




-
微管是由α、β-微管蛋白以异二聚体的形式组装而成的中空管状结构[1]。植物细胞中,微管可以快速的聚合或者解聚、迅速转变微管列阵,对细胞的正常扩张和分化具有重要作用。微管结合蛋白(MAP)是一类对微管的组装和功能具有调控作用的蛋白[2]。Changjie等最早从烟草(Nicotiana tabacum L.) BY-2 (‘Bright Yellow-2’)悬浮培养细胞中分离得到一类与微管紧密结合且分子量约为65 kD的蛋白,即MAP65[3]。MAP65蛋白通常定位于一种或几种微管列阵上,是一类可与微管交联并促进微管成束的微管结合蛋白[4]。
从烟草BY-2悬浮培养细胞中分离出小原生质体,制备胞质提取物,之后加入紫杉醇协助组装微管,再将微管和MAP低温高速离心以去组装,经过数次组装-去组装循环,分离得到3个烟草MAP65蛋白[5]。后用同样的方法在胡萝卜(Daucus carota L.)悬浮细胞中分离得到3个胡萝卜MAP65蛋白[6]。随着拟南芥(Arabidopsis thaliana (L.) Heynh.)与水稻(Oryza sativa L.)基因组测序的完成,拟南芥和水稻中分别鉴定出了9和11个MAP65蛋白家族成员[7-8]。小立碗藓 (Physcomitrella patens (Hedw.) Bruch & Schimp.)中克隆得到了5个MAP65基因[9]。对拟南芥MAP65蛋白家族的研究表明,9个成员之间氨基酸相似性为28%~79%,C端序列同源性尤其低,C端包含与微管特定结合的结构域,决定了微管聚合的速率,不同基因的C端微管结合结构域不同,故以不同的效率促进微管聚合[10]。ATMAP65-1和ATMAP65-2促进扩增细胞的轴向生长,但不是维持细胞营养生长和生殖生长所必需[11]。ATMAP65-3的缺失导致成膜体异常,胞质分裂不完全及植株发育异常[12]。ATMAP65-4和ATMAP65-3具有高度同源性,功能相似,交联成膜体中的相邻反向平行的微管,这2个基因的同时缺失会导致植株生长受到严重影响[13]。ATMAP65-5在整个细胞周期中均有表达,且对微管解聚药具有更强的耐受力[14]。ATMAP65-6对微管聚合没有促进作用,但可诱导微管形成网状结构,该蛋白定位于线粒体,可能参与线粒体相关功能的实现[15]。植物中MAP65基因家族的成员数量较多,且在细胞周期的不同微管列阵中具有不完全相同的定位模式,在不同组织中的表达量亦存在差异,这预示该基因家族在植物中具有功能的分化[10,16-17]。
毛果杨(Populus trichocarpa Torr. & Gray)作为第一个完成基因组测序的木本植物,为鉴定木本植物MAP65基因家族成员和分析其进化及功能提供了可能[18]。本研究在毛果杨中鉴定出9个MAP65,通过其理化性质分析、亚细胞定位的预测、系统进化分析、基因结构和保守结构域分析、共线性分析及同义突变频率 (Ks)、非同义突变频率(Ka)分析、组织表达量以及启动子顺式作用元件分析,探究了PtMAP65基因家族的进化扩张模式,以期揭示PtMAP65基因家族的进化生物学意义,并为进一步研究PtMAP65基因家族成员的潜在功能提供新的线索。
-
在拟南芥数据库(
https://www.arabidopsis.org )下载ATMAP65基因家族9个成员的蛋白质序列,将其作为靶序列,使用BLASTP方法在Phytozome(v12.1,https://phytozome.jgi.doe.gov/ )中的毛果杨v3.1蛋白质组数据库中进行搜索,期望阈值设为−10,其他参数默认;候选基因再通过在线工具SMART(http://smart.embl-heidelberg.de/ )和NCBI-CDD (https://www.ncbi.nlm.nih.gov )进行保守结构域鉴定,最后得到9个PtMAP65蛋白。利用Prot-Param(https://web.expasy.org/ )计算PtMAP65蛋白家族的分子量和等电点;利用Plant-mPLoc(http://www.csbio.sjtu.edu.cn/ )、LocTree3 (https://rostlab.org/ )和 Wolfpsort (https://www.genscript.com/ )预测PtMAP65蛋白家族的亚细胞定位。 -
利用MAGA7.0中的ClustalW程序,将毛果杨、拟南芥、烟草、水稻、玉米(Zea mays L.)、巨桉(Eucalyptus grandis Hill)、挪威云杉(Picea abies (L.) Karst.)、北美云杉(Picea sitchensis (Bongard) Carrière)、莱茵衣藻(Chlamydomonas reinhardtii P.A.Dangeard)等物种的MAP65-Like蛋白序列进行多重序列比对;以莱茵衣藻作为外类群,采用邻接法(NJ)构建系统进化树,模型为p-distance,成对删除,校验参数(Bootstrap)为1 000次和5 000次重复。毛果杨、拟南芥、烟草和水稻MAP65蛋白家族系统进化树构建方法同上。
-
利用GSDS (2.0,
http://gsds.cbi.pku.edu.cn/ )分析毛果杨、拟南芥、烟草和水稻MAP65s的基因结构;利用MEME (v5.1.1,http://meme-suite.org/ )分析其蛋白序列的保守结构域,最大结构域数目设置为15。 -
从毛果杨基因组注释文件中获得PtMAP65基因家族染色体定位的位置信息;在linux系统下利用MCScanX 进行毛果杨基因组内共线性分析,并使用TBtools (v0.6696)作图。利用BioEdit Sequence Alignment Editor比对复制基因对的CDS序列,再用DnaSP5计算复制基因对的Ks和Ka,根据Ks值计算进化时间(T):T = Ks/2λ,λ = 9.1 × 10−9[19]。
