




-
新疆野苹果(Malus sieversii (Ledeb.)Roem.)作为天山野果林的主要组成部分,在国内主要分布于新疆伊犁河谷地区[1]。该种是苹果(Malus domestica Borkh.)种植产业的重要选种来源[2],在品质改良、选育、良种驯化、杂交育种等生产实践和科学研究中占有重要基础地位[3-4]。然而自从1993年苹小吉丁虫(Agrilus mali Matsumura.)传入到新疆以后,对当地苹果树造成巨大的破坏,造成大量野苹果树枯死,使伊犁野苹果林和栽培果园遭受巨大破坏,且危害日益严重[5-6]。2003年开始在巩留县大面积发生[7]。
苹小吉丁虫在伊犁一年发生一代[8],以幼虫钻蛀枝条、主干的韧皮部和木质部进行危害,危害处通常会出现凹陷、失水及流胶等现象。成虫飞翔能力一般,活跃程度受温度影响,喜欢围绕树冠飞行[9]。成虫羽化后在羽化处啃咬一个“D”字状羽化孔后爬出,羽化出孔时间于6月中旬开始[6],羽化盛期为7月中旬[10],且具有取食苹果树叶片进行补充营养的习性[11]。苹小吉丁虫作为威胁伊犁乃至新疆林业生态建设和林果产业健康发展的重要害虫,做好监测预警工作至关重要。
昆虫的扩散行为通常会出现于昆虫种群数量上升后,一般以空间范围或距离单位作为扩散指标,同时,在单位时间内的扩散速度可作为昆虫扩散能力的评价依据[12]。以诱捕器作为回收装置,通过使用“标记-重捕”法,对埃及伊蚊(Aedes aegypti)的扩散行为研究发现,该蚊具有向西北方向进行扩散的趋势[13];葡萄浆果蛾(Endopiza viteana)的雄蛾在试验田间最远可自主扩散58.2 m,雌蛾可自主扩散41.2 m[14]。马铃薯甲虫(Leptinotarsa decemlineata)雌虫的扩散速度普遍大于雄虫,而扩散方向较为随机,无明显规律[15];光肩星天牛(Anoplophora glabripennis)种群扩散具有一定的方向性,且受风向影响较为明显[16]。
苹小吉丁虫的远距离扩散主要通过苗木调运等途径,而近距离传播主要通过其自身的飞行进行主动扩散,但是苹小吉丁随苗木调运等传播至果园后的扩散行为尚未见文献报导。而目前已有研究发现,浅绿色和蛋黄色粘虫板对苹小吉丁的诱捕作用较强[17],且该两种颜色粘虫板与植物源挥发物顺-3-己烯醇组合对苹小吉丁成虫具有良好的引诱效果[18],这为实现苹小吉丁成虫的扩散距离测定工作奠定了基础。通过使用“标记-重捕”法对苹小吉丁的林间扩散行为进行研究,虫源回收环节是关键,而已经研发出的以植物源挥发物为诱芯的诱捕器可为虫源回收提供技术支持。通过对苹小吉丁林间扩散能力进行测定,可以为预测苹小吉丁扩散趋势和划定监测预警范围提供科学依据。
-
试验地位于伊犁巩留县综合农场栽培果园,82°15′5076″ E;43°44′7858″ N。果园海拔高度为768.3 m,面积约为1.13 hm2,株行距约为3 m × 5 m,行为东西走向,整体呈东西长、南北短的矩形。主栽品种为嘎啦,树龄为8~10 a。林下套种中草药甘草(Glycyrrhiza uralensis Fisch.),该试验地以甘草为主要经济作物,而对苹果树未进行农事管理,苹小吉丁发生较为严重。该果园东西南北4个边缘均有以杨树和柳树为主的农田防护林带,呈单排种植,株距为3~5 m不等,期间偶有采伐留下的空缺。
