

 下载:
下载:

-
叶片作为重要的光合器官,是植物光合产物形成的主要场所。叶片干物质量与其叶面积之比定义为比叶质量(Leaf mass per area,LMA),可以理解为叶片水平上光的截留成本[1]。比叶质量是植物叶片关键叶性状之一,是植物生长的重要特征,也是植物生态学、农业和林业上广泛应用的,综合反应植物利用资源的能力以及适应环境的生长策略的重要指标[2-4]。比叶质量在植物生长策略中发挥核心作用,具有低比叶质量的物种倾向于快速的资源获取、快速代谢和增长,高比叶质量物种倾向于较强的资源节约、保护能力和持久性。国内外大量关于比叶质量(LMA)的文献数据分析表明,比叶质量在物种间的变化超过100倍[5]。
植物比叶质量是影响各种生态系统过程的重要因素[5],但不同植物功能群比叶质量随环境梯度可塑性变化的生理调节机制尚不清楚。Poorter等在室内控制条件下通过实验构建比叶质量随环境因子的响应曲线,结果显示,比叶质量随光照、温度和淹水变化而存在很大变化,随CO2浓度、养分及其水分胁迫变化比较适度[5]。Wright等基于全球175个样点2 548种植物的叶性分析表明,比叶质量随降雨量的减少和光照辐射的增加而增大[6]。薛立等研究发现6种阔叶幼苗叶片的比叶质量随幼苗种类和季节而变化,幼苗比叶质量与降雨量、平均气温和光照时间存在二项式关系[2]。
毛竹(Phyllostachys edulis (Carrière) J.Houz.)具有生长快、成材早、产量高、用途广、收益大等优点,是我国南方的重要森林资源。当今全球森林面积急剧下降,竹林面积却以3%的速度递增[7],这意味着竹林是一个不断增大的碳汇。然而,全球气候变化背景下,极端天气气候事件发生频度和强度越来越大,对生态系统结构和功能造成前所未有的威胁。2013年夏季,我国南方出现广泛的、持续时间较长的高温干旱,毛竹林生态系统也遭受严重破坏。水分作为植物生长的重要条件,与毛竹生产力有着密切联系,因此干旱对毛竹林生产力具有重要潜在影响。毛竹研究涉及内容较丰富[8-11],但作为重要生长策略指标,毛竹比叶质量随生长季节、冠层高度、年龄的动态变化及其对干旱适应性特征方面的研究较少[11]。因此,本研究结合毛竹野外人工截雨模拟干旱试验,针对毛竹比叶质量时空动态变化及其对模拟干旱响应进行系统研究,通过阐明不同生长季节、不同冠层部位叶片和不同年龄个体的比叶质量时空动态格局,间接揭示叶片比叶质量在毛竹适应光、水环境的调控策略,旨在探索毛竹季节性资源利用能力的快速判断指标,为全球气候背景下毛竹人工林可持续高效经营提供理论依据。
-
试验在国家林业和草原局钱江源森林生态系统定位观测研究站庙山坞辅站进行,位于杭州西郊富阳区境内(119°56′~120°02′E,30°03′~30°06′N)。该区域地形属浙西低山丘陵区天目山系余脉,属于中亚热带季风气候区,年平均气温16.1℃,最高气温40.2℃,最低气温-14.4℃,多年平均降水量1 441.9 mm。试验期间(2014—2018年),样地内年平均降水量1 461.8 mm。土壤属于酸性红壤。试验区毛竹林为20世纪60年代种植,处于粗放经营的自然更新林。
-
试验毛竹林大小年分明(2014年为小年,2015年为大年,以此类推)。建立6个面积为20 m×20 m的标准样地,样地坡度20°左右,坡向正南,海拔高度169 m。采用每木检测方法分别测定样地内竹株的胸径和竹高。样地内竹林密度为3 875株·hm-2,郁闭度0.95,竹株胸径为4.0~13.6 cm,平均胸径9.8 cm,平均竹高13.2 m。林下几乎无灌木和草本,但地表覆盖一定的凋落物,平均厚度约2.0 cm。每2 a劈山1次,不施肥,不翻耕,仅采挖春笋。
-
于2014年7月下旬(毛竹新竹高生长完成后),在6个标准样地中选择具有代表性、面积为10 m×10 m的地块6个,其中自然生长(对照)样地和截雨干旱样地各3块,且对照和干旱样地成对出现,分别进行样地本底调查。将干旱样地上的稀疏灌木去除,然后用顶棚法模拟截雨。具体方法:在距离地面上方1.5 m高度搭建PVC防水板材的截雨大棚(面积为11 m×11 m)进行截雨,用胶水粘合塑料布与毛竹结合处缝隙;为保证坡度、地形和林分条件与对照样地尽可能一致,大棚一边与等高线平行。在样地周围开挖深约50 cm、宽约20 cm的壕沟,并沿着壕沟埋入50 cm深的白铁皮,壕沟内部同时铺上塑料薄膜,用于阻止水分从旁边渗入,并能更好地排水。顶棚法模拟截雨后,样地内降雨量近乎为0。对照样地仅在周围开挖相似壕沟,不作其它处理。本人工截雨干旱试验目前仍在持续,其中截雨干旱样地内毛竹仍存活,但每年新竹数量相比对照处理有所下降。
-
于自然生长和人工截雨干旱处理试验2 a后,分别于2016年3月(春季)、8月(夏季)、10月(秋季)、12月(冬季)和翌年3月的每月上旬选择晴朗无云的天气,对不同处理(对照和截雨干旱)毛竹叶片比叶质量取样测定1次。具体方法:在每块样地中分别选择长势一致、胸径大小相似的不同年龄毛竹各2株。借助野外临时搭建的梯子,分别各龄毛竹上、中、下3个冠层部位的向阳位置,用镰刀勾取向阳枝条,迅速采摘所有叶片带回实验室,用于比叶质量和养分测定。每个处理(分别竹龄和冠层)各取10~15片叶片测定比叶质量,3次重复,最后取平均值。叶面积采用WinRHIZO(Pro 2007)软件扫描处理获得。取样叶在80℃下烘至恒质量,用电子天平(精确度0.001 g)称其干质量。比叶质量计算:LMA=叶片干质量/叶面积[12]。
