

-
根系呼吸是植物生命活动所需能量和物质的来源,对植物养分吸收、根系更新和植株生长具有重要意义[1-2]。植物根系呼吸是地下部分物质代谢的中心,因此,根系呼吸作用对植物体内的物质转化与合成具有直接影响,而在呼吸代谢过程中起重要作用的是呼吸代谢关键酶。环境因子可直接影响部分关键酶活性,对组织中新陈代谢具有间接影响[3-4]。
林木根系呼吸与其根系氮素浓度密切相关[5]。研究发现,根呼吸速率通常随着根系组织氮浓度的升高呈增加趋势[6],而引起根系组织氮浓度增加的主要外部因素是土壤氮浓度的增加[7]。当土壤中氮素获得量增加时,根系组织中氮浓度也随之提高,并且根系中相关维持蛋白以及结构蛋白的支出升高[8],从而使整个根系呼吸速率提高。Hawthorne等[9]通过研究哥伦比亚贝尔河营针叶林树种根系呼吸时发现,增施氮肥可以促进根系呼吸;Sorrenti等[10]采用沟槽法对桃(Amygdalus persica L.)根系呼吸进行了测定,发现根际土壤氮素浓度提高有利于桃的根系呼吸。Dyckmans等[11]通过研究欧洲山毛榉(Fagus sylvatica L.)林地土壤碳和氮养分循环对根系呼吸的影响,得出氮源的改变能够显著影响根系呼吸强度。Ceccon等[12]将欧洲山杨(Populus tremula L. Sp. Pl.)置于盆栽试验和田间试验条件下,发现根系呼吸与氮素浓度呈正相关。Kewald等[13]研究认为,土壤中增施氮肥对杨树(Populus Linn.)细根呼吸速率有显著影响,并且细根呼吸速率显著高于粗根呼吸速率。国内有关氮素浓度对根系呼吸影响的研究主要以水曲柳(Fraxinus mandschurica Rupr.)[14]、红桦(Betula albosinensis Burk.)[15]、杨树[16]、闽楠(Phoebe bournei(hemsl.)Yang.)[17]等用材林树种为主,而对经济林树种的研究鲜有报道。无论是经济树种还是用材树种,地上果实、地下块茎或块根、地上生物量、树干干材收获量等收获目标与经营管理措施的关系都极为密切,研究氮素对林木根系呼吸的影响规律,对于林木养分的科学管理都具有重要意义。
本研究以新疆南疆盆地核桃主栽品种‘新温185’(Juglans regia ‘Xinwen185’)为供试材料,通过设置田间施肥试验,运用离体根法测定根系呼吸,分析核桃在生长发育和组织更新代谢过程中施氮水平对细根呼吸速率及相关酶活性的影响,旨在进一步探讨氮素对核桃根系呼吸的调控机理,以期为核桃的高效施肥管理提供理论依据。
-
本试验于2017年3—9月在新疆阿克苏地区乌什县阿克托海乡阿特房子村核桃生产园(79°12′12.76″~79°13′57.87″ E,41°11′06.31″~41°12′47.74″ N;海拔1 394 m)进行。阿克托海乡位于天山南部、塔里木盆地的西北边缘处,气候带类型属于暖温带大陆性干旱气候,年平均气温为9.4℃,≥10℃的积温3 200~3 600℃,年平均降水量为91.5 mm,年均蒸发量达2 003.8 mm,年日照时数为2 750~2 850 h,无霜期250~286 d。
-
试验所用林地面积为0.3 hm2,主栽的核桃品种是‘新温185’,栽植方向为东西行向,株行距为5 m×6 m,选取树龄13 a、树体状况良好的核桃树。土壤类型为灌淤土,土层深厚,土壤有机质含量10.70 g·kg-1,全氮(N)含量2.62 g·kg-1,全磷(P)含量1.24 g·kg-1,全钾(K)含量8.79 g·kg-1,碱解N含量77.69 mg·kg-1,有效P含量14.34 mg·kg-1,速效K含量77.31 mg·kg-1。
-
在常规P肥和K肥施用量基础上设置4个施N水平,每个施氮水平重复3次,每次重复选取3株大小、长势一致的样株,在供试样园内选择面积一致、树木分布均匀的施肥样地。4个施氮水平(纯量)如下:N0(0 kg·株-1)、N1(0.75 kg·株-1)、N2(1.50 kg·株-1)和N3(2.25 kg·株-1)。氮肥用含N46%的尿素(CO(NH2)2),田间施肥于2017年3月进行,在核桃行间树冠2/3处挖50 cm深的环状沟,每一施氮水平中所有肥料一次性施入。
-
采用离体根法测定根系Q。分别在核桃雌花盛花期后的0、70、115、160、205、250 d,选择晴朗天气,在每个施氮水平样株同一方向上挖掘核桃根系,挖掘至60 cm土壤深度。根系挖掘在上午10:00—12:00(北京时间)进行,并在2 h内完成,每个施氮水平每次挖取3株。将挖出的根系先用细毛刷将其表面土壤刷去,使用游标卡尺测量选取直径<1 mm的根系[18-19],用于细根呼吸速率的测定。
为了测定Q,将直径<1 mm的根系迅速称取0.1 g,切成2 mm的小段,放入Oxytherm液相氧电极测定系统(英国Hansatech公司生产,仪器型号为Chlorolab-2)反应杯中,加盖并启动程序,开启电磁搅拌器和恒温水浴,放置3~4 min,当温度达到平衡后,用黑布遮住反应杯,开启记录仪,调好走纸速度,待信号稳定后,落下记录笔开始记录。每个样品测定3~5 min,重复测定3次。
Q(μmol·g-1·h-1)的计算公式如下:
$ Q=\frac{a \times n \times 1000}{A \times t \times 60} $
式中:a为记录纸每小格代表的氧量(μmol);n为测定呼吸速率时记录笔向左走的小格数;A为根系鲜质量(g);t为测定时间(min)。
-
