

 下载:
下载:




-
我国是农业大国,土壤是立农之本,却被动承载着包括重金属在内的大部分污染物。我国重金属污染的耕地面积达2 000万hm2,每年仅镉污染的粮食达1 200万t,造成的直接经济损失超过200亿元[1]。重金属元素的难降解、易富集及食物链的可传递性给生态环境和食品安全带来严重威胁,是人类生命健康和安全的“隐形杀手”[2]。植物修复技术是近年重金属污染土壤治理的研究热点[3],其中超积累型东南景天(Sedum alfredii Hance)是为数不多的镉超积累植物之一[4],可在地上部分累积镉15 500 μg·g−1(干质量)而不呈现任何毒害作用,是应用于土壤镉污染修复的宝贵材料[5];同时,超积累型东南景天的极强耐镉特性预示着其具有丰富的抗逆基因,是研究植物耐重金属机理的理想物种。
为解析其镉耐性及超积累的机制,大量的研究聚焦于超积累型东南景天的镉吸收、转运及解毒[6-8]。研究发现,超积累型东南景天可以通过高效的根-茎转运机制将高浓度的镉离子转运到地上部分,并最终储存于叶肉的薄壁细胞、茎的髓部及皮层组织中[9]。维持植物细胞内金属离子的平衡及内稳态是提高植物耐重金属的关键[10]。研究表明,超积累型东南景天中有多个基因被发现与镉离子转运等相关,包括SaHMA3(heavy metal ATPase, HMA)[11]、SaNRAMP3(natural resistance-associated macrophage protein, NRAMP) [12]、SaNRAMP6[13]、SaMTP1(metal tolerance protein, MTP)[14]、SaZIP4[15]以及SaCAX2(cation exchangers, CAX)[16]。
ATP结合盒(ATP-binding cassette,ABC)是目前发现的最大的、功能最广泛的蛋白超家族之一,因其在跨膜运输底物时需要借助水解ATP释放的能量完成而得名[17]。ABC基因家族包括全分子和半分子2种类型,全分子蛋白结构包括2个核苷酸结合域(NBD)和2个跨膜结构域(TMD),半分子的蛋白只有1个膜结构域(MSD)和1个NBD[17]。该基因家族广泛存在于各类生物体中,在哺乳动物[18]、微生物[19]、植物[20]等生物中鉴定出ABC家族的物种已超过100种,其中,人类基因组中发现48个ABC基因[18],参与细菌耐药性、次生代谢物积累和肿瘤细胞的抗药性等,受到国内外研究者广泛关注。在植物中,ABC家族被认为参与植物体内激素[21]、脂质[22]、金属离子[23]、次生代谢物[24]以及调控植物与病原体相互作用[25]。然而,目前在超积累型东南景天中尚未开展该基因家族成员的研究。本研究以前期超积累型东南景天组学测序结果为基础,借助同源比对鉴定超积累型东南景天ABC家族基因,并开展生物信息学分析。另外,借助实时定量技术分析干旱、盐碱等不同胁迫处理下ABC基因的表达情况,为该基因功能的深入验证奠定前期基础和理论依据。
-
本实验所选用的植物材料为超积累型东南景天,水培于1/2 Hoagland-Arnon营养液中并置于人工培养室中25℃、16 h光照、8 h黑暗培养,调节pH为6.0,每3 d更换一次水培营养液。
对于盐碱、干旱和脱落酸(ABA)激素胁迫,选取长势一致、根部发育健康、无发黄发红等现象,且主根长度约7 cm的植株进行胁迫处理,处理条件和浓度参照前期实验室的研究[26],具体处理方法为:将东南景天幼苗根部浸入含有100 mmol·L−1 NaCl、20%PEG6000、100 μmol·L−1 ABA的1/2 Hoagland 营养液中,对照幼苗在1/2 Hoagland 溶液中培养,然后于0、0.5、1、6 、12 h后分别取根系。每个时间点取3个生物学重复,用于提取RNA的根部质量约0.1 g。将来自3个生物学重复的所有样品在−80℃ 下冷冻保存以供进一步实验。
主要试剂:T4 DNA 连接酶与 pGEM®-T-easy 载体购自德国 Promega 公司;2×GoldStar Taq MasterMix、DM15000 DNA Marker和DM2000 DNA Marker购自北京康为科技有限公司;DNA 凝胶回收试剂盒、质粒 DNA 小量提取试剂盒购自 Axygen 公司;Trizol Reagent购自GIBCO BRL公司;反转录酶购自Invitrogen公司;实时荧光定量试剂盒购自大连宝生物工程有限公司(Takara);引物合成及测序由上海生工生物技术有限公司完成。
-
参照GIBCO BRL公司的Trizol Reagent操作说明书提取东南景天正常生长样品及盐碱、干旱和ABA激素胁迫处理下根部总RNA的提取。提取出的总RNA由1.2%琼脂糖凝胶电泳鉴定,并经过紫外分光光度计测定其纯度和浓度,使用TE Buffer按照1:100比例稀释RNA样品,然后测定其在260、280、320 nm下的吸光值,计算提取的总RNA纯度和含量。按照Invitrogen公司的反转录试剂盒操作说明书合成cDNA,并置于−80℃冰箱保存用于后续基因表达情况分析。
-
在超积累型东南景天转录组数据库中通过本地Blast(ftp://ftp.ncbi.nlm.nih.gov/)查找ABC家族基因的氨基酸及编码区序列[27],然后利用 HMM(http://www.ebi.ac.uk/Tools/hmmer/search/hmmsearch)在线分析工具鉴定候选的unigenes是否含有TMD和NBD结构域,并根据测序数据的序列号进行命名。
利用MUSCLE软件对SaABC蛋白氨基酸全长序列进行比对,随后在MEGA 7.0软件中用Neighbor-Joining(NJ)法进行系统发育树的构建,设置bootstrap测试为1 000个重复。用PFAM(https://pfam.xfam.org)和SMART(http://smart.embl-heidelberg.de/)在线预测网站确定候选基因的结构域组成,再将这些候选基因氨基酸序列在TMHMM v2.0 (http://www.cbs.dtu.dk/services/TMHMM/)网站上预测跨膜结构域(transmembrane domain, TM)。