-
使用TBtools (v0.6696)从毛果杨基因组序列文件中获取PtMAP65基因家族成员起始密码子上游2 000 bp的DNA序列作为启动子,使用plantCARE(
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )分析启动子中的顺式作用元件。 -
PtMAP65基因家族成员在不同组织中的表达数据来源于Poplar eFP Browser (
http://bar.utoronto.ca/ ),利用TBtools (v0.6696)分析制作基因表达热图,取log scal,极限值设置为2 000,其他参数默认。 -
通过在线BLASTP比对,从毛果杨中鉴定出14个候选的MAP65s。SMART和NCBI-CDD 分析结果显示:14条蛋白序列都具有完整或不完整的MAP65_ASE1 结构域 (PF03999);经过比对建树分析,删除影响树形的序列,最后得到9个MAP65s。根据其在染色体上的分布情况,将其命名为PtMAP65-1到PtMAP65-9(表1)。PtMAP65蛋白家族成员的氨基酸长度变化范围为570~730 aa,分子量变化范围为64.65~83.00 kDa,等电点变化范围为5.17~8.45。综合考虑PtMAP65蛋白家族成员用3种方法预测的亚细胞定位,最终预测:PtMAP65-1、PtMAP65-3和PtMAP65-9位于细胞核,PtMAP65-8位于叶绿体,其余成员位于细胞质。
表 1 PtMAP65基因家族及其理化性质
Table 1. The information of the PtMAP65 gene family
基因名称
Gene name基因登陆号
ID (Phytozome v12.1)DNA长度
DNA length/bpCDS长度
CDS length/bp氨基酸长度
amino acid length/aa分子量
Molecular weight/kDa等电点
pI预测的亚细胞定位
Predicted subcellular localizationPtMAP65-1 Potri.001G055100 4 089 1 809 602 68.73 5.70 细胞核 PtMAP65-2 Potri.001G356500 5 285 1 746 581 65.05 5.17 细胞质 PtMAP65-3 Potri.003G173300 4 551 1 779 592 67.62 5.87 细胞核 PtMAP65-4 Potri.003G192400 6 668 1 713 570 64.65 7.25 细胞质 PtMAP65-5 Potri.008G139700 7 530 1 803 600 68.12 7.14 细胞质 PtMAP65-6 Potri.011G092500 4 939 1 749 582 65.37 5.21 细胞质 PtMAP65-7 Potri.012G129600 4 717 1 755 584 67.08 5.41 细胞质 PtMAP65-8 Potri.014G070100 4 294 2 163 720 81.53 8.45 叶绿体 PtMAP65-9 Potri.015G131400 4 795 2 193 730 83.00 5.65 细胞核 -
为研究植物MAP65-Likes的系统进化关系,采用邻接法对毛果杨、拟南芥、烟草、水稻、巨桉、挪威云杉等物种的MAP65-Like蛋白构建系统进化树,校验参数(Bootstrap)分别为1 000次和5 000次重复,结果差异很小,图1中系统进化树的Bootstrap为1 000。从图1可以看出:该植物基因家族分为Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ5个亚组,裸子植物的MAP65-Likes仅分布于Ⅰ亚组和Ⅴ亚组中,且独立地聚为一簇;被子植物的MAP65-Likes在5个亚组中均有分布,且单子叶和双子叶植物MAP65s均匀地分布于各个亚组,这说明5个亚组的分化发生于裸子植物和被子植物分化之后、单子叶植物和双子叶植物分化之前。在Ⅰ、Ⅱ、Ⅲ和Ⅳ4个亚组中,单子叶植物和双子叶植物的MAP65-Likes各自聚为一簇,在Ⅴ亚组中,双子叶植物毛果杨、拟南芥和烟草的4个MAP65-Likes聚为一簇,其他8个MAP65-Likes与单子叶植物水稻的3个MAP65s聚为一簇。在Ⅰ亚组中,PtMAP65-2/PtMAP65-6组成旁系同源基因对,并与ATMAP65-1/ATMAP65-2这一对旁系同源基因聚为一支;在Ⅱ亚组中,PtMAP65-4/Eucgr.J00083.1组成直系同源基因对,并与ATMAP65-5聚为一支;在Ⅲ亚组中,PtMAP65-1/PtMAP65-3组成旁系同源基因对,并与Eucgr.B03489.1聚为一支;在Ⅴ亚组中,PtMAP65-7/PtMAP65-9组成旁系同源基因对,并与旁系同源基因对Eucgr.K00145.1/Eucgr.A02875.1聚为一支,另外,PtMAP65-8/ATMAP65-4组成直系同源基因对;在Ⅳ亚组中,PtMAP65-5/Eucgr.G02071.1组成直系同源基因对,这表明MAP65家族中毛果杨与巨桉的亲缘关系最近,这也与二者是多年生木本植物相一致。
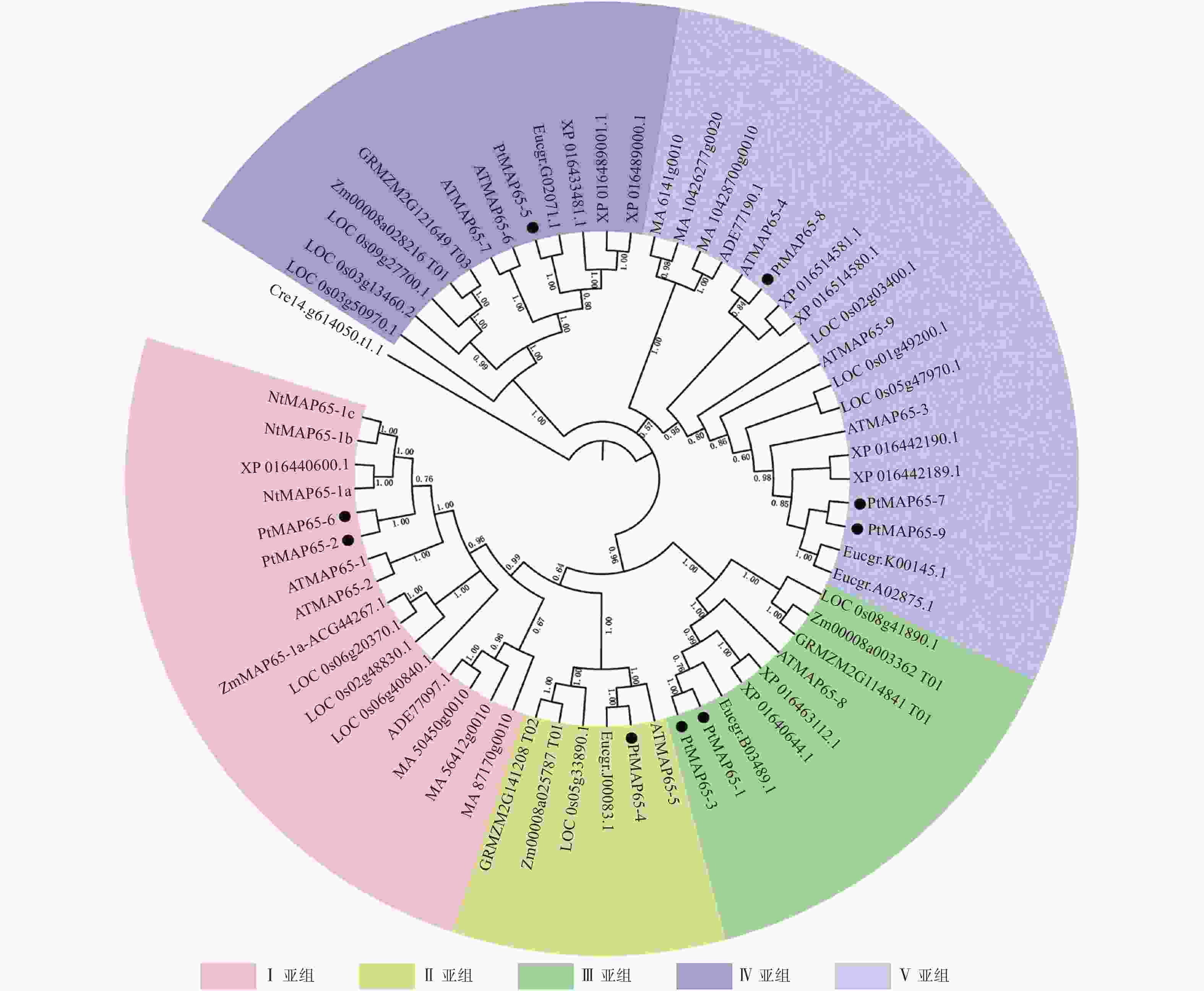
图 1 植物MAP65-Likes系统进化分析
Figure 1. Phylogenetic analysis of plant MAP65-Likes protein sequences
-