参照Bouma等[20]、毛志泉等[21]和高相彬等[22]方法测定根系呼吸速率。糖酵解途径(EMP)、三羧酸循环途径(TCA)和磷酸戊糖途径(PPP)分别用NaF(10 mmol·L-1)、丙二酸(50 mmol·L-1)和Na3PO4(10 mmol·L-1)3种抑制剂进行抑制(每种抑制剂用0.05 mol·L-1的磷酸缓冲液(pH=6.5)配制),然后用液相Oxy-Lab氧电极测定,每个样品测定5~10 min。测定时使用超级恒温器提供循环水,反应杯中的温度应保持在30℃,对照为不加抑制剂处理的样品。
-
参照Singla等[23]、Ling等[24]的方法测定。称取直径<1 mm的根系0.5 g,剪碎后加入2.5 mL的Tris-HCl(浓度0.1 mol·L-1,pH=7.5)缓冲液,冰浴研磨后,吸取至2 mL离心管中,4℃下以15 000 r·min-1离心15 min,取上清液备用(4℃保存)。取反应液2.9 mL加入酶提取液0.1 mL,再加入ATP溶液12 μL,立即放入340 nm处测初始吸光度,记录吸光度A1,取出比色皿放在25℃环境中反应10 min后,再放入340 nm下测吸光度,记录吸光度A2,对照为不加ATP的样液。
PFK活性(μ·g-1)计算公式如下:
$ {\rm PFK}活性=\frac{ΔA×V_{反总}×10^9×V_{样总}×t}{{ε×d×W×V_{样}}} $
式中:△A=A1-A2;V反总为反应体系总体积(mL);V样总为提取液总体积(mL);t为反应时间(min);ε为NADPH消光系数;d为比色皿光径(cm);W为根系鲜质量(g);V样为加样体积(mL)。
-
参照欧阳光察等[25]的方法测定。称取直径<1 mm的根系0.5 g,剪碎后加入1.5 mL Tris-HCl(浓度0.1 mmol·L-1,pH=7.5)缓冲液研磨,4℃下15 000 r·min-1离心15 min,取上清液备用(4℃保存)。取反应液2.7 mL,加入0.3 mL酶提取液,然后加入15 μL NADH,立即在340 nm下测初始吸光度,记录吸光度值A1,1 min后再测一次吸光度,记录吸光度值A2。
MDH活性(μ·g-1)计算公式如下:
$ {\rm MDH}活性=\frac{△A}{0.01×t×W} $
式中:△A=A1-A2;t为反应时间(min);W为根系鲜质量(g)。
-
按照薛应龙[26]和Brown等[27]的方法。称取直径<1 mm的根系0.5 g,剪碎后加入1 mLTris-HCl(浓度0.1 mg·L-1,pH=8.5)缓冲液研磨,4℃下15 000 r·min-1离心15 min,取上清液备用(4℃保存)。将反应液与酶提取液分别放置于30℃下保温,测量时取2.9 mL反应液和0.1 mL酶提取液混匀后(以2.9 mL反应液和0.1 mL提取缓冲液作为对照),立即在340 nm下测初始吸光度,记录吸光度值A1,5 min后再次测吸光度,记录吸光度值A2。
G-6-PDH活性(μ·g-1)计算公式如下:
$ {\rm G-6-PDH}活性=\frac{△A×V_{反总}}{ε×d×10×t^{-1}} $
式中:△A=A1-A2;t为反应时间(min);ε为NADPH消光系数;d为比色皿光径(cm); V反总为反应体系总体积(mL)。
-
采用二因素多响应变量方差分析检验花后时间、施氮水平以及二者的交互作用对核桃细根呼吸速率及其代谢相关酶活性是否存在影响。采用最小显著差异法(LSD)进行多重比较。采用Pearson相关计算响应变量之间的相关系数,采用双侧方法检验响应变量之间的相关性。显著性水平设定为α=0.05。
采用SSPS 21.0统计分析软件进行数据分析;绘图使用Microsoft Excel 2010软件。
-
二因素多响应变量方差分析结果(表 1)显示:除花后时间与施氮水平交互作用对MDH和PFK活性无显著影响(P>0.05)外,花后时间、施氮水平和二者的交互作用对TCA、EMP、PPP的呼吸速率和MDH、G-6-PDH、PFK活性以及细根Q均影响显著(P<0.05)。根据平方和大小可知:TCA、EMP、PPP的呼吸速率和MDH、G-6-PDH、PFK活性以及细根Q受花后时间影响较大。
表 1 花后时间和施氮水平对核桃细根呼吸速率及相关酶活性影响的二因素多响应变量方差分析
Table 1. Variance analysis of two multivariate responses to the effect of postoperative time and nitrogen application levels on root respiration rate and related enzyme activity
观测指标
Observation index变异来源
Source of variation平方和
SS自由度df 均方
MSF值
F valueP值
P valueTCA呼吸速率
Respiratory rate of TCA花后时间(T)Post flower time 0.002 4 0.001 113.225 <0.05 施氮水平(N)Nitrogen application levels 0.000 3 0.000 16.451 <0.05 花后时间(T)×施氮水平(N)Post flower time ×Napplication levels (T×N) 0.000 12 0.000 2.738 <0.05 EMP呼吸速率
Respiration rate of EMP花后时间(T)Post flower time 0.002 4 0.000 113.231 <0.05 施氮水平(N)Nitrogen application levels 0.000 3 0.000 16.362 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 0.000 12 0.000 2.672 <0.05 PPP呼吸速率