在前期转录组数据共表达网络分析获得的与镉胁迫关系密切的关键基因(hub类基因)[27]中查找隶属于ABC家族的基因,并下载该基因对应的关联基因(edge基因)及相关性系数,筛选出皮尔逊相关系数(Pearson Correlation Coefficient, PCC)阈值大于等于0.40的edge基因作为后续分析对象。利用基迪奥生物omicshare云平台(http://www.omicshare.com/tools/)中的GO富集分析功能对所有符合标准的edge基因进行GO分类。最后利用Cytoscape v3.6.1中的NetworkAnalyzer插件对hub基因与edge基因的相关性进行分析,并生成可视化图片。
-
使用NCBI网站的Primer-BLAST 在线工具(https://www.ncbi.nlm.nih.gov/)基于候选基因序列的非保守区设计实时荧光定量PCR(Quantitative Real-time PCR, qRT-PCR)的引物qSaABC-F(5’-GGAGCAAGATGACGTTCCTGA-3’)和qSaABC-R(5’-GCTTCACCGGTCCATCTTTTTC-3’)。利用大连宝生物工程有限公司(Takara)的TB GreenTM Premix Ex TaqTM II(Tli RNaseH Plus)(Code No. RR820A)试剂盒在Applied Biosystems 7300 实时荧光定量PCR仪中进行qRT-PCR反应。具体反应体系如下:2×TB GreenTM Premix Ex TaqTM II(Tli RNaseH Plus)10 μL,ROX Reference Dye 0.4 μL,Primer-qSa12F279-F(10 μmol·L−1)0.8 μL,Primer-qSa12F279-F(10 μmol·L−1)0.8 μL cDNA 2 μL,dH2O 6 μL。将上述反应物短暂离心后置于Applied Biosystems 7300 实时荧光定量PCR仪中,按照两步法设置实时荧光定量PCR扩增程序如下:阶段1:95℃ 30 s;阶段2:95℃ 5 s,60℃ 31 s(40 个循环);融解曲线:95℃ 15 s,60℃ 60 s,95℃ 15 s。
以SaUBC9基因作为内参基因[28],采用2−ΔΔCT的方法[29]计算候选SaABC基因在根中盐胁迫、干旱胁迫和ABA激素胁迫下的相对表达量。
-
在东南景天转录组数据库中运用本地Blast查找到表达水平受到镉胁迫调控的ABC基因。该基因的开放阅读框长度为4 497 bp,编码蛋白长度为1 498个氨基酸,相对分子量为167.1 KD,等电点为6.92,根据测序数据的编号将其命名为Sa12F279基因。
以模式生物拟南芥(Arabidopsis thaliana)的ABC家族成员为参照基因,构建系统进化树,鉴定Sa12F279基因的亚家族分类。进化树显示:拟南芥的ABC成员聚为11个亚类,Sa12F279基因与C亚类成员聚为一类,并与AtABCC9(AT3G601601)和AtABCC15(AT3G609701)2个基因关系最相近(图1)。

图 1 以拟南芥ABC家族基因为参照的Sa12F279基因的进化分析
Figure 1. Phylogenetic analysis of Sa12F279 with reference to AtABC family
-
基于在线网站pHammer等分析Sa12F279基因的结构域组成分布情况,鉴定其是否与拟南芥ABC家族中的C亚类成员类似,具有保守结构域排布。分析显示:Sa12F279基因的蛋白结构域构成呈现TMD-NBD-TMD-NBD的排布方式(图2)。Sa12F279基因的TMD和NBD与拟南芥ABC家族C亚类成员的相应结构域长度差异较小;除AtABCC15外,4个结构域在蛋白质一级结构中的定位位置也较为相近。结构域排布及分布的一致性也佐证了该基因为C亚类的一员,为其功能探索提供了方向。
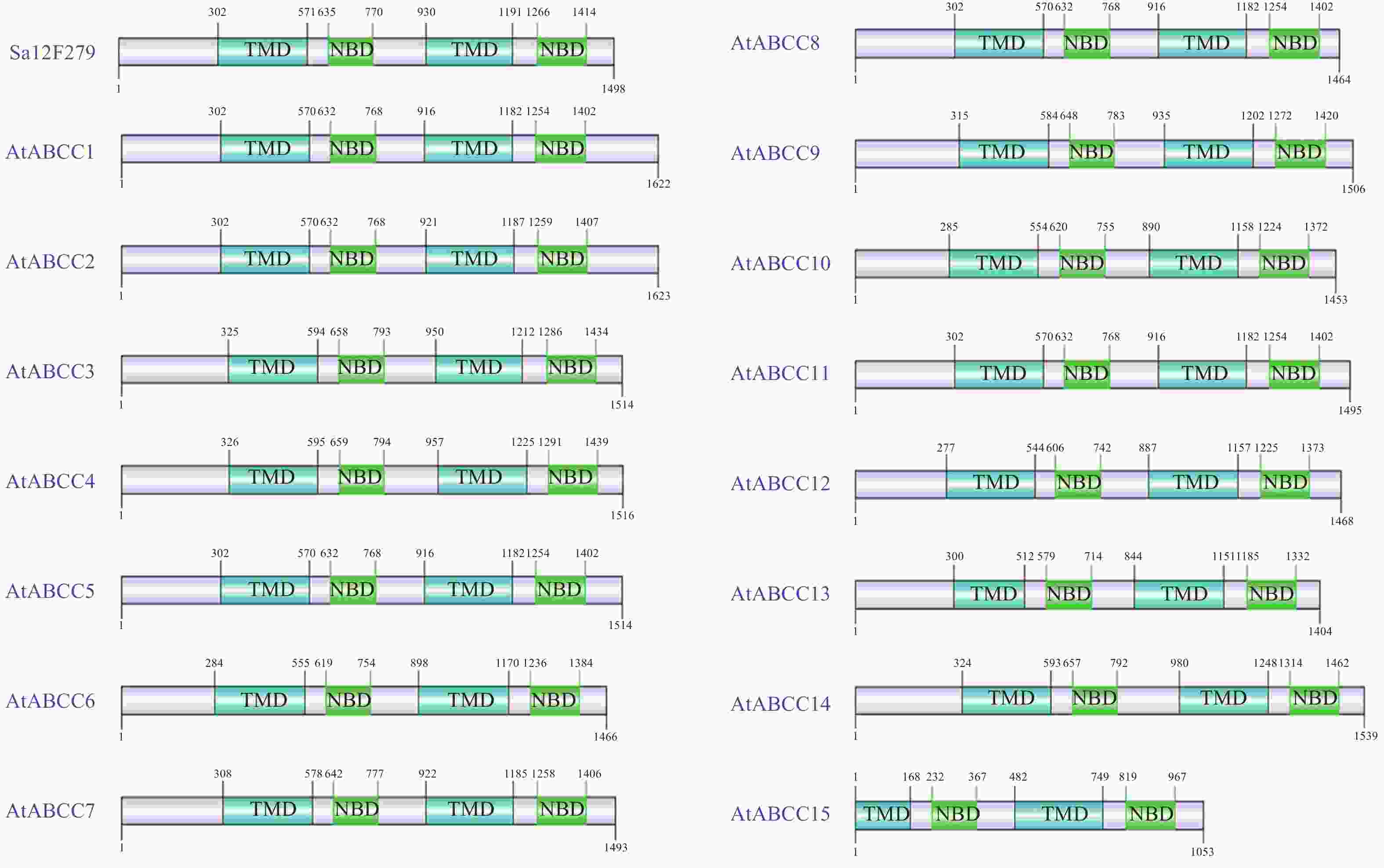
图 2 Sa12F279基因与拟南芥ABC家族C亚类成员的结构域组成比较
Figure 2. Comparison of domian structure among Sa12F279 and C-subfamily of AtABC
利用MegAlign软件进行同源比对分析显示:Sa12F279基因与拟南芥ABC家族的15个C亚类成员的氨基酸序列一致性在45.3%~77.1%,其中,2个NBD结构域和2个TMD结构域序列具有保守基序的一致性,NBD结构域也包含了Walker A、Walker B以及ABC 转运蛋白的一致基序(图3)。

图 3 Sa12F279基因与拟南芥ABC家族C亚类成员结构域的同源比对分析
Figure 3. Alignment of NBD and TMD domians among Sa12F279 and C-subfamily of AtABC
-
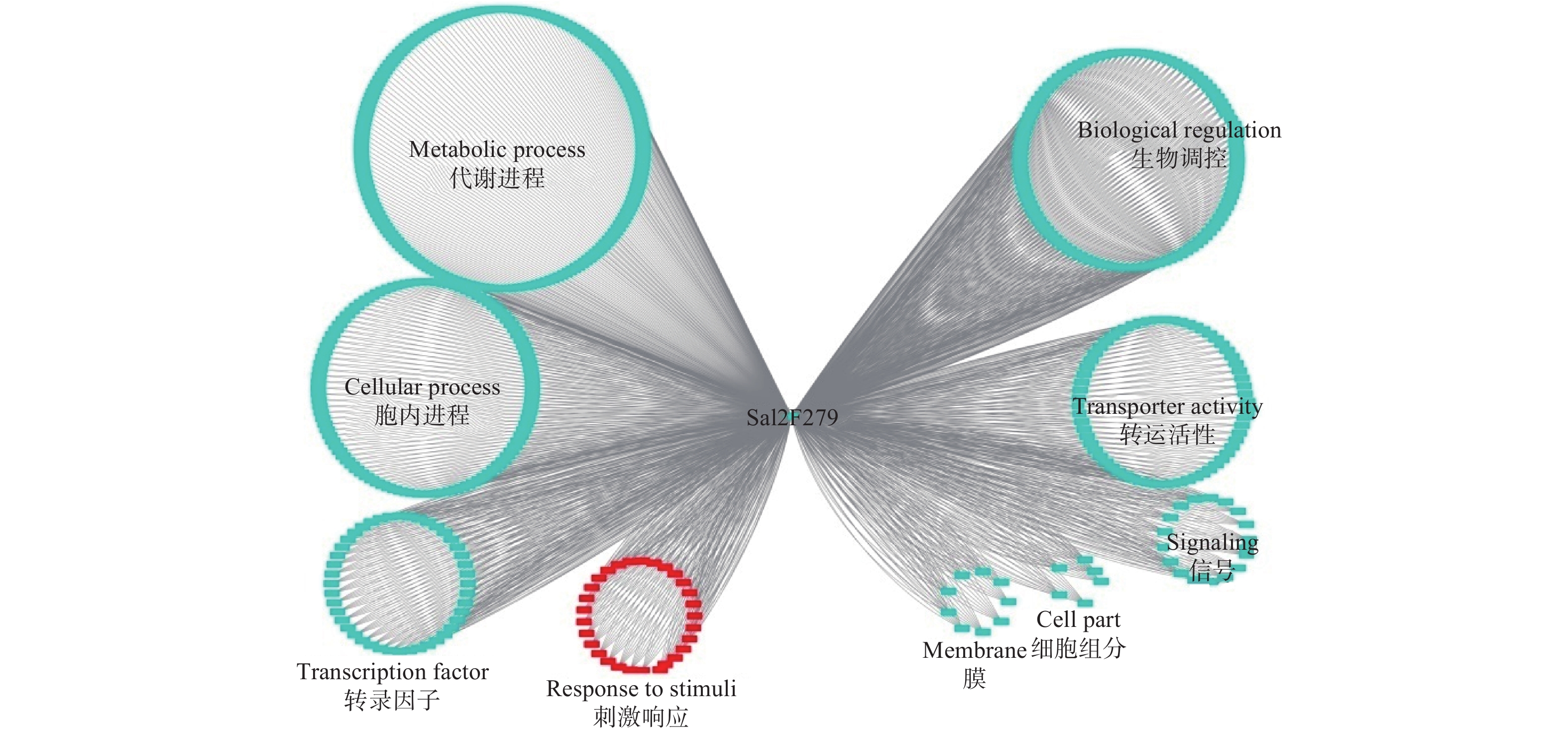
Sa12F279基因的共表达网络分析显示:其与567个基因存在功能上的关联(图4)。对这些基因进行功能注释,结果显示:39.8%的基因执行代谢相关功能(Metabolic process),29.3%的基因与细胞内进程相关(Cellular process),10.2%的基因涉及生物调控(Biological regulation),7.1%的基因参与转运活性(Transporter activity)。通过关联基因的功能,作者推测该基因更有可能担负着细胞内代谢物质等的运输,与超积累型东南景天的生长发育等进程密切相关。
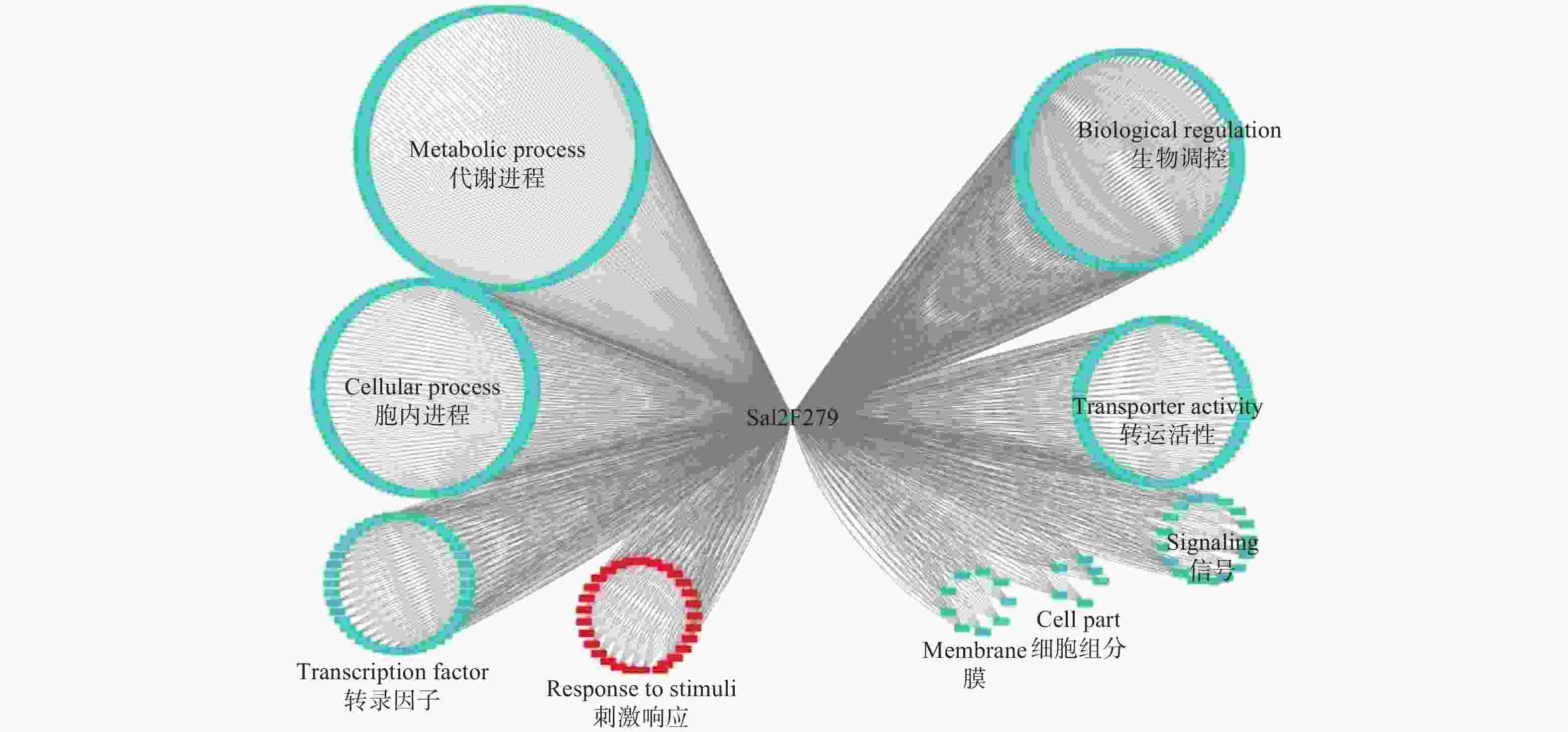
图 4 基于转录组数据的Sa12F279基因共表达网络分析
Figure 4. Co-expression network analysis of Sa12F279
-
