




-
许多植物中都存在绿色叶片出现黄色或白色斑块的突变体[1-7],这些突变植株是研究光合作用、叶绿素合成代谢和叶绿体发育等过程的极佳材料[8]。此外,这类表型也是许多观赏花卉的主要观赏性状,是园艺工作者对花卉品种的主要改良和育种目标之一[9-11]。因此,探究叶片黄色或白色的斑化形成机制具有重要的生物学意义和应用价值。
叶绿素是使植物叶片显绿色的主要色素,其在植物体内的生物合成通路已经得到了详细描述和报道[12-15]。许多研究表明,叶绿素生物合成途径中结构基因的表达量变化会直接决定叶绿素在组织中的积累量,从而影响叶片颜色的表型[16-17]。当这些结构基因表达量下降时,会导致中间产物无法被催化成叶绿素,最终使叶片失绿从而产生白色或黄色的斑块或条纹[15, 18-20];同时,造成组织失绿的具体机制在不同植物之间存在差异。因此,探索不同植物突变体叶片失绿的具体分子机制具有重要的意义,是对相关植物进行叶色园艺性状改良的理论基础和依据。
兰科(Orchidaceae)植物因其独特的花形、丰富的花色和迷人的花香而世界闻名,其许多种和栽培品种已成为广受大众欢迎的观赏花卉。叶片失绿的突变现象在兰花中被称为“叶艺”,其能够提高兰花的观赏价值,是兰花园艺工作者主要的选育目标之一[21]。但是,目前叶艺在兰科中的分子机制研究较少,仅在兰属(Cymbidium)[20, 22-23]和卡特兰属(Cattleya)[24]有过相关报道,大多研究主要在生理水平上对叶艺机制进行探索[10, 25-29]。近年来,许多石斛属(Dendrobium)的叶艺突变体在市场上越来越受欢迎。铁皮石斛(Dendrobium catenatum)在自然环境和人工栽培条件下常出现叶艺现象[30],这些叶艺铁皮石斛是石斛品种改良与培育的优质种质资源。此外,铁皮石斛的高质量基因组已经公布[31],这为研究铁皮石斛叶艺现象的分子机制提供了便利与基础。
-
供试材料为铁皮石斛叶艺突变体的无性系,其叶片上有大面积白色斑块或条纹。本研究所用材料都培养在中国林业科学研究院温室中,采光为自然光照,正常栽培管理。
-
分别取正常铁皮石斛(WT)、叶艺铁皮石斛叶子绿色组织(YYG)、叶艺铁皮石斛叶艺白色组织(YYW),用双面刀片快速切成薄片,放入有蒸馏水的玻璃皿中。选择其中最薄的透明度最大的薄片做成临时切片,在光学显微镜(Olympus,日本)下观察和拍照。
-
根据Li等[15]的方法,将实验材料制备成切片;利用透射电镜(JEOL,日本)对切片中的叶绿体显微结构进行观察和拍照。
-
本研究采用邵勤等[32]的叶绿素提取和含量计算方法,对不同处理称取等量实验材料并研磨至粉末,然后利用80%丙酮进行叶绿素萃取。通过UV2310Ⅱ分光光度计分别测定提取液波长在663、646、470 nm下的OD值。最后,根据相关计算公式得到不同实验处理的叶绿素含量。根据Bogorad[33]的方法对实验样品中胆色素原(Porphobilinogen, PBG)、尿卟啉原III (Uroporphyrinogen Ⅲ, UrogenIII)和粪卟啉III(Coproporphyrinogen Ⅲ, CoprogenⅢ)进行提取和含量测定。根据Lee等[34]的方法对实验样品的原卟啉IX (Protoporphyrin IX, Proto IX)、镁原卟啉IX(Mg-Protoporphyrin IX, Mg-Proto IX)和原叶绿素酸酯(Protochlorophyllide, Pchlide)进行提取和含量测定。每处理进行3个生物学重复测定,并利用Omiscshare tools (
http://omicshare.com/tools/ )对数据进行Student's paired t-test。 -
利用改良的SDS法[35]提取实验样品中的总RNA。利用NanoDrop 2000微量分光光度计(Thermo Scientific Inc., USA)对RNA的质量进行检测,OD260/280≥1.8、OD260/230≥1.8和浓度大于100 ng·μL−1的RNA被用于下一步的反转录。
-
利用EasyScript One Step gDNA Removal and cDNA Synthesis SuperMix qPCR(全式金,中国)反转录试剂合成RNA的cDNA第一链。从已发表的铁皮石斛基因组[31]中找与叶绿素合成相关的基因,利用NCBI的Primer-BLAST进行引物设计(表1)。以Dc18S作为内参基因对数据进行标准化。根据TaKaRa TB GreenTMPremixEx TaqTM试剂盒(TaKaRa, Japan)使用说明书设置LightCycler480 System(Roche, USA)的qRT-PCR运行程序进行基因的qRT-PCR定量。每个基因在每个处理中进行3个生物学重复测定,并利用Omiscshare tools (
http://omicshare.com/tools/ )对数据进行Student's paired t-test。表 1 叶绿素合成相关基因的qRT-PCR引物序列
