



-
细胞分裂素是植物生长过程中的一类重要调控激素,其主要通过双组分系统进行信号感知与传递,参与芽的分化、根的生长、种子发育等过程,且在低温、干旱及高盐等非生物胁迫的逆境响应中也发挥重要作用[1-4]。
植物双组分系统由组氨酸激酶(Histidine Kinase,HK)蛋白、组氨酸磷酸转移(Histidine Phosphotransfer,HPt)蛋白和反应调节(Response Regulator,RR)蛋白组成[5-6]。拟南芥(Arabidopsis thaliana)组氨酸激酶由AHK2、AHK3和AHK4(又名CRE1)编码,与细胞分裂素结合对下游的AHP(Arabidopsis HPts)蛋白和ARR(Arabidopsis Response Regulators)调节蛋白进行调节,进而传递植物激素(细胞分裂素、乙烯)和非生物胁迫信号[2, 7-11]。研究表明,组氨酸激酶基因在植物的生长发育中发挥重要的调控作用,如AHK4对植物根的发育具有明显的调控作用[12],AHK3则对植物叶片的衰老和细胞分化起主导作用[13-15],AHK2和AHK3共同调节拟南芥的种子萌发和芽的生长[16]。另外,组氨酸激酶基因在植物应对低温、干旱及高盐等非生物胁迫过程中也发挥重要作用[3-4]。Kang等[17]发现,拟南芥ahk2和ahk3单突变体比野生型个体抗旱性明显增强,并且ahk2/ahk3和ahk3/ahk4双突变体比相应的单突变体抗旱。低温胁迫时,ahk2/ahk3和ahk3/ahk4双突变体比野生型个体具有较高的抗冻性[18],ahk2和ahk3单突变体对干旱、盐胁迫、低温和强光等非生物胁迫的抗性比野生型植株显著增强[3, 18]。另外,通过萌发试验发现,ahk2、ahk3及ahk4单突变体对外源ABA高度敏感,外源ABA对拟南芥组氨酸激酶基因具有负调控作用[3]。通过ahk2/ahk3双突变体和野生型植株全基因组表达分析比较发现,组氨酸激酶基因在植物ABA调控的抗逆反应及非ABA调控的抗逆反应中均起重要作用[19]。在毛果杨(P. trichocarpa Torr. & Gray)基因组中鉴定出1个AHK2同源基因PtHK2、2个AHK3同源基因PtHK3a和PtHK3b,研究表明,毛果杨PtHK2、PtHK3a和PtHK3b在杨树形成层发育过程中具有重要调控作用[20]。然而,组氨酸激酶基因是否参与林木抗逆反应尚未见报道。
银腺杨‘84K’(P. alba × P. glandulosa ‘84K’)是我国从韩国引进的白杨派优良品种,生长快、材质好、抗性强、适应性广,是优良的绿化树种、生态树种和用材树种,且因其易于组培及遗传转化,成为林木基因工程的理想材料。本研究以银腺杨‘84K’为材料,克隆了拟南芥AHK3同源基因(PaHK3b)的启动子及全长CDS(coding sequence),对其启动子元件及蛋白结构域进行了分析,并对PaHK3b基因在高温、干旱、盐胁迫等非生物胁迫及不同植物激素处理下的表达进行了qPCR检测;同时,通过原核表达初步确定其生物学功能。该研究为利用组氨酸激酶基因进行杨树抗逆分子改良奠定了基础。
-
银腺杨‘84K’。
-
根据Phytozome网站上公布的毛果杨基因组信息,查找基因上游启动子序列,设计PaHK3b基因启动子区域特异性引物PaHK3b-pro-F和PaHK3b-pro-R(表1)。采用植物基因组DNA提取试剂盒(TIANGEN,北京)提取‘84K’杨叶片总DNA,用常规PCR方法进行目的片段扩增,用1.5%琼脂糖凝胶进行电泳检测,用胶回收试剂盒(Axygen,美国)回收并纯化目的片段,并连接到克隆载体pMDTM19-T Vector(Takara,日本)转化至大肠杆菌DH5α(TIANGEN,北京),经过蓝白斑培养基(Amp+)筛选,挑取阳性单克隆送至生物公司(中美泰和,北京)进行序列测定。利用在线软件plantCARE(
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )分析该基因启动子中含有的顺式作用元件。表 1 相关引物序列及产物大小
Table 1. The sequences and PCR product size of primers used in this study
基因 Gene 引物序列 Primer sequence (5'→3') 产物长度 Product length/bp PaHK3b-F ATGAGTTTGCTTCATGTATTTGGG 3 060 PaHK3b-R TAAATTCGCACTGGAAGTTGGC PaHK3b-pro-F GTGGCTTTGGGAGGAAGGAAT 1 600 PaHK3b-pro-R CCACATCTTGCCACCATCACC PaHK3b-q-F ATGAGTTTGCTTCATGTATTTGG 243 PaHK3b-q-R CTATAAATTCGCACTGGAAGTTG Actin-F AAACTGTAATGGTCCTCCCTCCG 193 Actin-R GCATCATCACAATCACTCTCCGA pET-28a-PaHK3b-F cgagctccgtcgacaagcttATGAGTTTGCTTCATGTATTTGGG 3 060 pET-28a-PaHK3b-R tcgagtgcggccgcaagcttTAAATTCGCACTGGAAGTTGGC -
采用EASY spin Plus植物RNA快速提取试剂盒(Aidlab,北京)提取‘84K’杨叶片总RNA,并用PrimeScript RT reagent Kit with gDNA Eraser (Perfect Real Time)(takara,日本)试剂盒进行反转录,合成单链cDNA。根据Phytozome网站上公布的毛果杨基因组信息,设计PaHK3b基因全长cDNA的PCR扩增引物PaHK3b-F和PaHK3b-R(表1),以单链cDNA为模板进行PCR扩增,扩增程序为95℃ 5 min;95℃ 30 s,57℃ 30 s,72℃ 3 min,38个循环;72℃ 7 min;4℃保温。用1.5%琼脂糖凝胶进行电泳检测,用胶回收试剂盒(Axygen,美国)回收纯化目的片段,并连接到克隆载体PLB-Vector(TIANGEN,北京)转化至大肠杆菌DH5α,涂含抗性(Amp+)平板,37℃过夜培养。挑取阳性单克隆送至生物公司(中美泰和,北京)进行序列测定。利用在线软件ExpasyProtParam(