-
粘虫板统一使用蛋黄色的粘虫板(580~590 nm),大小均为26 cm × 26 cm;植物源引诱剂统一使用YC;缓释瓶统一使用PVC缓释瓶(PVC材质,黑色,瓶身规格为25 mm × 11 mm,瓶口直径0.7 mm,容量1 mL);白色涂改液、捕虫网、记录本、碳素笔、G120BD手持GPS定位仪、放大镜、皮尺、自喷漆、数码相机(尼康公司)、PET材质透明塑料罐(罐底宽15.5 cm,高24.0 cm,口径11.2 cm)等。
-
以栽培果园中点的虫源释放罐为中心,分东、东南、东北、西、西南、西北、南、北8个方向,每个方向每隔5 m选择1株苹果树,统一在苹果树向阳处2~2.5 m高度枝条上悬挂一张苹小吉丁诱捕器(诱捕器为植物源引诱剂YC和蛋黄色粘虫板组合),东西方位设置5、10、15、20、25、30、35 m和40 m 8个距离梯度;其他方位均设置5、10、15、20、25、30 m和35 m 7个距离梯度。在样地东、南、西、北4个边缘防护林带各均匀选取5棵树,分别悬挂1个诱捕器,以对标记试虫和果园自然种群的潜在迁出行为进行初步探究。
-
通过使用涂改液对成虫的前胸背板进行标记,测试虫源释放位置位于果园中点,将装有虫源的塑料罐罐口朝上,任其自行爬出罐口进行飞行扩散。未成功飞行的试虫收回并记录为无效试虫。果园中点虫源释放罐中装有带标记的雌雄成虫共438头,其中雌成虫247头,雄成虫191头。于7月6日释放虫源,之后每隔3 d检查并记录各方位和距离诱捕器上诱捕到带标记和未标记的成虫,共调查4次,持续到7月22日。试验期间白天平均气温31.4 ℃,夜间平均气温18.3 ℃,以晴朗天气为主,未出现降水、大风等情况。及时补充引诱剂和更换粘虫板。
-
将不同方向诱捕器回收到的苹小吉丁标记虫数分开进行统计,同时下载试验期间的风向累计数据。
-
扩散速度Vi参考李超[19]的扩散速度计算方法,即用标记虫体与中心释放点的距离除以到达回捕点所用的时间。计算公式为:
$ {{Vi = }}\dfrac{{\sum {{{S_i}}} }}{{\sum {{{T_i}}} }} $
式中,Si为标记试虫扩散达到的位置;Ti为调查时间。
-
根据苹小吉丁标记试虫的扩散距离及回捕到的标记虫数计算移动距离。公式为:
$ {{D = }}\sqrt {\frac{{\sum {{{{d}}^{\rm{2}}}} }}{{{N}}}} $
式中,d为标记试虫扩散的距离,N为回捕到的苹小吉丁标记试虫数量[16]。
-
将不同调查时间内回捕到的标记虫数占释放标记虫数的比值作为回捕率。
-
分别记录不同距离处诱捕器回捕到的标记虫数,查看回捕虫数随诱捕器距离的变化趋势,同时将果园不同方向防护林中诱捕到的苹小吉丁虫数分开进行统计。
-
通过运用回捕到的标记成虫与未标记成虫数量估计田间种群数量N[20]。种群数量估计公式如下:
$ {{N = }}\frac{{{{Mn}}}}{{{m}}}$
式中,M为释放标记虫数,n为回收总虫数,m为回收标记虫数。
-
利用Microsoft Excel 2010软件对原始数据进行分析处理并绘制图表。试验期间相关的天气数据参考中国天气网(www.tianqi.com)。
-
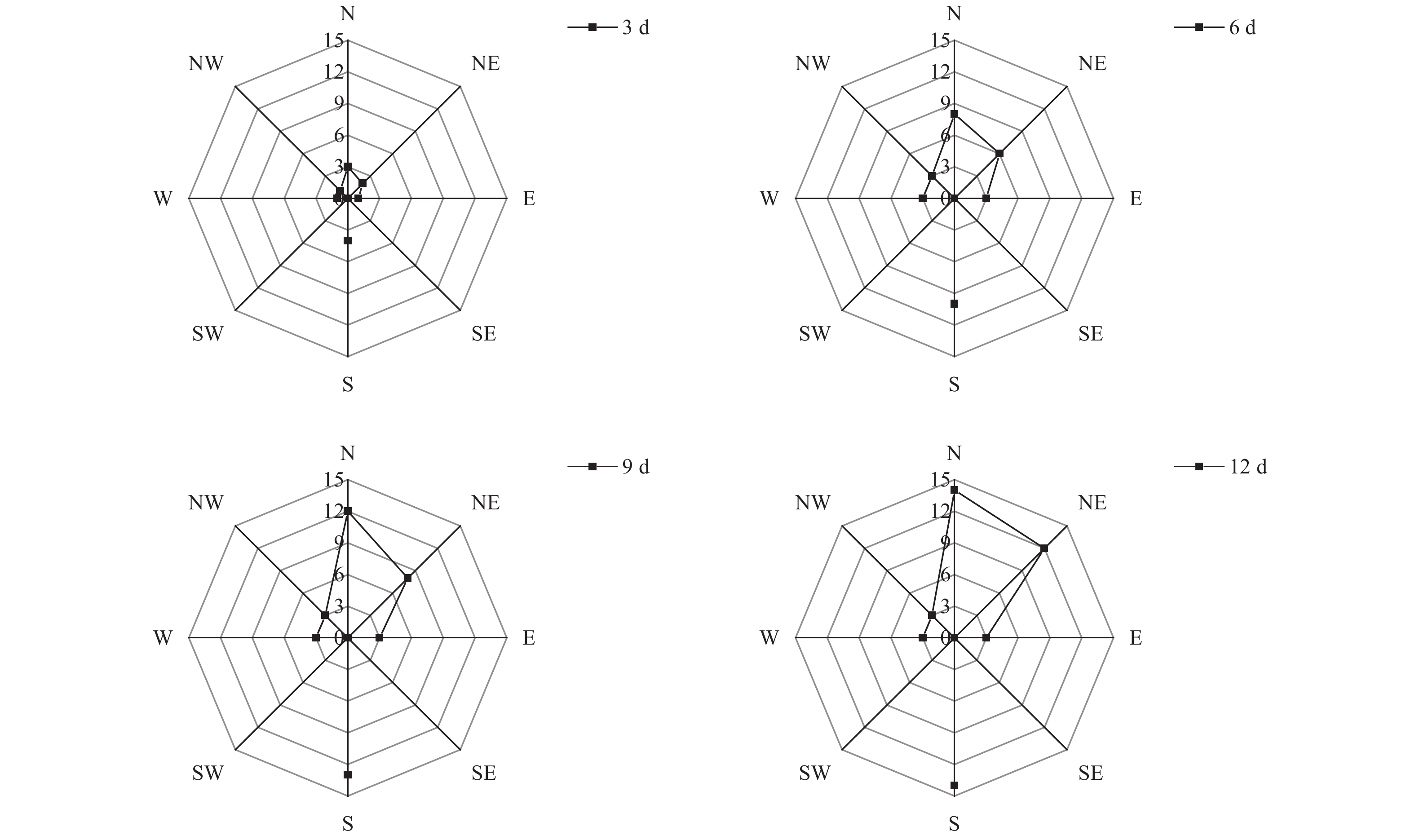
通过对不同方向12 d扩散总量进行统计,由图1可知,释放后3 d,于释放点东面、南面、西面、西北面、北面及东北面均已回收到标记试虫,其中南面扩散数量最多,共诱捕到4头;北面和东北面次之,依次诱捕到3头和2头。试虫释放后第6 d,南面、北面和东北面扩散数量增加明显,分别回收到10头、8头和6头;东面、西面和西北面均回收到3头,而东南面和西南面回收数量均为0。从12 d总扩散数量来看,苹小吉丁种群向北面、南面两个方位的扩散数量最多,均回收到14头;向东北面扩散的数量次之,累计回收到12头;东面、西面和西北面于第6 d累计回收到3头标记试虫后,回捕数量始终未增加。东南面和西南面始终未回收到标记试虫。从整体扩散趋势来看,成虫主要向南面、北面及东北面进行扩散。从试验期的累计风向来看(图2),南风向和北风向累计风力均为0,而南面和北面的扩散数量却最多,说明苹小吉丁喜欢向无明显风源的方向进行扩散,或者是因为本次试验使用的植物源引诱剂回捕法,苹小吉丁在无明显风力的情况下能够更为精确的对植物源引诱剂的位置进行定位,所以回捕数量更多。同时,由于果园行向为东西走向,东西长、南北短,试虫可能会选择距离较短的南北面向果园外进行扩散。
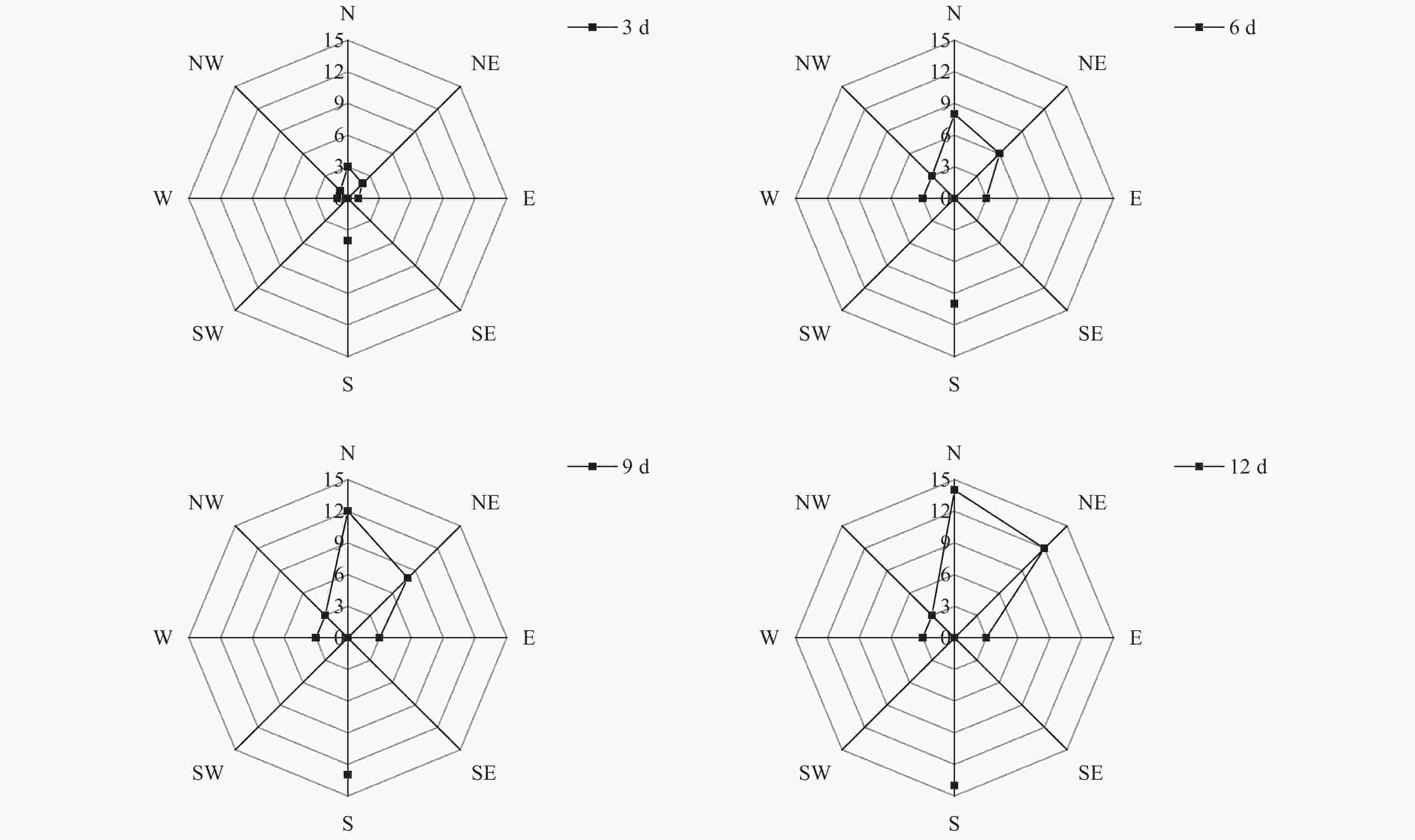
图 1 苹小吉丁在不同方向上的扩散数量
Figure 1. The number of diffusions in different directions of Agrilus mali Matsumura
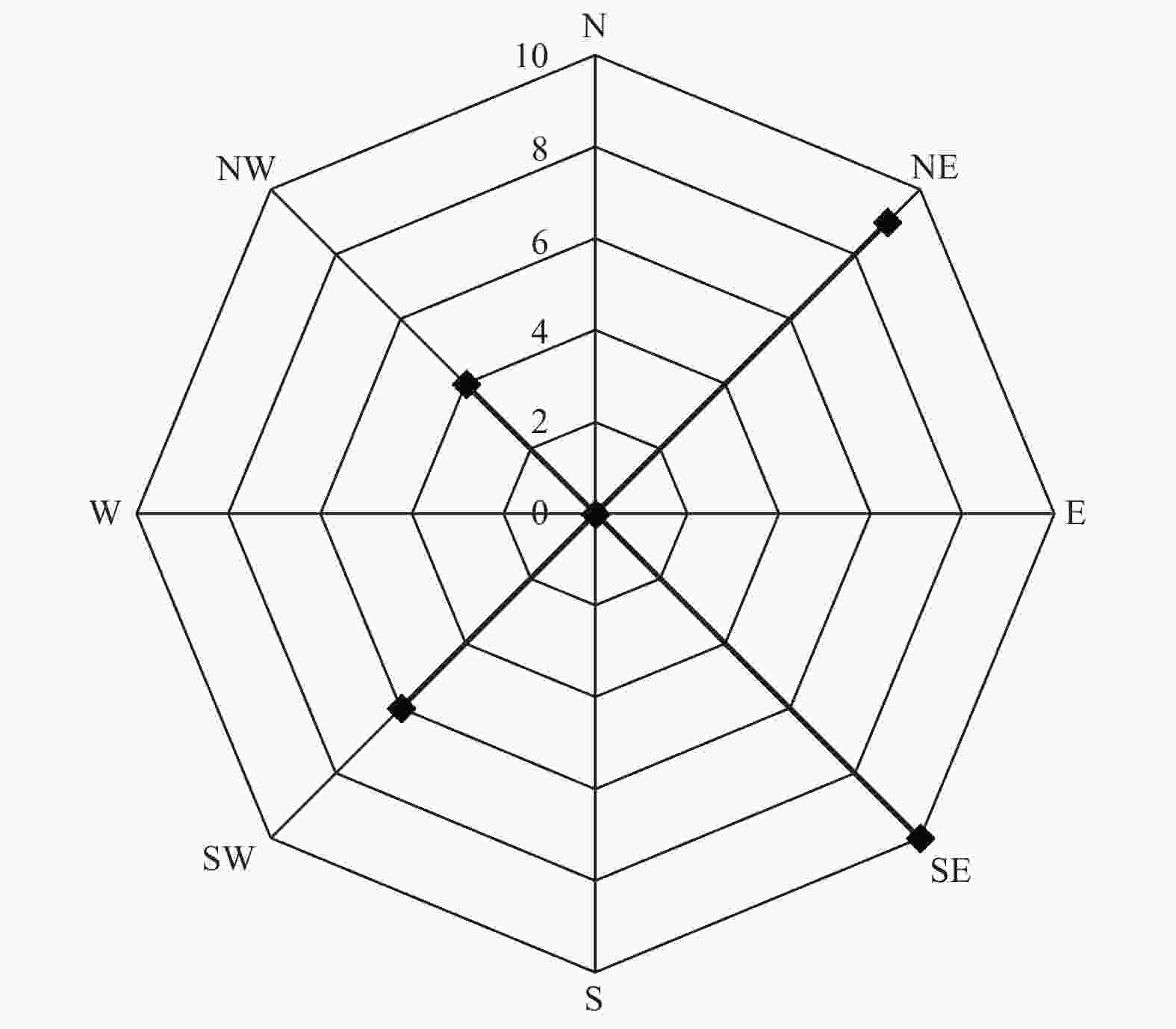
图 2 试验期风向累计
Figure 2. The wind direction accumulated during the test
-
标记试虫释放后,苹小吉丁不同性别成虫的扩散速度随着时间的推移具有不同的变化趋势(图3)。在标记试虫释放后第3 d,雌虫和雄虫的扩散速度一致,均为1.7 m·d−1。释放后第6 d,雌虫和雄虫的扩散速度均下降到最低,雌虫为1.3 m·d−1,雄虫为1.1 m·d−1。从第6 d开始,雌虫的扩散速度明显开始上升,并且与雄虫扩散速度差距不断增大。释放后第9 d雌虫扩散速度上升为2.2 m·d−1,在释放后第12 d其扩散速度达到最高,为2.5 m·d−1。雄性试虫在释放后第9 d其平均扩散速度又开始逐渐上升,为1.5 m·d−1,随后在第12 d又下降为1.4 m·d−1。雌雄试虫在12 d时扩散速度差异达到显著水平(P < 0.05)。从扩散速度可以看出,释放后前3 d是雄虫的主要扩散时期,而释放6 d以后是雌虫的主要扩散时期。