respiration rate of PPP花后时间(T)Post flower time 0.004 4 0.001 172.775 <0.05 施氮水平(N)Nitrogen application levels 0.000 3 0.000 12.062 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 0.000 12 0.000 4.005 <0.05 Q
Basal respiration rate花后时间(T)Post flower time 0.014 4 0.004 113.314 <0.05 施氮水平(N)Nitrogen application levels 0.002 3 0.001 16.400 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 0.001 12 0.000 2.664 <0.05 PFK活性
Activity ofPFK花后时间(T)Post flower time 3 258.907 4 814.727 135.915 <0.05 施氮水平(N)Nitrogen application levels 304.964 3 101.655 16.958 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 142.325 12 11.860 1.979 >0.05 MDH活性
Activity ofMDH花后时间(T)Post flower time 120.167 4 30.042 131.223 <0.05 施氮水平(N)Nitrogen application levels 7.444 3 2.481 10.839 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 4.322 12 0.360 1.573 >0.05 G-6-PDH活性
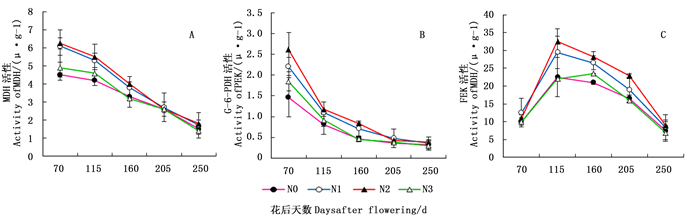
Activity of G-6-PDH花后时间(T)Post flower time 23.056 4 5.764 153.337 <0.05 施氮水平(N)Nitrogen application levels 1.484 3 0.495 13.195 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 1.295 12 0.108 2.870 <0.05 图 1A表明:不同施氮水平下,核桃细根Q在花后70~250 d总体呈下降趋势(图 1A),且根系TCA、EMP、PPP的呼吸速率分别占Q呼吸速率的40.23%、33.02%、20.47%。花后第70~160天,N0与N3水平下的细根Q均低于N2水平下细根Q,且N1水平下的细根Q介于N2与N3水平之间。
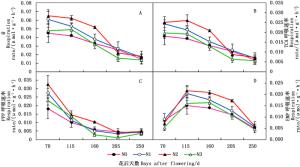
图 1 施氮水平对核桃细根呼吸速率的影响
Figure 1. The effect of nitrogen application levels on respiration rate of walnut fine roots
图 1B表明:在N2水平下,核桃细根TCA的呼吸速率呈先升后降趋势,即在花后70~115 d升高,之后开始下降;其它氮水平下TCA的呼吸速率呈下降趋势;花后70~160 d,N0与N3水平下TCA的呼吸速率均低于N2水平的,而N1水平下TCA的呼吸速率介于N2与N0水平间,其变化趋势与Q呼吸速率的变化趋势相似。
图 1C表明:核桃细根PPP的呼吸速率在花后70~205 d随花后天数的增加呈下降趋势,其中,N2水平下PPP的呼吸速率均高于其它施氮水平;205 d后PPP的呼吸速率缓慢回升。
图 1D表明:不同施氮水平下核桃细根EMP的呼吸速率均呈先升后降趋势。EMP的呼吸速率在花后70~115 d迅速升高,之后逐渐降低。花后115~250 d,N2水平下EMP的呼吸速率均高于其它施氮水平,且N1水平下EMP的呼吸速率介于N2与N3水平之间。
-
图 2A表明:在核桃生长发育过程中,不同施氮水平下核桃细根MDH活性总体呈下降趋势;花后70~160 d,N2与N1水平下MDH活性均高于N3和N0水平。图 2B表明:不同施氮水平下,核桃细根G-6-PDH活性随花后时间的增加总体呈下降趋势;花后70~160 d,N2与N1水平下G-6-PDH活性高于N3与N0水平。图 2C表明:不同施氮水平下,核桃细根PFK活性随花后时间的增加总体均呈先升后降的趋势,其中,N2水平下,PFK活性在花后115 d达到峰值;花后115~250 d,N2与N1水平下PFK活性均高于N3和N0水平。

图 2 施氮水平对核桃细根系呼吸速率及相关酶活性的影响
Figure 2. The effects of nitrogen application levels on respiration rate and related enzyme activities of walnut fine roots
-