为分析MAP65家族成员的基因结构特征,利用GSDS绘制了毛果杨、拟南芥、烟草和水稻MAP65家族成员的基因结构图。图2A表明:在Ⅰ亚组中,9个MAP65s有9个外显子、8个内含子,仅ATMAP65-1有8个外显子、7个内含子;在Ⅱ亚组中,MAP65有11个外显子、10个内含子;在Ⅲ亚组中,3个MAP65有11个外显子、10个内含子,LOC_Os08g41890.1有10个外显子、9个内含子;在Ⅳ亚组中,5个MAP65有11个外显子、10个内含子,仅LOC_Os03g50970.1有13个外显子、12个内含子;在Ⅴ亚组中,MAP65的外显子个数为9~12、内含子个数为8~11。另外,在各个亚组中,外显子的长度相似,内含子的长度差异较大,而同一物种的MAP65之间的内含子长度更相似。
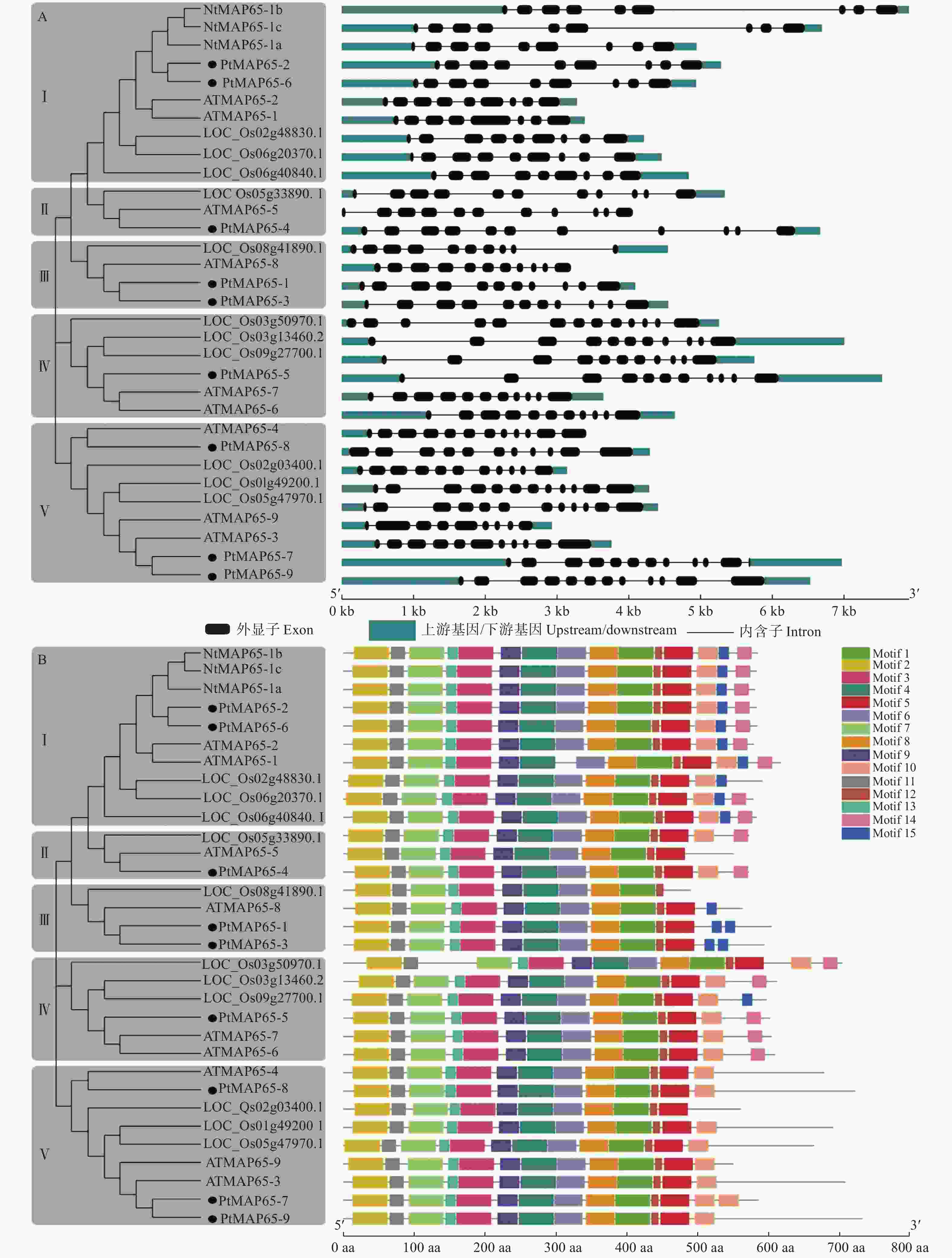
图 2 MAP65家族的基因结构和保守结构域分析
Figure 2. Gene structures and protein domains analysis of MAP65 gene family
利用MEME分析了MAP65蛋白家族的保守结构域。图2B表明:各个MAP65蛋白家族成员具有11~15个不等的结构域,第1、2、3、4、6、7、8、9、11、12和15个结构域较保守,在32条MAP65蛋白序列中均有分布。另外,植物MAP65蛋白序列的C端结构域较多变,各个亚组的MAP65蛋白具有相似的结构域分布。
-
根据毛果杨基因组序列文件和注释文件,分析了9个PtMAP65基因在染色体的定位情况。PtMAP65-1和PtMAP65-2位于Chr01,PtMAP65-3和PtMAP65-4位于Chr03,PtMAP65-5到PtMAP65-9分别位于Chr08、Chr11、Chr12、Chr14和Chr15。毛果杨基因组内共线性分析结果显示:MAP65基因家族中共有6对复制基因,且均为片段复制,其中PtMAP65-7、PtMAP65-8和PtMAP65-9互为复制基因(图3)。利用DnaSP5计算复制基因对的Ka和Ks,算得的Ks值为0.2341~1.9338,表明复制时间最早发生于106.25百万年前,最晚发生于12.86百万年前;除复制基因对PtMAP65-4/Potri.001G032566的Ka/Ks为0.617 3外,其余复制基因对的Ka/Ks < 0.25,表明PtMAP65基因家族在进化过程中受到强纯化选择的作用(表2)。
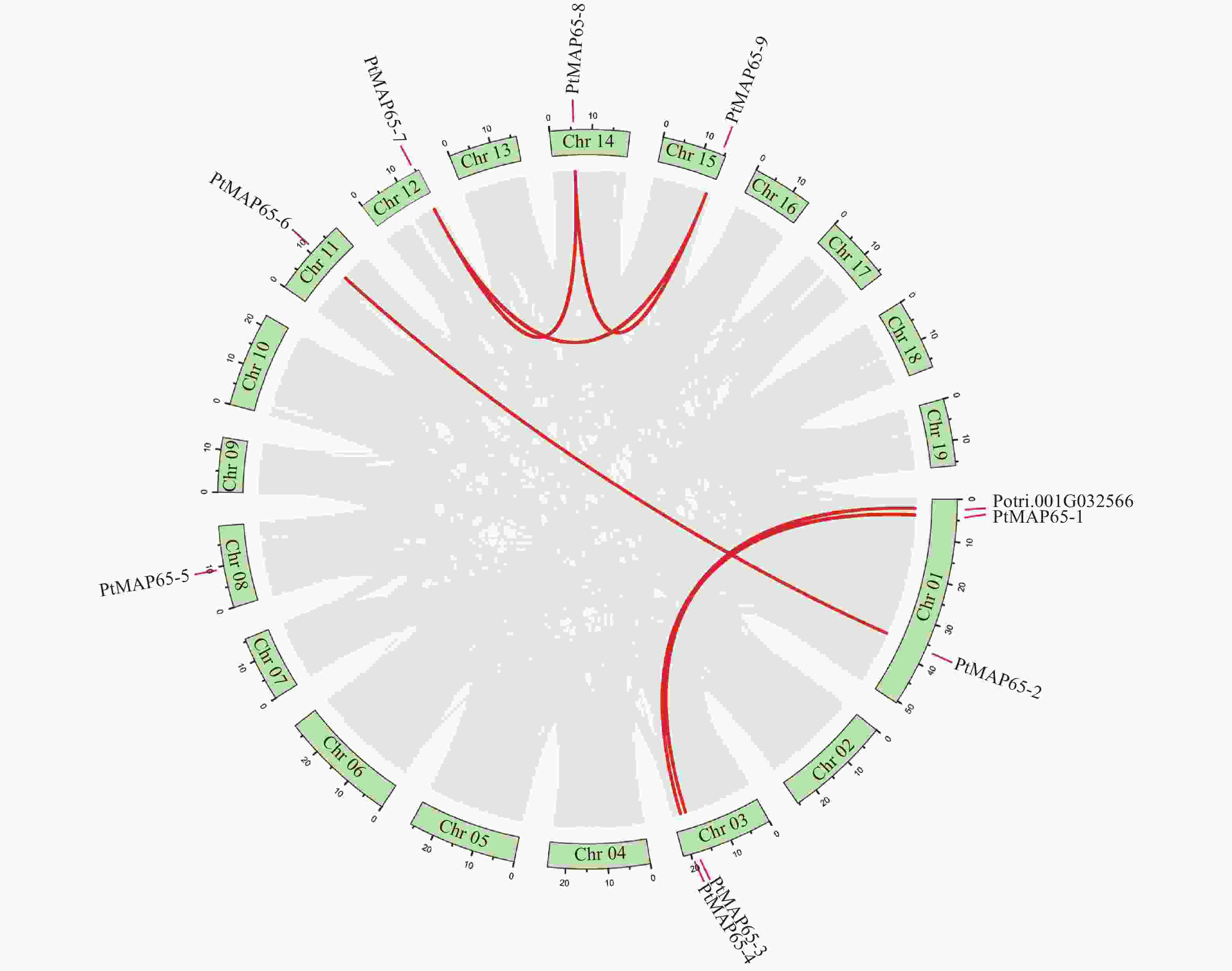
图 3 PtMAP65基因家族染色体分布与复制基因对
Figure 3. Chromosomal locations and duplicated gene pairs of PtMAP65 gene family
表 2 复制基因对的Ks、Ka分析
Table 2. Ks and Ka analysis of duplicated gene pairs
复制基因对
Pair of duplication gene同义突变频率
Ks非同义突变频率
Ka非同义突变率与同义突变率的比值
Ka/Ks复制时间/(百万年前)
Duplication time/(Million years ago)PtMAP65-1 PtMAP65-3 0.242 3 0.047 6 0.196 5 13.31 PtMAP65-2 PtMAP65-6 0.276 7 0.044 3 0.160 1 15.20 PtMAP65-4 Potri.001G032566 0.234 1 0.144 5 0.617 3 12.86 PtMAP65-7 PtMAP65-8 1.933 8 0.307 3 0.158 9 106.25 PtMAP65-7 PtMAP65-9 0.242 2 0.056 7 0.234 1 13.31 PtMAP65-8 PtMAP65-9 1.761 2 0.300 0 0.170 3 96.77 -
为了解转录调控机制,利用PlantCARE数据库分析PtMAP65基因家族启动子的顺式作用元件。根据顺式作用元件的功能,将其分为光响应元件、激素响应元件、逆境胁迫响应元件和植物生长发育元件。图4表明:在PtMAP65基因家族中,光响应元件和激素响应元件的个数较多,逆境胁迫响应元件和植物生长发育元件的个数较少;PtMAP65-3中的激素响应元件远多于其他基因,PtMAP65-4、PtMAP65-7和PtMAP65-8中未预测到植物生长发育元件。
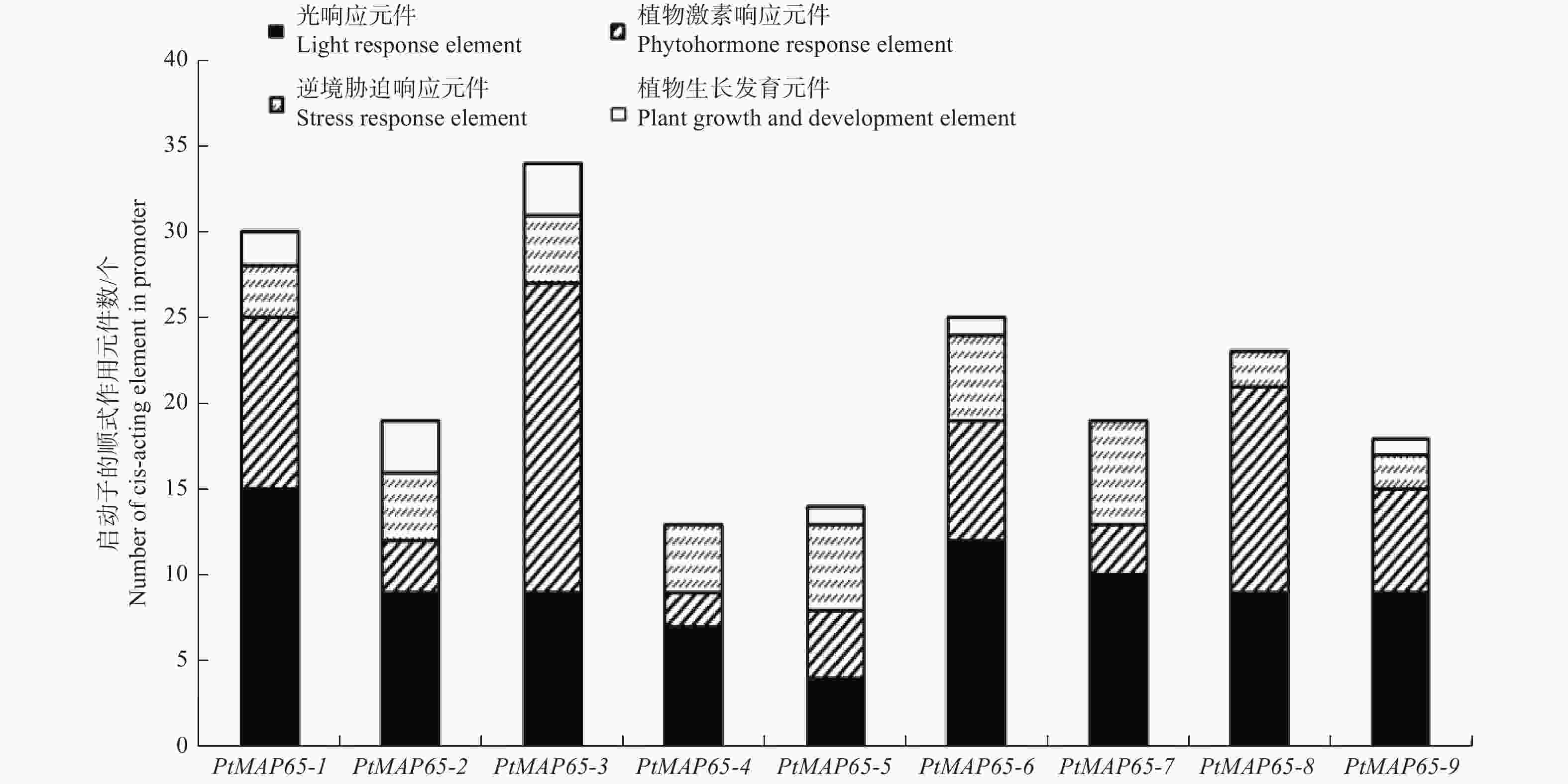
图 4 PtMAP65基因家族启动子的顺式作用元件分析
Figure 4. Cis-elements analysis of PtMAP65 gene family promoter
-
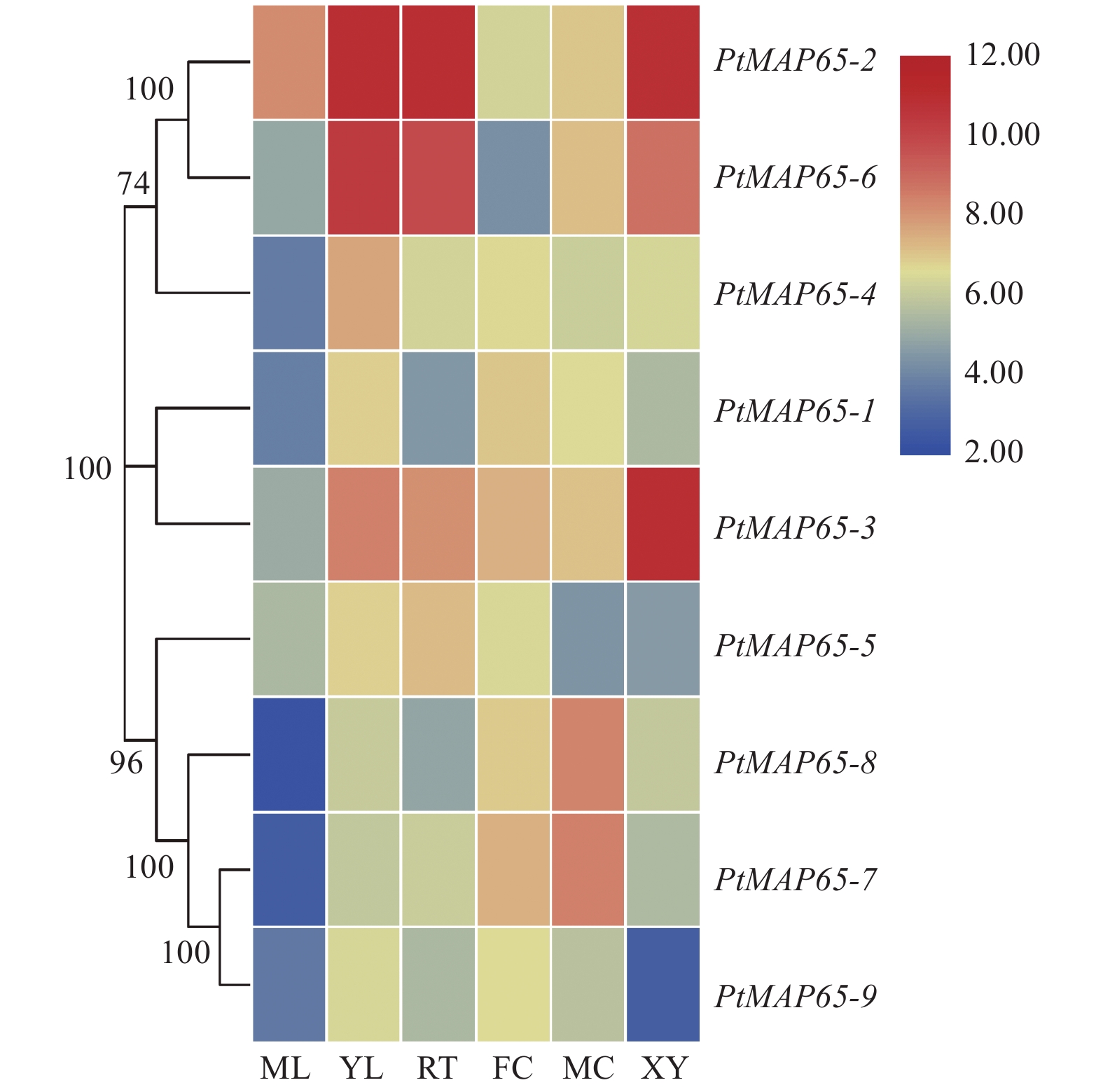
从Populus eFP browser数据库下载得到PtMAP65基因家族成员在成熟叶、幼叶、根、雌花、雄花和木质部中的表达数据,绘制热图。图5表明:复制基因对PtMAP65-2/PtMAP65-6在幼叶、根和木质部均有高度表达;复制基因对PtMAP65-7/PtMAP65-8在雌花和雄花中高度表达;PtMAP65-3在木质部中表达量最高;PtMAP65-1和PtMAP65-4主要在幼叶和雌花中表达;PtMAP65-5在幼叶和根中表达水平较高;PtMAP65-9在各个组织表达水平较低。

图 5 PtMAP65基因家族在不同组织中的表达模式
Figure 5. Expression profiles of PtMAP65 gene family in different tissues
-