-
在模拟截雨干旱和对照样地内分别采集4个土壤剖面作为重复测定土壤含水量,采样深度为0~20 cm,土壤装在铝盒中带回,用电子天平(精确度0.01 g)称取20 g鲜土,在80℃下烘干至恒质量。计算公式为:土壤含水量=(鲜土质量-干土质量)/鲜土质量×100%。该试验中对照和截雨干旱样地内0~20 cm深度土壤含水量存在季节差异,冬季和春季对照样地内土壤含水量最高(平均分别为29.82%和30.15%),其次为夏季(平均为27.12%),秋季(10月份)土壤含水量最低(平均为23.42%);截雨干旱样地土壤含水量季节差异不明显(各季节平均为4.44%~4.92%)。各生长季节对照样地土壤含水量显著高于截雨干旱样地。
-
不同年龄毛竹叶片比叶质量季节和冠层间的差异及其处理间差异,采用Excel 2007和SPSS16.0软件进行数据处理,各参数以平均值加减1个标准误(Mean±SE)表示。
-
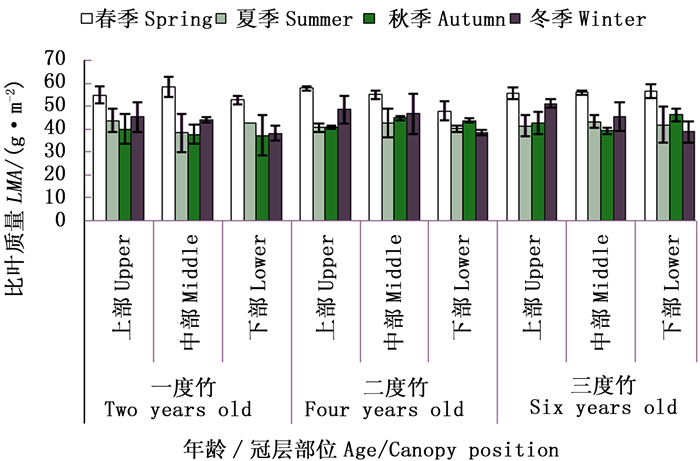
对截雨干旱与对照处理下毛竹各龄竹叶片平均比叶质量随生长季节的动态变化进行分析(图 1),截雨干旱与自然生长下,毛竹个体叶片比叶质量随生长季节更替均呈现出“先降后升”的变化趋势,即比叶质量在春季最高,其次为冬季,而在夏、秋季较低。在自然生长条件下,毛竹叶片比叶质量全年平均为41.08±7.94 g·m-2,四季比叶质量在33.37±2.62~50.08±2.50 g·m-2之间;干旱条件下,毛竹叶片比叶质量全年平均为46.44±6.44 g·m-2,四季比叶质量在41.70±1.14~55.96±2.27 g·m-2之间。可以看出,截雨干旱促使春、夏、秋季毛竹叶片比叶质量增加,尤其以夏季和秋季较为明显。方差分析表明(图 1),毛竹叶片比叶质量季节差异显著(P=0.046 4 < 0.05),但对照与截雨干旱处理之间差异不显著。
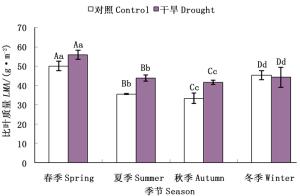
图 1 毛竹个体叶片比叶质量的季节变化
Figure 1. The seasonal change of LMA for different ages of Ph. edulis between control and drought stress
-
对不同竹龄个体叶片比叶质量季节变化的研究表明(图 2),自然生长条件下,不同生长季节毛竹叶片比叶质量均随着竹龄增加而增加。各龄毛竹叶片比叶质量均值的季节变化均表现为春季>冬季>秋季>夏季,其中,各龄竹叶片春季比叶质量均值分别比夏季和秋季高92.74%~130.78%和47.96%~58.73%。方差分析表明(图 2),毛竹同龄个体叶片比叶质量生长季节差异极显著(P < 0.01),相同季节的不同竹龄个体之间差异显著(P=0.026 0 < 0.05)。
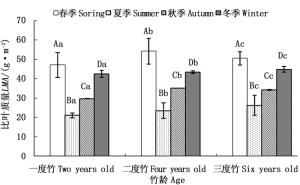
图 2 自然生长下各龄毛竹叶片比叶质量季节变化
Figure 2. Seasonal variation of LMA for different ages of Ph. edulis under natural condition
-
由图 3可以看出,与对照一致,截雨干旱下,各生长季节的毛竹叶片比叶质量均随着竹龄增加而增加。截雨干旱下各竹龄毛竹叶片比叶质量均值随生长季节变化也均表现为春季>冬季>秋季>夏季的趋势。由此可知,截雨干旱与否,毛竹各龄竹叶片比叶质量在春季最高,冬季其次,而夏季最低。方差分析表明(图 3),截雨干旱下,毛竹叶片比叶质量生长季节差异也极显著(P < 0.01),在不同竹龄个体之间差异显著(P < 0.05)。
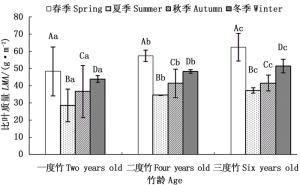
图 3 截雨干旱下各竹龄毛竹叶片比叶质量季节变化
Figure 3. Seasonal variation of LMA of each age of Ph. edulis under drought stress
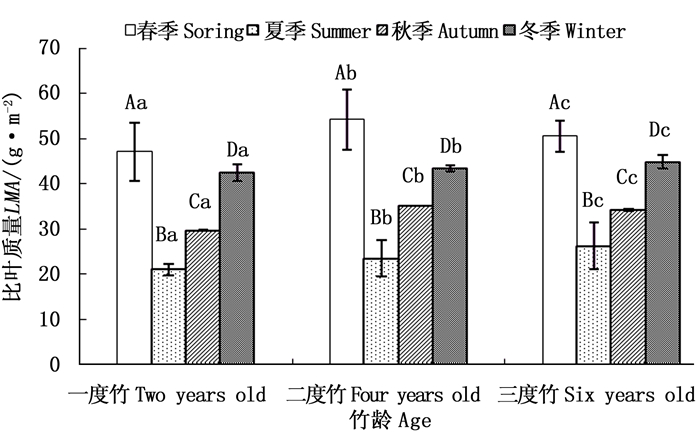
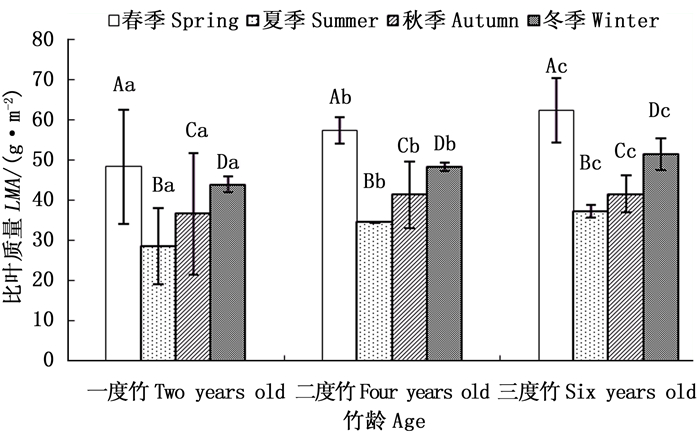
由图 4可以看出,截雨干旱处理下毛竹各竹龄叶片比叶质量在不同生长季节均表现出高于对照的趋势(图 4a、b、c),各龄竹叶片全年平均比叶质量也高于对照(图 4d)。截雨干旱与否,毛竹叶片全年比叶质量均值均随竹龄增加而增加,表现为三度竹>二度竹>一度竹(图 4d)。
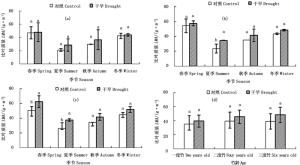
图 4 截雨干旱下毛竹叶片比叶质量季节变化
Figure 4. Seasonal variation of LMA of Ph. edulis under control and drought stress