核桃细根Q和TCA、EMP、PPP的呼吸速率与相关酶活性的相关性分析结果(表 2)显示:细根Q与G-6-PDH活性和MDH活性呈极显著正相关(P<0.01),但与PFK活性的相关性不显著(P>0.05);TCA的呼吸速率与G-6-PDH、MDH活性呈极显著正相关(P<0.01),但与PFK活性不显著相关(P>0.05);EMP的呼吸速率与PFK活性呈极显著正相关(P<0.01),与MDH活性不显著正相关(P>0.05),与G-6-PDH活性呈不显著负相关(P>0.05);PPP的呼吸速率与MDH、G-6-PDH活性呈极显著正相关(P<0.01),和PFK活性呈不显著负相关(P>0.05)。
表 2 核桃细根Q和TCA、EMP、PPP的呼吸速率与酶活性的相关系数
Table 2. Correlation of respiration rate and enzyme activity between Q and TCA, EMP and PPP of walnut fine root
测定指标
Observation indexMDH活性
Activity of MDHG-6-PDH活性
Activity of G-6-PDHPFK活性
Activity of PFKQ
Basal respiration rate0.974** 0.834** 0.414 TCA呼吸速率
Respiratory rate of TCA0.970** 0.821** 0.430 EMP呼吸速率
Respiratory rate of EMP0.358 -0.096 0.990** PPP呼吸速率
Respiratory rate of PPP0.851** 0.986** -0.141 注:**在0.01水平上极显著相关(双尾),*在0.05水平上显著相关(双尾)。
Notes:** Correlation is highly significant at the 0.01 level(double tail); * Correlation is significant at the 0.05 level(double tail). -
本研究结果表明,在Q中,核桃细根TCA的呼吸速率占比最高,PPP的呼吸速率占比最小。这是因为TCA途径是为植物生活提供所需能量的最有效途径,由于其产生的ATP效率极高,其速率下降致使ATP生成量下降,从而无法满足植物根系正常的生理生化需要[22, 28]。这也说明根系对氮营养的适应,需通过促进TCA呼吸途径才能有效地吸收氮素并为其生长提供重要的营养物质[29]。
较低的氮素浓度有利于提高根系的呼吸速率,氮素浓度过高会抑制根系的呼吸速率,这与桤木(Alnus cremastogyne Burk.)[30]、杨树[12]上的研究结果类似。当土壤中氮含量供给过高时,植物将降低向根系分配碳量,并且降低根系呼吸强度,尤其是细根的生长呼吸强度[31]。由于细根是植物典型的吸收根[32],这类根系细胞生理代谢旺盛[33],与粗根相比具有最大的呼吸速率,其呼吸对根系组织氮浓度的增加或降低极为敏感。因此,当土壤中的氮含量低于植物需求量时,植物为了保证自身的生长发育而获得足够的氮,通过提高向根系分配碳的方式进而促进根系生长[34],最终提高植物氮素吸收的能力,同时也提高了根系的呼吸速率;但是氮素的过量施用也会抑制根系对磷、钾等元素的吸收和利用[35],从而降低根系呼吸速率。
与对照相比,不同施氮水平均使核桃细根呼速率增加。这种作用与相关呼吸酶活性受到促进有关,因为PFK、G-6-PDH和MDH分别是调控EMP、PPP和TCA三种呼吸途径的关键酶,其活性的高低直接影响呼吸速率的强弱[36-37]。随着核桃生长发育的变化,其活性的变化趋势与相应呼吸途径的呼吸速率时节变化趋势相同。本研究结果与樱桃(Cerasus pseudocerasus(Lindl.) G.Don)[38]、平邑甜茶(Malus hupehensis Rehd.)[22]的研究结果相似。本试验是在大田条件基础上开展的,只是研究了施氮水平对核桃细根呼吸速率的影响,在后续的研究中将会考虑环境中多重因素对细根呼吸的影响,以便为解释林木细根呼吸提供更多的理论依据。
-
核桃细根呼吸过程包括3个呼吸途径,即:TCA途径、EMP途径和PPP途径,其中,TCA途径呼吸速率在基础呼吸速率中占比高于EMP和PPP途径的呼吸速率;核桃细根TCA、PPP的呼吸速率与MDH、G-6-PDH活性极显著正相关,EMP的呼吸速率与PFK活性极显著正相关,调节各呼吸速率的关键酶活性高低直接影响呼吸速率的强弱。适宜的施N量可以促进核桃细根Q和EMP、TCA、PPP的呼吸速率及其相关酶活性,过高或过低都会对核桃细根呼吸速率和相关酶活性产生抑制。
施氮水平对核桃细根呼吸速率及相关酶活性的影响
Effects of Nitrogen Application Levels on Respiration Rate and Related Enzyme Activity of Walnut Root
-
摘要:
目的 以新疆南疆盆地核桃主栽品种‘新温185’(Juglans regia ‘Xinwen185’)为试验材料,探究不同施氮水平对核桃生长发育过程中细根呼吸速率及相关酶活性的影响,旨在进一步加深氮素对根系呼吸调控机理的认识。 方法 通过设置田间施肥试验,采用离体根测定法,分析核桃在不同施氮水平下细根基础呼吸速率(Q)和糖酵解途径(EMP)、三羧酸循环途径(TCA)、磷酸戊糖途径(PPP)呼吸速率的动态变化及磷酸果糖激酶(PFK)、苹果酸脱氢酶(MDH)和葡萄糖-6-磷酸脱氢酶(G-6-PDH)活性的动态变化以及细根呼吸速率与酶活性之间的相关性。 结果 随着花后时间的变化,核桃细根Q和TCA、PPP呼吸速率总体呈下降趋势,EMP呼吸速率呈先升后降趋势;细根TCA、EMP、PPP呼吸速率分别占Q呼吸速率的40.23%、33.02%、20.47%;N2水平的细根Q和TCA、PPP、EMP呼吸速率以及MDH、G-6-PDH和PFK活性高于其它施氮水平。 