MAP65蛋白最早在烟草中发现,是一类具有稳定微管功能的蛋白[3]。本研究在毛果杨中鉴定出9个PtMAP65基因,保守结构域分析发现,MAP65_ASE(pfam03999)结构域普遍存在于拟南芥、毛果杨、水稻和烟草MAP65蛋白家族中[16],暗示MAP65_ASE为植物MAP65家族行使共同功能的主要结构域。研究发现,植物MAP65的C端包含一个高度分化的微管结合结构域[20],这个结构域参与微管聚合、成束,且对其亚细胞定位有一定影响[21],在有丝分裂中期,C端蛋白的磷酸化可及时地下调MAP65与微管的结合[22-23]。进一步分析发现,毛果杨PtMAP65蛋白的C端同样包含一个高度分化的微管结合结构域,这可能与毛果杨MAP65家族的功能分化相关。
片段复制和串联复制是基因家族扩张的主要动力[24]。毛果杨基因组内共线性分析发现,PtMAP65基因家族中共有6对复制基因,且均为片段复制,这说明片段复制是PtMAP65基因家族进化扩张的主要动力。毛果杨经历了3次全基因组复制事件,分别是古老的复制事件、100~120百万年前的真蔷薇分支复制事件和60~65百万年前的杨柳科复制事件[18]。分析PtMAP65复制基因发生时间发现,PtMAP65-7/PtMAP65-8、PtMAP65-8/PtMAP65-9的复制时间与真蔷薇分支复制事件接近,PtMAP65-7/PtMAP65-9的复制大约发生于为13百万年前,由此推测,位于Chr14的PtMAP65-8经过2次片段复制或者三倍化复制得到位于Chr12的PtMAP65-7和位于Chr15的PtMAP65-9。最新研究表明,被子植物在20百万年前亦发生了1次全基因组复制事件,当时气温和CO2浓度均较低,保留下来的基因多与盐胁迫、低温胁迫、干旱胁迫等非生物胁迫有关[25]。分析发现,PtMAP65-1/PtMAP65-3、PtMAP65-2/PtMAP65-6、PtMAP65-4/ Potri.001G032566.2,PtMAP65-7/PtMAP65-9这4对基因的复制时间均与此次全基因组复制事件的时间相吻合。
通过对植物MAP65系统发育关系的研究发现,PtMAP65多与ATMAP65或巨桉MAP65聚为一支,这说明毛果杨和拟南芥、巨桉的亲缘关系最相近。前人研究表明:ATMAP65-1定位于除萼片和花药以外的所有植物器官和组织,且在根表皮细胞、子叶和下胚轴细胞中发现ATMAP65-1蛋白与微管相结合,在体外ATMAP65-1在微管之间形成25 nm的交联桥,促进微管成束[26];ATMAP65-2在调节细胞的轴向生长方面具有重要作用;在ATMAP65-1和ATMAP65-2双突变体中,没有观察到明显的膨胀、变形和其他形态学缺陷的细胞,但植株生长显著迟缓[11]。分析发现,PtMAP65-2、PtMAP65-6与ATMAP65-1、ATMAP65-2聚为一支,位于Ⅰ亚组,且其在雄花和雌花中表达量低,在其他组织中表达量均较高,这暗示PtMAP65-2和PtMAP65-6可能通过参与细胞骨架的构建进而影响植株形态发生。ATMAP65-3与有丝分裂微管列阵紧密相关,在有丝分裂过程中,该蛋白组织纺锤体形态发生和成膜体形成,ATMAP65-3基因突变体根尖、茎尖分裂能力弱,植株矮小[27];引入ATMAP65-4拷贝基因的植株显著抑制了由ATMAP65-3突变引起的细胞质分裂和幼苗生长缺陷,而ATMAP65-3和ATMAP65-4的同时突变将导致植物死亡[13]。分析发现,PtMAP65-7/PtMAP65-9与ATMAP65-3聚为一簇,PtMAP65-8与ATMAP65-4聚为一簇,位于Ⅴ亚组,暗示PtMAP65-7、PtMAP65-8和PtMAP65-9这3个基因通过参与细胞分裂过程决定植物的生长发育情况,但这3个基因在毛果杨的各个组织中表达量较低,其中的调控方式尚需进一步实验来明确。ATMAP65-8在与周质微管的结合较弱,可能仅结合周质微管中的某一部分[28]。分析发现,PtMAP65-1/PtMAP65-3与ATMAP65-8聚为一支,位于Ⅲ亚组,这对基因的复制发生于20百万年前的全基因组复制事件,预测的亚细胞定位在细胞核,特别的是:PtMAP65-3启动子上的激素响应元件远远高于其他基因,且该基因在木质部高度表达,推测PtMAP65-3可能通过参与各种非生物胁迫响应而影响植物次生生长。综上,PtMAP65-1、PtMAP65-2、PtMAP65-3、PtMAP65-6、PtMAP65-7、PtMAP65-8和PtMAP65-9可能紧密结合微管,通过影响细胞分裂进而影响植物生长,具体的作用机制及生物学功能仍需进一步研究。
-
本研究利用BLASTP比对,在毛果杨中鉴定了9个PtMAP65s;构建多个物种的MAP65s系统进化树,将其分为5个亚组;按不同亚组将基因结构图和保守结构域图归类比较,同一亚组的MAP65s具有相似的基因结构和保守结构域;利用毛果杨基因组内共线性及Ka、Ks分析PtMAP65基因家族的进化,片段复制是其扩张的主要动力,且复制基因在进化过程中受到强纯化选择的作用;亚细胞定位预测、基于芯片数据的表达量和启动子的顺式作用元件综合分析表明:在PtMAP65基因家族中,不同亚组间的PtMAP65s已经存在功能分化,同一亚组内的PtMAP65s即复制基因对之间仍存在功能冗余。
毛果杨MAP65基因家族的扩张与表达分析
Expansion and Expression Analysis of PtMAP65 Gene Family in Populus trichocarpa
-
摘要:
目的 以林木模式植物毛果杨为研究材料,旨在研究毛果杨MAP65基因家族成员的扩张与表达情况,为MAP65的功能研究提供参考。 方法 利用BLASTP基于Phytozome数据库鉴定毛果杨的MAP65基因家族成员,采用Prot-Param、Plant-mPLoc、LocTree3、Wolfpsort、MAGA7.0、GSDS、MEME、MCScanX、TBtools、BioEdit Sequence Alignment Editor、DnaSP5、plantCARE等工具分析其理化性质,预测其亚细胞定位,构建系统进化树,绘制基因结构图,分析保守结构域、基因复制事件、启动子元件,并基于基因芯片数据分析PtMAP65基因家族成员在不同组织中的表达量。 结果 利用BLASTP比对方法鉴定得到了9个MAP65基因。系统进化分析结果显示:植物MAP65基因家族分为5个亚组,且每个亚组都具有相似的基因结构和保守结构域。毛果杨基因组内共线性分析及同义突变频率 (Ks)、非同义突变频率(Ka)分析结果表明:PtMAP65基因家族中有6对片段复制基因,且复制基因在进化过程中受到强纯化选择的作用。PtMAP65基因家族成员的表达数据分析显示:该基因家族成员在不同组织中有不同的表达模式,暗示其功能的分化。启动子的顺式作用元件分析表明:PtMAP65s的启动子中有较多的光响应元件和激素响应元件。 结论 片段复制是PtMAP65基因家族进化扩张的主要动力,推测基因家族成员之间存在功能分化。 Abstract:Objective To study the expansion and expression of the MAP65 gene family members by using Populus trichocarpa, a model forest plant, as research material. Method Using BLASTP to identify the members of PtMAP65 gene family based on Phytozome database, and using tools such as Prot-Param, Plant-mPLoc, LocTree3, Wolfsort, MAGA7.0, GSDS, MEME, MCScanX, TBtools, BioEdit Sequence Alignment Editor, DnaSP5 and plantCARE to analyze the physical and chemical properties, predict subcellular locations, construct a phylogenetic tree, draw a gene structure diagram, analyze conserved domains, gene duplication events and promoter elements; and analyze the expression of PtMAP65 gene family members in different tissues based on gene chip data. Result Nine MAP65 genes were identified by BLASTP comparison. Phylogenetic analysis showed that the plant MAP65 gene family can be divided into 5 groups, and each group had a similar gene structure and conserved domain. The results of collinearity analysis and Ka and Ks analysis of the P. trichocarpa genome indicated that there were six pairs of segmental duplication genes in the PtMAP65 gene family, and the duplication genes were subject to strong purification selection in the evolutionary process. Analysis of the expression data showed that PtMAP65 gene family members had different expression patterns in different tissues, suggesting their functional divergence. Cis-acting element analysis of the promoter showed that PtMAP65s promoter had more light response and hormone response elements than stress response, growth and development elements. Conclusion Segmental duplication is a main force driving the evolution and expansion of PtMAP65 gene family, and it is speculated that there has been functional divergence among the PtMAP65 gene family members. -
Key words:
- Populus trichocarpa
- / PtMAP65
- / phylogenetic analysis
- / gene structure
-
图 1 植物MAP65-Likes系统进化分析
Figure 1. Phylogenetic analysis of plant MAP65-Likes protein sequences