Table 1. The primer sequences of chlorophyll biosynthesis related genes for qRT-PCR
目的基因
Gene引物名称
Primer name引物序列
Sequence(5‘-3’/0DcHEMB FP-HEMB GCTTACTAGACGTCCTCGTCG RP-HEMB AAGCAACCTGGCATAGCTCC DcHEMC FP-HEMC TCCCTTGTGAGAATCGGCAC RP-HEMC GCTCCGAGTGTGTTGCCATA DcHEME FP-HEME CCTATGGCGAGCGAACAGAG RP-HEME CAGAGAACCCACTTCTCGGC DcHEMF FP-HEMF ACGCTCACTCCTCTCAATGC RP-HEMF TGTATCAGGGCGCTCGTTTT DcHEMG FP-HEMG GGCCCTAATAGCTTCCAGCC RP-HEMG GGCCCTAATAGCTTCCAGCC DcCHLD FP-CHLD GGGATCGCCATTTCGGGTAA RP-CHLD GGACGGGTCTGCATTTGCTA DcCHLH FP-CHLH CTTCGAGCTCGTCGGATACC RP-CHLH ATGAAAATGTTGGCGGTGGC DcCHLM FP-CHLM GTCGTCCGCGAGTACTTCAA RP-CHLM CTTGTTGACGCCTTCGGTTG Dc18S FP-18S CCAGGTCCAGACATAGTAAG RP-18S GTACAAAGGGCAGGGACGTA -
利用SPSS 13.0与Excel 2010对生理指标进行统计和方差分析,基因定量采用Comparative CT (−ΔΔCt) 法进行计算[36]。
-
正常的铁皮石斛叶片为全绿色(图1a),而叶艺铁皮石斛的叶片上则出现了大面积的白色条纹或斑块(图1b、c)。对叶片横切面的显微观察结果显示:正常铁皮石斛(WT)和叶艺铁皮石斛叶片绿色组织(YYG)的部位中充满了绿色细胞(图1d、e),而叶艺铁皮石斛叶艺白色组织(YYW)的部位中则基本上看不到绿色细胞(图1f)。对这些组织细胞的超显微结构观察显示:WT和YYG的叶绿体呈饱满的半球形,其体内富含排列整齐的基粒片层和类囊体基粒,淀粉颗粒少(图2);而YYW的叶绿体则呈干瘪的三角形,体内基粒片层和类囊体基粒数量稀疏且排列无规则,淀粉颗粒多(图2)。这些结果显示叶子呈绿色的WT和YYG富含大量的绿色叶绿体且具有相似的叶绿体细胞结构,而YYW则与前两者相反。

图 1 叶艺和正常铁皮石斛的形态特征和叶片显微观察
Figure 1. The morphological characteristics and microscopic structure of normal and mutant D. catenatum leaves

图 2 石斛叶片叶绿体的超显微结构观察
Figure 2. The ultrastructure of chloroplast in D. catenatum leaves
-
叶绿素和类胡萝卜素含量测定显示:WT和YYG之间的叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量无明显差异,但它们都显著高于YYW(图3)。这一结果显示YYW的叶绿素合成受到了阻碍,并支持了其白化表型特征和显微观察YYW中几乎没有绿色细胞的结果。

图 3 野生型和突变型铁皮石斛叶片中光合色素的含量
Figure 3. Photosynthetic pigments content of WT and mutant leaves
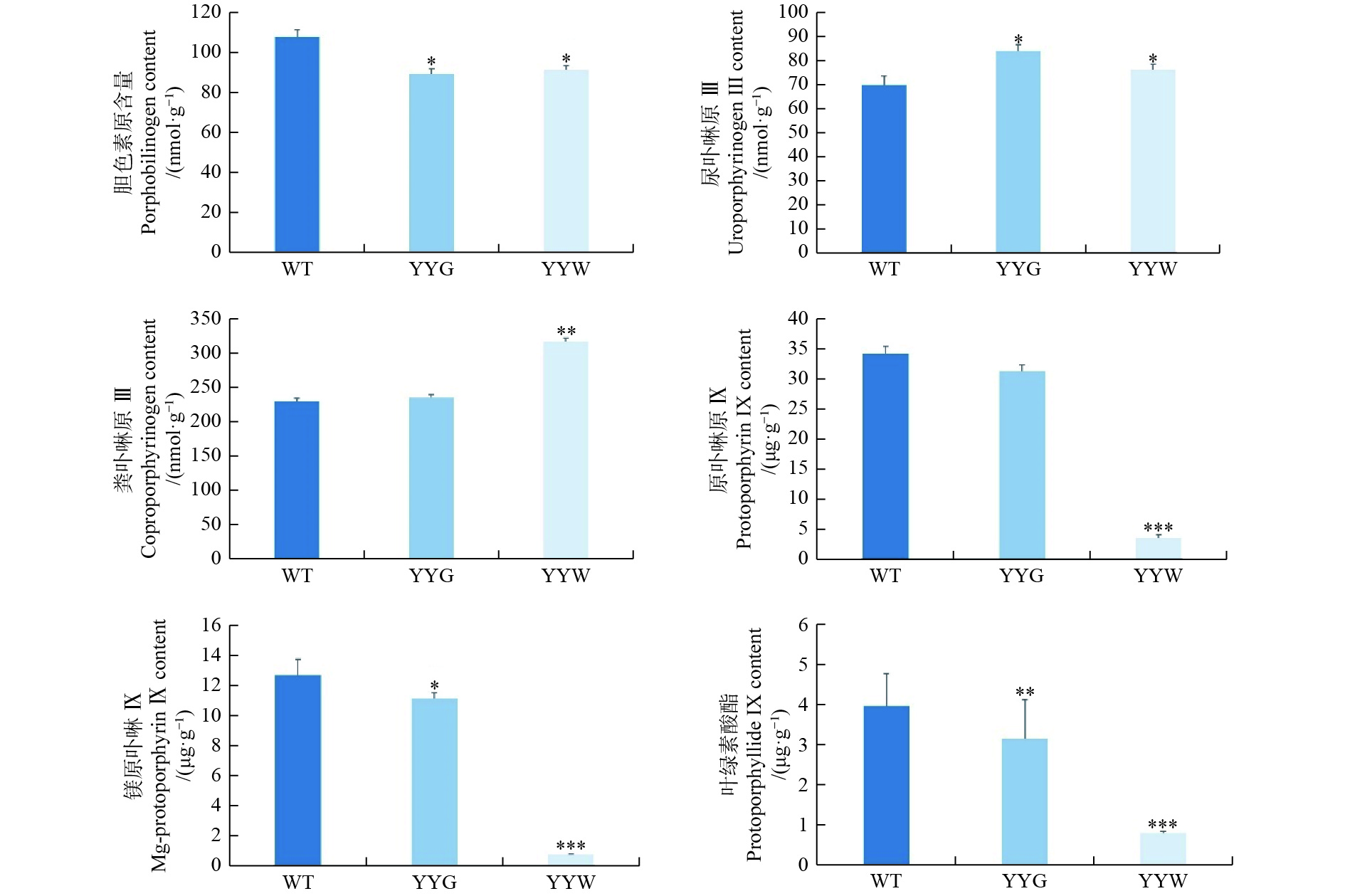
叶绿素合成的上游中间代谢物中,YYW的胆色素原含量是WT的84%,而尿卟啉原Ⅲ和粪卟啉原Ⅲ含量则分别是WT的109%和137%(图4)。在叶绿素合成下游中间前体物质中,WT和YYG的原卟啉IX、镁原卟啉IX和原叶绿素酸酯含量是YYW的4.9~17.1倍,而它们二者之间的差距相较于YYW则较小(图4)。粪卟啉原Ⅲ是原卟啉IX和镁原卟啉IX重要合成前体[15]。因此,这些结果显示:YYW在合成叶绿素的过程中,原卟啉IX的合成受到了阻碍,中间产物无法正常流向叶绿素合成。
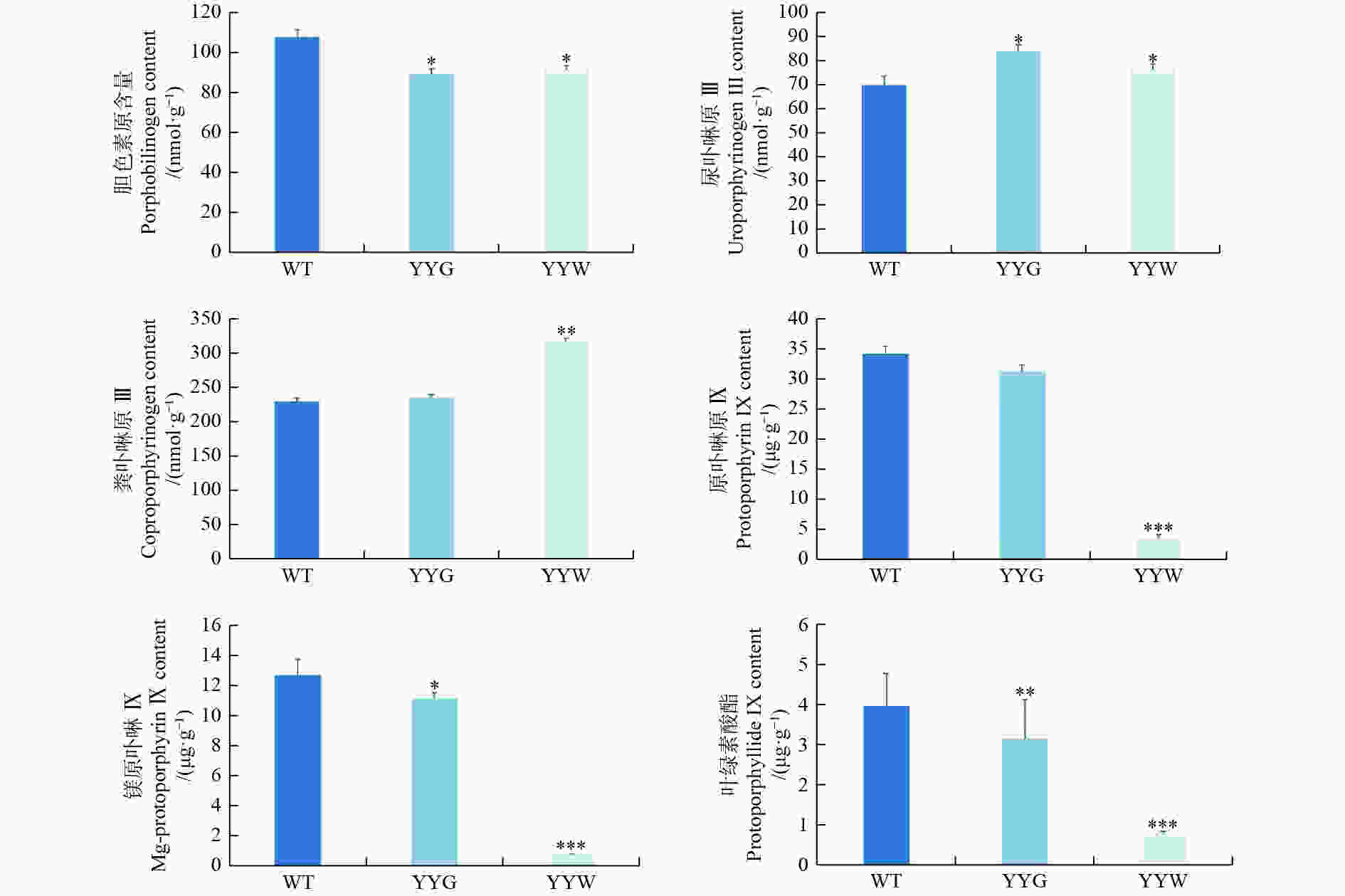
图 4 野生型和突变型铁皮石斛叶片中叶绿素合成的中间产物含量
Figure 4. The content of chlorophyll intermediates in WT and mutant leaves
-