http://web.expasy.org/protparam/ )对PaHK3b基因序列特征进行分析,使用GOR4在线工具(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_gor4.pl )进行蛋白的二级结构进行预测,利用NCBI在线工具(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )进行蛋白结构域分析。 -
将带有顶端的‘84K’杨组培苗嫩茎(2~3 cm)切下,转接到生根培养基(1/2MS+0.02 mg·L−1 NAA+0.05 mg·L−1IBA)中,培养于温度24℃、光周期16 h/8 h(光照/黑暗)、光照强度为50 μmol·m−2·s−1人工气候培养室,28 d后,选取生长状态一致的组培苗,转移到装有5 mL 1/2MS液体培养基的玻璃管(直径4 cm,高20 cm)中继续培养。培养5 d后,进行非生物胁迫处理和植物激素处理。非生物胁迫处理分别为42℃高温、0℃低温、200 mmol·L−1 NaCl和5% PEG6000处理,植物激素为脱落酸(ABA)、6-苄氨基嘌呤(6-BA)、吲哚丁酸(IBA)、赤霉素(GA3)及水杨酸(SA),浓度均为10 μmol·L−1 。NaCl、PEG6000及植物激素处理方法为将药剂添加到1/2MS液体培养基中,以培养于1/2MS培养基中的组培苗为对照。各处理的时间均为3 h,每个处理3个生物学重复,处理后取成熟叶片,液氮速冻保存于 −80℃超低温冰箱。
-
提取‘84K’杨根、茎、叶以及不同处理植株叶片总RNA,反转录为cDNA。设计PaHK3b基因实时定量PCR(qRT-PCR)引物PaHK3b-q-F和PaHK3b-q-R(表1)。将合成的cDNA稀释10倍作为实时定量PCR模板,参照TB Green TM Premix Ex TaqTM II(TliRNaseH Plus)(TaKaRa,日本)试剂盒说明配制反应体系:TB Green Premix TaqII(TliRNaseH Plus,2 × )10 μL,Primer-F(10 μmol·L−1) 0.8 μL,Primer-R(10 μmol·L−1) 0.8 μL,cDNA模板2 μL,ddH2O补足至20 μL。用Roche Light Cycle 480Ⅱ型荧光定量PCR仪(Roche,瑞士)进行qRT-PCR反应,反应程序为:预变性95℃30 s,变性95℃ 5 s,退火60℃ 30 s,40个循环;溶解曲线为95℃ 5 s,65℃ 1 min。以Actin为内参基因,采用2−ΔΔCT算法计算PaHK3b基因的相对表达量[21]。利用Excel2010和Spass23.0软件对数据进行整理制图和差异分析。
-
根据TIANGEN公司在线无缝克隆引物设计工具(http://123.56.75.19)设计含酶切位点的原核表达载体引物pET-28a-PaHK3b-F和pET-28a-PaHK3b-R(表1),PCR扩增‘84K’杨PaHK3b基因的cDNA序列,回收扩增产物。原核表达载体pET-28a(索莱宝,北京)经Hind III(Thermo Fisher Scientific,美国)单酶切,胶回收纯化,根据EasyGeno快速重组克隆试剂盒(TIANGEN,北京)操作流程,将二者回收产物进行重组反应,将重组质粒转化至大肠杆菌DH5α感受态细胞中,经抗性(Kan+)筛选,挑取单克隆进行PCR验证,并送至生物公司测序验证,提取阳性菌株质粒,获得重组载体(pET-28a-PaHK3b)并转化至表达菌株BL21(DE3)(TIANGEN,北京)。
-
将转化后的表达菌株BL21(DE3)接种于5 mL含有50 mg·L−1的卡那霉素的LB液体培养基,37℃摇床震荡培养至OD600 0.6~0.8,加入终浓度为0.5 mmol·L−1的IPTG诱导4 h,稀释菌液104倍,取5 μL至含有0、50、100、150、200 mmol·L−1 NaCl LB固体培养基上涂抹直径约为1 cm的9个点,超净台吹干菌液,37℃倒置过夜培养[22]。同时,取诱导后的产物1 mL加入到1 mL含有PEG6000的LB液体培养基中使PEG终浓度为5%(W/V),放置37℃摇床震荡培养,0~7 h内每隔1 h测1次OD600值,每个时间点测3个重复[23-25]。
-
通过PCR扩增、克隆得到‘84K’杨PaHK3b基因全长CDS。CDS全长为3060 bp,编码1019个氨基酸,蛋白分子量为113564.85 kDa,等电点为6.53,蛋白不稳定指数为37,为稳定蛋白;脂肪族指数为92.31,蛋白疏水性平均值为−0.088。蛋白主要以无规则卷曲为主,α-螺旋与延伸链则散布在蛋白中,其中,无规则卷曲416个(40.82%)、α-螺旋377个(37%)、延伸链则有226个(22.18%)。蛋白结构域分析结果表明,PaHK3b蛋白具有典型的细胞分裂素受体结构域,包含CHASE (Cyclaseand histidine kinase associated sensing extracellular)结构域、HisKA (His kinase A domain)结构域及REC(CheY-homologous receiver domain)结构域,其中,CHASE结构域使细胞分裂素与其相应的受体结合,HisKA结构域感知信号和His残基自身磷酸化,REC结构域与磷酸基团相结合,并向下游的信号蛋白进行传递。
克隆得到全长1600 bp的‘84K’杨PaHK3b基因启动子序列,并采用plantCARE对其进行生物信息学分析。结果表明:PaHK3b启动子不仅具有TATA-box和CAAT-box核心基本元件,还包括G-box、GA-motif、TCT-motif等光响应元件。另外,PaHK3b启动子区域还包含多个与逆境响应及激素调控相关的顺式作用元件,如低温响应元件LTR、防御与胁迫响应元件TC-rich repeats、赤霉素响应元件GARE-motif、水杨酸响应元件TCA-element等(表2),表明该基因可能参与杨树逆境胁迫及激素信号响应。
表 2 PlantCARE预测PaHK3b启动子的主要顺式作用元件
Table 2. The main cis-elements in PaHK3b promoter predicted by PlantCARE
顺式作用元件
Cis-element起始位点(方向)
Start site/bp(direction)序列
Sequence特性