利用前期获得的镉胁迫前后超积累型东南景天的转录组数据,进行Sa12F279基因镉胁迫表达分析(图5)。结果显示:在镉胁迫24 h的样品中,Sa12F279基因在根、茎、叶3种组织中均出现了下调的应答模式,与对照组相比,根和茎中的下调比叶中更显著。在镉胁迫96 h的样品中,Sa12F279基因的表达水平与镉胁迫24 h相比未出现显著变化。基于此,作者推测Sa12F279基因有可能不直接参与镉胁迫应答,有可能与其它胁迫相关,或参与正常生理进程的维持。

图 5 基于转录组数据的Sa12F279基因胁迫24、96 h下根、茎、叶组织的镉响应模式
Figure 5. The 24 h and 96 h Cd responsive patterns of Sa12F279 in the root, stem and leaf based on the transcriptomic data
-
为进一步分析Sa12F279基因对其它代表性非生物胁迫的响应情况,采用实时定量聚合酶链式反应(qRT-PCR)方法,研究干旱、盐碱及ABA激素处理下Sa12F279基因在根中的响应模式,并借助SPSS软件进行T-test显著性分析(图6)。结果显示:在ABA激素处理0.5 h时,Sa12F279基因出现表达量的显著下调,随后出现表达量的上调应答,在1 h和12 h处理条件下呈现表达量的上升(图6A)。在盐胁迫处理条件下,Sa12F279基因在处理前期未有明显的表达量变化,而在12 h处理时出现表达量的显著上调(图6B)。在干旱胁迫处理下,Sa12F279基因在1 h处理时显著下调,其他时间点表达量升高或变化不明显(图6C)。综合3种胁迫处理,作者发现,Sa12F279基因对不同胁迫的响应趋势存在差异,但均未有极显著的应答产生,这意味着Sa12F279基因可能更重要的功能是参与生长发育的调控,但具体功能仍需更有力的实验结果进一步证实。

图 6 Sa12F279基因在ABA激素、盐碱及干旱胁迫下根中的响应模式。
Figure 6. The responsive patterns of Sa12F279 in the root under the abiotic stresses including ABA, salt and drought.
-
超积累型东南景天是镉超积累植物,蕴含了大量与镉耐性、吸收相关的基因[8, 30]。它既是研究镉耐性及富集机制的宝贵材料,同时在植物修复中具有巨大的应用潜力。ABC类蛋白作为介导物质运输的重要家族,其功能研究一直受到广泛的关注[31-32]。在人类中,研究者针对ABC家族的C亚类转运蛋白的底物特异性、对组织表达以及转运动力学开展广泛的研究,而这些研究对探索细胞功能、新药物研发和药物代谢研究等都具有非常重要的意义[25]。在植物中,ABC家族的C亚类转运蛋白最初被发现是位于液泡上,负责转运谷胱甘肽复合物[33];后续随着研究的深入,该家族成员被发现参与重金属污染物的摄取[34]、盐胁迫的抵御[35]、根际分泌物的运输[36]以及离子通道的调节[34]。然而该家族成员在超积累型东南景天中尚缺乏相关研究。
本研究在前期镉胁迫东南景天转录组测序数据中[27]鉴定了属于ABC家族的1个基因,并将其命名为Sa12F279基因。对该基因的进化及蛋白结构的研究有助于提供该基因功能的线索。通过进化分析,作者发现,Sa12F279基因与拟南芥ABC转运蛋白家族中的C亚类聚为一簇,同时也呈现TMD和NBD结构域的保守排布形式。这意味着Sa12F279基因的功能或许更倾向于C亚类成员的功能,然而同一基因家族成员之间功能也存在着差异或冗余[31, 37],需要开展深层次的功能研究。
转录组测序反映了整体水平上mRNA的表达及不同基因之间的关联[38-39]。通过分析Sa12F279基因在转录组数据中的关联基因,作者发现其功能关联的基因多参与代谢途径、细胞内进程以及生物调控等;同时,其对镉胁迫的响应呈现下调的应答模式。这意味着Sa12F279基因的功能可能更偏重于参与生长、发育等进程,在镉胁迫下,其表达受到抑制,便于植物体调控更为核心的ABC家族成员响应胁迫。同样,在拟南芥中,不同的ABCC成员的功能也存在较大的差异,并非所有的成员都与重金属的转运或耐性相关,如AtABCC4调控气孔的运动[40],AtABCC6被发现与根的次生生长相关[41]。
另外,作者发现,Sa12F279基因对ABA激素、盐碱和干旱3种非生物胁迫的应答模式也存在差异,且表达量的变化幅度较平缓,这在另一方面佐证了作者对Sa12F279基因的功能推测。然而,表达水平的变化受到多种因素的影响[42],仅通过镉、干旱、ABA激素及盐碱胁迫响应模式断定其是否参与重金属离子的转运及解毒尚不够充分,更强有力的实验证据源自基因的异源表达及转基因材料的镉耐性及镉含量。而作者的研究工作将为超积累型东南景天ABC转运蛋白的研究提供前期基础和参考依据。
-
在东南景天转录组数据库中,作者通过序列比对获得1个ABC家族成员的基因,并将其命名为Sa12F279基因。Sa12F279基因在进化上与C亚类成员聚为一簇,且在结构域构成上符合ABC家族的保守排列。Sa12F279基因的共表达网络分析显示:与其功能关联的基因多参与代谢途径、细胞内进程以及生物调控等。转录组数据显示:Sa12F279基因对镉胁迫呈现下调响应,对ABA激素、盐碱和干旱胁迫的应答模式也存在差异,且表达量的变化幅度较平缓。在ABA激素处理下,呈现先下调后上升的趋势;在盐胁迫下,呈现早期响应不显著而于胁迫后期出现上调应答;在干旱处理下,呈现先升后降又升的趋势。基于上述研究,作者推测Sa12F279基因可能更多地参与生长、发育等代谢进程,而非镉胁迫或其它非生物胁迫的主要响应因子,更加精确的功能有待更进一步的功能研究。
超积累型东南景天Sa12F279基因的抗逆表达响应及功能关联分析
Characterization of Expression Patterns of Sa12F279 from Sedum alfredii under Abiotic Stress and Its Functional Interaction Analysis
-
摘要:
目的 对超积累型东南景天ABC转运蛋白家族成员Sa12F279开展生物信息学分析及表达分析,为探究ABC类转运蛋白在镉、干旱、盐碱等非生物胁迫中的功能提供参考。 方法 通过对超积累型东南景天转录组数据库进行比对分析,获得1个ABC家族成员的基因。通过生物信息学方法分析Sa12F279基因的进化关系、蛋白结构域构成、核心结构域同源比对及在组学数据中相关的互作蛋白分类;借助实时定量PCR技术分析该基因在盐碱、干旱及ABA激素胁迫下根中的表达变化。 结果 通过比对分析获得ABC家族成员Sa12F279基因,该基因的开放阅读框长度为4 497 bp,编码蛋白长度为1 498个氨基酸,相对分子量为167.1 KD,等电点为6.92。进化分析显示:Sa12F279基因与C亚类成员聚为一簇,且在结构域构成上符合ABC家族的保守排布。共表达网络分析显示:Sa12F279与 567个基因存在功能上的关联,对这些基因进行功能注释发现,其中39.8%的基因执行代谢相关功能,29.3%的基因与细胞内进程相关,10.2%的基因涉及生物调控,7.1%的基因参与转运活性。对镉胁迫前后转录组数据分析显示,Sa12F279基因在根、茎、叶3种组织中,取样点24 h和96 h的表达量均呈现对镉胁迫下调响应。实时定量结果显示:该基因对ABA激素、盐碱和干旱不同胁迫的应答模式存在差异,且应答响应较平缓。在ABA激素处理下,呈现先下调后上升的趋势;在盐胁迫下,呈现早期响应不显著而于胁迫后期出现上调应答;在干旱处理下,呈现先升后降又升的趋势。 结论 鉴定了超积累型东南景天ABC家族的1个Sa12F279基因,揭示了该基因在不同非生物胁迫下的表达模式。 Abstract:Objective In order to elucidate the functions of the ATP-binding cassette (ABC) protein in Sedum alfredii, a Cd hyperaccumulator, the bioinformatics analysis and expression pattern study were performed on Sa12F279 belonging to ABC protein family. Methods A local blast was carried out in the transcriptomic datasets of S. alfredii and produced a transcript categorized to ABC protein family. Further bioinformatics analysis was conducted on the phylogenetic clustering, the protein domain structures and classification of possible interaction proteins. Quantitative real-time PCR (qRT-PCR) was applied to uncover the expression patterns of Sa12F279 in the root under drought, salt, abscisic acid stresses. Result In the present work, a gene belonging to C subfamily of ABCs was identified in S. alfredii using local blast and designated as Sa12F279. The length of Sa12F279 open reading frame was 4 497 bp, coding a protein of 1 498 amino acids with pI being 6.92. The phylogenetic analysis showed that Sa12F279 was clustered with the C-subfamily of ABC protein, exhibiting uniform domain structure of TMD-NBD-TMD-NBD. Co-expression network analysis indicated that Sa12F279 served as hub genes mostly associated with metabolic process (39.8%), cellular process (29.3%), biological regulation (10.2%) and transporter activity (7.1%). The expression data from the transcriptomic datasets showed that Sa12F279 was suppressed by Cd stress in the three tissues (root, stem and leaf) at the sampling points of 24 h and 96 h. Transcriptional expression profiles responsive to drought, salt, abscisic acid (ABA) stress illustrated different response tendencies showing moderate induction. Under the treatment of ABA, the expression of Sa12F279 exhibited the tendency of decrease to elevation. Under the salt stress, Sa12F279 was up-regulated at the late stage of treatment while under the drought stress, Sa12F279 displayed the irregular tendency