WT、YYG和YYW中叶绿素生物合成途径中8个结构基因的qRT-PCR结果(图5)显示:YYW中的尿卟啉原脱羧酶(Uroporphyrinogen decarboxylase, DcHEME)、粪卟啉原氧化脱羧酶(Coproporphyrinogen III oxidase, DcHEMF)、原卟啉原氧化酶(Protoporphyrinogen oxidase, DcHEMG)和镁螯合酶H和镁螯合酶H亚基(Mg chelatase H subunit, DcCHLH)的表达量都显著低于WT和YYG。羟甲基后胆色素原合酶(Porphobilinogen deaminase, DcHEMC)的表达量在三者之间并无明显差异,而YYG和YYW中的胆色素原合酶(Porphobilinogen Synthase, DcHEMB)、镁螯合酶D亚基(Mg chelatase D subunit, DcCHLD)和镁原卟啉IX甲基转移酶(Mg protoporphyrin IX methyltransferase, DcCHLM)的表达量则都显著低于WT。值得注意的是,HEMF和HEMG是合成原卟啉IX的关键酶,而CHLH和CHLD则是合成镁原卟啉IX的关键酶[15]。
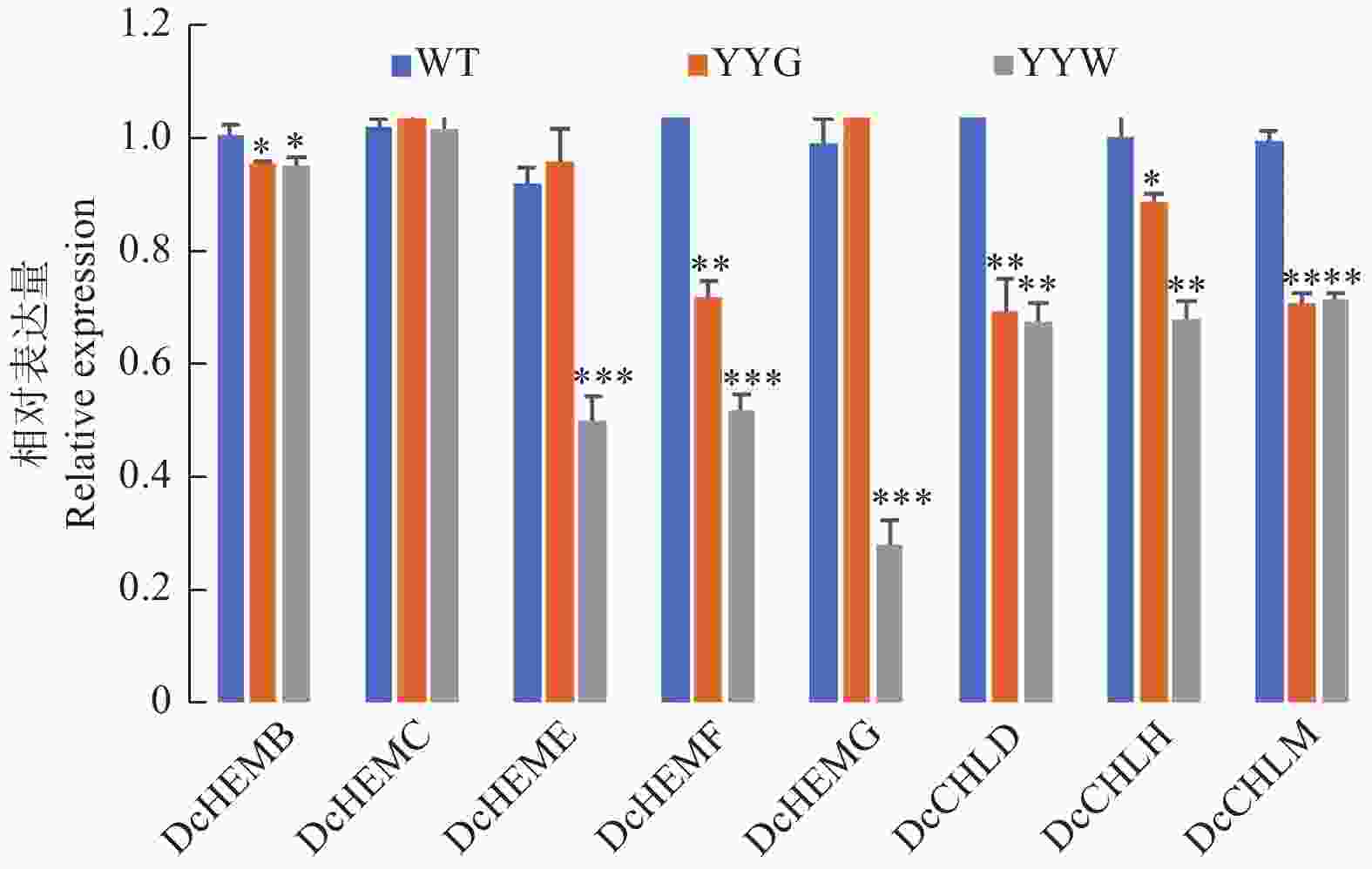
图 5 野生型和突变体石斛叶片中叶绿素合成相关基因的表达量变化
Figure 5. The relative expression of some chlorophyll biosynthesis relative genes in WT and mutant leaves
-
大多数植物的叶片为绿色,但许多植物都有叶片白化的突变体,而这种叶片出现白化的突变现象是许多观赏植物的重要观赏性状,具有很高的观赏价值[15]。叶绿素是植物进行光合作用的主要物质,同时它也是使植物组织呈现绿色的重要色素[37]。在许多植物的白化突变体中,其叶子的白化区域都出现了叶绿素积累量显著下降[13, 38-40]。本研究通过显微观察发现,YYW基本没有绿色细胞,同时其叶绿体细胞结构畸形且体内基粒片层和类囊体基粒数量稀疏且排列无规则。许多研究表明,植物叶色的突变都伴随着叶绿体细胞结构的改变和体内类囊体基粒的无序排列[11, 41-42]。因此,叶绿素含量的下降是导致叶艺铁皮石斛叶片出现白色条纹或斑块的主要原因。此外,粪卟啉原Ⅲ在YYW中的含量显著高于WT和YYG,而粪卟啉原Ⅲ的下游产物原卟啉IX、镁原卟啉IX和原叶绿素酸酯在WT和YYG中的含量是YYW的4.7~17.1倍。这进一步表明,YYW的叶绿素合成从粪卟啉原Ⅲ下游产物合成开始就受到了阻碍,而这可能是导致YYW中叶绿素无法正常合成的主要原因。
叶绿素生物合成途径中关键酶编码基因表达量的变化会直接影响叶绿素在植物组织的积累量,从而改变植物组织的颜色表型[15-17, 43]。本研究中,叶绿素合成途径中的DcHEMF、DcHEMG、DcCHLH和DcCHLM在YYW中的表达量显著低于正常叶片。在叶绿素合成途径中,粪卟啉原Ⅲ经过HEMF和HEMG的催化后会生成原卟啉,然后分别通过CHLD、CHLH和CHLI进一步形成镁原卟啉IX[17]。值得注意的是,叶艺铁皮石斛中粪卟啉原Ⅲ含量显著高于正常叶片,而原卟啉和镁原卟啉IX含量则显著低于正常叶片。前人研究表明,金鱼草(Antirrhinum majus)叶片的黄绿表型是由Tam3插入使ChlH基因失去活性所导致[44]。水稻(Oryza sativa)叶片的突变体中,由于T-DNA插入到line 9-07117的镁螯合酶大亚基ChlH中,使其无法正常转录翻译,导致水稻叶片失绿[18]。银杏(Ginkgo biloba)叶片的黄化突变体中,GbPPO、GbCHLD、GbCHLH和GbCHLI在黄化叶片中的表达量下调,同时其粪卟啉原Ⅲ含量显著高于正常叶片,而原卟啉IX和镁原卟啉IX的含量则相反[15]。本研究中,DcHEMF、DcHEMG和DcCHLH在YYW中的低表达使粪卟啉原Ⅲ无法正常流向叶绿素合成,造成叶绿素的积累量减少,最终导致叶片失绿。这些研究结果为今后石斛属的叶艺性状改良提供了理论依据和分子基础。
-
DcHEMF、DcHEMG和DcCHLH在叶艺铁皮石斛叶片白色区域中的低表达使粪卟啉原Ⅲ无法正常流向叶绿素合成,这导致了叶绿素含量的下降,是使铁皮石斛出现叶艺表型的主要原因。
铁皮石斛(Dendrobium catenatum)叶艺形成机制的初探
A Preliminary Study of Yeyi Mechanism in Dendrobium catenatum Leaves
-
摘要:
目的 探究铁皮石斛(Dendrobium catenatum)叶艺形成的分子机制,为培育叶艺兰花品种提供理论基础。 方法 以正常铁皮石斛(WT)、叶艺铁皮石斛叶片绿色组织(YYG)、叶艺铁皮石斛叶艺白色组织(YYW)为研究对象,通过显微结构观察、叶绿素及其前体物质含量的测定、叶绿素合成相关基因的qRT-PCR定量来探究叶艺形成的分子机制。 结果 YYW的总叶绿素含量明显低于WT和YYG,同时其叶绿体细胞结构出现了畸形现象。在叶绿素合成前体物质中,YYW中粪卟啉原Ⅲ的含量显著高于WT和YYG,而原卟啉IX、镁原卟啉IX和原叶绿素酸酯在WT和YYG中的含量则高于YYW。此外,qRT-PCR结果显示:粪卟啉原氧化脱羧酶(DcHEMF)、原卟啉原氧化酶(DcHEMG)、镁螯合酶H亚基(DcCHLH)在YYW中的表达量显著下调。 