结论 TCA途径呼吸速率在基础呼吸速率中占比高于EMP和PPP途径呼吸速率,适量施用氮肥能够提高细根MDH和PFK活性,促进核桃细根呼吸,有利于核桃的生长,施氮水平过高或过低均会抑制细根呼吸速率,对核桃的生长造成不利影响。 Abstract:Objective To further understand the mechanism of nitrogen regulation on root respiration of Juglans regia 'Wen185', the main walnut cultivar in South Xinjiang Basin. Method J. regia 'Wen185' was used as experimental material to study the effect of different nitrogen application levels on the fine root respiration rate and related enzyme activity during the growth and development of walnut. Field fertilizer experiment setting and in vitro root determination method were conducted to analyse the fine root's respiration rate (Q) and glycolytic pathway (EMP), Citric acid cycle Pathway (TCA), pentose phosphate pathway (PPP) respiratory rate and the dynamics of phosphofructokinase (PFK), malate dehydrogenase (MDH) and glucose-6-phosphate dehydrogenase (G-6-PDH) activity, as well as the fine root respiration rate and the correlation between enzyme activity under different nitrogen levels. Result With the changes of post-flowering time, the respiration rate of Q, TCA and PPP generally decreased, while the respiration rate of EMP increased at first and then decreased. The respiratory rates of TCA, EMP and PPP accounted for 40.23%, 33.02% and 20.47% of Q respectively. When the nitrogen level (pure quantity) was N2, the respiration rate of Q and TCA, PPP and EMP, as well as the activity of MDH, G-6-PDH and PFK were significantly higher than that of the other nitrogen application levels(P < 0.05). Conclusion TCA pathway respiratory rate occupies a higher proportion in the basic respiratory rate than EMP and PPP pathway. Proper application of nitrogen application levels could improve MDH and PFK activity, and promote root respiration of walnut, and be beneficial to the growth of walnut. Extremely low and high nitrogen application would inhibit root respiration rate and even adversely affect the growth of walnut. -
Key words:
- Juglans regia
- / walnut
- / nitrogen application level
- / fine root
- / respiratory rate
- / enzyme activity
-
图 1 施氮水平对核桃细根呼吸速率的影响
Figure 1. The effect of nitrogen application levels on respiration rate of walnut fine roots
图 2 施氮水平对核桃细根系呼吸速率及相关酶活性的影响
Figure 2. The effects of nitrogen application levels on respiration rate and related enzyme activities of walnut fine roots
表 1 花后时间和施氮水平对核桃细根呼吸速率及相关酶活性影响的二因素多响应变量方差分析