CharacteristicARE 1 150(+) AAACCA 厌氧诱导 Essential for the anaerobic induction GARE-motif 346(−) TCTGTTG 赤霉素响应 Gibberellin-responsive element LTR 318(−) CCGAAA 低温响应 Low-temperature responsiveness TC-rich repeats 88(+) ATTCTCTAAC 防卫及胁迫响应 Defense and stress responsiveness TCA-element 288(−)/818(+) CCATCTTTTT 水杨酸响应 Salicylic acid responsiveness GATA-motif 171(−) GATAGGA 光响应元件 Light responsive element GATA-motif 439(+) GATAGGG TCCC-motif 1511(+) TCTCCCT TCT-motif 1 462(+)/1 551(−) TCTTAC GT1-motif 677(+) GTGTGTGA GT1-motif 1 409(−)/1 353(−) GGTTAA GA-motif 706(+) ATAGATA G-box 9(−) CACGAC CAAT-box 619(−)/575(−)/993(−)/103(−) CCAAT 启动子和增强子区域常见的顺式作用元件
Common cis-acting element in promoter and enhancer regionsTATA-box 27(+)/195(+)/150(+)/1 067(−) TATA 核心启动子
Core promoter element around −30 of transcription start148(−)/1 065(+)/193(−) TATAAA 147(−) TATAAAT 136(+) ATATAA 1 109(−) ccTATAAAaa 192(−) TATAAAA -
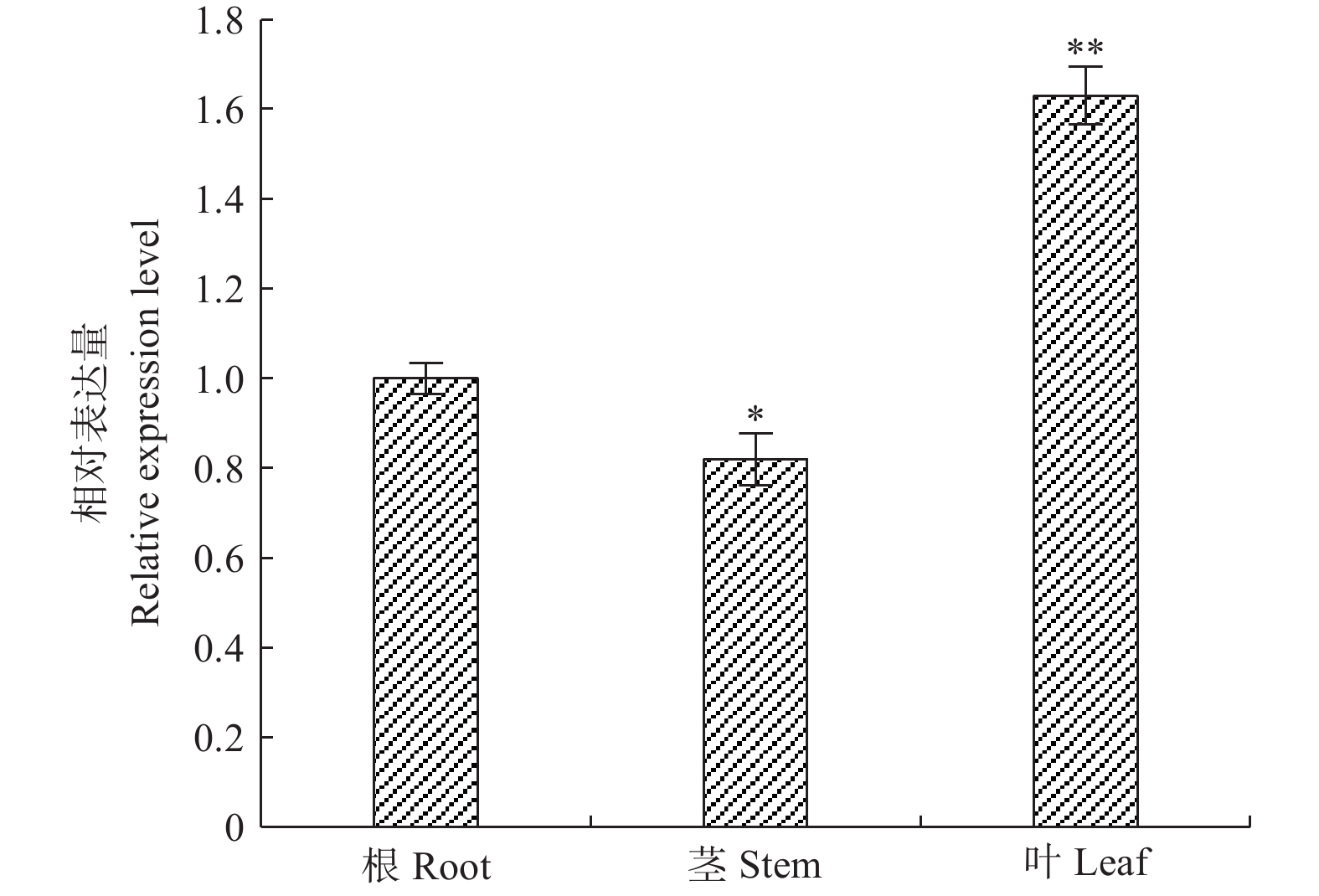
采用qRT-PCR技术对‘84K’杨PaHK3b基因在根、茎、叶的表达进行检测。结果表明:该基因在根、茎、叶中均有表达,以根表达量作为参照,茎表达量低于根组织,为根表达量的82%,叶片的表达量高于根,为根表达量的1.63倍(图1)。以上结果表明,PaHK3b基因在‘84K’杨叶片表达量最高。
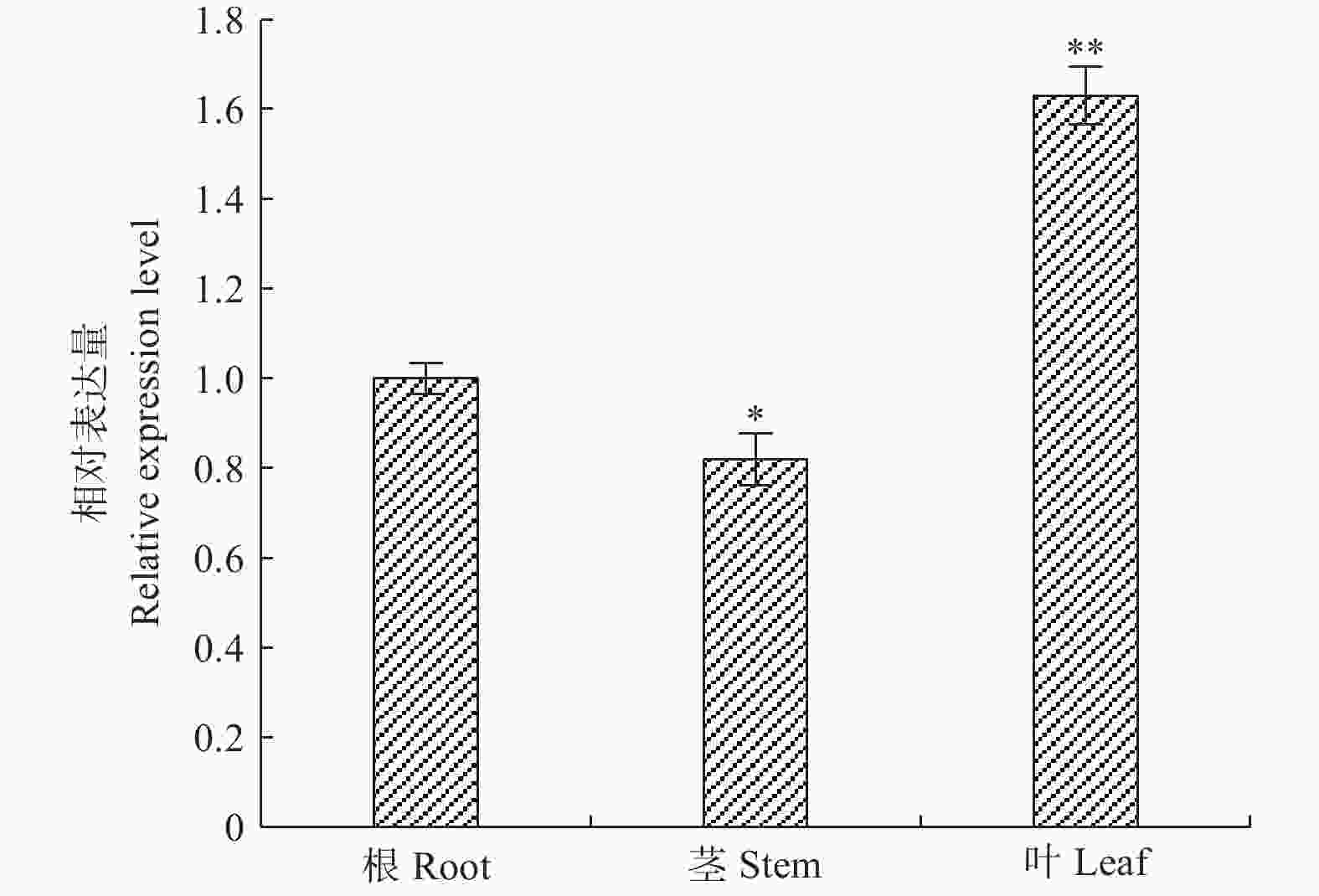
图 1 ‘84K’杨PaHK3b基因在根、茎、叶片中的表达差异
Figure 1. Expression difference of PaHK3b gene in root,stem and leaf of ‘84K’ poplar
-
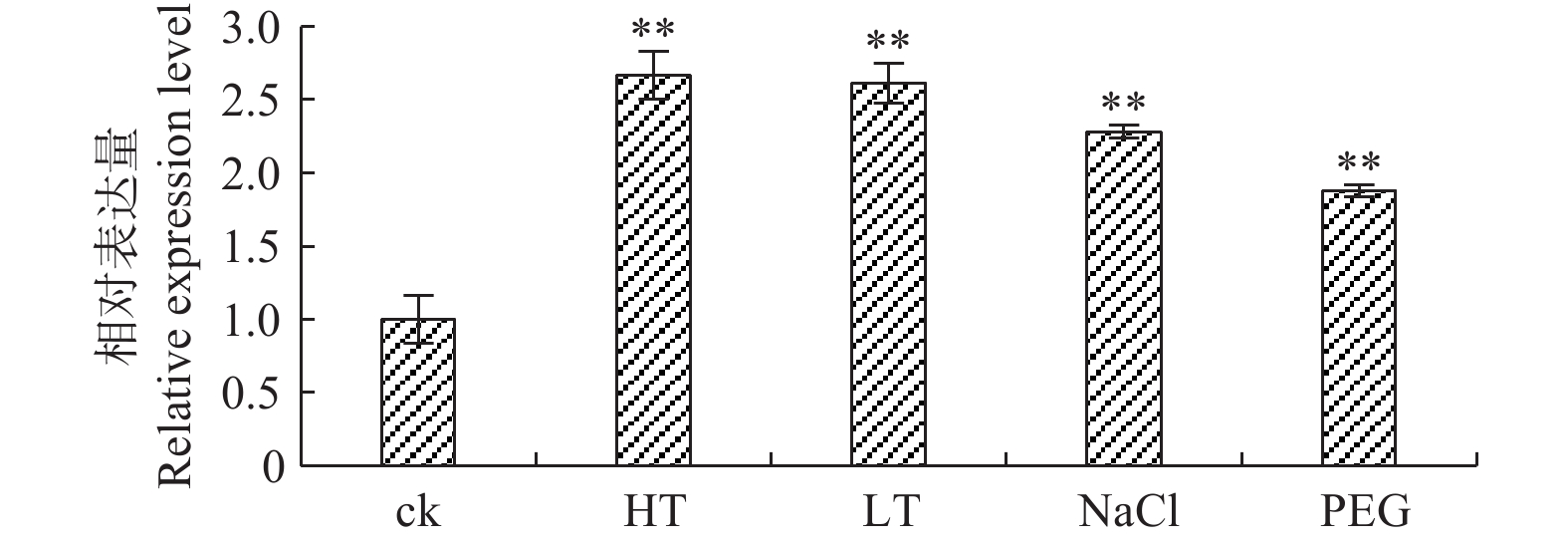
高温、低温、盐胁迫和干旱胁迫条件下,‘84K’杨PaHK3b基因表达量均明显高于对照,并且温度胁迫下PaHK3b基因表达量高于盐胁迫和干旱胁迫下的表达量(图2),其中,42℃高温胁迫、0℃低温胁迫时,PaHK3b基因表达量分别为对照的2.67、2.61倍;200 mmol·L−1 NaCl胁迫、5%PEG6000胁迫时,PaHK3b基因表达量分别为对照的2.28、1.87倍。以上结果说明,PaHK3b基因参与了非生物胁迫响应,在杨树的逆境胁迫反应过程中起作用。
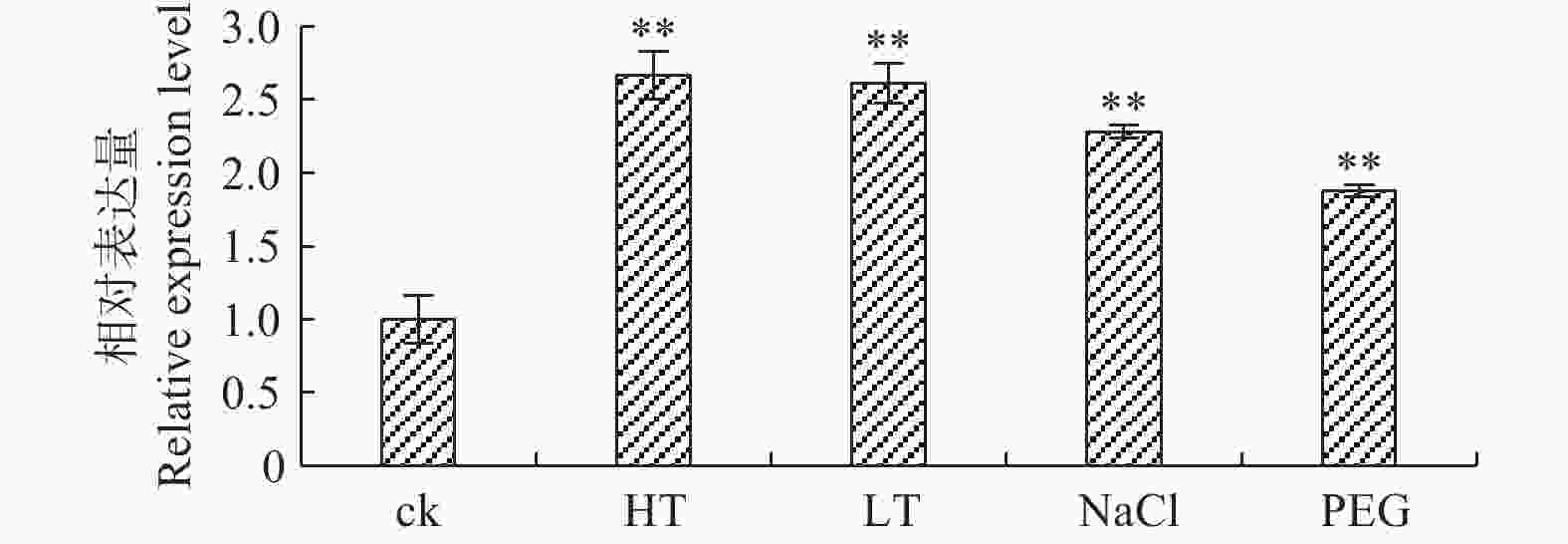
图 2 非生物胁迫处理下‘84K’杨叶片PaHK3b基因的表达差异
Figure 2. Expression difference of PaHK3b in leaf of ‘84K’ poplar under abiotic stresses
-
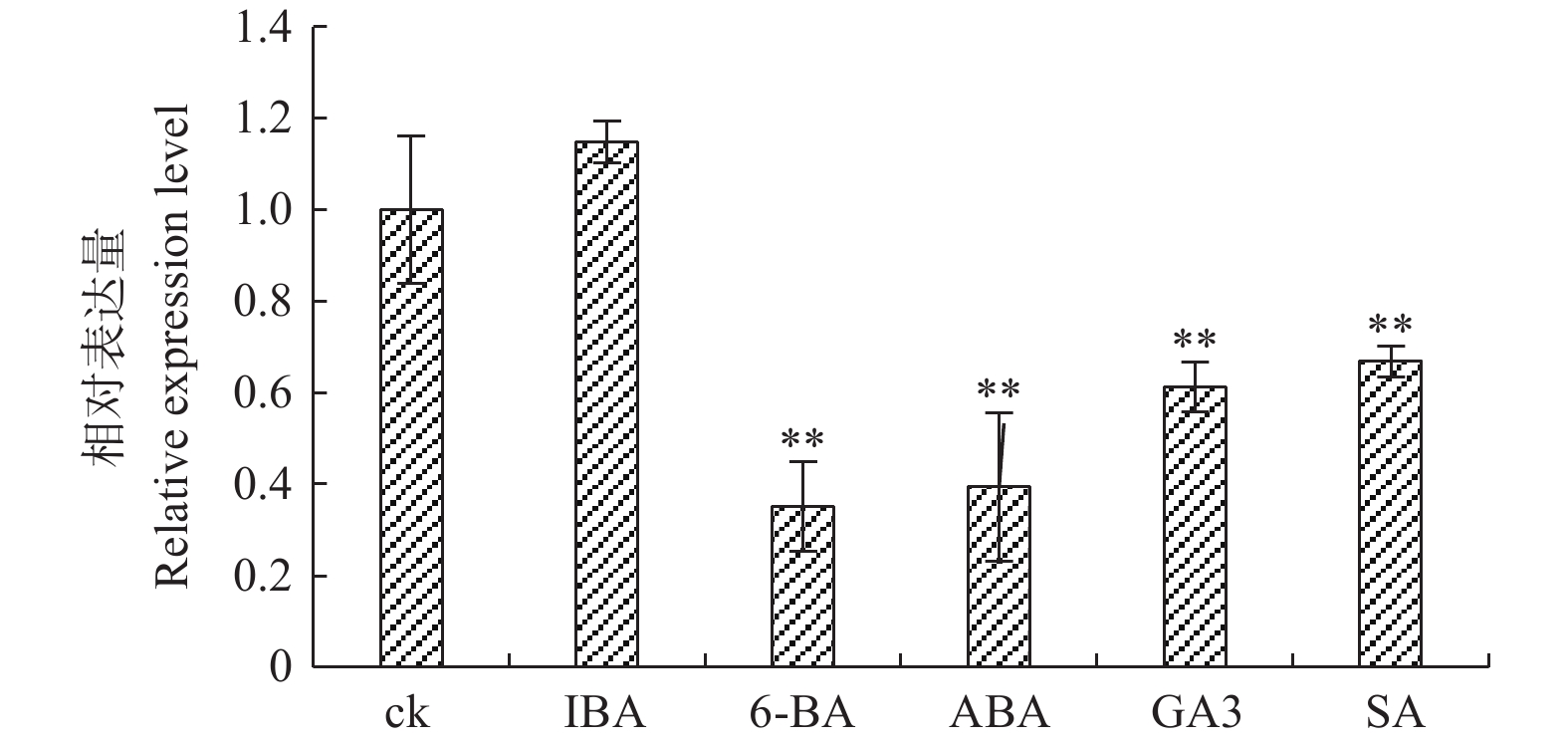
图3表明:PaHK3b基因对不同植物激素的响应有差异。生长素IBA处理时,PaHK3b基因表达量与对照相似,而其他激素处理时,PaHK3b基因表达量均不同程度的低于对照,其中,6-BA、ABA、GA3、SA处理时,PaHK3b基因表达量分别为对照的35%、39%、61%、66%。可见,外源赤霉素、脱落酸、细胞分裂素及水杨酸均能抑制杨树PaHK3b基因的表达,表明PaHK3b基因参与植物激素响应。
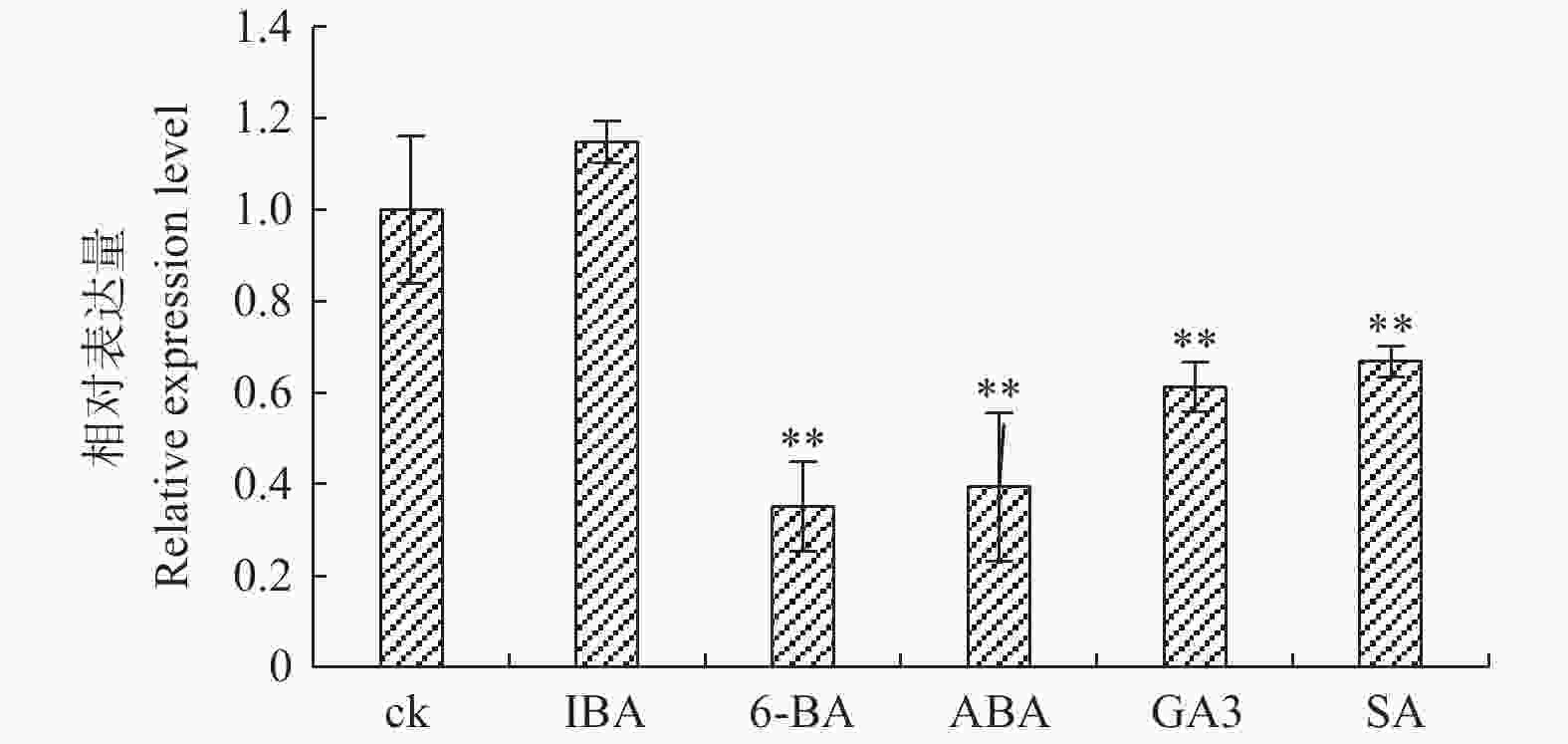
图 3 不同植物激素处理下‘84K’杨叶片PaHK3b基因的表达差异
Figure 3. Expression difference of ‘84K’PaHK3b under several plant hormone treatments
-
利用斑点法对PaHK3b基因的抗盐性进行了检测。结果表明:在未添加NaCl的LB固体培养基上,转入pET-28a-PaHK3b重组质粒的大肠杆菌BL21菌株(重组菌株)与转入pET-28a空载体的菌株(对照菌株)生长速度基本一致。在含有50、100、150 mmol·L−1的NaCl LB固体培养基上,重组菌株与对照菌株生长均受到抑制,且随着NaCl浓度的增加菌落生长减少;但重组菌生长速度较对照快,菌落多且菌斑大。当培养基中NaCl浓度为200 mmol·L−1时,2种菌株均停止生长(图4)。因此,PaHK3b基因通过原核表达能够提高大肠杆菌菌株的抗盐能力。
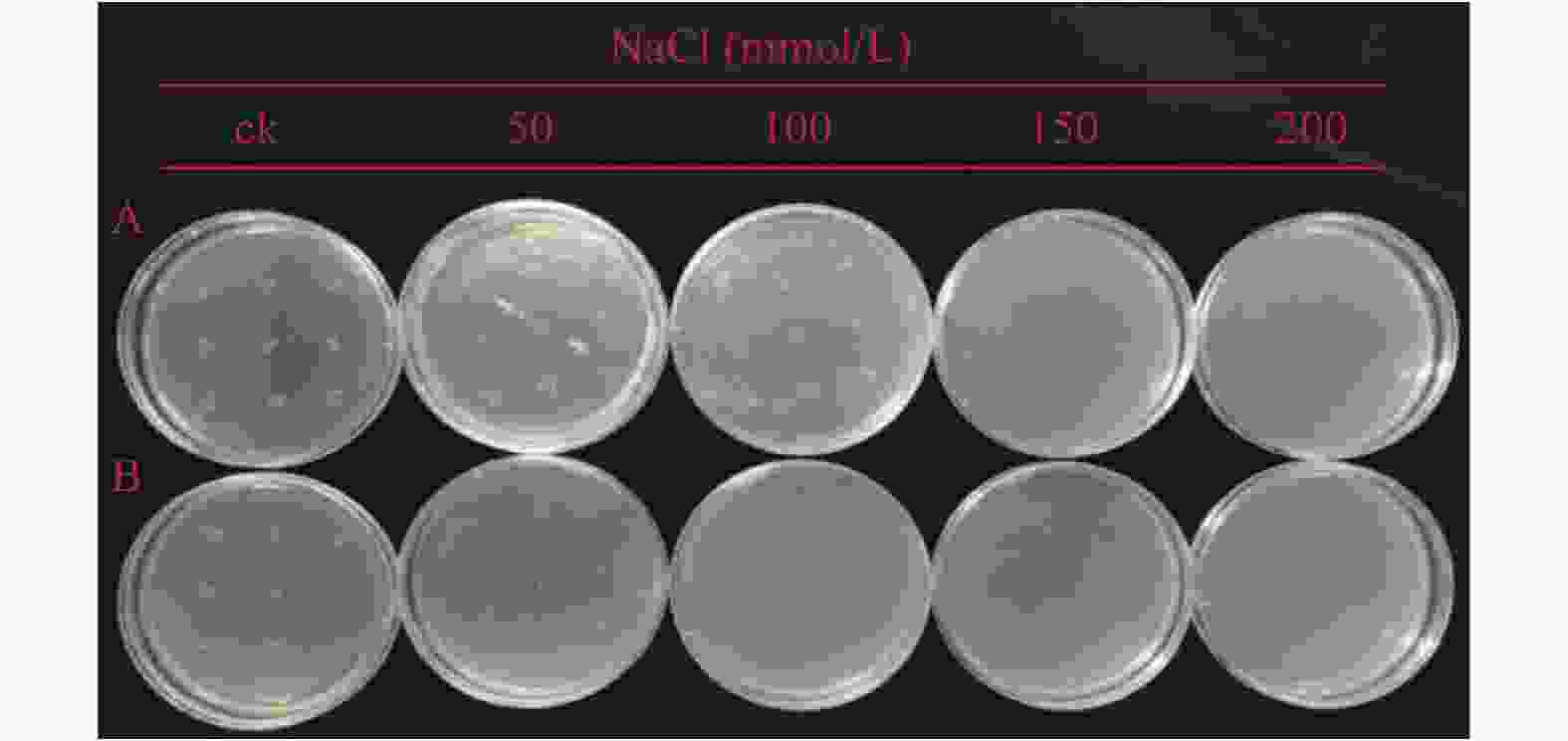
图 4 大肠杆菌E.coli BL21(pET-28a)和E.coli BL21(pET-28a-PaHK3b)菌株耐盐性差异
Figure 4. Difference in salt tolerance of E.coli BL21(pET-28a)and E.coli BL21 (pET-28a-PaHK3b)
利用PEG模拟干旱胁迫对PaHK3b基因的抗旱性进行评价,结果表明:在LB液体培养基中,重组菌株的生长速度较对照菌株要快;在添加5%PEG6000的LB液体培养基中,重组菌株和对照菌株生长均受到抑制。培养1 h时,重组菌株和对照菌株生长没有差异,之后转入pET-28a-PaHK3b重组质粒的菌株较转入pET-28a空载的大肠杆菌生长速度快,培养7 h后,含重组质粒菌株的OD600值是初始的1.8倍,含空载质粒菌株的OD600值是初始的1.1倍(图5)。因此,PaHK3b基因原核表达不仅能够提高大肠杆菌菌株生长速度,而且还能够提高其抗脱水能力。
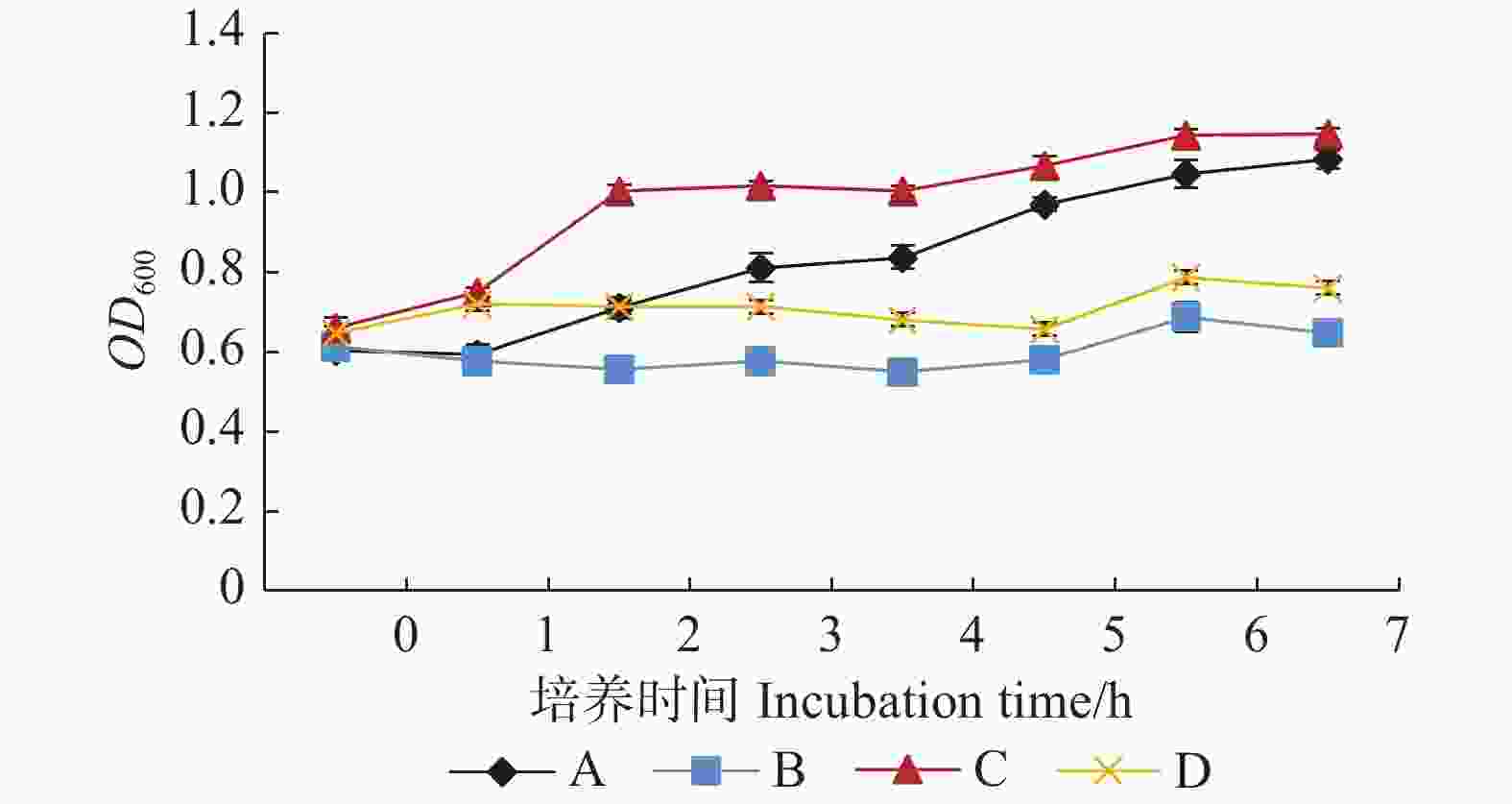
图 5 大肠杆菌E.coli BL21(pET-28a)和E.coli BL21(pET-28a-PaHK3b)菌株在LB及LB-5%PEG6000液体培养基生长曲线