结论 YYW中DcHEMF、DcHEMG和DcCHLH表达量的下调使粪卟啉原Ⅲ无法正常流向叶绿素合成,这导致了其叶绿素含量的下降,是使铁皮石斛叶片出现了叶艺表型的主要原因。 Abstract:Objective To find the mechanism of Yeyi (a mutation of de-greening) of Dendrobium catenatum leaves and provide information for breeding novel orchid cultivar. Method The normal D. catenatum leaf (WT), the green parts of mutant leaf (YYG) and the white parts of mutant leaf (YYW) were used to study the molecular mechanism of Yeyi formation by observing the microstructure, the quantification of chlorophyll and its precursor, and the quantitative real-time PCR (qRT-PCR) of chlorophyll biosynthesis related genes . Result There was almost no green cell and the cellular structure of chloroplasts appeared abnormal in YYW. Among the precursors of chlorophyll, the coproporphyrinogen Ⅲ content in YYW was significant lower than that in WT and YYG, whereas the contents of protoporphyrin IX, Mg- Protoporphyrin IX and Protochlorophyllide in WT and YYG were higher than that in YYW. In addition, the qRT-PCR result suggested that the expressional levels of DcHEMF, DcHEMG, and DcCHLH were significantly down-regulated in YYW. Conclusion The down-regulated expression of DcHEMF, DcHEMG, and DcCHLH prevents coproporphyrinogen Ⅲ from flowing into chlorophyll biosynthesis in YYW, which will lead to the decrease of chlorophyll content and is the main reason for appearing the phenotype of Yeyi in D. catenatum. -
Key words:
- Yeyi
- / ornamental characteristic
- / Dendrobium catenatum
- / chlorophyll
- / gene expression
-
图 1 叶艺和正常铁皮石斛的形态特征和叶片显微观察
Figure 1. The morphological characteristics and microscopic structure of normal and mutant D. catenatum leaves
图 2 石斛叶片叶绿体的超显微结构观察
Figure 2. The ultrastructure of chloroplast in D. catenatum leaves
图 3 野生型和突变型铁皮石斛叶片中光合色素的含量
Figure 3. Photosynthetic pigments content of WT and mutant leaves
图 4 野生型和突变型铁皮石斛叶片中叶绿素合成的中间产物含量
Figure 4. The content of chlorophyll intermediates in WT and mutant leaves
图 5 野生型和突变体石斛叶片中叶绿素合成相关基因的表达量变化
Figure 5. The relative expression of some chlorophyll biosynthesis relative genes in WT and mutant leaves
表 1 叶绿素合成相关基因的qRT-PCR引物序列
Table 1. The primer sequences of chlorophyll biosynthesis related genes for qRT-PCR
目的基因
Gene引物名称
Primer name引物序列