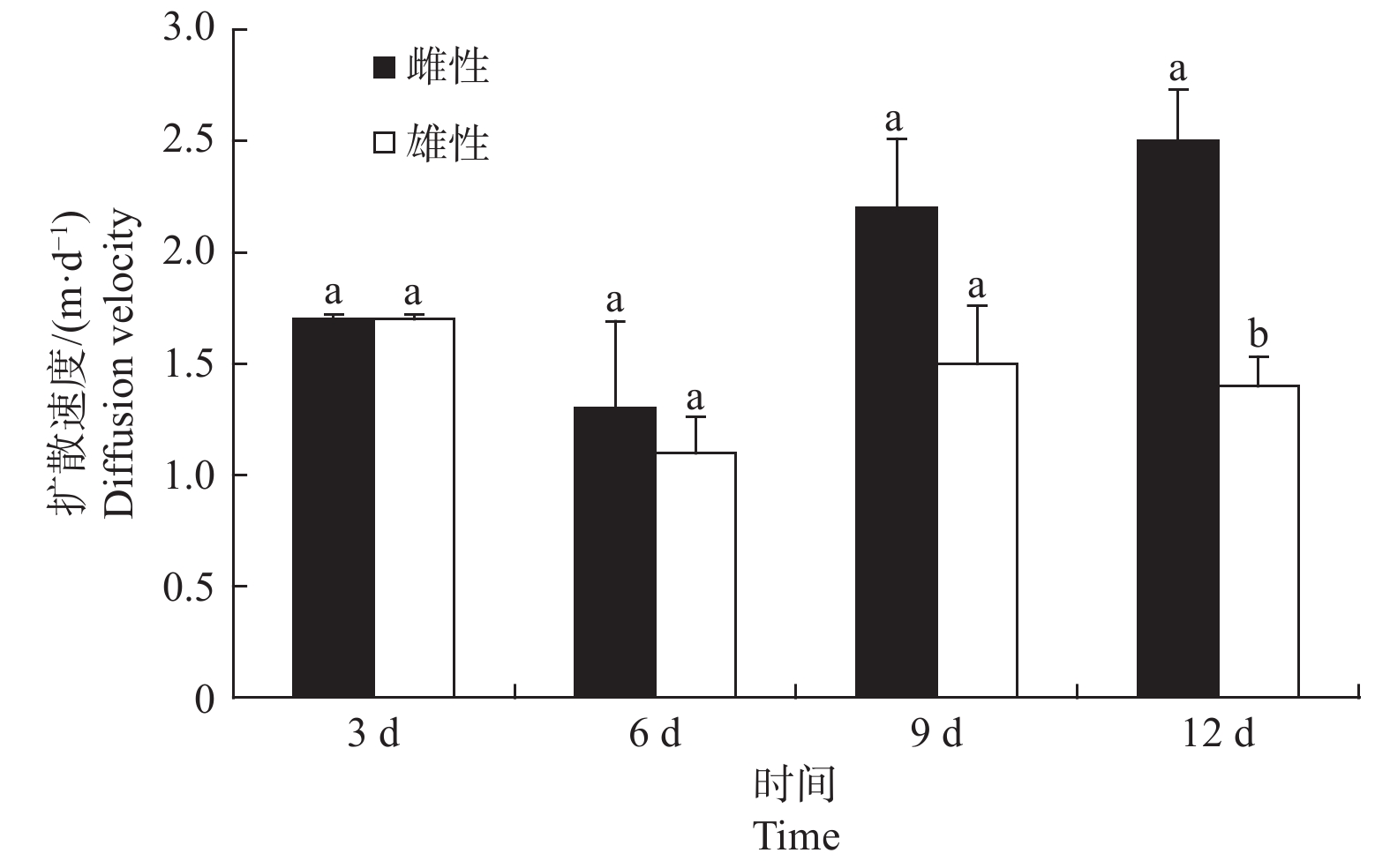
图 3 不同性别苹小吉丁成虫扩散速度
Figure 3. Different sex Agrilus mali Matsumura adult diffusion rate
-
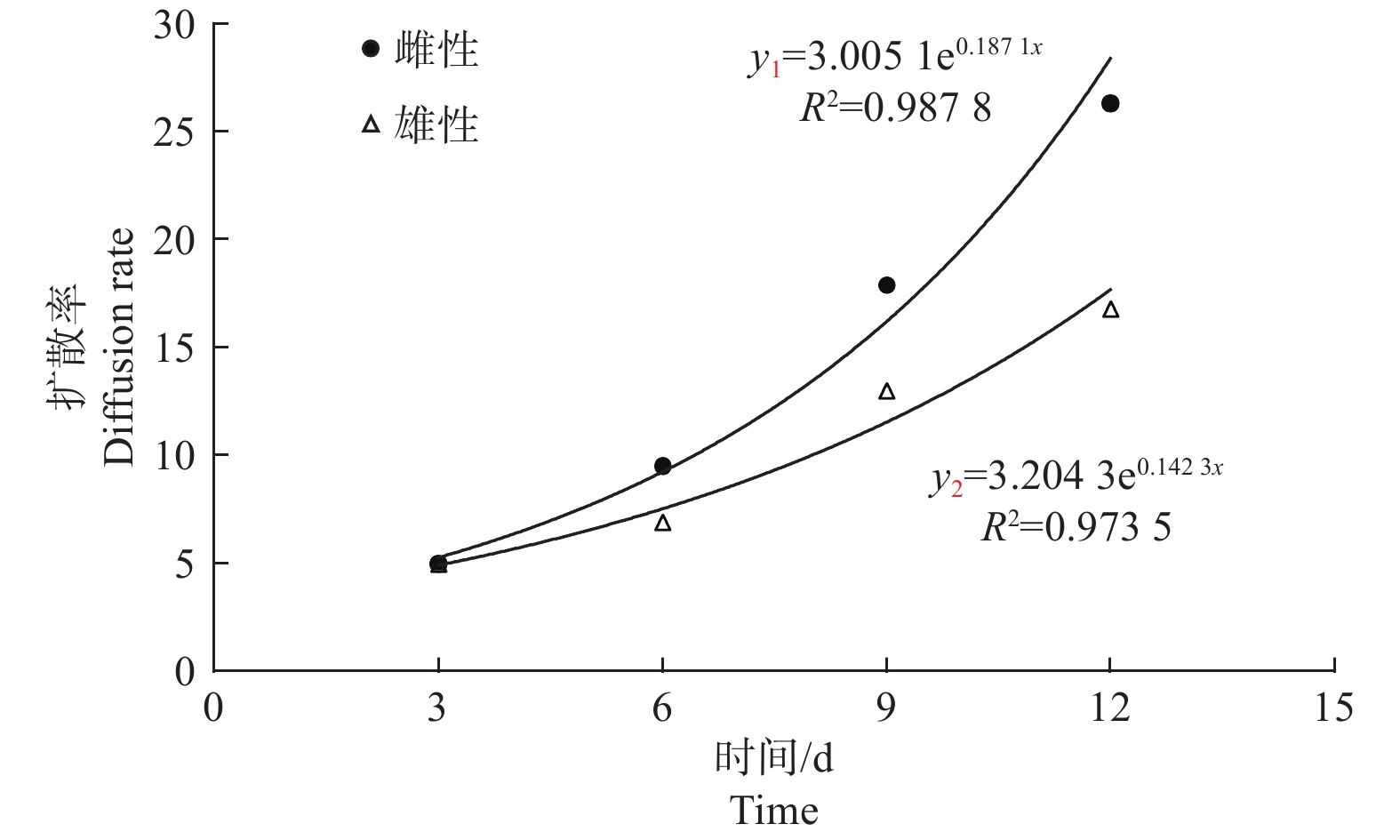
本试验为3 d调查一次回捕情况,从图4可以看出,苹小吉丁雌虫的扩散率随着释放时间的推移不断上升,第3、6、9、12 d的扩散率分别为5.0、9.5、17.9、26.3。从扩散率的上升趋势来看,前9 d雌虫的扩散率上升速率较快,第12 d的扩散率上升速度开始放缓。雄虫的扩散率变化趋势与雌虫基本相同,均随着释放时间的推移而逐渐上升,第3、6、9、12 d的扩散率分别为5.0、6.9、13.0、16.8。雄虫前9 d扩散率的上升趋势同样较快,第12 d的扩散率上升速度也开始放缓。

图 4 不同释放时间下苹小吉丁扩散率
Figure 4. The diffusivity of different release time inAgrilus mali Matsumura
-
标记试虫释放后,苹小吉丁雌雄虫在不同回捕时间的回捕率变化趋势基本一致。在释放后第6 d,苹小吉丁雌雄虫回捕率均为最高,分别达到4.5%和5.2%。雌虫在释放后第3 d的回捕率次之,为3.2%;并且雌虫在第9 d和第12 d的回捕率一致,均为1.6%;雄虫在释放后第9 d和第12 d的回捕率也逐渐降低,分别为2.6%和1.5%。从调查结果来看,无论是雄虫还是雌虫,其回捕率的变化趋势均为先迅速上升后逐渐下降(图5)。
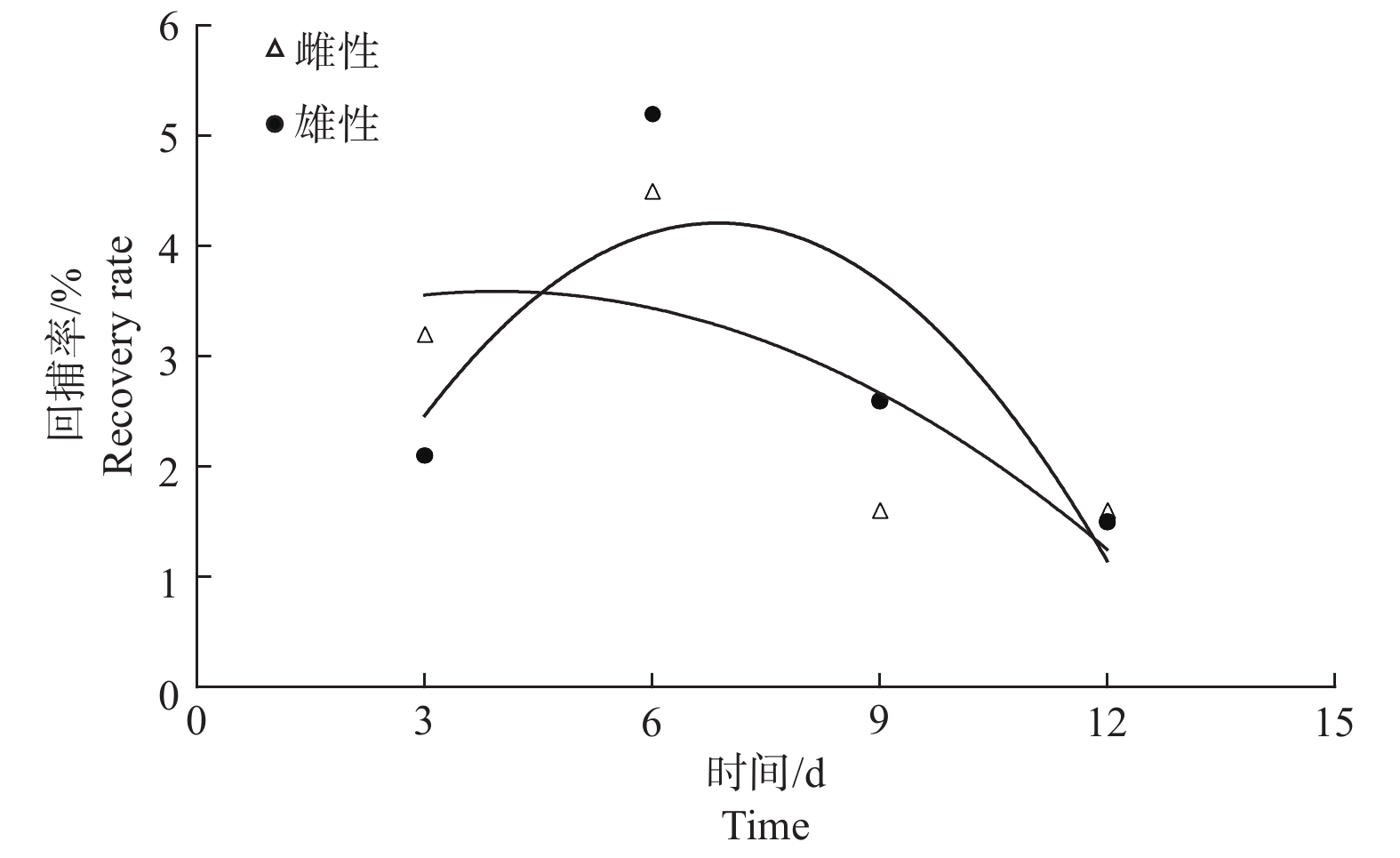
图 5 苹小吉丁不同时间回捕率差异
Figure 5. Different time recovery rate of Agrilus mali Matsumura
-
(1)不同距离回捕数量
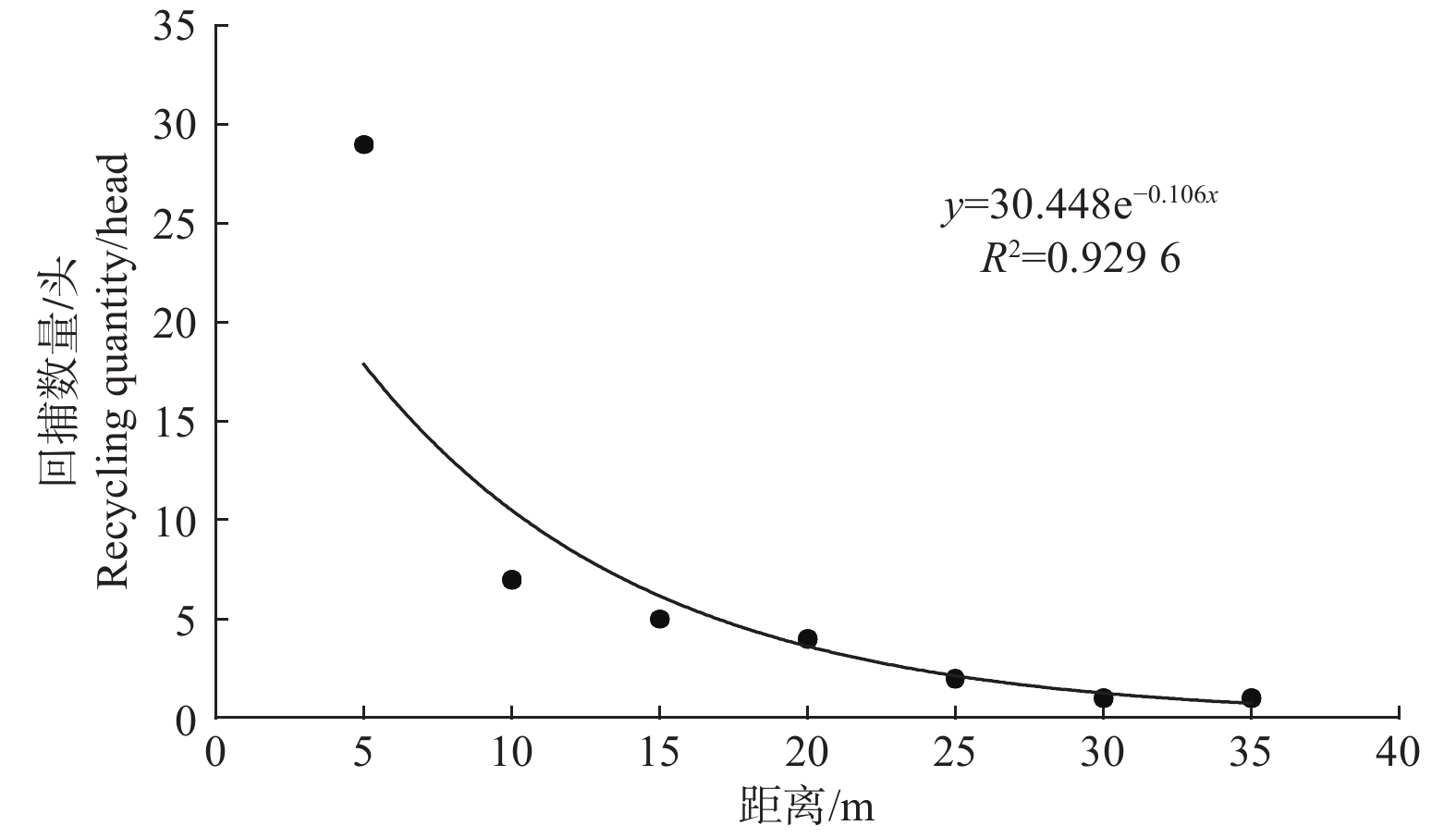
田间调查发现,苹小吉丁释放种群扩散至距释放点5 m处的数量最多,共诱捕到29头,随着距离的增加,扩散数量逐渐降低。在其他距离中,7头扩散至10 m,5头扩散至15 m,4头扩散20 m,2头扩散至25 m,在30 m和35 m处均诱捕到1头,说明苹小吉丁释放种群在种植寄主植物的栽培果园内具有一定的扩散能力,部分个体表现出向外扩散的趋势(图6)。
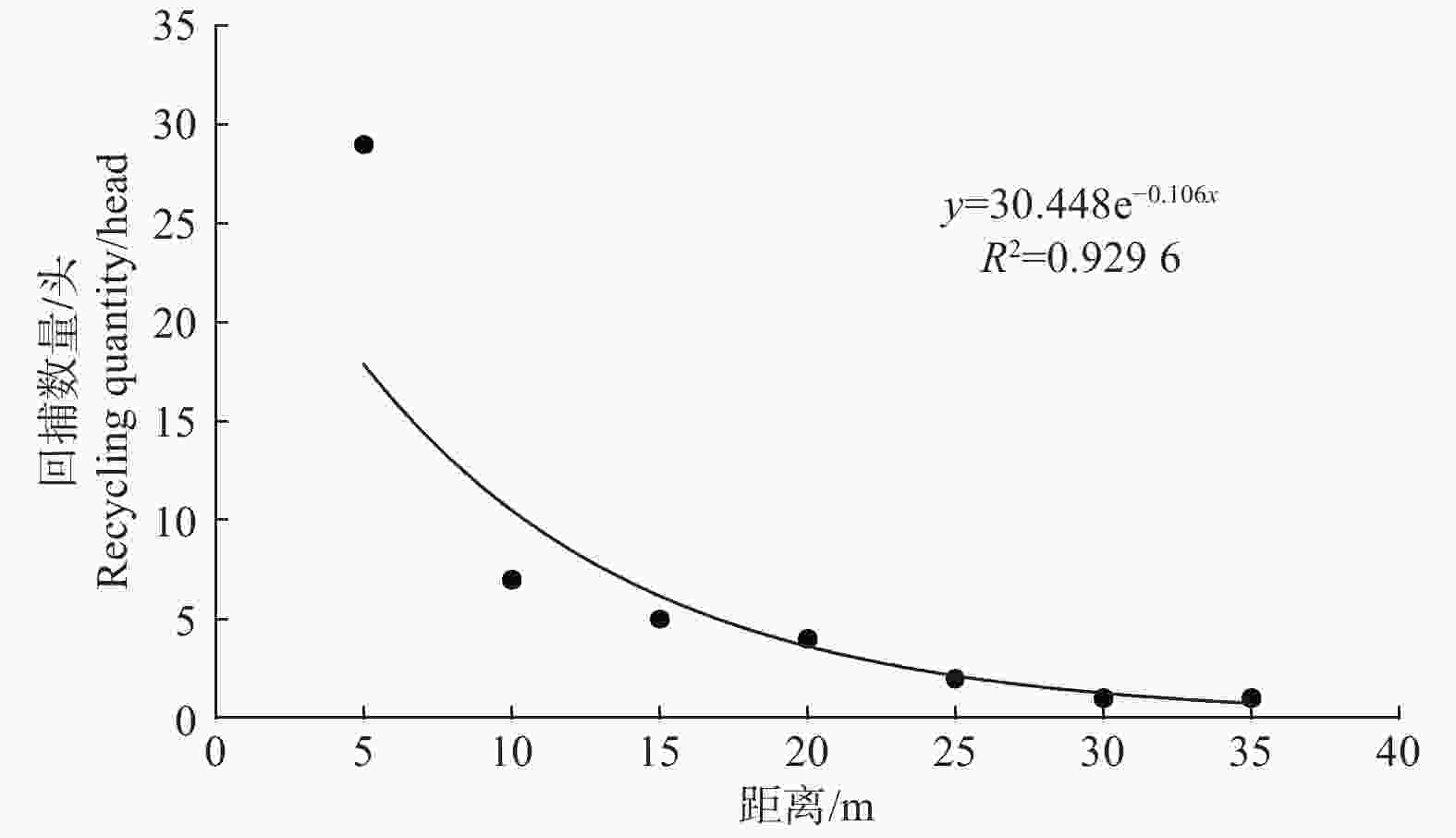
图 6 苹小吉丁不同距离回捕数量
Figure 6. The number of catches from different distances of Agrilus mali Matsumura
对苹小吉丁释放种群在栽培果园的扩散距离进行了方程拟合,结果表明,指数方程的R2值最高,达到0.9296;多项式方程的R2值次之,为0.8452;线性方程的R2值最低,仅为0.5893。说明从苹小吉丁释放种群在栽培果园的扩散距离来看,较为符合指数分布规律(详见表1)。