图 2 MAP65家族的基因结构和保守结构域分析
Figure 2. Gene structures and protein domains analysis of MAP65 gene family
图 3 PtMAP65基因家族染色体分布与复制基因对
Figure 3. Chromosomal locations and duplicated gene pairs of PtMAP65 gene family
图 4 PtMAP65基因家族启动子的顺式作用元件分析
Figure 4. Cis-elements analysis of PtMAP65 gene family promoter
图 5 PtMAP65基因家族在不同组织中的表达模式
Figure 5. Expression profiles of PtMAP65 gene family in different tissues
表 1 PtMAP65基因家族及其理化性质
Table 1. The information of the PtMAP65 gene family
基因名称
Gene name基因登陆号
ID (Phytozome v12.1)DNA长度
DNA length/bpCDS长度
CDS length/bp氨基酸长度
amino acid length/aa分子量
Molecular weight/kDa等电点
pI预测的亚细胞定位
Predicted subcellular localizationPtMAP65-1 Potri.001G055100 4 089 1 809 602 68.73 5.70 细胞核 PtMAP65-2 Potri.001G356500 5 285 1 746 581 65.05 5.17 细胞质 PtMAP65-3 Potri.003G173300 4 551 1 779 592 67.62 5.87 细胞核 PtMAP65-4 Potri.003G192400 6 668 1 713 570 64.65 7.25 细胞质 PtMAP65-5 Potri.008G139700 7 530 1 803 600 68.12 7.14 细胞质 PtMAP65-6 Potri.011G092500 4 939 1 749 582 65.37 5.21 细胞质 PtMAP65-7 Potri.012G129600 4 717 1 755 584 67.08 5.41 细胞质 PtMAP65-8 Potri.014G070100 4 294 2 163 720 81.53 8.45 叶绿体 PtMAP65-9 Potri.015G131400 4 795 2 193 730 83.00 5.65 细胞核  下载: 导出CSV
下载: 导出CSV
表 2 复制基因对的Ks、Ka分析
Table 2. Ks and Ka analysis of duplicated gene pairs
复制基因对
Pair of duplication gene同义突变频率
Ks非同义突变频率
Ka非同义突变率与同义突变率的比值
Ka/Ks复制时间/(百万年前)
Duplication time/(Million years ago)PtMAP65-1 PtMAP65-3 0.242 3 0.047 6 0.196 5 13.31 PtMAP65-2 PtMAP65-6 0.276 7 0.044 3 0.160 1 15.20 PtMAP65-4 Potri.001G032566 0.234 1 0.144 5 0.617 3 12.86 PtMAP65-7 PtMAP65-8 1.933 8 0.307 3 0.158 9 106.25 PtMAP65-7 PtMAP65-9 0.242 2 0.056 7 0.234 1 13.31 PtMAP65-8 PtMAP65-9 1.761 2 0.300 0 0.170 3 96.77
下载: 导出CSV
-
[1] 张少斌, 刘 曦, 张立军, 等. 植物微管结合蛋白[J]. 植物生理学通讯, 2009, 45(3):291-298. [2] Hamada T. Microtubule-associated proteins in higher plants[J]. Journal of Plant Research, 2007, 120(1): 79-98. doi: 10.1007/s10265-006-0057-9 [3] Changjie J, Sonobe S. Identification and preliminary characterization of a 65 kDa higher-plant microtubule-associated protein[J]. Journal of Cell Science, 1993, 105(1): 891-901. [4] Hamada T. Microtubule organization and microtubule-associated proteins in plant cells[J]. International Review of Cell and Molecular Biology, 2014, 312C: 1-52. [5] Smertenko A, Saleh N, Igarashi H, et al. A new class of microtubule-associated proteins in plants[J]. Nature Cell Biology, 2000, 2(10): 750-753. doi: 10.1038/35036390 [6] Chan J, Rutten T, Lloyd