Figure 5. Growth curve of E.coli BL21(pET-28a)and E.coli BL21(pET-28a-PaHK3b)in LB and LB-5%PEG6000 liquid medium respectively
-
组氨酸激酶基因在植物激素信号响应及非生物逆境胁迫响应中发挥重要作用[3, 26]。如拟南芥组氨酸激酶AHK3参与其对干旱、高盐及冷胁迫的抗逆反应[3-4],转玉米(Zea mays L.)组氨酸激酶基因ZmHK9的拟南芥抗旱性明显增强[27]。本研究中,在高温、低温、高盐和PEG模拟干旱等处理条件下,PaHK3b基因表达量均增加,其中,基因上调程度呈高温>低温>盐胁迫>干旱胁迫的模式;在6-BA、ABA、GA3及SA处理条件下,PaHK3b基因表达量均下降,基因下调程度呈6-BA>ABA>GA3>SA的模式。因此表明,PaHK3b基因能够不同程度的参与植物激素信号响应及非生物胁迫信号响应。另外,SA是植物防卫反应的重要内源植物激素,作为一种信号分子,在植物生物胁迫响应过程中发挥重要作用[28-29]。ABA在植物抗逆胁迫反应及在生物胁迫和非生物胁迫中均具有重要调控作用。因此,PaHK3b基因可能通过ABA及SA途径的负调节提高了杨树对逆境胁迫的耐受性。
通过分析PaHK3b基因CDS序列发现,‘84K’杨组氨酸激酶PaHK3b蛋白具有典型的CHASE细胞分裂素受体结构域。在植物外源激素6-BA处理下,PaHK3b基因下调表达,表明该基因能够感知细胞分裂素信号并参与响应,这与组氨酸激酶基因具有CHASE结构域,通过该结构域与细胞分裂素的结合,感知植物激素信号并进行传导是一致的[7, 10]。对PaHK3b基因启动子序列分析发现,该区域包含低温响应、防御与胁迫响应、赤霉素响应、水杨酸响应等多个参与胁迫响应及激素调控的顺式作用元件。这与组氨酸激酶基因启动子区域有多个植物激素信号响应及非生物逆境胁迫相关的顺式作用元件是一致的[30-31]。另外,结合在植物激素6-BA、ABA、GA3、SA和非生物胁迫高温、低温、高盐、PEG处理下,PaHK3b基因均不同程度参与响应的表达,表明PaHK3b基因参与植物激素信号响应及非生物胁迫响应与其基因结构密切相关。
已有研究表明,通过构建基因的原核表达载体,并利用大肠杆菌表达系统可以对基因的功能进行初步鉴定。如将小麦(Triticum aestivum L.)脱水蛋白编码基因——DHN14基因的原核表达载体转入大肠杆菌后,大肠杆菌菌株对重金属胁迫及过氧化氢胁迫的耐受性显著增强,推测小麦脱水蛋白基因DHN14在其响应非生物胁迫的过程中起作用[23]。转入谷子(Setaria italica L.)类受体蛋白激酶基因SiRLK35原核表达载体的大肠杆菌菌株生长状态较阴性对照好,同时获得的转SiRLK35基因水稻植株对盐胁迫的耐受性高于对照[22]。以上研究说明,原核表达结果能对克隆基因功能进行有效鉴定。在本研究中,转入PaHK3b基因原核表达载体的菌株在50~150 mmol·L−1 NaCl的LB固体培养基及5% PEG6000的LB液体培养基中生长状态均好于转入空载体的对照菌株。因此可得,杨树PaHK3b基因能够促进大肠杆菌的生长,同时能够提高其耐盐和抗旱能力,推测该基因可能在杨树生长及抗逆境胁迫的过程中发挥作用。
-
本研究从‘84K’杨中克隆了组氨酸激酶基因PaHK3b及其启动子,生物信息学分析表明,该基因编码蛋白具有典型的细胞分裂素受体结构域,其启动子区域有多个参与胁迫响应和激素信号转导的顺式作用元件。在高温、低温、高盐和PEG模拟干旱等非生物胁迫下,PaHK3b基因表达量增加,而在6-BA、ABA、GA3及水杨酸等植物激素处理条件下,其表达量下降。原核表达PaHK3b基因可以提高受体大肠杆菌的生长及抗旱耐盐能力。初步生物学功能研究表明,该基因参与杨树植物激素信号响应,并在其抗逆境胁迫过程中发挥重要调控作用。本研究结果为深入研究PaHK3b基因在激素信号转导及逆境胁迫下杨树生长发育的调控作用提供线索,为杨树抗逆分子育种及品种改良奠定基础。
‘84K’杨组氨酸激酶基因PaHK3b的克隆及功能分析
Cloning and Functional Analysis of Histidine Kinase Gene PaHK3b in Poplar '84K'
-
摘要:
目的 本研究克隆了银腺杨‘84K’(Populus alba × P. glandulosa ‘84K’)组氨酸激酶基因PaHK3b启动子及编码区,并对其表达进行检测及功能鉴定,为深入研究PaHK3b基因在杨树生长发育的调控作用提供线索,为杨树分子育种及品种改良奠定基础。 方法 根据毛果杨(P. trichocarpa Torr. & Gray)基因组信息,设计引物克隆‘84K’杨组氨酸激酶基因PaHK3b启动子及CDS序列,并对其保守结构域和启动子顺式作用元件进行分析;同时,对‘84K’杨进行植物激素处理(10 μmol·L−1ABA、10 μmol·L−16-BA、10 μmol·L−1 IBA、10 μmol·L−1GA3及10 μmol·L−1 水杨酸(SA))及非生物胁迫处理(42℃高温、0℃低温、200 mmol·L−1 NaCl和5% PEG6000),利用实时定量PCR(qRT-PCR)方法检测PaHK3b基因表达情况与表达响应差异,并采用原核表达方法初步确定PaHK3b基因的生物学功能。 结果 PaHK3b基因编码框区长度为3060 bp,编码1019个氨基酸,PaHK3b蛋白具有CHASE、HisKA和REC等典型的细胞分裂素受体结构域。PaHK3b基因启动子序列中不仅含有大量TATA框和CAAT框常见核心元件,还包含低温响应元件LTR、防御与胁迫响应元件TC-rich repeats、赤霉素响应元件GARE-motif、水杨酸响应元件TCA-element等顺式作用元件,这些元件与杨树的激素响应和逆境胁迫响应密切相关。qRT-PCR分析表明:PaHK3b基因在叶片中表达最高,根部中等,茎中最少;另外,与正常条件下相比,在高温、低温、NaCl及PEG处理时,PaHK3b基因表达量与对照明显增高,分别为对照的2.67、2.61、2.28、1.87倍;用IBA诱导处理时,基因表达量与对照相比差异不大,而在6-BA、ABA、GA3及SA处理时,基因表达量与对照相比均呈下调表达;在添加5%PEG6000的LB液体培养基中,转入PaHK3b基因原核表达载体的大肠杆菌菌株生长速度显著高于对照,在添加50~150 mmol·L−1NaCl的LB固体培养基上,转入PaHK3b基因原核表达载体的大肠杆菌菌株单克隆生长均好于对照。 