表 1 苹小吉丁扩散距离拟合方程
Table 1. The diffusion distance fitting equation of Agrilus mali Matsumura
模型
Model回归方程
The regression equationR2 多项式 Polynomial y = 0.053 8x2−2.8595x+37.286 0.8452 线性 Linear y = −0.7071x+21.143 0.5893 指数 Index y = 30.448e−0.106x 0.9296 (2)不同方向防护林回捕数量
通过在果园东西南北4个边缘的防护林悬挂苹小吉丁诱捕器,对具有迁出行为的苹小吉丁成虫进行诱捕监测(表2)。果园自然种群中,在诱捕器悬挂的第3 d,南缘、西缘和北缘防护林各监测到1头苹小吉丁成虫,其中南缘和西缘为雄虫,北缘为雌虫。东缘分别监测到1头雌虫和3头雄虫。南缘防护林在第6 d诱捕到数量最多,雌雄成虫分别为4头和2头;西缘和东缘也在第6 d诱捕数量达到最多,分别为3头雄虫和2头雄虫。北缘防护林在第9 d诱捕成虫数量最多,分别为2头雌虫和3头雄虫。从迁出数量来看,果园苹小吉丁自然种群向北缘防护林的迁出数量最多,南缘次之,东缘和西缘较少。说明该样地苹小吉丁具有向北面扩散及迁出的趋势。
表 2 果园边缘防护林未标记成虫诱捕数量
Table 2. The number of insect traps in the orchard edge shelterbelt of not marked
方向 Direction 回捕数量 Number of recovered 合计 Total 3 d 6 d 9 d 12 d ♀ ♂ ♀ ♂ ♀ ♂ ♀ ♂ 东 East 1 3 0 2 1 0 0 0 7 南 South 0 1 4 2 1 0 1 0 9 西 West 0 1 0 3 0 0 0 0 4 北 North 1 0 2 1 2 3 0 1 10 -
此次标记释放的438头苹小吉丁成虫中,在12 d试验期内共有49头标记试虫被回收,其中雌成虫共回收到27头,雄成虫共回收到22头。此外,田间未标记种群共诱捕到203头,雌成虫共诱捕到111头,雄成虫共诱捕到92头(表3)。
表 3 苹小吉丁虫回收数量及种群估计
Table 3. The recovery quantity and population estimation of Agrilus mali Matsumura
释放虫数
Release insect
count累计回收虫数
Cumulative number of recovered估计数量
Estimated
number标记
Tag未标记
Untag雌虫
Female247 27 111 1 262 雄虫
Male191 22 92 989 合计
Combined438 49 203 2 253 由种群数量预估方程计算得知(表3),栽培果园试验田间苹小吉丁估计种群数量为2 253头,雌成虫估计值为1 262头,雄成虫估计值为989头。预估结果虽然不能精确量化田间苹小吉丁成虫的种群数量,但能为初步了解田间发生情况提供参考依据。
