


-
在全球气候变化的大背景下,森林作为陆地上最大的生态系统,其吸收、储存二氧化碳的机制和过程被广泛认为是最有效的固碳方式,其碳汇作用受到越来越多国家和地区的重视[1-3]。迄今为止,国内外已建立了各种形式的生物量模型[4-6]。已建立的生物量模型大多采用基于幂函数形式的异速生长方程[7-9]。异速模型中作为解释变量的胸径和树高等测树因子,其本身就是生长的结果,所建模型虽精度较高,但模型的建立与建模所选样本的范围和区域有关,缺乏一定的生物学意义[10-11],且在实际应用中,胸径和树高等测树因子往往需要样木的野外复测,再代入到相应的异速模型中,费时费力。
近年来,越来越多的学者将时间变量考虑到方程中,构建了含有年龄变量的异速生长方程。如薛春泉等[12]基于樟树、木荷和枫香3个广东省主要乡土阔叶树种的生物量实测数据,构建了含有年龄和胸径的单木生物量异速模型,同时还考虑了单木干材、干皮、树枝和树叶与地上部分总量相兼容的问题。随后,薛春泉等[13]又基于Schumacher,Chapman-Richards, Logistic和Korf等4种理论生长方程构建了不同起源(天然林和人工林)的地上与地下部分单木生物量模型,并通过联立方程组控制总量的方法解决了地上部分各组分生物量的相容性问题。考虑时间因素的异速模型生物量始终保持幂指数生长的趋势,生物量随年龄变化生长的生物学意义难以体现,且模型仅局限在林龄已知的人工林和碳汇造林的生物量估计,模型的应用和推广有一定的限制。具有生物学意义的理论生长方程在森林生长与收获中应用较广[14-16]。理论生长方程多用来描述胸径和树高等测树因子的生长趋势[17-19],很少用来描述生物量与时间的理论生长关系。由于观测数据获取的困难性和立地条件的复杂性等,使得关于生物量生长模型的研究鲜有报道,尤其是区域尺度生物量生长模型的相关研究。
广东省的森林类型主要以常绿阔叶林为主,主要包括栎类(Quercus spp.)和木荷(Schima superba Gardn. et Champ.)等阔叶树种,蓄积量大,分布范围广,是广东省碳汇造林的主要树种。本研究基于广东省1997至2017年的森林资源连续清查数据,筛选有五期连续生长30株以上保留木的固定样地30个(共计1 412株样木),构建了基础生物量生长模型、含有林分特征的生物量生长模型、含有林分特征和立地条件的生长模型等不同形式的林分生物量生长模型,以期为广东省区域尺度森林生物量和碳储量的估算提供模型和方法,提高区域尺度森林生物量和碳储量估算的准确性,也为生物量生长模型在其他区域的应用提供参考。
-
30个固定样地的样木数据来源于广东省森林资源连续清查数据,固定样地面积约为0.067 hm2,植被类型属于常绿阔叶林,包括栎类和木荷等常绿阔叶树种,样地林分起源全部为天然林,地类为乔木林地,样地海拔高度在300 ~ 1 300 m之间,地形主要以低山丘陵为主,土壤质地以壤土和砂壤土为主。30个固定样地在广东省的分布如图1所示。

图 1 30个样地在广东省的分布
Figure 1. Distribution of 30 fixed plots in Guangdong province
-
表1给出了30个固定样地五期相应的林分统计量,包括林分平均胸径Dg(定义为均方直径)、林分胸高断面积(Ba)以及林分密度指数(SDI)[20]等。样木类型包括进界木和保留木等。表2中列出了30个样地建模样本的株数和林分平均胸径(均方直径)的统计量。
表 1 30个固定样地不同年份的样木数据统计量
Table 1. Descriptive statistics of sample trees in 30 permanent plots at different years
变量 Variable 时间 Year 平均值 Mean 标准差 SD 最小值 Min. 最大值 Max 样木数量
No. of tree1997 104 30.46 58 175 2002 122 33.15 44 196 2007 126 38.41 53 193 2012 122 35.51 64 191 2017 124 42.21 62 231 林分平均胸径Dg
Quadratic mean diameter at breast height/cm1997 11.58 2.63 7.72 18.24 2002 12.55 2.56 9.51 18.60 2007 13.23 2.68 9.72 20.35 2012 14.04 2.61 10.59 20.25 2017 14.84 2.95 10.04 22.01 林分胸高断面积BA
Stand basal area/(m2·hm−2)1997 16.79 7.81 5.22 38.02 2002 22.01 7.01 8.21 36.12 2007 24.83 6.82 11.65 38.69 2012 27.42 7.46 15.01 42.04 2017 30.38 7.93 17.18 44.53 林分密度指数 SDI
Stand density index1997 43.34 17.00 15.71 83.67 2002 55.72 14.38 20.93 81.56 2007 61.90 14.27 28.74 84.50 2012 66.74 15.63 36.57 92.25 2017 72.47 16.40 40.49 106.67 表 2 建模样本数量和林分平均胸径统计量