方差分析表明,一度竹叶片各生长季节比叶质量在对照和截雨干旱之间差异不显著(P>0.05)(图 4a);二度竹叶片比叶质量截雨干旱与对照间差异仅在夏季显著(P < 0.05)(图 4b);三度竹叶片比叶质量在春、夏季截雨干旱与对照间差异均显著(P < 0.05)(图 4c)。可见,毛竹叶片比叶质量截雨干旱与对照间差异显著性受竹龄影响,随着竹龄增加,处理间比叶质量显著差异出现的季节范围增加。
-
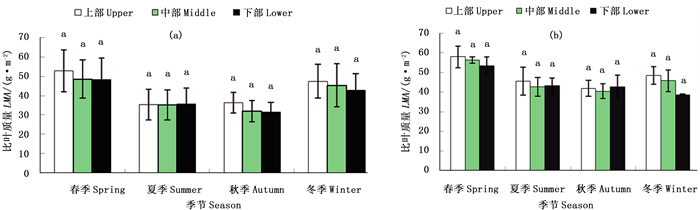
图 5a显示自然生长下不同生长季节毛竹全竹叶片平均比叶质量冠层差异。结果表明,不同生长季节毛竹比叶质量随冠层高度的垂直变化均表现出冠层上部>中部>下部,即各生长季节叶片比叶质量均随着冠层高度增加而增加。

图 5 不同生长季节毛竹叶片比叶质量冠层差异
Figure 5. The canopy difference of LMA for different growth stage of Ph. edulis
与毛竹所有年龄个体叶片比叶质量的季节变化相似,各冠层叶片比叶质量均随着生长季节增加而呈现“先降后升”的趋势。冠层上部叶片,其比叶质量随季节变化表现为春季>冬季>秋季>夏季;中部和下部叶片则为春季>冬季>夏季>秋季。可见,无论竹龄和冠层部位,春季和冬季毛竹个体叶片比叶质量均较高,而在夏季和秋季其比叶质量较低。方差分析表明,自然生长条件下,不同冠层部位之间叶片比叶质量差异不显著。
图 5b显示截雨干旱下不同生长季节毛竹比叶质量冠层差异。结果表明,截雨干旱下,毛竹不同冠层部位比叶质量差异随生长季节变化不同。春季和冬季,比叶质量随冠层变化为冠层上部>中部>下部,夏季和秋季其比叶质量以冠层上部最高,中部和下部差异较小。
相同冠层部位,其比叶质量随季节变化趋势不尽相同。冠层上部和中部叶片比叶质量随季节递增呈现“先降后升”趋势,即春季>冬季>夏季>秋季的大小变化趋势;冠层下部叶片比叶质量随生长季节递增呈现逐渐下降趋势。方差分析表明,截雨干旱下不同冠层之间比叶质量差异也不显著。由图 5a、b可以看出,不同冠层,各生长季节毛竹叶片比叶质量均为干旱处理高于对照。
-
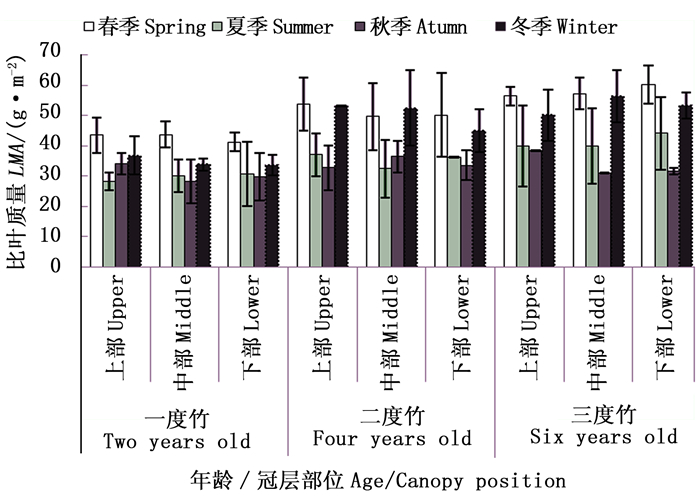
对不同竹龄毛竹叶片比叶质量冠层差异随生长季节的变化研究表明(图 6),自然生长条件下,不同竹龄毛竹各冠层部位叶片比叶质量随季节变化趋势一致,均为“先降后升”,仅季节排序不同。相同生长季节,毛竹各冠层部位叶片比叶质量均随着竹龄增加逐渐上升。
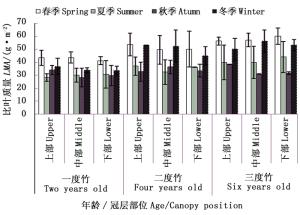
图 6 自然生长下毛竹叶片比叶质量冠层差异随竹龄和生长季节的变化
Figure 6. The change of canopy difference for LMA with seasons and ages of Ph. edulis under control treatment
一度竹春、夏季比叶质量随着冠层增加而增加,秋、冬季节比叶质量随着冠层增加而下降。二度竹春季和冬季比叶质量随着冠层高度增加而下降。三度竹比叶质量冠层差异因季节不同而不同,春季和夏季比叶质量随着冠层高度增加而增加,秋季随着冠层高度增加而下降,冬季随着冠层高度增加先升后降。
-
截雨干旱下,除二度竹、三度竹冠层下部叶片外,不同竹龄毛竹各冠层部位叶片比叶质量随季节变化趋势一致,均为“先降后升”(图 7)。相同冠层部位,不同竹龄之间差异较小。截雨干旱下,毛竹比叶质量冠层差异也受生长季节和竹龄双重影响。春季,一度竹比叶质量冠层中部最高,二度竹为冠层上部>中部>下部,三度竹相反。夏季,二度竹和三度竹比叶质量冠层梯度分布相似,为中部最高,一度竹中部最低。秋季,二度竹和三度竹比叶质量随着冠层增加逐渐增加,一度竹则不断下降。冬季,各龄竹比叶质量均随冠层增加逐渐下降。
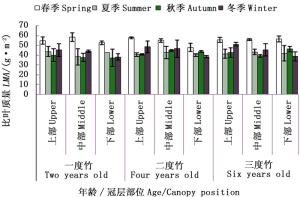
图 7 截雨干旱下毛竹叶片比叶质量冠层差异随竹龄和生长季节的变化
Figure 7. The change of canopy difference for LMA with seasons and ages of Ph. edulis under drought treatment
对于同一冠层毛竹而言,干旱处理下各竹龄毛竹上下冠层比叶质量增幅由大到小均表现为三度竹>一度竹>二度竹。综合分析表明,截雨干旱与否,各龄竹叶片比叶质量在冠层之间差异均不显著,但相同冠层比叶质量季节差异极显著或显著;而干旱缓解了毛竹各冠层比叶质量的季节差异。毛竹冠层高度对叶片比叶质量的影响受竹龄和季节的限制。
-
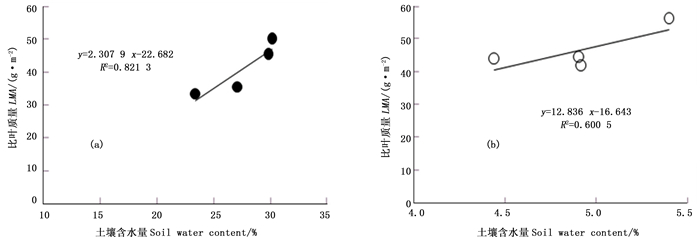
由于截雨干旱处理后毛竹叶片比叶质量适宜性响应变化,且截雨干旱与对照土壤含水量显著差异,对不同处理样地内毛竹叶片比叶质量随土壤含水量的变化分别进行相关性分析。由图 8a可以看出,自然生长条件下,毛竹不同生长季节叶片比叶质量与试验样地内各生长季节土壤含水量之间存在显著的线性相关(R2 = 0.821 3)。说明自然生长下毛竹在不考虑个体年龄对比叶质量的影响情况下,其各生长季节叶片比叶质量会随土壤含水量增加而增加。由图 8b可以看出,截雨干旱下,毛竹叶片比叶质量也与土壤含水量呈显著正相关(R2 = 0.600 5),但相关性低于自然生长条件。