-
在一定的作用范围内,寄主植物会引诱昆虫进行主动扩散,且寄主与虫源的距离对诱集数量会产生显著的影响,一般情况下,两者间的距离越近,寄主对虫源的诱集数量越多[21-22]。此次田间试验发现,苹小吉丁成虫扩散迁移对方位具有一定的选择性,雌虫和雄虫均主要向北面、东北面和南面进行扩散。在扩散距离上,苹小吉丁释放种群扩散至距释放点5 m处的数量最多,共诱捕到29头。随着距离的增加,扩散数量逐渐降低,这可能是由于苹小吉丁飞翔能力一般,喜欢选择距离较近的寄主植物进行补充营养以节省能量消耗,同时受到植物源诱剂与寄主植物的双重引诱作用,离试虫释放点的距离越近、引诱作用越强,所以5 m距离处诱捕数量最多。最远在北面35 m处诱捕到1头,说明标记释放的苹小吉丁成虫由果园北面迁出的潜在能力较大。这与李国宏对光肩星天牛(Anoplophora glabripennis)扩散距离的研究结果基本一致[16]。从苹小吉丁释放种群扩散距离的变化趋势来看,基本符合指数分布,这与沙蒿金叶甲(Chrysolina aeruginosa)在沙蒿林的扩散规律基本一致,沙蒿金叶甲的扩散规律与该虫本身的迁移特性有关,沙蒿金叶甲喜欢爬行移动而很少飞行,所以扩散能力一般,扩散速度较慢[23]。苹小吉丁可能由于个体体型较小、飞翔能力较弱,所以随着回捕点距离的增加扩散数量迅速降低。
本次研究发现,试虫释放前3 d苹小吉丁雄虫的扩散速度高于试验后期;而雌虫的扩散速度在后期即第12 d达到最高。由于雌性成虫需要通过不断取食来为产卵积累足够的营养,同时为了寻找合适的产卵位置,也需要不断进行补充营养来为其飞行提供糖类和脂类营养物质[24-25],所以雌虫的扩散速度随着释放时间的推移呈上升趋势。同种昆虫中,有时体型较大的个体表现出的飞行能力更强[26-27],与苹小吉丁雄虫相比,雌虫的扩散速度更快,可能是由于雌虫的体型更大,所以飞行能力更强。苹小吉丁雌雄成虫在释放后第6 d回捕率达到最高。不同方位防护林的回捕数量差异可能是与果园间苹小吉丁的分布模式有关,苹小吉丁在林间一般呈聚集性分布[10],果园北面和南面防护林的回捕数量更多可能是因为这两个方位边缘的虫口密度更大,这有待于进一步调查证实。由种群数量预估方程计算得知,栽培果园试验田间苹小吉丁估计种群数量为2 253头,雌成虫估计值为1 262头,雄成虫估计值为989头。说明田间自然种群中,苹小吉丁雌虫数量稍高于雄虫,这也可能是害虫在田间容易暴发的主要原因之一。
研究昆虫野外扩散行为一般通过虫源标记-释放-再回收等步骤进行试验,在试验过程中要着重考虑目标虫源、虫源释放时间和试验过程中的天气等条件[28]。本次试验试探性利用植物源诱剂对苹小吉丁林间扩散能力进行初步测定,试验周期较短,所以只考虑了风向对其扩散的影响。以后可以延长试验周期,同时评价温度、湿度及降水等因子对其扩散能力的影响作用,深入揭示苹小吉丁扩散行为的影响机制。埃及伊蚊具有向西北方向进行扩散的趋势[13],这与本研究结果基本一致。除了以诱捕器作为回收装置进行虫源收集外,还可以直接进行人为释放和再捕捉,通过人为释放-再捕捉法对果蝇的扩散行为进行研究时也取得了良好的预期结果[29]。