Table 2. Descriptive statistics of number and stand diameter at breast height of sample treesfor model data
变量 Variable 时间 Year 平均值 Mean 标准差 SD 最小值 Min. 最大值 Max 样木数 No. of tree ─ 47 13.13 31 82 林分平均胸径
Stand diameter /cm1997 11.79 3.12 7.68 20.02 2002 14.14 3.24 9.62 22.17 2007 15.56 3.27 10.68 23.13 2012 16.88 3.39 11.55 24.00 2017 18.38 3.62 12.19 25.77 -
同一时间序列下,对于保留木来说,只需确定第一期样地的林分平均年龄,再加上相应时间序列的时间间隔,即可推算出每期每块样地的林分平均年龄。根据相应时间序列首期和末期样地平均胸径的平均生长速率
$ \overline{v} $ ,每块样地第一期(1997)林分平均年龄t按以下公式确定:$ \overline{v}={(D}_{g}^{\left(p\right)}-{D}_{g}^{\left(q\right)})/\Delta t $
(1) $ t=\sum\nolimits_{i=1}^{k}\left(\frac{{D}_{gi}}{\overline{v}}·{n}_{i}\right)/\sum\nolimits_{i=1}^{k}{n}_{i} $
(2) 式中,
$ {D}_{g}^{\left(p\right)},{D}_{g}^{\left(q\right)} $ 分别是样地相应时间序列首期和末期的林分平均胸径,$ \Delta t $ 是相应的间隔时间,i=1, 2, 3…k代表第一期(1997)相应的检尺类型(进界木,保留木等),$ {D}_{gi} $ 代表第i类检尺类型的平均胸径,$ {n}_{i} $ 是第i类检尺类型的样木株数,t为样地的平均年龄,按每期5年的间隔时间依次确定建模样本每期每块样地的林分平均年龄。 -
我国已发布的13个常见树种立木生物量模型和碳计量参数的行业标准提供了不同区域单株立木生物量的异速模型,包括一元模型和二元模型以及地上、地下生物量模型[21-23],其他文献关于广东省区域内相应树种的立木生物量异速模型也均有报道[24]。
-
本研究选择Richards理论生长方程作为基础方程[25],模型结构如下:
$ Y=a{(1-{\rm{e}}^{-bt})}^{c} $
(3) 式中,Y为样地生物量,t为样地年龄,a, b, c为模型参数。
-
为解释不同林分密度状况对林分生物量生长的影响,刻画不同林分结构条件下生物量生长的差别,基于Richards基础方程,将方程中相关的参数描述为与林分平均胸高断面积和林分密度指数相关的参数方程,将式(3)扩展成含有林分特征的生物量生长模型,模型形式如下:
$ Y=a·{\left(Ba\right)}^{d}{(1-{\rm{e}}^{-k·{\left(SDI\right)}^{b}t})}^{c} $
(4) 式中,Ba为林分胸高断面积(m2·hm−2),SDI为林分密度指数,k为常数,其他参数意义同式(3)。
-
式(3)中参数a和c是生长曲线的最大值和相应的形状,分别对参数a和c进行有序样本聚类,将30块样地依次划分为3种类别,以此将区域内不同林分立地状况分为3类,代表3种不同立地等级和生产力(优、良和差),依次求得每个样地参数a和c的所属类别,将相应的哑变量加入到式(4)中,得到含有林分特征和立地条件的分级生物量生长模型,其形式如下[26-27]:
$ Y=({a}_{1}+{a}_{2}{S}_{11}+{a}_{3}{S}_{12})·{\left(Ba\right)}^{d}{(1-{\rm{e}}^{-k·{\left(SDI\right)}^{b}t})}^{({c}_{1}+{c}_{2}{S}_{21}+{c}_{3}{S}_{22})} $
(5) 式中,
$ {S}_{11} $ ,$ {S}_{12} $ 是参数a的哑变量,$ {S}_{21} $ ,$ {S}_{22} $ 是参数c的哑变量。a1, a2, a3, c1, c2, c3分别是相应的参数值。 -
采用决定系数(R2)、调整决定系数(Ra2),估计值的标准误(SEE),总相对误差(TRE)、平均系统误差(ASE)、平均百分标准误差(MPSE)、赤池信息准则AIC和BIC评价生物量生长模型的拟合效果,公式详见文献[28]。关于样本数据是否要划分为建模样本和检验样本的问题,仍具有争议[28-31]。本研究构建的林分生物量模型是以30个样地保留木数据为基础,为了充分利用30个样地林分生物量信息,不区分建模样本和检验样本[28, 32-33],将30个样地生物量的建模样本作为检验样本对所建的模型进行检验和评价。
-