图 8 毛竹叶片比叶质量与土壤含水量的相关性
Figure 8. The correlation between LMA and soil water content for P.edulis
-
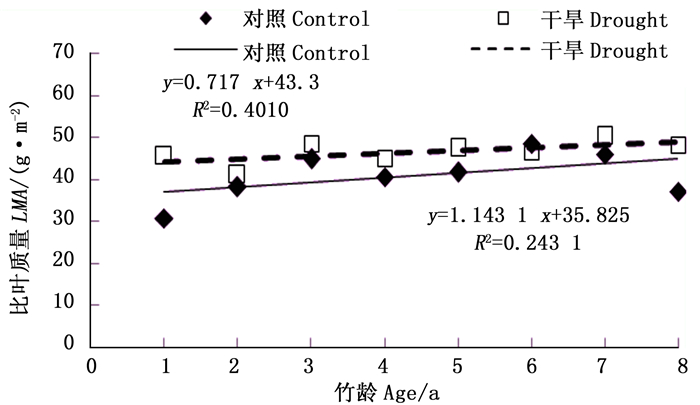
由图 9可以看出,毛竹叶片比叶质量也与竹龄大小呈一定的正相关(对照:R2=0.243 1;截雨干旱:R2=0.401 0),即随着竹龄增加,毛竹叶片比叶质量增加,但截雨干旱处理叶片比叶质量与竹龄的相关性程度高于对照。

图 9 毛竹叶片比叶质量与竹龄的相关性
Figure 9. The correlation between LMA and ages for P.edulis
-
比叶质量不仅存在物种和功能群间的内在差异,同时也随生长环境因子差异发生变化,并存在时间和空间上的变化[5]。为保证植物本身不受外界环境因子的伤害,同时使自身具有更快速生长的可能[13],比叶质量会随环境的变化而变化。因此,植物比叶质量的变化趋势具有重要生态学意义,其变化一定程度反映了植物性能及其对环境的可塑性适应程度。
毛竹作为重要固碳树种,生长迅速,每年生物量积累巨大。其生长独特,从幼笋出土至长成幼竹只需短短几个月的时间即完成秆行生长[14]。因此,研究毛竹比叶质量时空变化特性及其对环境变化的响应对于揭示毛竹生态适应性及生产力提升机理具有重要科学意义。大部分陆地物种的比叶质量介于30~330 g·m-2之间,生长缓慢的物种比快速生长的物种具有更高的比叶质量[5]。本研究发现,截雨干旱与否,毛竹比叶质量处于大部分物种比叶质量分布范围区间的较低端,这也充分说明毛竹生长迅速。
与前人比叶质量随年龄增加而增加的研究结论一致[5, 15],本研究发现,截雨干旱与否,毛竹叶片比叶质量在不同生长季节均随竹龄增加而增加。就比叶质量随生长季节变化而言,其各龄竹随生长季节的变化趋势均为春季>冬季>秋季>夏季,且截雨干旱与否,不同竹龄毛竹比叶质量存在显著季节差异。说明截雨干旱与否,在夏季、秋季毛竹鞭根系统旺盛生长季节,毛竹通过拥有较低的比叶质量来获取更多资源满足生长需求,与作者研究结论较为一致,即生长速率与比叶质量显著相关[16]。而春季和冬季,毛竹通过增加比叶质量来增强对春旱和冬季低温的抵御能力。因此,生产实践中可通过简易测定叶片比叶质量来判断毛竹生长状况。这也说明,通过测定不同生长季节毛竹比叶质量大小可用来判断该季节毛竹生长情况,进而可为优化生长调控措施提供直接依据。
-
比叶质量可用来反映植物对资源的综合利用情况[17-19]。造成比叶质量差异的因素有很多,除了植物内在变化影响比叶质量外[12],环境条件也产生重要影响。Shipley对30种草本植物的研究中发现,尽管比叶质量大部分变异是由物种(70%)引起,但同一个体叶片中存在相当大的比叶质量变异(26%)[20]。植物个体内部的差异较大,如红杉树(Sequoia sempeervirens (Lamb.) Endl.)比叶质量随树冠高度增加而增加[21],因为光照梯度对确定比叶质量起着至关重要的作用。与荫蔽环境下叶片相比,处于较高光照条件下的叶片具有较大的比叶质量[17, 22]。何春霞等通过测定树冠内外部及4个方位上的比叶质量随树冠开度的变化以及方位差异[18],研究显示叶片比叶质量随树冠开度增加而增大;叶片比叶质量的方位变化则是南向>西向>北向>东向,与叶片所接受到的光强变化规律一致。表明树冠外围和南向、西向的叶片由于接受到的光能较多、温度高、相对湿度小等,其叶片会增大单位面积的质量,从而减少对光能的吸收,是叶片对周围小气候的形态适应。董伊晨等通过分析不同苗龄红松(Pinus koraiensi Sieb. et Zucc.)幼苗在控制温度和光照条件下比叶质量等叶性状的变化差异,发现幼苗比叶质量受到温度、光照和苗龄的影响,比叶质量与温度和苗龄呈正相关性[15]。
与前人研究结果一致,本研究发现,自然生长条件下,各生长季节毛竹叶片比叶质量均随着冠层高度增加而增加,冠层差异较显著。截雨干旱下,毛竹不同冠层部位叶片比叶质量差异不显著,基本以冠层上部最高;其中,春季和冬季,毛竹叶片比叶质量随冠层变化表现为冠层上部>中部>下部,夏季和秋季其比叶质量以冠层上部最高,中部和下部差异较小。Sterck等将田间数据与基于过程的植物模型相结合,研究表明比叶质量是高光照条件下植物生长的种间变异的最佳预测因子[23]。因此,本研究对毛竹叶片比叶质量随冠层梯度分布的特性研究可用于指导不同生长光环境或不同经营密度下毛竹个体生长差异的预测因子。
-
研究认为,除了光照影响比叶质量外,其它因素与冠层光照梯度共同作用影响,最显著的是水分亏缺(干燥空气)。Koch等研究认为,较高树木的水分利用率降低是比叶质量冠层差异的主要因素[21];Wright等研究表明比叶质量随着降雨量的减少和光照辐射的增加而增大[6]。这是由于在低水分条件下生长发育的叶片其膨胀率较低,叶片细胞更小,排列更紧密,叶片细胞壁也增厚,共同导致叶片密度增加[6]。高比叶质量的植物叶片的细胞壁组分和碳含量较高,其叶片水分含量、氮含量较低。