Table 1. Variance analysis of two multivariate responses to the effect of postoperative time and nitrogen application levels on root respiration rate and related enzyme activity
观测指标
Observation index变异来源
Source of variation平方和
SS自由度df 均方
MSF值
F valueP值
P valueTCA呼吸速率
Respiratory rate of TCA花后时间(T)Post flower time 0.002 4 0.001 113.225 <0.05 施氮水平(N)Nitrogen application levels 0.000 3 0.000 16.451 <0.05 花后时间(T)×施氮水平(N)Post flower time ×Napplication levels (T×N) 0.000 12 0.000 2.738 <0.05 EMP呼吸速率
Respiration rate of EMP花后时间(T)Post flower time 0.002 4 0.000 113.231 <0.05 施氮水平(N)Nitrogen application levels 0.000 3 0.000 16.362 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 0.000 12 0.000 2.672 <0.05 PPP呼吸速率
respiration rate of PPP花后时间(T)Post flower time 0.004 4 0.001 172.775 <0.05 施氮水平(N)Nitrogen application levels 0.000 3 0.000 12.062 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 0.000 12 0.000 4.005 <0.05 Q
Basal respiration rate花后时间(T)Post flower time 0.014 4 0.004 113.314 <0.05 施氮水平(N)Nitrogen application levels 0.002 3 0.001 16.400 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 0.001 12 0.000 2.664 <0.05 PFK活性
Activity ofPFK花后时间(T)Post flower time 3 258.907 4 814.727 135.915 <0.05 施氮水平(N)Nitrogen application levels 304.964 3 101.655 16.958 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 142.325 12 11.860 1.979 >0.05 MDH活性
Activity ofMDH花后时间(T)Post flower time 120.167 4 30.042 131.223 <0.05 施氮水平(N)Nitrogen application levels 7.444 3 2.481 10.839 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 4.322 12 0.360 1.573 >0.05 G-6-PDH活性
Activity of G-6-PDH花后时间(T)Post flower time 23.056 4 5.764 153.337 <0.05 施氮水平(N)Nitrogen application levels 1.484 3 0.495 13.195 <0.05 花后时间(T)×施氮水平(N)Post flower time ×N application levels (T×N) 1.295 12 0.108 2.870 <0.05  下载: 导出CSV
下载: 导出CSV
表 2 核桃细根Q和TCA、EMP、PPP的呼吸速率与酶活性的相关系数
Table 2. Correlation of respiration rate and enzyme activity between Q and TCA, EMP and PPP of walnut fine root
测定指标
Observation indexMDH活性
Activity of MDHG-6-PDH活性
Activity of G-6-PDHPFK活性
Activity of PFKQ
Basal respiration rate0.974** 0.834** 0.414 TCA呼吸速率
Respiratory rate of TCA0.970** 0.821** 0.430 EMP呼吸速率
Respiratory rate of EMP0.358 -0.096 0.990** PPP呼吸速率
Respiratory rate of PPP0.851** 0.986** -0.141 注:**在0.01水平上极显著相关(双尾),*在0.05水平上显著相关(双尾)。
Notes:** Correlation is highly significant at the 0.01 level(double tail); * Correlation is significant at the 0.05 level(double tail).