表3给出了3种不同形式生物量生长模型拟合结果的统计指标。基础生物量生长模型的调整决定系数Ra2不到0.48,平均相对误差(MPSE)高达37.40%,AIC和BIC值也均达到了1 000以上,拟合效果最差。将相应林分特征加入基础生物量生长模型后,决定系数R2提高到了0.836,估计值的标准差(SEE)下降到了36.92 kg,总相对误差(TRE)、平均系统误差(ASE)、平均百分标准误差(MPSE)和平均预估误差(MPE)分别下降了0.93%,4.17%,16.36%和3.25%,AIC和BIC也均明显降低,拟合效果得到一定程度改善。含有林分特征的生长模型中通过哑变量反映不同的立地条件后,拟合效果得到进一步提高,决定系数R2达到了0.961,TRE、均ASE接近于0,SEE、MPSE、MPE、AIC以及BIC值均达到最小。
表 3 3种不同形式生物量生长模型拟合效果的评价统计指标
Table 3. Statistics indices of goodness-of-fit for three different forms of biomass growth function
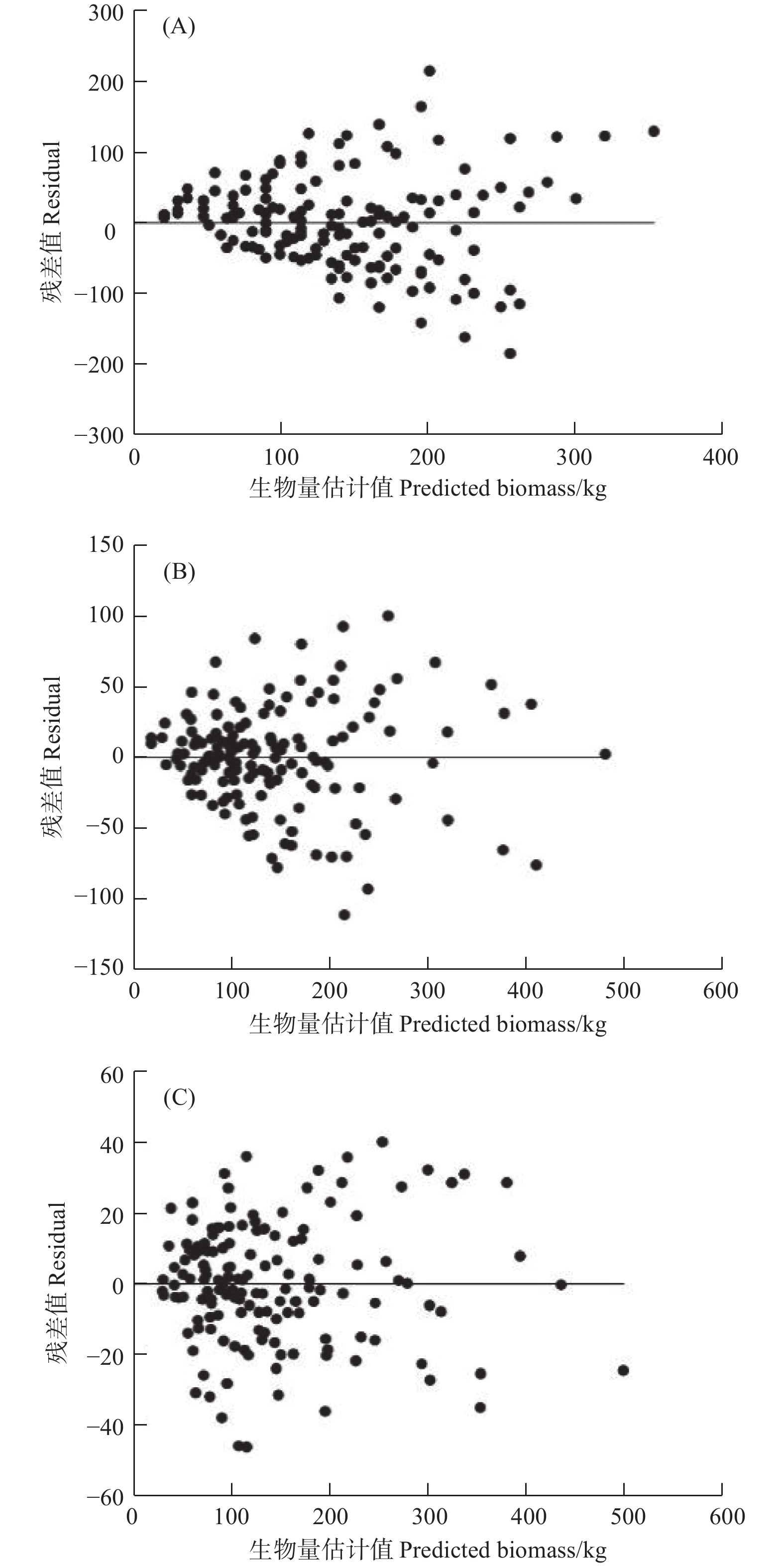
模型Model R2a SEE/kg TRE/% ASE/% MPSE/% MPE/% AIC BIC Eq. (3) 0.474 51 66.17 1.25 6.55 37.40 7.35 1 006.11 1 018.15 Eq. (4) 0.836 43 36.92 0.32 2.38 21.04 4.10 919.10 935.14 Eq. (5) 0.961 45 17.92 −0.03 0.17 11.31 1.99 812.60 844.68 图2给出了3种不同形式生物量生长模型的残差图,可以看到式(3)模型拟合结果的残差(图2(A))表现出明显的“喇叭状”规律,随着样地平均年龄的增大,生物量估计值的变动情况变大,呈现出明显的异方差性,拟合效果最差。式(4)模型拟合结果残差图(图2(B))“喇叭状”虽没有式(3)明显,但也呈现出一定程度的异方差性。式(5)模型拟合的残差图(图2(C))在x轴两侧的分布较为均匀,残差随样地年龄的增加变动幅度较小,无明显的异方差现象,拟合效果最优。
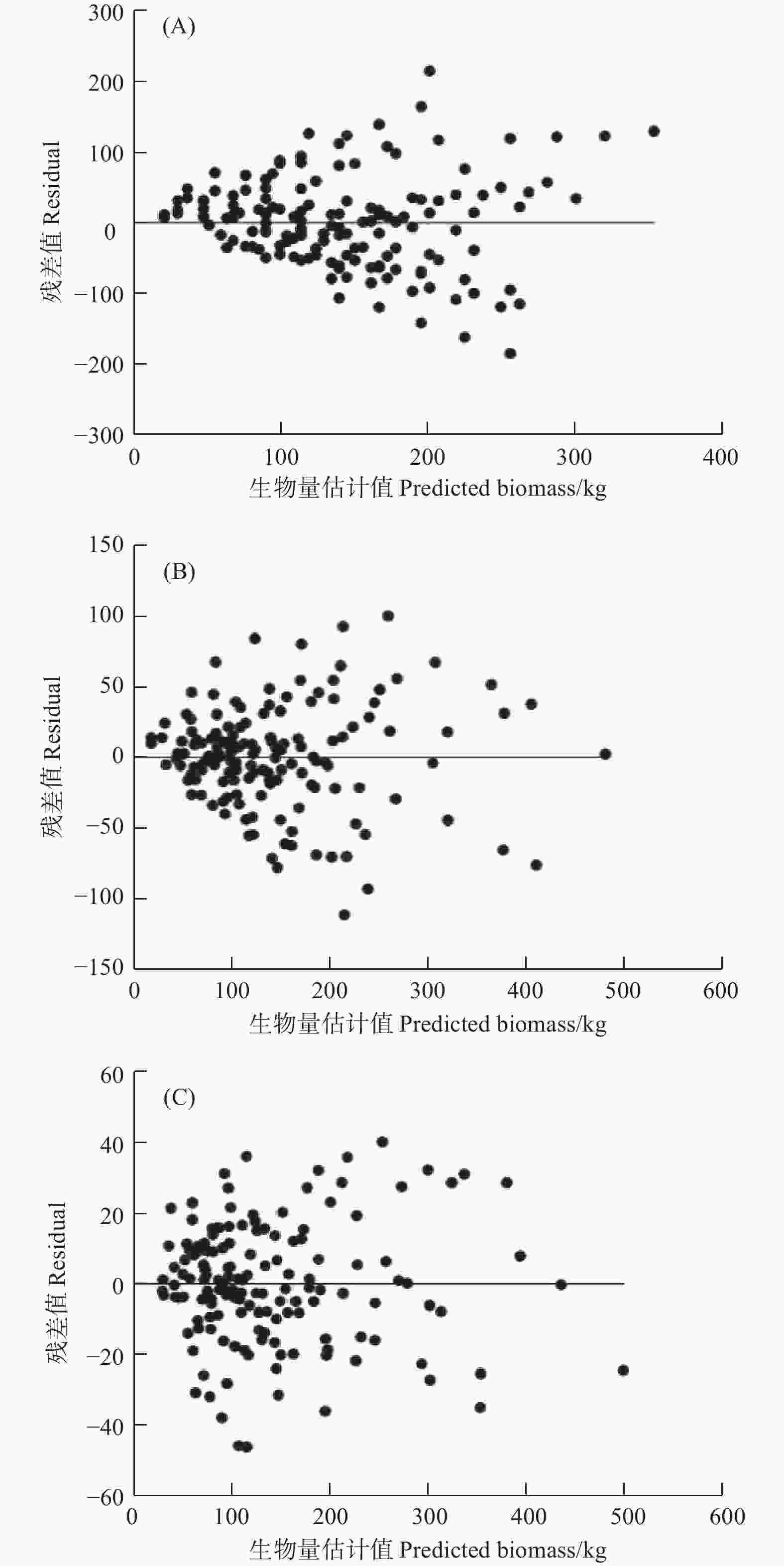
图 2 3种不同形式生物量生长模型的残差分布(A)、(B)和(C)表示式(3)、式(4)和式(5)
Figure 2. Residuals of three different forms of biomass growth functions (A), (B), (C) represents Eq. (3), Eq. (4) and Eq. (5)
图3给出了不同生长模型实际值与观测值之间的关系。式(5)模型拟合的估算值与实际值的坐标点均匀分布在坐标轴对角线两侧(图3(C)),模型的估算值与实际值相差较小,模型拟合效果最优。而式(3)与式(4)模型拟合的相应坐标点零散分布在坐标轴对角两侧(图3(A)、(B)),模型估算值与实际值相差较大,尤其是式(3)模型,明显高估了生物量较小的样地,而严重低估了生物量较大的样地,拟合效果最差。
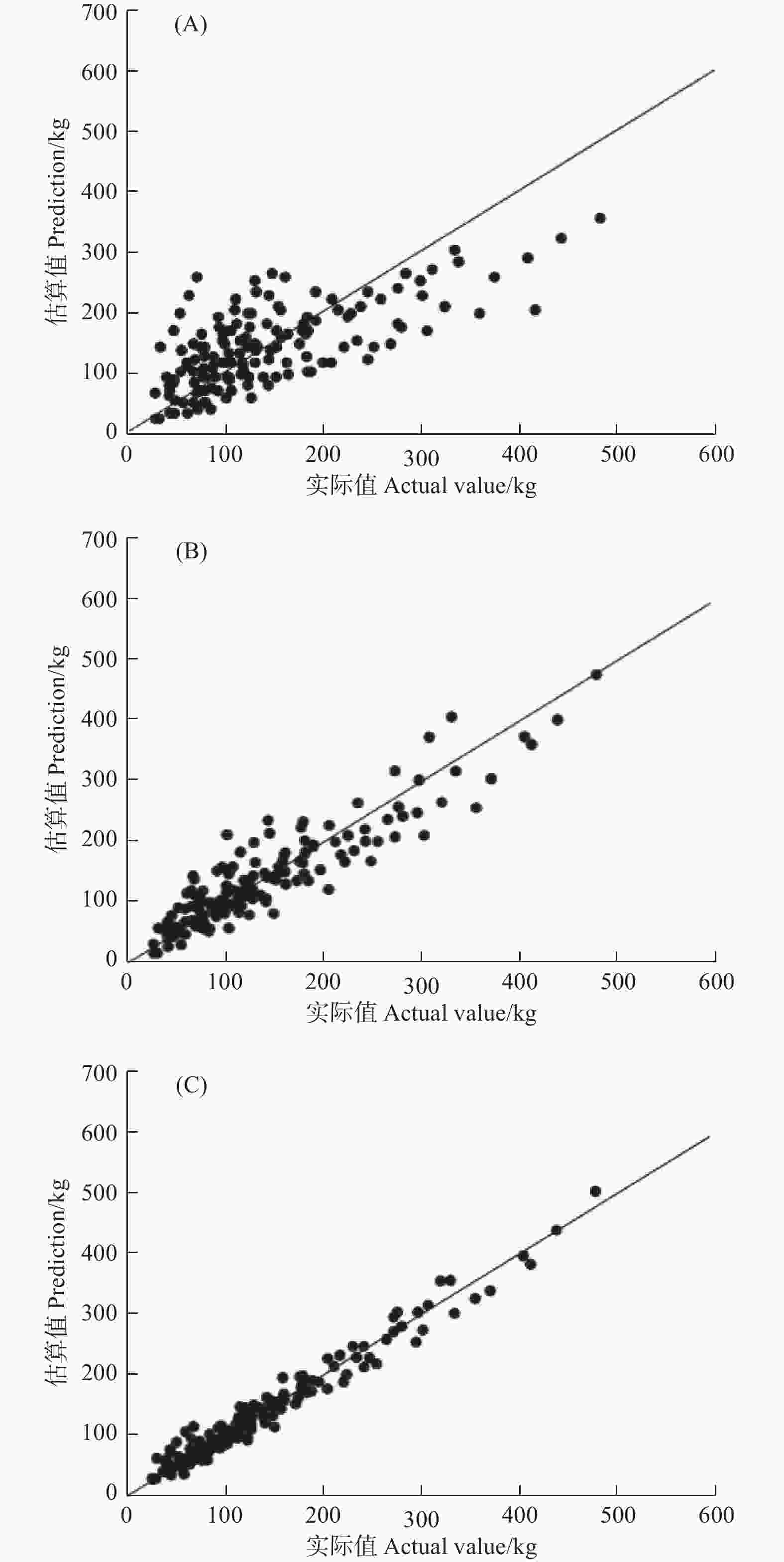
图 3 3种不同形式生物量生长模型实际值与估算值关系 (A)、(B)和(C)表示式(3)、式(4)和式(5)
Figure 3. Relationofactual value and prediction for three biomass growth models. (A), (B), (C) represents Eq. (3), Eq. (4) and Eq. (5)
基础生长模型仅仅考虑了林木自身的生长特性,仅以此建立的生物量生长方程不能很好地解释林分生物量的累积变化,拟合效果最差,且出现严重的异方差现象;林分胸高断面积和林分密度指数是广泛使用的林分密度指标,将相应林分密度指标体现在模型参数上,很好地解释了林分内林木的拥挤状况等林分特征对林木生长的影响,使得模型拟合效果得到了很大提高,一定程度上改善了模型的异方差现象;另外,由于林分所处地理环境的差异,林分本身供给林木生长的养分不同,即不同的立地条件下,林分具有不同的立地生产力。基于哑变量方法,通过划分不同立地等级,在模型参数上体现了不同立地的生产力,解释了由于立地生产力的不同而导致林分生长变化的影响,大大提高了生长模型的拟合精度(表4)。
表 4 最优生物量生长模型参数估计值的统计量
Table 4. Statistics indices of parameter estimate for the optimal biomass growth model
参数 Parameter 估计值 Estimate 渐进标准差 SD 变异系数 CV/ % a1 458.819 840 122.642 30 26.73 a2 590.385 661 152.798 11 25.88 a3 761.193 906 203.097 68 26.68 b −1.084 128 0.221 10 −20.39 c1 1.252 284 0.071 39 5.71 c2 1.287 937 0.077 12 5.99 c3 1.354 630 0.092 47 6.83 d 1.442 239 0.224 53 15.57 -