植物比叶质量与叶片密度和叶片厚度两者大小密切相关[5, 24]。往往水分含量较低环境下植物具有较高的比叶质量和较大叶片厚度[24]。与前人研究结论一致[5-6],本研究发现,截雨干旱处理下,随着干旱胁迫增加,毛竹叶片比叶质量比对照(自然生长下)显著增加。可能是低水分条件下生长发育的毛竹叶片其膨胀率较低,叶片细胞排列更紧密,叶片细胞壁也增厚,共同导致叶片密度增加,此原因影响机制需要今后进一步探讨。
具有低比叶质量的物种倾向于快速的资源获取、快速代谢和增长。本研究中毛竹比叶质量较低,处于多数物种比叶质量范围下限,说明毛竹属于低比叶质量物种,在较好资源环境条件下具有较强的适应性和生长优势。截雨干旱促使毛竹叶片比叶质量增加,这是毛竹应对截雨干旱作出的典型形态学反应,反映了毛竹在截雨干旱缺水的不利生长环境下,可通过增加比叶质量来增强干旱防御能力和生存持久能力。前期研究发现,无论生长季节,不同林龄毛竹个体叶片比叶质量随冠层高度垂直变化趋势基本与叶片净光合速率日均值的冠层梯度变化趋势较为一致[11]。本研究表明截雨干旱处理毛竹各冠层比叶质量均值高于对照处理,截雨干旱增加毛竹叶片比叶质量。干旱胁迫是否通过影响比叶质量对毛竹光合能力产生影响有待进一步研究。
-
比叶质量是植物重要生长策略指标,其时空变化一度程度反映了植物性能及其对环境的可塑性适应程度。毛竹属于低比叶质量物种,因此在较好资源环境条件下,具有较强的适应性和生长优势。毛竹比叶质量存在显著季节差异和年龄差异。毛竹比叶质量随冠层垂直梯度分布,但冠层差异不明显,且受生长季节变化影响。毛竹比叶质量冠层差异较小而季节差异较显著,说明与光照相比,土壤水分条件对毛竹比叶质量产生重要影响。截雨干旱促使毛竹叶片比叶质量增加,这是毛竹应对截雨干旱作出的典型形态学反应,反映了毛竹在截雨干旱缺水的不利生长环境下,可通过增加比叶质量来增强干旱防御能力。但其影响机制需要今后进一步探讨。
毛竹比叶质量时空变化及对截雨干旱的响应
Seasonal and Canopy Variation of Leaf Mass Per Area for Phyllostachys edulis Leaves and its Response to Drought Stress
-
摘要:
目的 定量分析毛竹叶片比叶质量随生长季节、冠层部位和竹龄的变化及其对截雨干旱的响应,为全球气候变化下毛竹人工林可持续高效经营提供理论依据。 方法 以毛竹为研究对象,通过人工截雨干旱试验,对毛竹叶片比叶质量时空特性进行测定分析。 结果 (1)截雨干旱与自然生长下,毛竹叶片比叶质量存在显著季节差异和年龄差异;不同生长季节,毛竹叶片比叶质量均随竹龄增加而增加;各龄竹比叶质量随生长季节的变化趋势均为春季>冬季>秋季>夏季。(2)截雨干旱与自然生长下,各龄竹比叶质量冠层差异不显著;自然生长下,比叶质量随冠层高度的垂直变化在不同生长季节均为冠层上部>冠层中部>冠层下部;截雨干旱下,比叶质量随冠层高度的变化因生长季节变化而不同。(3)截雨干旱下,各龄竹不同季节的比叶质量均高于自然生长下的比叶质量。干旱缓解了各冠层比叶质量的季节差异。截雨干旱与自然生长间比叶质量差异显著性受竹龄影响。(4)不同处理下各季节毛竹叶片比叶质量与土壤含水量、竹龄呈线性正相关。 结论 毛竹属于低比叶质量物种,生长季节显著影响其比叶质量变化;与光照相比,土壤水分条件对毛竹比叶质量产生重要影响。 Abstract:Objective To quantitatively analyze the seasonal change of leaf mass per area (LMA) along canopy height for different aged moso bamboo (Phyllostachys edulis) in natural and drought stress growth environment, and to clarify the response characteristics of LMA to the throughfall exclusion for drought stress, aiming at providing references for sustainable and efficient management of Ph. edulis plantations under the global climate background. Method The spatial and temporal dynamic change of LMA is detected for moso bamboo under two different treatments with rain shelter or not. Result (1) Regardless of throughfall exclusion or not, the LMA increased with the age of bamboo during different growing seasons, and the seasonal change trend from maximum to minimum was