结论 ‘84K’杨PaHK3b基因启动子含有逆境和激素响应元件,表明PaHK3b基因与杨树植物激素信号及非生物胁迫信号响应密切相关。经非生物胁迫处理、激素处理及原核表达证实,杨树PaHK3b基因参与杨树植物激素信号响应,并在其抗逆境胁迫过程中发挥重要调控作用。 Abstract:Objective Using Poplar '84K' (Populus alba × P. glandulosa) as material to clone the promoter region and coding sequence (CDS) of histidine kinase gene PaHK3b, then to detect its expression and identify its functions, so as to provide clues for analyzing the function of PaHK3b in growth and development regulation of poplars and for molecular breeding and genetic improvement of poplar. Method Based on the published genome information of P. trichocarpa Torr. & Gray, the promoter region and CDS were cloned using the specific primers designed. The conservative structure domain and promoter cis-acting element were analyzed. At the same time, in vitro plants of Poplar '84K' were treated with several plant hormones (10 μmol·L−1 ABA, 10 μmol·L−1 6-BA, 10 μmol·L−1 IBA, 10 μmol·L−1 GA3, and 10 μmol·L−1 SA) under various abiotic stresses (42℃, 0℃, 200 mmol·L−1 NaCl, and 5% PEG6000). Differences in expression and response of PaHK3b gene were detected. Furthermore, the prokaryotic expression system was used to study the biological function of PaHK3b gene in vitro. Result The PaHK3b gene was 3060 bp in length and encoded 1019 amino acids. PaHK3b protein had a typical cytokinin receptor domain, a CHASE domain, a HisKA domain, and a REC domain. Besides, a large number of common core elements of TATA box and CAAT box, a number of stress-related and hormone-related regulatory elements, such as low temperature response element LTR, defense and stress responsive element TC-rich repeats, gibberellin responsive element GARE-motif, salicylic acid responsive element TCA-element etc. were predicted in the promoter sequence of PaHK3b gene, that closely related to the plant hormone and stress response regulations. The qRT-PCR results showed that PaHK3b gene was expressed the highest in leaf, medium in root, and least in stem. The transcriptional levels of PaHK3b gene were about 2.67, 2.61, 2.28, 1.87 times that of the control respectively under 42℃, 0℃, 200 mmol·L−1 NaCl, and 5% PEG treatments. Under IBA treatment, the transcripts of PaHK3b were not significantly different from that of the control. The expression of PaHK3b were down-regulated under the treatments of 6-BA, ABA, GA3 and SA respectively. The growth of the Escherichia coli strain with PaHK3b prokaryotic expression construction in LB liquid medium supplemented with 5% PEG6000 were faster than that of the control. Meanwhile, the growth of the E. coli strain with PaHK3b prokaryotic expression construction on solid LB contained 50~150 mmolL−1 NaCl was better than that of the control. Conclusion The promoter of poplar '84K' PaHK3b gene contained cis-regulatory elements, such as stress response element and hormone responsive element etc, showing that PaHK3b gene is closely related to the response of poplar to hormones and abiotic stress. It is confirmed by abiotic stress treatment, hormone treatment and prokaryotic expression that PaHK3b gene participates in the signal response of plant hormones and plays an important role in poplar response to abiotic stresses. -