单木生物量的估算一般基于异速生长方程,对样地所有单木进行汇总或者取标准木的方法,得到林分水平上的生物量[34]。而区域尺度上生物量的估算一般基于生物量与材积的比值,包括IPCC法,转换因子连续函数法和加权生物量回归模型法[35-36],但建模过程复杂,需要大规模重复的野外测量数据。本研究基于Richards理论生长方程,通过构建含有林分年龄的生物量生长模型,从而实现了对特定时间下区域尺度上的生物量预估,可用于区域尺度生物量和碳储量的估算和碳汇潜力的评估。
生物量理论生长方程通过建立区域尺度生物量与时间(年龄)之间的关系,反映了区域尺度下生物量生长的变化趋势,可用于估算出某个年份下区域的森林生物量。区域尺度上林分基准年龄的确定是关键。龙时胜等[36]通过设置首期不同年龄采集样地内各树种的直径与年龄坐标信息,并基于树高生长至1.3 m处的年龄,采用树高生长方程准确定位,从而估算异龄林林分年龄。随后,龙时胜等[37]又根据相同直径的优势木和被压木的关系,通过年龄变化范围内的随机模拟,估计初始阶段的立木年龄推算林分年龄。本研究确定初始阶段林分年龄与龙时胜等的方法类似。
Richards基础生长模型不能很好的解释生物量生长过程,需要考虑林分特征和立地等因素解释其生物量的生长变化。林分密度是影响林分生长的重要因素之一,也是营林工作中能够有效控制的因子。慧淑荣等[38]在Richards生长方程的基础上,将立地指数和林分密度引入到方程中,构建了日本落叶松林分胸径、树高、断面积和蓄积的理论生长方程,精度检验和估测效果均较好;Sharma等[19]分龄级和森林类型建立了包含林分密度,林分胸高断面积以及地位指数等林分密度指标在内的北美短叶松和黑云杉胸径-树高Richards理论生长方程。
混合效应和哑变量模型兼顾了不同区域的现实条件,是考虑到了不同情况相互兼容的两种建模方法。但有研究表明,利用哑变量模型的拟合效果优于混合效应模型[39]。本研究针对不同的立地条件,通过构建哑变量反映不同样地类别的划分,兼顾到了不同样地之间的差异。这种差异可能来源于不同的立地因子如坡度、坡向和海拔等,立地因子反映了立地生产力的不同,在模型上主要体现在相应哑变量参数值的不同。理论生长方程中参数a是方程的最大值,一般认为与立地条件和生产力有着密切关系。划分的3种不同立地代表了30个样地的3种不同立地状况,同时也体现了3种不同类别立地潜在生产力。划分不同立地等级,同时考虑林分特征变量,兼顾了不同立地的潜在生产力,拟合精度较高,预估效果较好,可满足区域尺度森林生物量和碳储量的估算要求,另一方面,林分密度变量可以为森林经营管理者提供控制林分密度的参考,为合理评价区域碳汇潜力和提高固碳速率提供一定借鉴。
应用在区域尺度下林分生物量模型,样本单元数至关重要,大尺度范围应用的立木生物量模型,样本单元数的增加可以提高模型的准确度和通用性[6]。本研究仅选取了广东省森林资源清查30个固定样地数据进行建模,在样本单元数上仅仅满足了区域大样本的要求[6],为得到更准确的生物量模型,需要覆盖全省范围尽可能多的固定样地数据。
-
本研究基于五期(1997, 2002, 2007, 2012, 2017)广东省森林资源连续清查资料,通过30个固定样地五期保留木数据,采用Richards理论生长方程,分别构建和比较了基础生长方程、含有林分特征的生长方程和含有林分特征和立地条件的生长方程等不同形式的生物量生长模型。在林分生物量生长模型中,引入林分胸高断面积和林分密度指数等指标以及反映不同立地生产力差异的哑变量,极大地改善了模型的拟合效果,提升了模型预测能力,为广东省区域尺度常绿阔叶林生物量和碳储量估算提供了借鉴和参考。
广东省常绿阔叶林生物量生长模型
Biomass Growth Models for Evergreen Broad-leaved Forests in Guangdong
-
摘要:
目的 研究林分生物量生长模型,以期估算区域尺度上的森林生物量。 方法 利用广东省五期森林资源连续清查数据(1997, 2002, 2007, 2012, 2017),以30个固定样地为研究对象(每块样地均有30株以上五期保留木,共计1 412株样木)。以Richards理论生长方程为基础,分别构建了基础生物量生长模型、含有林分特征的生物量生长模型、含有林分特征和立地条件的生长模型等不同形式的林分生物量生长模型,比较和评价了不同林分生物量生长模型的拟合效果。 结果 基础生物量生长模型拟合效果最差,调整决定系数Ra2仅为0.475;将林分密度指标引入基础生物量生长模型后,拟合效果得到极大改善,Ra2提高到了0.836;将哑变量引入含有林分特征的生长方程进行立地条件划分后,Ra2达到0.961,拟合效果达到最优。 结论 含有林分特征的生物量生长方程一定程度反映了林分生物量生长与林分密度之间的关系,在此基础上划分样地类别进行参数分级,进一步提高了生物量生长模型的拟合精度,反映了不同立地条件下生产力之间的差别。 