spring, winter, autumn and then summer in turn. There were significant differences of LMA among different seasons and treatments for the same age of bamboo (P < 0.05), regardless of throughfall exclusion or not. (2) Regardless of throughfall exclusion or not, the differences of LMA were not significant among canopy for various ages of bamboo, while it was significant among seasons within the same canopy. In natural growth condition, the LMA of each season increased along with the canopy height and its vertical change showed the trend of the upper canopy > middle canopy > lower canopy successively, and the value of each canopy decreased at first and then increased as the growing season. There was significant difference among different seasons (P < 0.01) for LMA of different ages of bamboo, while there was no significant difference among different canopy layers under throughfall exclusion. (3) Under the drought stress, the annual average value and each individual value of the LMA were all higher than the control in every season. The drought stress relieved seasonal difference of LMA in various canopy. The seasonal change of the LMA within the same canopy was also affected by age of bamboo. (4) With or without drought stress, there was linear positive correlation between the LMA and soil water content in each season, but the correlation under drought stress was lower than that of the control. The positive correlation between LMA and ages of bamboo under drought stress was higher than that of the control. Conclusion It is concluded that moso bamboo belongs to the species with lower LMA, and the growth season has significant effect on the change of LMA. Compared with the light, the soil moisture condition has more important influence on the LMA of the moso bamboo. -
Key words:
- Phyllostachys edulis
- / drought stress
- / leaf mass per area
- / season
- / age
- / canopy height
-
图 1 毛竹个体叶片比叶质量的季节变化
Figure 1. The seasonal change of LMA for different ages of Ph. edulis between control and drought stress
图 2 自然生长下各龄毛竹叶片比叶质量季节变化
Figure 2. Seasonal variation of LMA for different ages of Ph. edulis under natural condition
图 3 截雨干旱下各竹龄毛竹叶片比叶质量季节变化
Figure 3. Seasonal variation of LMA of each age of Ph. edulis under drought stress