C. Isolation of microtubule-associated proteins from carrot cytoskeletons: a 120 kDa MAP decorates all four microtubule arrays and the nucleus[J]. The Plant Journal, 1996, 10(2): 251-259. doi: 10.1046/j.1365-313X.1996.10020251.x [7] Hussey P J, Hawkins T J, Igarashi H, et al. The Plant cytoskeleton: recent advances in the study of the plant microtubule-associated proteins MAP-65, MAP-190 and the Xenopus MAP215-like protein, MOR1[J]. Plant Molecular Biology, 2002, 50(6): 915-924. doi: 10.1023/A:1021236307508 [8] Guo L, Ho C K, Kong Z, et al. Evaluating the microtubule cytoskeleton and its interacting proteins in monocots by mining the rice genome[J]. Annals of Botany, 2009, 103(3): 387-402. doi: 10.1093/aob/mcn248 [9] Kosetsu K, De Keijzer J, Janson M E, et al. MICROTUBULE-ASSOCIATED PROTEIN65 is essential for maintenance of phragmoplast bipolarity and formation of the cell plate in Physcomitrella patens[J]. Plant Cell, 2013, 25(11): 4479-4492. doi: 10.1105/tpc.113.117432 [10] Smertenko A, Kaloriti D, Chang H, et al. The C-terminal variable region specifies the dynamic properties of Arabidopsis microtubule-associated protein MAP65 isotypes[J]. The Plant Cell, 2008, 20(12): 3346-3358. doi: 10.1105/tpc.108.063362 [11] Lucas J R, Courtney S, Hassfurder M, et al. Microtubule-associated proteins MAP65-1 and MAP65-2 positively regulate axial cell growth in etiolated Arabidopsis hypocotyls[J]. The Plant Cell, 2011, 23(5): 1889-1903. [12] Sabine Müller, Smertenko A, Wagner V, et al. The plant microtubule-associated protein ATMAP65-3/PLE is essential for cytokinetic phragmoplast function[J]. Current Biology, 2004, 14(5): 412-417. doi: 10.1016/j.cub.2004.02.032 [13] Li H, Sun B, Sasabe M, et al. Arabidopsis MAP65‐4 plays a role in phragmoplast microtubule organization and marks the cortical cell division site[J]. New Phytologist, 2017, 215(1): 187-201. doi: 10.1111/nph.14532 [14] Van Damme D, Van Poucke K, Boutant E, et al. In vivo dynamics and differential microtubule-binding activities of MAP65 proteins[J]. Plant Physiology, 2004, 136(4): 3956-3967. doi: 10.1104/pp.104.051623 [15] Mao T, Jin L, Li H, et al. Two microtubule-associated proteins of the Arabidopsis MAP65 family function differently on microtubules[J]. Plant Physiology, 2005, 138(2): 654-662. doi: 10.1104/pp.104.052456 [16] Pellman D, Bagget M, Tu Y H, et al. Two microtubule-associated proteins required for anaphase spindle movement in Saccharomyces cerevisiae[J]. Journal of Cell