Sequence(5‘-3’/0DcHEMB FP-HEMB GCTTACTAGACGTCCTCGTCG RP-HEMB AAGCAACCTGGCATAGCTCC DcHEMC FP-HEMC TCCCTTGTGAGAATCGGCAC RP-HEMC GCTCCGAGTGTGTTGCCATA DcHEME FP-HEME CCTATGGCGAGCGAACAGAG RP-HEME CAGAGAACCCACTTCTCGGC DcHEMF FP-HEMF ACGCTCACTCCTCTCAATGC RP-HEMF TGTATCAGGGCGCTCGTTTT DcHEMG FP-HEMG GGCCCTAATAGCTTCCAGCC RP-HEMG GGCCCTAATAGCTTCCAGCC DcCHLD FP-CHLD GGGATCGCCATTTCGGGTAA RP-CHLD GGACGGGTCTGCATTTGCTA DcCHLH FP-CHLH CTTCGAGCTCGTCGGATACC RP-CHLH ATGAAAATGTTGGCGGTGGC DcCHLM FP-CHLM GTCGTCCGCGAGTACTTCAA RP-CHLM CTTGTTGACGCCTTCGGTTG Dc18S FP-18S CCAGGTCCAGACATAGTAAG RP-18S GTACAAAGGGCAGGGACGTA  下载: 导出CSV
下载: 导出CSV
-
[1] 苗 晗, 顾兴芳, 张圣平, 等. 蔬菜叶色突变体研究进展[J]. 中国蔬菜, 2007, 1(6):39-42. doi: 10.3969/j.issn.1000-6346.2007.06.015 [2] 吕 明, 刘海衡, 毛虎德, 等. 芥菜型油菜黄化突变体叶片叶绿素合成代谢变化[J]. 西北植物学报, 2010, 30(11):2177-2183. [3] 袁丽钗, 李雪平, 彭镇华, 等. 菲白竹组培苗白化、绿化突变体的超微结构及15个叶绿体编码基因的表达[J]. 植物学报, 2010, 45(4):451-459. doi: 10.3969/j.issn.1674-3466.2010.04.008 [4] Deng X J, Zhang H Q, Wang Y,<italic> et al</italic>. Mapped clone and functional analysis of leaf-color gene <italic>Ygl7</italic> in a rice hybrid (<italic>Oryza sativa</italic> L. ssp. indica)[J]. PLoS One, 2014, 9(6): e99564. doi: 10.1371/journal.pone.0099564 [5] Waters B M, McInturf S A, Amundsen K. Transcriptomic and physiological characterization of the <italic>fefe</italic> mutant of melon (<italic>Cucumis melo</italic>) reveals new aspects of iron-copper crosstalk[J]. New Phytologist, 2014, 203(4): 1128-1145. doi: 10.1111/nph.12911 [6] 杨佳秀, 杜丽芬, 刘录祥, 等. 小麦旗叶黄化转绿突变体的生理分析及细胞学研究[J]. 西北植物学报, 2015, 35(12):2455-2461. doi: 10.7606/j.issn.1000-4025.2015.12.2455 [7] Li X, Kanakala S, He Y,<italic> et al</italic>. Physiological characterization and comparative transcriptome tnalysis of white and green leaves of <italic>Ananas comosus</italic> var. <italic>Bracteatus</italic>[J]. PLoS One, 2017, 12(1): e0169838. doi: 10.1371/journal.pone.0169838 [8] 何 冰, 刘玲珑, 张文伟, 等. 植物叶色突变体[J]. 植物生理学通讯, 2006, 42(1):1-9. [9] 夏宜平, 李钱鱼, 常 乐, 等. 玉簪属(<italic>Hosta</italic>)植物的花叶嵌合体特性研究[J]. 浙江大学学报:农业与生命科学版, 2008, 34(2):193-199. [10] 王彩霞, 田韦韦, 田 敏, 等. 文心兰黄化突变体的初步研究[J]. 