图 4 截雨干旱下毛竹叶片比叶质量季节变化
Figure 4. Seasonal variation of LMA of Ph. edulis under control and drought stress
图 5 不同生长季节毛竹叶片比叶质量冠层差异
Figure 5. The canopy difference of LMA for different growth stage of Ph. edulis
图 6 自然生长下毛竹叶片比叶质量冠层差异随竹龄和生长季节的变化
Figure 6. The change of canopy difference for LMA with seasons and ages of Ph. edulis under control treatment
图 7 截雨干旱下毛竹叶片比叶质量冠层差异随竹龄和生长季节的变化
Figure 7. The change of canopy difference for LMA with seasons and ages of Ph. edulis under drought treatment
图 8 毛竹叶片比叶质量与土壤含水量的相关性
Figure 8. The correlation between LMA and soil water content for P.edulis
-
[1] Gutschick V P, Wiegel F W. Optimizing the canopy photosynthetic rate by patterns of investment in specific leaf mass[J]. The American Naturalist, 1988, 132(1):67-86. doi: 10.1086/284838 [2] 薛立, 张柔, 奚如春, 等.华南地区6种阔叶幼苗叶片形态特征的季节变化[J].生态学报, 2012, 32(1):123-134 [3] Lambers H, Poorter H. Inherent variation in growth rate between higher plants:a search for physiological causes and ecological consequences[J]. Advances in Ecological Research, 1992, 22(1):187-261. [4] Westoby M, Falster D S, Moles A T, et al. Plant ecological strategies:some leading dimensions of variation between species[J]. Annual Review of Ecology & Systematics, 2002, 33(1):125-159. [5] Poorter H, Niinemets U, Poorter L, et al. Causes and consequences of variation in leaf mass per area (LMA):a meta-analysis[J]. New Phytologist, 2009, 182(3):565-588. doi: 10.1111/j.1469-8137.2009.02830.x [6] Wright I J, Reich P B, Westoby M, et al. The worldwide leaf economics spectrum[J]. Nature, 2004, 428(8985):821-827. [7] 唐晓鹿, 范少辉, 漆良华, 等.采伐对幕布山区毛竹林土壤呼吸的影响[J].林业科学研究, 2013, 26(1):52-57 doi: 10.3969/j.issn.1001-1498.2013.01.009 [8] Wen G S, Zhang L Y, Zhang R M, et al. Temporal and spatial dynamics of carbon fixation by moso bamboo(Phyllostachys pubescens) in subtropical China[J]. Botanical Review, 2011, 77(3):271-277. doi: 10.1007/s12229-011-9068-x [9] 应叶青, 郭璟, 魏建芬, 等.干旱胁迫对毛竹幼苗生理特性的影响[J].生态学杂志, 2011, 30(2):262-266. [10] 袁佳丽, 温国胜, 张明如, 等.毛竹快速生长期的水势变化特征[J].浙江农林大学学报, 2015, 32(5):722-728. [11] 曹永慧, 周本智, 王小明, 等.冠层高度对毛竹叶片光合生理特性的影响[J].西北植物学报, 2016, 36(11):2256-2266 doi: 10.7606/j.issn.1000-4025.2016.11.2256 [12] 郑淑霞, 上官周平.