的dsRNA就有明显的沉默效果[31],在甜菜夜蛾(Spodoptera exigua Hiibner)中需要大量的dsRNA才有较为显著的干扰效果[32]。是本实验表明,当菌液浓度为6.2 ng·mL-1时,对紫胶虫的基因表达量及泌胶量影响最为明显。本实验对需要以虫体涂抹法进行RNAi实验的浓度阈值提供了参考。
RNAi的时效性一直备受关注,不同的物种其RNAi持续的时间也不尽相同,因而检测RNAi效果的时间点就显得尤为重要[31]。检测的时间过早,很有可能RNAi效应还未开始,则检测不到预期的干扰效果;检测时间过晚,则可能由于mRNA恢复至正常水平,干扰效果可能大幅度降低,也不能检测到RNAi的效果[33]。本实验在大量重复预实验的基础上,发现FAD基因最佳干扰效果出现在干扰之后12 h左右。
紫胶树脂是紫胶的主要成分,由于其本身的复杂性,对于树脂的各组分酸含量不同的研究者所得的结果不同,但大多数研究者认为紫胶酮酸(9, 10, 16-三羟基棕榈酸)和壳脑醛酸是树脂的主要成分[2]。脂肪酸去饱和酶(FAD)被认为是长链多不饱和脂肪酸合成的关键酶,在脂肪酸代谢过程中发挥了重要作用[34]。Dallerac等的研究结果表明,fad1具有Δ9功能特性,能通过激素来调控果蝇的脂质代谢[13],其最优底物为棕榈酸,而在紫胶树脂中紫胶桐酸(9, 10, 16-三羟基棕榈酸)是棕榈酸的主要成分,因此,我们推测FAD基因对紫胶合成通路起着重要的调控作用。为了验证这一猜想,通过构建FAD基因RNA干扰载体并将其转染到紫胶虫体内,从而对其功能进行初步验证,实验结果表明,干扰后FAD基因表达量明显降低,其中低浓度在12 h和中浓度在24 h时相对于L4440-HT115菌液对照组有显著差异,分别降低了90.90%和86.52%。不同处理组的个体泌胶量相比ck组个体泌胶量有所差异,其中,低浓度处理对紫胶虫个体泌胶量有明显影响。表明FAD基因对紫胶虫泌胶有直接的调控作用。
-
本研究在中华紫胶虫FAD基因功能验证中引入了细菌表达dsRNA的RNAi系统,并利用虫体涂抹法将其成功导入虫体内,为探索高效、便捷的昆虫功能基因的研究方法提供参考,将为后续解析紫胶合成的分子机理提供技术基础与科学依据。
中华紫胶虫FAD基因RNA干扰载体构建与功能初步分析
Construction of RNA Interference Vector and Function Analysis of FAD Gene in Kerria chinensis
-
摘要:
目的 引入细菌表达dsRNA干扰系统对中华紫胶虫FAD基因进行RNA干扰,通过检测干扰后FAD基因表达量与个体泌胶量动态变化,对FAD基因功能进行初步分析,为验证紫胶合成相关基因功能提供科学基础。 方法 中华紫胶虫FAD基因和L4440载体经双酶切之后,利用T4连接酶连接,构建FAD-L4440重组质粒,并转入HT115感受态细胞中,IPTG诱导表达dsRNA后,通过虫体涂抹法将dsRNA转染到紫胶虫体内。用RT-qPCR检测干扰后FAD基因的表达量并测定干扰后个体泌胶量的变化。 结果 RNA干扰后FAD基因表达量明显降低,其中,以低浓度菌液处理后12 h和中浓度菌液处理后24 h相对于对照组有显著差异,分别降低了90.90%和86.52%,中浓度在72 h时干扰效率高于其他两个组,个体泌胶量与对照组比较有显著下降。 结论 本研究构建了中华紫胶虫FAD基因的RNA干扰载体,在基因功能验证中引入了细菌表达dsRNA的RNAi系统,使用虫体涂抹法将其导入虫体内引起FAD基因表达量与紫胶虫个体泌胶量显著下降,为RNAi转染过程中不适用注射法、喂食等转染方法的昆虫提供技术参考,为后续解析紫胶合成的分子机理提供技术基础与科学依据。 Abstract:Objective In order to verify the function of FAD gene of Kerria chinensis, the RNAi system by bacterial expression of dsRNA was used and the function of FAD gene was preliminarily analyzed to provide references for verifying the function of genes related to lac synthesis. Method The FAD gene of K. chinensis and L4440 vector were digested by double enzymes primarily, then the FAD-L4440 recombinant plasmid was constructed by T4 ligase ligation and transferred into HT115 competent cells as well. After the expression of dsRNA induced by IPTG, the dsRNA was transfected into lac insect by spreading it to the trunks. Detected the FAD gene expression after RNAi by RT-qPCR and the amount of individual lac secreted was also measured timely. Result The expression of FAD gene significantly decreased. There was a significant difference between the control group and the low concentration bacterial solution treated for 12 hours and the medium concentration bacterial solution treated for 12 hours, which decreased by 90.79% and 85.46% respectively. The interference efficiency of medium concentration at 72 hours was higher than that of the other two groups and there was a significant difference in the amount of individual lac secretion in comparison with the control group. Conclusion In this study, the RNAi vectors of FAD gene of K. chinensis were successfully constructed. In the verification of gene function, the RNAi system was used for bacterial expression of dsRNA. The expression of FAD gene and the amount of individual lac secretion decreased significantly after imported into the insect body. It provides a new technical reference for insects which not suitable for injection and feeding for RNAi transfection and it also provides a technical and scientific basis for the verification of molecular mechanism related to lac biosynthesis. -