核农学报, 2013, 27(12):1845-1852. doi: 10.11869/hnxb.2013.12.1845 [11] Yang Y X, Chen X X, Xu B,<italic> et al</italic>. Phenotype and transcriptome analysis reveals chloroplast development and pigment biosynthesis together influenced the leaf color formation in mutants of <italic>Anthurium andraeanum</italic> ‘Sonate’[J]. Frontiers in Plant Science, 2015, 6: e139. [12] Beale S I. Green genes gleaned[J]. Trends in plant science, 2005, 10(7): 309-312. doi: 10.1016/j.tplants.2005.05.005 [13] Wu Z, Zhang X, He B,<italic> et al</italic>. A Chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis[J]. Plant Physiology, 2007, 145(1): 29-40. doi: 10.1104/pp.107.100321 [14] Wang L, Yue C, Cao H,<italic> et al</italic>. Biochemical and transcriptome analyses of a novel chlorophyll-deficient chlorina tea plant cultivar[J]. BMC Plant Biology, 2014, 14(1): 352-365. doi: 10.1186/s12870-014-0352-x [15] Li W X, Yang S B, Lu Z G,<italic> et al</italic>. Cytological, physiological, and transcriptomic analyses of golden leaf coloration in <italic>Ginkgo biloba</italic> L.[J]. Horticulture Research, 2018, 5: 12.DOI:10.1038/s41438-018-0015-4. [16] Adhikari N D, Froehlich J E, Strand D D,<italic> et al</italic>. GUN4-porphyrin complexes bind the ChlH/GUN5 subunit of Mg-Chelatase and promote chlorophyll biosynthesis in <italic>Arabidopsis</italic>[J]. Plant Cell, 2011, 23(4): 1449-1467. doi: 10.1105/tpc.110.082503 [17] Nagata N, Tanaka R, Satoh S,<italic> et al</italic>. Identification of a vinyl reductase gene for chlorophyll synthesis in <italic>Arabidopsis thaliana</italic> and implications for the evolution of <italic>Prochlorococcusspecies</italic>[J]. Plant Cell, 2005, 17(1): 233-240. doi: 10.1105/tpc.104.027276 [18] Jung K H, Hur J, Ryu C H,<italic> et al</italic>. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system[J]. Plant Cell Physiology, 2003, 44(5): 463-472. doi: 10.1093/pcp/pcg064 [19] Kim C, Apel K. Substrate-dependent and organ-specific chloroplast protein import in planta[J]. Plant Cell, 2004, 16(1): 88-98. doi: 10.1105/tpc.015008 [20] Zhu G, Yang F, Shi S,<italic> et al</italic>. Transcriptome characterization of <italic>Cymbidium sinense</italic> ‘Dharma’ Using 454 pyrosequencing and its application in the identification of genes associated with leaf color variation[J]. PLoS One, 2015, 10(6): e0128592. doi: 10.1371/journal.pone.0128592 [21] 刘仲健. 