Abstract:Objective To establish stand biomass growth models and estimate the forest biomass on the regional scale so as to provide supports and references for carbon measurement and accounting. Method The forest inventory data of five phases in Guangdong province (1997, 2002, 2007, 2012, and 2017) were used to select thirty fixed plots (with more than thirty reserved sample trees for five phases in each plot, totally 1 412 trees). Based on Richards theoretical growth function, the basic biomass growth model, stand-character-based biomass growth model, and stand-character-based and site-condition-based biomass growth model were established respectively using dummy variables. The biomass growth models established for stand level were evaluated and compared in this study. Result The basic biomass growth model performed the worst with the lowest Ra2 of 0.475. When the stand density was added to the basic biomass growth model, the performance got improved greatly with the R2 of 0.836. When the dummy variable were added to the stand-character-based biomass growth function, the performance got best with the highest R2 of 0.961. Conclusion Based on the stand characters, the biomass growth function can reflect the relationship between biomass growth and stand density to some extents and got better goodness-of-fit. The introduction of the dummy variable to the stand-character-based biomass growth functions can improve the fitting accuracy further, indicating the difference of stand condition and production among different stand types. -
图 2 3种不同形式生物量生长模型的残差分布(A)、(B)和(C)表示式(3)、式(4)和式(5)
Figure 2. Residuals of three different forms of biomass growth functions (A), (B), (C) represents Eq. (3), Eq. (4) and Eq. (5)
图 3 3种不同形式生物量生长模型实际值与估算值关系 (A)、(B)和(C)表示式(3)、式(4)和式(5)
Figure 3. Relationofactual value and prediction for three biomass growth models. (A), (B), (C) represents Eq. (3), Eq. (4) and Eq. (5)
表 1 30个固定样地不同年份的样木数据统计量
Table 1. Descriptive statistics of sample trees in 30 permanent plots at different years
变量 Variable 时间 Year 平均值 Mean 标准差 SD 最小值 Min. 最大值 Max 样木数量
No. of tree1997 104 30.46 58 175 2002 122 33.15 44 196 2007 126 38.41 53 193 2012 122 35.51 64 191 2017 124 42.21 62 231 林分平均胸径Dg
Quadratic mean diameter at breast height/cm1997 11.58 2.63 7.72 18.24 2002 12.55 2.56 9.51 18.60 2007 13.23 2.68 9.72 20.35 2012 14.04 2.61 10.59 20.25 2017 14.84 2.95 10.04 22.01 林分胸高断面积BA
Stand basal area/(m2·hm−2)1997 16.79 7.81 5.22 38.02 2002 22.01 7.01 8.21 36.12 2007 24.83 6.82 11.65 38.69 2012 27.42 7.46 15.01 42.04 2017 30.38 7.93 17.18 44.53 林分密度指数 SDI