Biology, 1995, 130(6): 1373-1385. doi: 10.1083/jcb.130.6.1373 [17] Jiang W, Jimenez G, Wells N J, et al. PRC1: A human mitotic spindle-associated CDK substrate protein required for cytokinesis[J]. Molecular Cell, 1999, 2(6): 877-885. [18] Tuskan G A, Difazio S P, Jansson S, et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray)[J]. Science, 2006, 313(5793): 1596-1604. doi: 10.1126/science.1128691 [19] Lynch M, Conery J S. The evolutionary fate and consequences of duplicate genes[J]. Science, 2000, 290(5494): 1151-1155. doi: 10.1126/science.290.5494.1151 [20] Gaillard J, Neumann E, Van Damme D, et al. Two microtubule-associated proteins of Arabidopsis MAP65s promote antiparallel microtubule bundling[J]. Molecular Biology of the Cell, 2008, 19(10): 4534-4544. [21] Smertenko A, Chang H, Sonobe S, et al. Control of the AtMAP65-1 interaction with microtubules through the cell cycle[J]. Journal of Cell Science, 2006, 119(15): 3227-3237. doi: 10.1242/jcs.03051 [22] Mao G, Chan J, Calder G, et al. Modulated targeting of GFP-AtMAP65-1 to central spindle microtubules during division[J]. Plant Journal, 2010, 43(4): 469-478. [23] Sasabe, M. Phosphorylation of NtMAP65-1 by a MAP kinase down-regulates its activity of microtubule bundling and stimulates progression of cytokinesis of tobacco cells[J]. Genes & Development, 2006, 20(8): 1004. [24] Cannon S B, Mitra A, Baumgarten A, et al. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana[J]. BMC Plant Biology, 2004, 4(1): 10-10. doi: 10.1186/1471-2229-4-10 [25] Wu S, Han B, Jiao Y, et al. Genetic contribution of paleopolyploidy to adaptive evolution in angiosperms[J]. Molecular Plant, 2020, 13(1): 59-71. doi: 10.1016/j.molp.2019.10.012 [26] Smertenko A, Chang H, Wagner V, et al. The Arabidopsis microtubule-associated protein AtMAP65-1: molecular analysis of its microtubule bundling activity[J]. The Plant Cell, 2004, 16(8): 2035-2047. [27] Michal Q, Baurès Isabelle, Caroline H, et al. The Arabidopsis microtubule-associated protein MAP65-3 supports infection by filamentous biotrophic pathogens by down-regulating salicylic acid-dependent defenses[J]. Journal of Experimental Botany, 2016(6): 1731-1743. [28] Damme D V, Bouget F Y, Poucke K V, et al. Molecular dissection of plant cytokinesis and phragmoplast structure: a survey of GFP-tagged proteins[J]. Plant Journal, 2004, 40(3): 386-398. doi: 10.1111/j.1365-313X.2004.02222.x -

 点击查看大图
点击查看大图

计量
- 文章访问数: 6442
- HTML全文浏览量: 3621
- PDF下载量: 72
- 被引次数: 0