不同功能型植物光合特性及其与叶氮含量、比叶质量的关系[J].生态学报, 2007, 27(1):171-181 doi: 10.3321/j.issn:1000-0933.2007.01.020 [13] Wright I J, Wearoby M. Cross-species relationship between seedling relative growth rate, nitrogen productivity and root vs. leaf function in 28 Australian woody species[J]. Functional Ecology, 2000, 14(1):97-107. doi: 10.1046/j.1365-2435.2000.00393.x [14] Isagi Y. Ecological study of bamboo communities:a resource-based approach[J]. Bull Fac Int Arts Sci Hiroshima Univ, 1995, 4(21), 235-238(in Japanese). [15] 董伊晨, 刘艳红.红松不同苗龄幼苗叶性状对温度和光照变化的响应[J].生态学报, 2017, 37(17):5662-5672 [16] Poorter L. Leaf traits show different relationships with shade tolerance in moist versus dry tropical forests[J]. New Phytologist, 2009, 181(4):890-900. doi: 10.1111/j.1469-8137.2008.02715.x [17] 冯玉龙, 曹坤芳, 冯志立, 等.四种热带雨林树种幼苗比叶质量, 光合特性和暗呼吸对生长光环境的适应[J].生态学报, 2002, 22(6):901-910 doi: 10.3321/j.issn:1000-0933.2002.06.015 [18] 何春霞, 李吉跃, 张燕香, 等.5种绿化树种叶片比叶重、光合色素含量和δ13C的开度与方位差异[J].植物生态学报, 2010, 34(2):134-143 doi: 10.3773/j.issn.1005-264x.2010.02.004 [19] 王子奇, 查天山, 贾昕, 等.油蒿光合参数季节动态及其与叶氮含量和比叶面积的关系[J].生态学杂志, 2017, 36(4):916-924. [20] Shipley B. Structured interspecific determinants of specific leaf area in 34 species of herbaceous angiosperms[J]. Functional Ecology, 1995, 9(2):312-319. doi: 10.2307/2390579 [21] Koch G W, Sillett S C, Jennings G M, et al. The limits to tree height[J]. Nature, 2004, 428(6985):851-854. doi: 10.1038/nature02417 [22] Cao K F. Leaf anatomy and chlorophyll content of 12 woody species in contrasting light conditions in a Bornean heath forest[J]. Canadian Journal of Botany, 2000, 78(10):1245-1253. doi: 10.1139/b00-096 [23] Sterck F J, Poorter L, Schieving F. Leaf traits determine the growth-survival trade-off across rain forest tree species[J]. American Naturalist, 2006, 167(5):758-765. doi: 10.1086/503056 [24] Puglielli G, Crescente M F, Frattaroli A R, et al. Leaf mass per area (LMA) as a possible predictor of adaptive strategies in two species of Sesleria (Poaceae):analysis of morphological, anatomical and physiological leaf traits[J]. Annales Botanici Fennici, 2015, 52(1-2):135-143. doi: 10.5735/085.052.0201 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4250
- HTML全文浏览量: 2025
- PDF下载量: 61
- 被引次数: 0