Stand density index1997 43.34 17.00 15.71 83.67 2002 55.72 14.38 20.93 81.56 2007 61.90 14.27 28.74 84.50 2012 66.74 15.63 36.57 92.25 2017 72.47 16.40 40.49 106.67  下载: 导出CSV
下载: 导出CSV
表 2 建模样本数量和林分平均胸径统计量
Table 2. Descriptive statistics of number and stand diameter at breast height of sample treesfor model data
变量 Variable 时间 Year 平均值 Mean 标准差 SD 最小值 Min. 最大值 Max 样木数 No. of tree ─ 47 13.13 31 82 林分平均胸径
Stand diameter /cm1997 11.79 3.12 7.68 20.02 2002 14.14 3.24 9.62 22.17 2007 15.56 3.27 10.68 23.13 2012 16.88 3.39 11.55 24.00 2017 18.38 3.62 12.19 25.77
下载: 导出CSV
表 3 3种不同形式生物量生长模型拟合效果的评价统计指标
Table 3. Statistics indices of goodness-of-fit for three different forms of biomass growth function
模型Model R2a SEE/kg TRE/% ASE/% MPSE/% MPE/% AIC BIC Eq. (3) 0.474 51 66.17 1.25 6.55 37.40 7.35 1 006.11 1 018.15 Eq. (4) 0.836 43 36.92 0.32 2.38 21.04 4.10 919.10 935.14 Eq. (5) 0.961 45 17.92 −0.03 0.17 11.31 1.99 812.60 844.68
下载: 导出CSV
表 4 最优生物量生长模型参数估计值的统计量
Table 4. Statistics indices of parameter estimate for the optimal biomass growth model
参数 Parameter 估计值 Estimate 渐进标准差 SD 变异系数 CV/ % a1 458.819 840 122.642 30 26.73 a2 590.385 661 152.798 11 25.88 a3 761.193 906 203.097 68 26.68 b −1.084 128 0.221 10 −20.39 c1 1.252 284 0.071 39 5.71 c2 1.287 937 0.077 12 5.99 c3 1.354 630 0.092 47 6.83 d 1.442 239 0.224 53 15.57
下载: 导出CSV
-
[1] Cao M, Woodward F I. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change[J]. Global Change Biology, 1998, 4(2): 185-198. doi: 10.1046/j.1365-2486.1998.00125.x [2] 胡海清, 罗斯生, 罗碧珍, 等. 林火干扰对广东省2种典型针叶林森林生物碳密度的影响[J]. 林业科学研究, 2020, 33(1):19-27. [3] 张少伟, 张弓乔, 惠刚盈. 内蒙古大兴安岭森林净初级生产力时空格局分析[J]. 林业科学研究, 2019, 32(5):74-82. [4] Jenkins, J C, Chojnacky, D C, Heath, L S, et al. National-scale biomass estimators for United States tree species[J]. Forest Science, 2004, 49(1): 12-35. [5] Ter-Mikaelian M T, Korzukhin M D. Biomass equations for sixty-five North American tree species[J]. Forest Ecology and Management, 1997, 97: 1-24. doi: 10.1016/S0378-1127(97)00019-4 [6] 曾伟生. 全国立木生物量方程建模方法研究[D]. 