Key words:
- Kerria chinensis
- / RNA interference
- / FAD gene
- / L4440
-
图 2 干扰载体构建和dsRNA诱导表达
Figure 2. The construction of RNAi plasmid and the induction dsRNA
图 1 干扰载体构建及dsRNA诱导表达流程
Figure 1. Schematic presentation of RNAi plasmid construction and dsRNA producing process
图 3 不同浓度菌液涂抹对中华紫胶FAD基因的影响
Figure 3. The effects of smering stem different E· coli on the FAD gene of K. chinensis
表 1 dsRNA和RT-qPCR的引物
Table 1. Primers used for dsRNA synthesis and RT-qPCR
基因
Gene引物
Primers引物序列
Sequence of primer(5’→3’)长度
Product/bpFAD ds-f ATGAGCTCGCAATGATACAACGAACCA 266 ds-R ATGTCGACGAACGATGTGACCATAAGC RT-F CATCGTTCTTACAAGGCTAA 142 RT-R TATGTGGATCGGCATTCG β-actin RT-F ATCGTGCTGAGTGAGGAA 143 RT-R CGCTTCGCTGATTATCGTA  下载: 导出CSV
下载: 导出CSV
-
[1] 陈晓鸣.紫胶虫生物多样性研究[M].昆明:云南科技出版社, 2005, 1-109. [2] 陈晓鸣, 陈又清, 张弘, 等.紫胶虫培育与紫胶加工[M].北京:中国林业出版社, 2008, 170-179. [3] 陈航, 陈晓鸣, 冯颖, 等.紫胶虫主要生产种的RAPD分子标记分析[J].林业科学研究, 2006, 19(4):423-430. doi: 10.3321/j.issn:1001-1498.2006.04.003 [4] 刘凤书, 易鹏, 李绍家.紫胶虫的泌胶性能与寄主植物化学成份的关系[J].林业科学, 1988, 24(1):106-112. [5] Chen H, Chen X M, Feng Y, et al. Molecular phylogeny and biogeography of lac insects (Hemiptera:Kerriidae) inferred from nuclear and mitochondrial gene sequences[J]. Molecular Biology Reports, 2013, 40(10):5943-5952. doi: 10.1007/s11033-013-2701-5 [6] Ranjan S K, Mallick C B, Saha D, et al. Genetic variation among species, races, forms and inbred lines of lac insects belonging to the genus Kerria (Homoptera, Tachardiidae)[J]. Genetics & Molecular Biology, 2011, 34(3):511-519. [7] Ding B J, Carraher C, Löfstedt C. Sequence variation determining stereochemistry of a Δ11 desaturase active in moth sex pheromone biosynthesis[J]. Insect Biochem Molecular Biology, 2016, 74:68-75. doi: 10.1016/j.ibmb.2016.05.002 [8] Matousková P, Pichová I, Svatos A. Functional characterization of a desaturase from the tobacco hornworm moth (Manduca sexta) with bifunctional Z11-and 10, 12-desaturase activity[J]. Insect Biochem Molecular Biology, 2007, 37(6):601-610. doi: 10.1016/j.ibmb.2007.03.004 [9] Lee D W, Kim Y, Koh Y H. RNA interference of PBAN receptor suppresses expression of two fatty acid desaturases in female Plutella xylostella[J]. Journal of Asia-Pacific Entomology, 2011, 14(4):405-410. doi: 10.1016/j.aspen.2011.05.004 [10] Wang Y, Da Cruz T C, Pulfemuller A, et al. Inhibition of fatty acid desaturases in Drosophila melanogaster larvae blocks feeding and developmental progression[J]. Archives of Insect Biochemistry and Physiology, 2016, 92(1):6-23. [11] Alonso D L, Maroto F G, Ruiz J R, et al. Evolution of the membrane-bound fatty acid desaturases[J]. Biochemical Systematics and Ecology, 2003, 31(10):1111-1124. doi: 10.1016/S0305-1978(03)00041-3 [12] Wicker-Thomas C, Henriet C, Dallerac R. Partial characterization of a fatty acid desaturase gene in Drosophila melanogaster[J]. Insect Biochemistry & Molecular Biology, 1997, 27(11):963. [13] Dallerac R, Labeur C, Jallon J M, et al. A delta 9 desaturase gene with a different substrate specificity is responsible for the cuticular diene hydrocarbon polymorphism in Drosophila melanogaster[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(17):9449-9454. doi: 10.1073/pnas.150243997 [14] Fire A, Xu S Q, Montgomery M K, et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans[J]. Nature, 1998, 391(6669):806-811. doi: 10.1038/35888 [15] Kennerdell J R, Carthew R W. Use of dsRNA-mediated genetic interference