图 1 ‘84K’杨PaHK3b基因在根、茎、叶片中的表达差异
Figure 1. Expression difference of PaHK3b gene in root,stem and leaf of ‘84K’ poplar
图 2 非生物胁迫处理下‘84K’杨叶片PaHK3b基因的表达差异
Figure 2. Expression difference of PaHK3b in leaf of ‘84K’ poplar under abiotic stresses
图 3 不同植物激素处理下‘84K’杨叶片PaHK3b基因的表达差异
Figure 3. Expression difference of ‘84K’PaHK3b under several plant hormone treatments
图 4 大肠杆菌E.coli BL21(pET-28a)和E.coli BL21(pET-28a-PaHK3b)菌株耐盐性差异
Figure 4. Difference in salt tolerance of E.coli BL21(pET-28a)and E.coli BL21 (pET-28a-PaHK3b)
图 5 大肠杆菌E.coli BL21(pET-28a)和E.coli BL21(pET-28a-PaHK3b)菌株在LB及LB-5%PEG6000液体培养基生长曲线
Figure 5. Growth curve of E.coli BL21(pET-28a)and E.coli BL21(pET-28a-PaHK3b)in LB and LB-5%PEG6000 liquid medium respectively
表 1 相关引物序列及产物大小
Table 1. The sequences and PCR product size of primers used in this study
基因 Gene 引物序列 Primer sequence (5'→3') 产物长度 Product length/bp PaHK3b-F ATGAGTTTGCTTCATGTATTTGGG 3 060 PaHK3b-R TAAATTCGCACTGGAAGTTGGC PaHK3b-pro-F GTGGCTTTGGGAGGAAGGAAT 1 600 PaHK3b-pro-R CCACATCTTGCCACCATCACC PaHK3b-q-F ATGAGTTTGCTTCATGTATTTGG 243 PaHK3b-q-R CTATAAATTCGCACTGGAAGTTG Actin-F AAACTGTAATGGTCCTCCCTCCG 193 Actin-R GCATCATCACAATCACTCTCCGA pET-28a-PaHK3b-F cgagctccgtcgacaagcttATGAGTTTGCTTCATGTATTTGGG 3 060 pET-28a-PaHK3b-R tcgagtgcggccgcaagcttTAAATTCGCACTGGAAGTTGGC  下载: 导出CSV
下载: 导出CSV
表 2 PlantCARE预测PaHK3b启动子的主要顺式作用元件
Table 2. The main cis-elements in PaHK3b promoter predicted by PlantCARE
顺式作用元件
Cis-element起始位点(方向)
Start site/bp(direction)序列
Sequence特性
CharacteristicARE 1 150(+) AAACCA 厌氧诱导 Essential for the anaerobic induction GARE-motif 346(−) TCTGTTG 赤霉素响应 Gibberellin-responsive element LTR 318(−) CCGAAA 低温响应 Low-temperature responsiveness TC-rich repeats 88(+) ATTCTCTAAC 防卫及胁迫响应 Defense and stress responsiveness TCA-element 288(−)/818(+) CCATCTTTTT 水杨酸响应 Salicylic acid responsiveness GATA-motif 171(−) GATAGGA 光响应元件 Light responsive element GATA-motif 439(+) GATAGGG TCCC-motif 1511(+) TCTCCCT TCT-motif 1 462(+)/1 551(−) TCTTAC GT1-motif 677(+) GTGTGTGA GT1-motif 1 409(−)/1 353(−) GGTTAA GA-motif 706(+) ATAGATA G-box 9(−) CACGAC CAAT-box 619(−)/575(−)/993(−)/103(−) CCAAT 启动子和增强子区域常见的顺式作用元件
Common cis-acting element in promoter and enhancer regionsTATA-box 27(+)/195(+)/150(+)/1 067(−) TATA 核心启动子
Core promoter element around −30 of transcription start148(−)/1 065(+)/193(−) TATAAA 147(−) TATAAAT 136(+) ATATAA 1 109(−) ccTATAAAaa 192(−) TATAAAA
下载: 导出CSV
-