下载: 导出CSV
-
[1] 唐晓鹿, 范少辉, 漆良华, 等.采伐对幕布山区毛竹林土壤呼吸的影响[J].林业科学研究, 2013, 26(1):52-57. [2] 丁杰萍, 罗永清, 周欣, 等.植物根系呼吸研究方法及影响因素研究进展[J].草业学报, 2015, 24(5):206-216. [3] 李志霞, 秦嗣军, 吕德国, 等.植物根系呼吸代谢及影响根系呼吸的环境因子研究进展[J].植物生理学报, 2011, 47(10):957-966. [4] Nakamura T, Nakamura M. Root respiratory costs of ion uptake, root growth, and root maintenance in wetland plants:efficiency and strategy of O2 use for adaptation to hypoxia[J].Oecologia, 2016, 182(3):667-678. [5] 陈惠哲, 朱德峰, 林贤青, 等.土壤透水状况对水稻根系生长与氮素利用的影响[J].中国生态农业学报, 2007, 15(5):34-37. [6] Hao Q, Jiang C.Contribution of root respiration to soil respiration in a rape (Brassica campestris L.) field in Southwest China[J].Plant Soil and Environment, 2014, 60(1):8-14. [7] Burton A J, Jarvey J C, Jarvi M P, et al.Chronic N deposition alters root respiration-tissue N relationship in northern hardwood forests[J]. Global Change Biology, 2012, 18(1):258-266. [8] 周睿智.氮对小叶白蜡、银杏、金叶女贞和小叶黄杨幼树根系呼吸的影响[D].北京: 北京林业大学, 2009. [9] Hawthorne I, Johnson M S, Jassal R S, et al.Application of biochar and nitrogen influences fluxes of CO2, CH4and N2O in a forest soil[J].Journal of Environmental Management, 2017, 192:203-214. [10] Sorrenti G, Buriani G, Gaggia F, et al.Soil CO2emission partitioning, bacterial community profile and gene expression of Nitrosomonas spp.and Nitrobacter spp.of a sandy soil amended with biochar and compost[J].Applied Soil Ecology, 2017, 112:79-89. [11] Dyckmans J, Flessa H.Influence of internal nitrogen reserves on the response of beech (Fagus sylvatica)trees to elevated atmospheric carbon dioxide concentration[J].Tree Physiol, 2002, 22(1):41-49. [12] Ceccon C, Tagliavini M, Schmitt A O, et al.Untangling the effects of root age and tissue nitrogen on root respiration in Populus tremuloides at different nitrogen supply[J].Tree Physiol-ogy, 2016, 36:618-627. [13] Kewald B, Kunze M E, Godbold D L.NH4:NO3, nutrition influence on biomass productivity and root respiration of poplar and willow clones[J].Global Change Biology Bioenergy, 2016, 8(1):51-58. [14] 任军, 徐程杨, 林玉海, 等.不同供氮水平下水曲柳幼苗根系呼吸季节动态[J].生态学报, 2009, 29(8):4169-4178. [15] 孙誉育, 尹春英, 贺合亮, 等.红桦幼苗根系对水-氮耦合效应的生理响应[J].生态学报, 2016, 36(21):6758-6765. [16] 闫美芳, 张新时, 周广胜, 等.不同树龄杨树人工林的根系呼吸季节动态[J].生态学报, 2010, 30(13):3449-3456. [17] 王东光, 尹光天, 邹文涛, 等.氮素营养对闽楠幼苗生长及光合特性的影响[J].林业科学研究, 2013, 26(1):70-75. [18] 刘俊萍, 喻苏琴, 游璐, 等.油茶细根时空分布动态对施钾水平的响应[J].