中国兰花色叶艺研究及色叶复合艺名品鉴赏[M]. 北京: 中国林业出版, 2002: 53. [22] Yukawa T, Stern W L. Comparative vegetative anatomy and systematics of <italic>Cymbidium</italic> (Cymbidieae: Orchidaceae)[J]. Botanical Journal of the Linnean Society, 2002, 138(4): 383-419. doi: 10.1046/j.1095-8339.2002.00038.x [23] Zhang J, Wu K, Zeng S,<italic> et al</italic>. Transcriptome analysis of <italic>Cymbidium sinense</italic> and its application to the identification of genes associated with floral development[J]. BMC Genomics, 2013, 14(1): 1-17. doi: 10.1186/1471-2164-14-1 [24] Li Y H, Imai K, Ohno H,<italic> et al</italic>. Effects of acclimatization temperatures on antioxidant enzyme activities in mericlones of a <italic>Cattleya</italic> hybrid[J]. Engei Gakkai zasshi, 2004, 73(4): 386-392. doi: 10.2503/jjshs.73.386 [25] 范燕萍, 李慧玲, 李浩建. 几种花叶线艺兰叶片色斑色素组成和叶绿体超微结构研究[J]. 华南农业大学学报, 2006, 27(2):8-12. doi: 10.3969/j.issn.1001-411X.2006.02.003 [26] 蒋 彧, 何俊蓉, 熊剑锐, 等. 中国兰叶色突变体生理生化分析[J]. 北方园艺, 2015, 39(7):65-68. [27] 苏 畅, 李枝林, 付永恒. 叶艺兰辐射突变体的气孔特征与染色体核型分析[J]. 西部林业科学, 2016, 45(6):89-94. [28] 王 雁, 周进昌, 郑宝强, 等. 石斛兰[M]. 北京: 中国林业出版社, 2015: 20–30. [29] 蒋 彧, 陶 炼, 何俊蓉. 兰属春剑叶艺突变体叶片结构的研究[J]. 植物科学学报, 2018, 36(1):112-118. doi: 10.11913/PSJ.2095-0837.2018.10112 [30] 陈少宏. 线艺兰初探-试论国兰的遗传变异与选育之三[J]. 广东园林, 2002(3):45-47,44. doi: 10.3969/j.issn.1671-2641.2002.03.016 [31] Zhang G Q, Xu Q, Bian C,<italic> et al</italic>. The <italic>Dendrobium catenatum</italic> Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution[J]. Scientific Reports, 2016, 6(19029): 19029. [32] 邵 勤, 于泽源, 李兴国, 等. 叶色黄化突变体甜瓜叶片内部生理生化变化的研究[J]. 中国蔬菜, 2013(14):59-65. doi: 10.3969/j.issn.1000-6346.2013.14.012 [33] Bogorad L. Methods in enrymology [M]. New York: Academic Press, 1962: 885-891. [34] Lee H J, Mark D B, Parham R,<italic> et al</italic>. Chloroplast biogenesis 65: enzymic conversion of protoporphyrin Ⅸ to Mg-protoporphyrin Ⅸ in a subplastidic membrane fraction of cucumber etiochloroplasts[J]. Plant Physiology, 1992, 99(3): 1134-1140. doi: 10.1104/pp.99.3.1134 [35] 黄昕蕾. 基于转录组测序的鼓槌石斛花色花香形成分子调控机理研究[D]. 