of elevation-decrease-elevation. Conclusion Based on the studies above, the authors characterize a gene named Sa12F279 belonging to the ABC protein family in S. alfredii and performs a comprehensive analysis including phylogenetic analysis, protein structure and co-expression network. Meanwhile, the expression profiles under different abiotic stress are also uncovered. The results could provide reference for the further studies on the ABC protein family. -
Key words:
- Sedum alfredii
- / ABC transporters
- / abiotic stress
- / expression pattern
-
图 1 以拟南芥ABC家族基因为参照的Sa12F279基因的进化分析
Figure 1. Phylogenetic analysis of Sa12F279 with reference to AtABC family
图 2 Sa12F279基因与拟南芥ABC家族C亚类成员的结构域组成比较
Figure 2. Comparison of domian structure among Sa12F279 and C-subfamily of AtABC
图 3 Sa12F279基因与拟南芥ABC家族C亚类成员结构域的同源比对分析
Figure 3. Alignment of NBD and TMD domians among Sa12F279 and C-subfamily of AtABC
图 5 基于转录组数据的Sa12F279基因胁迫24、96 h下根、茎、叶组织的镉响应模式
Figure 5. The 24 h and 96 h Cd responsive patterns of Sa12F279 in the root, stem and leaf based on the transcriptomic data
-
[1] Tang X, Li Q, Wu M, et al. Review of remediation practices regarding cadmium-enriched farmland soil with particular reference to China[J]. Journal of Environmental Management, 2016, 181: 646-662. doi: 10.1016/j.jenvman.2016.08.043 [2] Chaparro Leal L T, Guney M, Zagury G J. In vitro dermal bioaccessibility of selected metals in contaminated soil and mine tailings and human health risk characterization[J]. Chemosphere, 2018, 197: 42-49. doi: 10.1016/j.chemosphere.2018.01.008 [3] Ali H, Khan E, Sajad M A. Phytoremediation of heavy metals--concepts and applications[J]. Chemosphere, 2013, 91(7): 869-881. doi: 10.1016/j.chemosphere.2013.01.075 [4] Li J T, Gurajala H K, Wu L H, et al. Hyperaccumulator plants from China: a synthesis of the current state of knowledge[J]. Environmental Science & Technology, 2018, 52(21): 11980-11994. [5] Deng D M, Deng J C, Li J T, et al. Accumulation of zinc, cadmium, and lead in four populations of Sedum alfredii growing on lead/zinc mine spoils[J]. Journal of Integrative Plant Biology, 2008, 50(6): 691-698. doi: 10.1111/j.1744-7909.2008.00669.x [6] Tian S, Lu L, Labavitch J, et al. Cellular sequestration of cadmium in the hyperaccumulator plant species Sedum alfredii[J]. Plant Physiology, 2011, 157(4): 1914-1925. doi: 10.1104/pp.111.183947 [7] Tao Q, Jupa R, Luo J, et al. The apoplasmic pathway via the root apex and lateral roots contributes to Cd hyperaccumulation in the hyperaccumulator Sedum alfredii[J]. Journal of Experimental Botany, 2017, 68(3): 739-751. [8] Liu M, He X, Feng T, et al. cDNA Library