林业科学研究, 2019, 32(1):31-38. [19] 罗达, 史彦江, 宋锋惠, 等.平欧杂种榛细根空间分布特征[J].林业科学研究, 2019, 32(1):81-89. [20] Bouma T J, Yanai R D, Elkin A D, et al. Estimating aged-dependent costs and benefits of roots with contrasting life span:comparing apples and oranges[J].New Phytologist, 2001, 150:685-695. [21] 毛志泉, 王丽琴, 沈向, 等.有机物料对平邑甜茶实生苗根系呼吸强度的影响[J].植物营养与肥料学报, 2004, 10(2):171-175. [22] 高相彬, 赵凤霞, 沈向, 等.肉桂酸对平邑甜茶幼苗根系呼吸速率及相关酶活性的影响[J].中国农业科学, 2009, 42(12):4308-4314. [23] Singla N K, Jain V, Jain S, et al.Activities of glycolytic enzymes in leaves and roots of contrasting cultivars of sorghum during flooding[J].Biologia Plantarum, 2003, 47(4):555-560. [24] Ling K H, Paetkau V, Marcus F, et al.Phosphofructokinase fromskeletal muscles[M].New York:Academy Press, 1996, 9:425-426. [25] 欧阳光察.苹果酸脱氢酶活性测定[M].上海:上海科学技术出版社, 1985:179-181. [26] 薛应龙.植物生理学实验手册[M].上海:上海科学技术出版社, 1985:68-69. [27] Brown A P, Wary J L. Correlated changes of some enzyme activities and cofactor and substrate contents of pea cotyledon tissue during germination[J].Biochemical Journal, 1968, 108:437-444. [28] 王海波, 王孝娣, 史祥宾, 等.葡萄冬芽自然休眠诱导期间的呼吸代谢变化[J].中国农业科学, 2015, 48(16):3287-3295. [29] Vega-Mas I, Cukier C, Coleto I, et al. Isotopic labelling reveals the efficient adaptation of wheat root TCA cycle flux modes to match carbon demand under ammonium nutrition[J]. Scientific reports, 2019, 9(1):8925. [30] 饶龙兵, 李霞, 段红平, 等.不同供氮水平对4种桤木幼苗生长的影响[J].林业科学研究, 2012, 25(2):246-253. [31] Ryan M G, Hubbard R M, Pongracic S, et al. Foliage, fine root, woody-tissue and stand respiration in Pinus radiata in relation to nitrogen status[J].Tree Physiology, 1996, 16(3):333-343. [32] Lambers H, Chapin F S, Pons T L.Plant Physiological Ecology[M].New York:Springer-Verlag, 1998:122-133. [33] Lux A, Luxova M, Abe J, et al.Root cortex:structural and functional variability and responses to environmental stress[J].Root Research, 2004, 13(3):117-131. [34] 范志强, 王政权, 吴楚, 等.不同供氮水平对水曲柳苗木生物量、氮分配及其季节变化的影响[J].应用生态学报, 2004, 15(9):1497-1503. [35] 胡智勇, 陈孝杨, 陈敏, 等.土壤与大气温度及CO2浓度变化对根系呼吸影响研究进展[J].江苏农业科学, 2018, 46(11):1-5. [36] 张鹏, 秦嗣军, 周文杰, 等.荧光假单胞菌对樱桃根系呼吸和幼苗生长的影响[J].中国农业科学, 2014, 47(19):3857-3865. [37] 孙晓莉, 田寿乐, 沈广宁, 等.干旱胁迫下H2S对板栗幼苗根系抗氧化特性及呼吸相关酶活性的影响[J].核农学报, 2019, 33(5):1024-1031. [38] 王闯, 胡艳丽, 高相彬, 等.硝态氮对淹水条件下甜樱桃根系呼吸速率及相关酶活性的影响[J].植物营养与肥料学报, 2009, 15(6):1433-1438. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4586
- HTML全文浏览量: 2178
- PDF下载量: 52
- 被引次数: 0