北京: 中国林业科学研究院, 2018. [36] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2<sup>-ΔΔCT </sup>method[J]. Methods, 2001, 25(4): 402-408. doi: 10.1006/meth.2001.1262 [37] Ohmiya A, Hirashima M, Yagi M,<italic> et al</italic>. Identification of genes associated with chlorophyll accumulation in flower petals[J]. PLoS One, 2014, 9(12): e113738. doi: 10.1371/journal.pone.0113738 [38] 刘艳霞, 林冬枝, 董彦君. 水稻温敏感叶色突变体研究进展[J]. 中国水稻科学, 2015, 29(4):439-446. doi: 10.3969/j.issn.1001-7216.2015.04.014 [39] Li Y H, Wang B H, Dai Z Y,<italic> et al</italic>. Morphological structure and genetic mapping of new leaf-color mutant gene in rice (<italic>Oryza sativa</italic>)[J]. Rice Science, 2012, 19(2): 79-85. doi: 10.1016/S1672-6308(12)60025-0 [40] Maekawa S, Takabayashi A, Reyes T H,<italic> et al</italic>. Pale-green phenotype of <italic>atl31</italic> <italic>atl6</italic>double mutant leaves is caused by disruption of 5-aminolevulinic acid biosynthesis in <italic>Arabidopsis thaliana</italic>[J]. PLoS One, 2015, 10(2): e0117662. doi: 10.1371/journal.pone.0117662 [41] Yoo S C, Cho S H, Sugimoto H,<italic> et al</italic>. Rice virescent3 and stripe1encoding the large and small subunits of ribonucleotide reductase are required for chloroplast biogenesis during early leaf development[J]. Plant Physiology, 2009, 150(1): 388-401. doi: 10.1104/pp.109.136648 [42] Yang H Y, Xia X W, Fang W,<italic> et al</italic>. Identification of genes involved in spontaneous leaf color variation in <italic>Pseudosasajaponica</italic>[J]. Genetics and Molecular Research, 2015, 14(4): 11827-11840. doi: 10.4238/2015.October.2.16 [43] Zhang H, Li J, Yoo J,<italic> et al</italic>. Rice chlorine-1 and chlorine-9 encode <italic>Ch1D</italic> and <italic>CH1I</italic> subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development[J]. Plant Molecular Biology, 2006, 62(3): 325-337. doi: 10.1007/s11103-006-9024-z [44] Hudson A, Carpenter R, Doyle S,<italic> et al</italic>. Olive: a key gene required for chlorophyll biosynthesis in <italic>Antirrhinum majus</italic>[J]. The EMBO Journal, 1993, 12(10): 3711-3719. doi: 10.1002/j.1460-2075.1993.tb06048.x -

 点击查看大图
点击查看大图

计量
- 文章访问数: 5242
- HTML全文浏览量: 2667
- PDF下载量: 54
- 被引次数: 0