for mining functional genes in Sedum alfredii Hance related to cadmium tolerance and characterization of the roles of a novel SaCTP2 gene in enhancing cadmium hyperaccumulation[J]. Environmental Science & Technology, 2019, 53(18): 10926-10940. [9] Tian S, Xie R, Wang H, et al. Uptake, sequestration and tolerance of cadmium at cellular levels in the hyperaccumulator plant species Sedum alfredii[J]. Journal of Experimental Botany, 2017, 68(9): 2387-2398. doi: 10.1093/jxb/erx112 [10] Conte S S, Walker E L. Transporters contributing to iron trafficking in plants[J]. Molecular Plant, 2011, 4(3): 464-476. doi: 10.1093/mp/ssr015 [11] Zhang J, Zhang M, Shohag M J, et al. Enhanced expression of SaHMA3 plays critical roles in Cd hyperaccumulation and hypertolerance in Cd hyperaccumulator Sedum alfredii Hance[J]. Planta, 2016, 243(3): 577-589. doi: 10.1007/s00425-015-2429-7 [12] Feng Y, Wu Y, Zhang J, et al. Ectopic expression of SaNRAMP3 from Sedum alfredii enhanced cadmium root-to-shoot transport in Brassica juncea[J]. Ecotoxicology and Environmental Safety, 2018, 156: 279-286. doi: 10.1016/j.ecoenv.2018.03.031 [13] Chen S, Han X, Fang J, et al. Sedum alfredii SaNramp6 metal transporter contributes to Cadmium accumulation in transgenic Arabidopsis thaliana[J]. Scientific Reports, 2017, 7(1): 13318. doi: 10.1038/s41598-017-13463-4 [14] Zhang M, Senoura T, Yang X, et al. Functional analysis of metal tolerance proteins isolated from Zn/Cd hyperaccumulating ecotype and non-hyperaccumulating ecotype of Sedum alfredii Hance[J]. FEBS Letters, 2011, 585(16): 2604-2609. doi: 10.1016/j.febslet.2011.07.013 [15] Yang Q Y, Ma X X, Luo S, et al. SaZIP4, an uptake transporter of Zn/Cd hyperaccumulator Sedum alfredii Hance[J]. Environmental and Experimental Botany, 2018, 155: 107-117. doi: 10.1016/j.envexpbot.2018.06.021 [16] Zhang M, Zhang J, Lu L L, et al. Functional analysis of CAX2-like transporters isolated from two ecotypes of Sedum alfredii[J]. Biologia Plantarum, 2016, 60(1): 37-47. doi: 10.1007/s10535-015-0557-3 [17] Beis K. Structural basis for the mechanism of ABC transporters[J]. Biochem Soc Trans, 2015, 43(5): 889-893. doi: 10.1042/BST20150047 [18] Dean M, Hamon Y G C. The human ATP-binding cassette (ABC) transporter superfamily[J]. J Lipid Res, 2001, 42(7): 1007-1017. [19] Kumari S, Kumar M, Khandelwal N K, et al. ABC transportome inventory of human pathogenic yeast Candida glabrata: Phylogenetic and expression analysis[J]. Plos One, 2018, 13(8): e0202993. doi: 10.1371/journal.pone.0202993 [20] Cakir B, Kilickaya O. Whole-genome survey of the putative ATP-binding cassette transporter family