[1] Muller B, Sheen J. Arabidopsis cytokinin signaling pathway[J]. Science Signaling STKE, 2007(407): cm5. [2] Werner T, Schmulling T. Cytokinin action in plant development[J]. Current Opinion in Plant Biology, 2009, 12(5): 527-538. doi: 10.1016/j.pbi.2009.07.002 [3] Tran L S, Urao T, Qin F, et al. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis[J]. PNAS, 2007, 104(51): 20623-20628. doi: 10.1073/pnas.0706547105 [4] Jeon J, Kim N Y, Kim S, et al. A subset of cytokinin two-component signaling system plays a role in cold temperature stressresponse in Arabidopsis[J]. Journal of Biological Chemistry, 2010, 285(30): 23371-23386. doi: 10.1074/jbc.M109.096644 [5] Maruyama-Nakashita A, Nakamura Y, Yamaya T, et al. A novel regulatory pathway of sulfate uptake in Arabidopsis roots: implication of CRE1/WOL/AHK4 mediated cytokinin-dependent regulation[J]. Plant Journal, 2004, 38(5): 779-789. doi: 10.1111/j.1365-313X.2004.02079.x [6] Chefdor F, Héricourt F, Koudounas K, et al. Highlighting type A RRs as potential regulators of the dkHK1 multi-step phosphorelay pathway in Populus[J]. Plant Science Elsevier, 2018, 277: 68-78. doi: 10.1016/j.plantsci.2018.09.010 [7] Hwang I, Chen H C, Sheen J. et al. Two-component signal transduction pathways in Arabidopsis[J]. Plant Physiol, 2002, 129(5): 500-515. [8] Schaller F, Zerbe P, Reinbothe S, et al. The allene oxide cyclase family of Arabidopsis thaliana localization and cyclization[J]. Febs Journal, 2008, 275: 2428-2441. doi: 10.1111/j.1742-4658.2008.06388.x [9] Shan X, Yan J, Xie D, et al. Comparison of phytohormone signaling mechanisms[J]. Curr Opi Plant Biol, 2012, 15(10): 84-91. [10] Hwang I, Sheen J, Müller B, et al. Cytokinin signaling networks[J]. Annual Review Plant Biology, 2012, 63(1): 353-380. doi: 10.1146/annurev-arplant-042811-105503 [11] Argyros R D, Mathews D E, Chiang Y H, et al. Type B response regulators of Arabidopsis play key roles in cytokinin signaling and plant development[J]. Plant Cell, 2008, 20(8): 2012-2116. [12] Masayuki H, Melissa S. Pischke, et al. In planta functions of the Arabidopsis cytokinin receptor family[J]. PNAS, 2004, 101(23): 8821-8826. doi: 10.1073/pnas.0402887101 [13] Kim H J, Ryu H, Hong S H, et al. Cytokinin-mediated control of leaf longevity by AHK3 through phosphorylation of ARR2 in Arabidopsis[J]. Proc Natl Acad Sci USA, 2006, 103(3): 814-819. doi: 10.1073/pnas.0505150103 [14] Riefler M, Novsk O, Strnad M, et al. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism[J]. Plant Cell, 2006, 18(1): 40-54. doi: 10.1105/tpc.105.037796 [15] Dello I R, Linhares F S, Scacchi E, et al. Cytokinins determine Arabidopsis root meristem size by controlling cell differentiation[J]. Current Biology, 2007, 17(8): 678-682. doi: 10.1016/j.cub.2007.02.047 [16] Bartrina I, Jensen H, Novák O, et al. Gain-of-function mutants of the cytokinin receptors AHK2 and AHK3 regulate plant organ size, flowering time and plant longevity[J]. Plant Physiology, 2007, 173(3): 