北京: 中国林业科学研究院, 2001. [7] Kittredge J. Estimation of the amount of foliage of trees and stands[J]. Journal of Forestry, 1944, 42(12): 905-912. [8] Wang C K. Biomass allometric equations for 10 co-occurring tree species in chinese temperate forests[J]. Forest Ecology and Management, 2006, 222(1): 9-16. [9] Zianis D, Xanthopoulos G, Kalabokidis K, et al. Allometric equations for aboveground biomass estimation by size class for Pinus brutia Ten.trees growing in north and south Aegean Islands, Greece[J]. European Journal of Forest Research, 2011, 130(2): 145-160. doi: 10.1007/s10342-010-0417-9 [10] Ozdemir E, Makineci E, Yilmaz E, et al. Biomass estimation of individual trees for coppice-originated oak forests[J]. European Journal of Forest Research, 2019, 138(4): 623-637. doi: 10.1007/s10342-019-01194-2 [11] 罗 红, 李百炼. 异速生长模型研究概述[J]. 生态学杂志, 2011, 30(9):2060-2065. [12] 薛春泉, 徐期瑚, 林丽平, 等. 广东主要乡土阔叶树种含年龄和胸径的单木生物量模型[J]. 林业科学, 2019, 55(2):97-108. doi: 10.11707/j.1001-7488.20190210 [13] 薛春泉, 徐期瑚, 林丽平, 等. 广东主要乡土阔叶树种单木生物量生长模型[J]. 华南农业大学学报, 2019, 40(2):65-75. doi: 10.7671/j.issn.1001-411X.201806031 [14] Rohner B, Waldner P, Lischke H, et al. Predicting individual-tree growth of central European tree species as a function of site, stand, management, nutrient, and climate effects[J]. European Journal of Forest Research, 2018, 137(1): 29-44. doi: 10.1007/s10342-017-1087-7 [15] Zeide B. Analysis of growth equations[J]. Forest Science, 1993, 39(3): 594-616. doi: 10.1093/forestscience/39.3.594 [16] 张建国, 段爱国. 理论生长方程与直径结构模型的研究[M]. 北京: 科学出版社, 2004. [17] Russell M B, Weiskittel A R, Kershaw J A. Comparing strategies for modeling individual-tree height and height-to-crown base increment in mixed-species Acadian forests of northeastern North America[J]. European Journal of Forest Research, 2014, 133(6): 1121-1135. doi: 10.1007/s10342-014-0827-1 [18] Wensel L, Meerschaert W, Biging G. Tree height and diameter growth models for northern California conifers[J]. Hilgardia, 1987, 55(8): 1-20. doi: 10.3733/hilg.v55n08p020 [19] Sharma M, Yin Zhang S. Height–diameter models using stand characteristics for Pinus banksiana and Picea mariana[J]. Scandinavian Journal of Forest Research, 2004, 19(5): 442-451. doi: 10.1080/02827580410030163 [20] Reineke, L. H. Perfecting a stand-density index for even-aged forests[J]. Journal of Agricultural Research, 1933, 46(7): 627-638. [21] 国家林业局. 立木生物量模型及碳计量参数-马尾松: LY/T 2263-2014[S]. 北京: 中国林业出版社, 2014. [22] 国家林业局. 立木生物量模型及碳计量参数-栎树: LT/T 2658-2016[S]. 北京: 中国林业出版社, 2016a. [23] 国家林业局. 