genes in Vitis vinifera[J]. Plos One, 2013, 8(11): e78860. doi: 10.1371/journal.pone.0078860 [21] Cho M, Cho H T. The function of ABCB transporters in auxin transport[J]. Plant Signaling & Behavior, 2013, 8(2): e22990. [22] da Costa K M, Valente R C, Salustiano E J, et al. Functional characterization of ABCC proteins from Trypanosoma cruzi and their involvement with thiol transport[J]. Frontiers in Microbiology, 2018, 9: 205. doi: 10.3389/fmicb.2018.00205 [23] Park J, Song W Y, Ko D, et al. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury[J]. The Plant Journal, 2012, 69(2): 278-288. doi: 10.1111/j.1365-313X.2011.04789.x [24] Nagy R, Grob H, Weder B, et al. The Arabidopsis ATP-binding cassette protein AtMRP5/AtABCC5 is a high affinity inositol hexakisphosphate transporter involved in guard cell signaling and phytate storage[J]. The Journal of Biological Chemistry, 2009, 284(48): 33614-33622. doi: 10.1074/jbc.M109.030247 [25] Gu X, Manautou J E. Regulation of hepatic ABCC transporters by xenobiotics and in disease states[J]. Drug Metab Rev, 2010, 42(3): 482-538. doi: 10.3109/03602531003654915 [26] Chen S S, Jiang J, Han X J, et al. Identification, expression analysis of the Hsf family, and characterization of Class A4 in Sedum alfredii Hance under cadmium stress[J]. International Journal of Molecular Sciences, 2018, 19(4): 1216. doi: 10.3390/ijms19041216 [27] Han X, Yin H, Song X, et al. Integration of small RNAs, degradome and transcriptome sequencing in hyperaccumulator Sedum alfredii uncovers a complex regulatory network and provides insights into cadmium phytoremediation[J]. Plant Biotechnology Journal, 2016, 14(6): 1470-1483. doi: 10.1111/pbi.12512 [28] Sang J, Han X, Liu M, et al. Selection and validation of reference genes for Real-Time Quantitative PCR in hyperaccumulating ecotype of Sedum alfredii under different heavy metals stresses[J]. Plos One, 2013, 8(12): e82927. doi: 10.1371/journal.pone.0082927 [29] Liu M, Jiang J, Han X, et al. Validation of reference genes aiming accurate normalization of qRT-PCR data in Dendrocalamus latiflorus Munro[J]. Plos One, 2014, 9(2): e87417. doi: 10.1371/journal.pone.0087417 [30] 刘明英, 乔桂荣, 蒋 晶, 等. 矿山型东南景天cDNA表达文库构建与耐镉基因筛选[J]. 林业科学研究, 2012, 25(3):332-338. doi: 10.3969/j.issn.1001-1498.2012.03.010 [31] Aryal B, Laurent C, Geisler M. Correction: Learning from each other: ABC transporter regulation by protein phosphorylation in plant and mammalian systems[J]. Biochem Soc Trans, 2016, 44(2): 663-673. doi: 10.1042/BST20150128_2 [32] Lane T S, Rempe C S, Davitt J, et al. Diversity of ABC transporter genes across the plant kingdom and their potential utility in biotechnology[J]. BMC Biotechnol, 2016, 16(1): 47. doi: 10.1186/s12896-016-0277-6 [33] Lu Y P, Li Z S, Rea P A. AtMRP1 gene of Arabidopsis encodes a glutathione S-conjugate pump: isolation and functional definition of a plant ATP-binding cassette transporter gene[J]. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(15): 8243-8248. doi: 10.1073/pnas.94.15.8243 [34] Suh S J, Wang Y F, Frelet A, et al. The ATP binding cassette transporter AtMRP5 modulates anion and calcium channel activities in Arabidopsis guard cells[J]. The Journal of Biological Chemistry, 2007, 282(3): 1916-1924. doi: 10.1074/jbc.M607926200 [35] Lee E K, Kwon M, Ko J H, et al. Binding of sulfonylurea by AtMRP5, an Arabidopsis multidrug resistance-related protein that functions in salt tolerance[J]. Plant Physiology, 2004, 134(1): 528-538. doi: 10.1104/pp.103.027045 [36] Badri D V, Loyola-Vargas V M, Broeckling C D, et al. Altered profile of secondary metabolites in the root exudates of Arabidopsis ATP-binding cassette transporter mutants[J]. Plant Physiology, 2008, 146(2): 762-771. doi: 10.1104/pp.107.109587 [37] Baker A, Carrier D J, Schaedler T, et al. Peroxisomal ABC transporters: functions and mechanism[J]. Biochem Soc Trans, 2015, 43(5): 959-965. doi: 10.1042/BST20150127 [38] Gao J, Sun L, Yang X, et al. Transcriptomic analysis of cadmium stress response in the heavy metal hyperaccumulator Sedum alfredii Hance[J]. Plos One, 2014, 8(6): e64643. [39] Halimaa P, Lin Y F, Ahonen V H, et al. Gene expression differences between Noccaea caerulescens ecotypes help to identify candidate genes for metal phytoremediation[J]. Environmental Science & Technology, 2014, 48(6): 3344-3353. [40] Klein M, Geisler M, Suh S J, et al. Disruption of AtMRP4, a guard cell plasma membrane ABCC-type ABC transporter, leads to deregulation of stomatal opening and increased drought susceptibility[J]. The Plant Journal, 2004, 39(2): 219-236. doi: 10.1111/j.1365-313X.2004.02125.x [41] Gaillard S, Jacquet H, Vavasseur A, et al. AtMRP6/AtABCC6, an ATP-binding cassette transporter gene expressed during early steps of seedling development and up-regulated by cadmium in Arabidopsis thaliana[J]. BMC Plant Biology, 2008, 8: 22. doi: 10.1186/1471-2229-8-22 [42] Ubrihien R P, Ezaz T, Taylor A M, et al. The response of Isidorella newcombi to copper exposure: Using an integrated biological framework to interpret transcriptomic responses from RNA-seq analysis[J]. Aquatic Toxicology, 2017, 185: 183-192. doi: 10.1016/j.aquatox.2017.02.014 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4851
- HTML全文浏览量: 2098
- PDF下载量: 76
- 被引次数: 0