1783-1797. [17] Kang N Y, Cho C, Kim N Y, et al. Cytokinin receptor-dependent and receptor-independent pathways in the dehydration response of Arabidopsis thaliana[J]. Plant Physiol, 2012, 169(14): 1382-1391. doi: 10.1016/j.jplph.2012.05.007 [18] Cortleven A, Nitschke S, Klaumünzer M, et al. A novel protective function for cytokinin in the light stress response is mediated by the Arabidopsis histidine kinase2 and Arabidopsis histidine kinase3 receptors[J]. Plant Physiology, 2014, 164(3): 1470-1483. doi: 10.1104/pp.113.224667 [19] Tran L S, Shinozaki K, Yamaguchi-Shinozaki K, et al. Role of cytokinin responsive two-component system in ABA and osmotic stress signaling[J]. Plant Signal & Behavior, 2010, 5(2): 148-150. [20] Nieminen K, Immanen J, Laxell M, et al. Cytokinin signaling regulates cambial development in poplar[J]. Proceedings of the National Academy of Sciences, 2008, 105(50): 20032-20037. doi: 10.1073/pnas.0805617106 [21] Sun J, Niu Q W, Tarkowski P, et al. The Arabidopsis AtIPT8/PGA22 gene encodes an isopentenyl transferase that is involved in de novo cytokinin biosynthesis[J]. Plant Physiology, 2003, 131(1): 167-176. doi: 10.1104/pp.011494 [22] 王一帆, 李 臻, 潘教文, 等. 谷子SiRLK35基因克隆及功能分析[J]. 遗传, 2017, 39(5):413-422. [23] 史学英, 田 野, 李 核, 等. 小麦K2型脱水蛋白DHN14响应非生物胁迫的功能分析[J]. 西北农林科技大学学报, 2019, 47(5):1-10. [24] 林 琳, 李 健, 李慧玉, 等. 逆境胁迫下柽柳脂质转运蛋白基因(ThLTP)的克隆与功能初步分析[J]. 林业科学研究, 2012, 25(4):492-499. doi: 10.3969/j.issn.1001-1498.2012.04.013 [25] 雷世康, 徐 刚. 麻疯树BRI1基因的鉴定及其在不同发育时期花蕾中的表达分析[J]. 林业科学研究, 2019, 32(5):121-129. [26] Ramsong N, Praveen S, Ratna K, et al. Histidine kinases in plants: Cross talk between hormone and stress responses[J]. Plant Signaling & Behavio, 2012, 7(10): 1230-1237. [27] Wang B, Guo B, Xie X, et al. A novel histidine kinase gene, ZmHK9, mediate drought tolerance through the regulation of stomatal development in Arabidopsis[J]. Gene, 2012, 501(2): 171-179. doi: 10.1016/j.gene.2012.04.012 [28] Bilal A, Farrukh A, Muhammad A A, et al. Genome-wide identification and expression analysis of two component system genes in Cicer arietinum[J]. Genomics, 2020, 112(2): 1371-1382. doi: 10.1016/j.ygeno.2019.08.006 [29] He Y, Liu X, Ye L, et al. Genome-wide identification and expression analysis of two-component system genes in tomato[J]. International journal of molecular sciences, 2016, 17(8): 1204. doi: 10.3390/ijms17081204 [30] Vernooij B, Friedrich L, Weymann K, et al. A central role of salicylic acid in plant disease resistance[J]. Science (Washington D C), 1994, 266(5188): 1247-1250. doi: 10.1126/science.266.5188.1247 [31] Yusuf M, Hasan S A, Ali B, et al. Effect of salicylic acid on salinity-induced changes inBrassica juncea[J]. Journal of Integrative Plant Biology, 2008, 50(9): 1096-1102. doi: 10.1111/j.1744-7909.2008.00697.x -

 点击查看大图
点击查看大图
计量
- 文章访问数: 7385
- HTML全文浏览量: 2979
- PDF下载量: 109
- 被引次数: 0