to demonstrate that frizzled and frizzled 2 act in the wingless pathway[J]. Cell, 1998, 95:1017-1026. doi: 10.1016/S0092-8674(00)81725-0 [16] Hu Z, Lin Q, Chen H, et al. Identification of a novel cytochrome P450 gene, CYP321E1 from the diamondback moth, Plutella xylostella (L.) and RNA interference to evaluate its role in chlorantraniliprole resistance[J]. Bulletin of Entomological Research, 2014, 104:716-723. doi: 10.1017/S0007485314000510 [17] Guo Z J, Kang S, Zhu X, et al. The novel ABC transporter ABCH1 is a potential target for RNAi based insect pest control and resistance management[J]. Scientific Reports, 2015, 10.1038/srep13728. doi: 10.1038/srep13728 [18] Xu H J, Xue J, Lu B, et al. Two insulin receptors determine alternative wing morphs inplanthoppers[J]. Nature, 2015, doi:10.1038/nature14286. [19] Yang M M, Zhao L N, Shen Q D, et al. Knockdown of two trehalose-6-phosphate synthasesseverely affects chitin metabolism gene expression in the brown planthopper Nilaparvata lugens[J]. Pest Management Science, 2017, 73:206-216. doi: 10.1002/ps.4287 [20] Tang B, Wei P, Zhao L N, et al. Knockdown of five trehalase genes using RNA interference regulates the gene expression of the chitin biosynthesis pathway in Tribolium castaneum[J]. BMC Biotechnology, 2016, 16:67. doi: 10.1186/s12896-016-0297-2 [21] Kong Y, Liu X P, Wan P J, et al. The P450 enzyme Shade mediates the hydroxylation of ecdysone to 20-hydroxyecdysone in the Colorado potato beetle, Leptinotarsa decemlineata[J]. Insect Molecular Biology, 2014, 23:632-643. doi: 10.1111/imb.12115 [22] Xu L, Duan X, Lv Y, et al. Silencing of an aphid carboxylesterase gene by use of plant-mediated RNAi impairs Sitobion avenae tolerance of Phoxim insecticides[J]. Transgenic Research, 2014, 23(2):389-396. doi: 10.1007/s11248-013-9765-9 [23] 刘吉升, 朱文辉, 廖文丽, 等.昆虫RNA干扰中双链RNA的转运方式[J].昆虫学报, 2016, 59(6):682-691. [24] 王伟伟, 刘妮, 陆沁, 等. RNAi技术的最新研究进展[J].生物技术通报, 2017(11):41-46. [25] Tenllado F, MartínezGarcía, Belén, Vargas M, et al. Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections[J]. BMC Biotechnology, 2003, 3(1):3. [26] 王根洪, 祝慧敏, 罗会松, 等.细菌表达dsRNA介导的家蚕FTZ-F1基因的RNA干扰[J].昆虫学报, 2011, 54(5):596-601. [27] Timmons L, Court D L, Fire A. Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans[J]. Gene, 2001, 263(1):103-112. [28] Zhang X, Liu X, Ma J, et al. Silencing of cytochrome P450 CYP6B6 gene of cotton bollworm (Helicoverpa armigera) by RNAi[J]. Bulletin of entomological research, 2013, 103(5):1-8. [29] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method[J]. Methods, 2001, 25(4):402-408. doi: 10.1006/meth.2001.1262 [30] 刘妮, 陆沁, 张金稳, 等.外源保幼激素类似物对白蜡虫泌蜡和发育的影响[J].林业科学研究, 2018, 31(2):118-123. [31] Levin D M, Breuer L N, Zhuang S, et al. A hemocyte-specific integrin required for hemocytic encapsulation in the tobacco hornworm, Manduca sexta[J]. Insect Biochemistry & Molecular Biology, 2005, 35(5):0-380. [32] Terenius O, Papanicolaou A, Garbutt J S, et al.RNA interference in Lepidoptera:An overview of successful and unsuccessful studies and implications for experimental design[J]. Journal of Insect Physiology, 2011, 57(2):231-245. doi: 10.1016/j.jinsphys.2010.11.006 [33] 刘婷, 秦国华, 张建珍, 等.东亚飞蝗谷胱甘肽S-转移酶RNA干扰效率研究[J].应用昆虫学报, 2011, 48(4):820-825. [34] 曾硕士, 江黎明, 元冬娟.脂肪酸去饱和酶的研究进展[J].生命科学, 2008, 20(5):816-820. doi: 10.3969/j.issn.1004-0374.2008.05.031 -

 点击查看大图
点击查看大图

图(4) / 表(1)
计量
- 文章访问数: 5302
- HTML全文浏览量: 2361
- PDF下载量: 68
- 被引次数: 0