立木生物量模型及碳计量参数-木荷: LY/T2660-2016[S]. 北京: 中国林业出版社, 2016b. [24] 曹 磊, 李海奎. 两种相容性生物量模型的比较—以广东省3个阔叶树种为例[J]. 生态学杂志, 2019, 38(6):1916-1925. [25] Richards F J. A flexible growth function for empirical use[J]. Journal of Experimental Botany, 1959, 10(2): 290-301. doi: 10.1093/jxb/10.2.290 [26] 唐守正, 李 勇, 符利勇. 生物数学模型的统计学基础[M]. 北京: 科学出版社, 2015. [27] 唐守正, 郎奎建, 李海奎. 统计和生物数学模型计算(ForStat教程)[M]. 北京: 科学出版社, 2009. [28] 曾伟生, 唐守正. 立木生物量方程的优度评价和精度分析[J]. 林业科学, 2011, 47(11):106-113. doi: 10.11707/j.1001-7488.20111117 [29] Berk K N. Validating regression procedures with new data[J]. Technimetrics, 1984, 26(4): 331-338. doi: 10.1080/00401706.1984.10487985 [30] Kozak A, Kozak R. Does cross validation provide additional information in the evaluation of regression models[J]. Canadian Journal of Forest Research, 2003, 33(6): 976-987. doi: 10.1139/x03-022 [31] Ketterings Q M, Coe R, van Noordwijk M, et al. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests[J]. Forest Ecology and Management, 2001, 146(1-3): 199-209. doi: 10.1016/S0378-1127(00)00460-6 [32] 曾伟生, 唐守正. 非线性模型对数回归的偏差校正及与加权回归的对比分析[J]. 林业科学研究, 2011, 24(2):137-143. [33] 曾伟生, 骆期邦, 贺东北. 论加权回归与建模[J]. 林业科学, 1999, 35(5):5-11. doi: 10.3321/j.issn:1001-7488.1999.05.002 [34] Zeng W S, Zhang H R, Tang S Z. Using the dummy variable model approach to construct compatible single-tree biomass equations at different scales—a case study for Masson pine (Pinus massoniana) in southern China[J]. Canadian Journal of Forest Research, 2011, 41(7): 1547-1554. doi: 10.1139/x11-068 [35] Fang J Y, Chen A P, Peng C, et al. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 2001, 262(5525): 2320-2322. [36] 龙时胜, 曾思齐, 甘世书, 等. 基于林木多期直径测定数据的异龄林年龄估计方法[J]. 中南林业科技大学学报: 自然科学版, 2018, 38(9):1-8. [37] 龙时胜, 曾思齐, 甘世书, 等. 基于林木多期直径测定数据的异龄林年龄估计方法Ⅱ[J]. 中南林业科技大学学报: 自然科学版, 2019, 39(6):23-29. [38] 惠淑荣, 于洪飞. 日本落叶松林分生长量Richards生长方程的建立与应用[J]. 生物数学学报, 2003, 18(2):204-206. doi: 10.3969/j.issn.1001-9626.2003.02.011 [39] Wang M, Borders B E, Zhao D. An empirical comparison of two subject-specific approaches to dominant heights modeling: The dummy variable method and the mixed model method[J]. Forest Ecology and Management, 2008, 255(7): 2659-2669. doi: 10.1016/j.foreco.2008.01.030 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 5992
- HTML全文浏览量: 5611
- PDF下载量: 42
- 被引次数: 0