田间扩散行为研究一般在自然寄主田进行试验,但也可以在非寄主田或空白地块进行对比研究,探究目标害虫在无寄主植物条件下和有寄主植物条件下的扩散行为差异[14]。此次试验在已发生苹小吉丁危害的栽培苹果园进行,试虫可能容易受到寄主植物的影响,更喜欢停留在寄主植物上进行补充营养和栖息,降低了向田外扩散的可能性。可以于空白地或非寄主林分进行扩散行为研究,进一步探究在缺乏寄主植物的情况下苹小吉丁虫的扩散规律,或在空白荒地的条件下苹小吉丁的被迫迁飞行为。同时,本试验在栽培果园为第一次进行,在时间和条件允许的情况下可重复进行,更为精确的掌握其在田间的扩散规律。
-
从扩散方向来看,在样地中点虫源释放罐的北面、东北面和南面回捕到的苹小吉丁标记试虫数量较多,说明标记试虫主要向该3个方向进行扩散。从扩散距离来看,5 m处回捕到的标记试虫数量最多,达到29头,随着距离的增加回捕数量逐渐降低,最远可扩散至35 m,且雌虫的扩散速度强于雄虫。雌雄试虫的回捕率变化趋势基本一致,均为先上升后降低;而扩散率均随着时间的推移逐渐上升。于样地北面防护林回捕到的自然种群数量最多,说明该样地苹小吉丁向样地北面进行扩散及迁出的可能性最大。也可利用回收标记虫数对样地自然种群数量进行预测,为评价样地苹小吉丁发生情况进行初步评估。本研究结论也可为苹小吉丁林间扩散路径及监测预警区域划分提供一定科学指导依据。
苹小吉丁林间扩散行为的初步研究
A Preliminary Study on the Field-Diffusion Behavior of Agrilus mali Matsumura (Coleoptera: Bupresidea)
-
摘要:
目的 掌握苹小吉丁成虫在林间的自然扩散行为规律,以制定合理的害虫监控策略。 方法 于伊犁巩留县五乡栽培苹果园内采用标记-重捕法监测苹小吉丁雌雄试虫在不同方向、不同距离的扩散数量及影响因子进行了研究,对扩散速度进行了分析,同时对林间种群数量进行了预估。 结果 结果表明:苹小吉丁在林间的扩散对方向具有一定的选择性,主要向北面、南面及东北面进行扩散,分别回捕到标记释放总虫数的3.2%、3.2%和2.7%;东南面和西南面均未回捕到标记试虫。随着回捕点距离的增加,标记试虫回捕数量逐渐降低,扩散至5 m处的数量最多,占标记释放总虫数的6.6%,最远扩散至北面35 m,说明标记试虫由果园北面迁出的潜在性较大,扩散距离整体呈指数分布趋势(R2 = 0.9262)。苹小吉丁雄虫扩散速度高峰出现在第3 d,为1.7 m·d−1;而雌虫扩散速度高峰出现在第12 d,达到2.5 m·d−1。随着释放时间的推移苹小吉丁雌雄虫的扩散率逐渐上升。由种群数量预估方程计算得知,试验田间苹小吉丁估计种群数量为2253头,雌成虫估计值为1262头,雄成虫估计值为989头。 结论 苹小吉丁在林间具有一定的自然扩散能力,最远可达35 m,且雌虫的扩散能力要强于雄虫。林间种群数量预估结果能为苹小吉丁发生动态预测和控制策略制定提供参考依据。 Abstract:Objective To understand the natural diffusion behavior of Agrilus mali Matsumura, in order to develop reasonable pest control strategy. Method In an apple orchard at Yili of Xinjiang Uygur Autonomous Region, the method of marking and re-catching was used to monitor the amount of male and female A. mali spreading in different directions and distances. The influencing factors were studied, the diffusion velocity was analyzed, and the insect population was estimated. Result The results showed that the diffusion of A. mali among forests was selective in direction. It diffused mainly to the north, south and northeast, the insect re-caught in the three directions occupied respectively by 3.2%, 3.2% and 2.7% of the total amount of the marked insect released. No marked specimens were re-caught in the directions of southeast and southwest. With the increase of the distance to re-catching point, the amount of re-caught insects decreased, the maximum amount of diffusion was at 5 m, which accounted for 6.6% of the total amount of released A. mali. It was found the farthest distance of diffusion was 35 m to the north, indicating that there was a greater potential for the marked insect to migrate from the north of the orchard. The diffusion distance shows an exponential distribution trend. The peak diffusion rate of the male appeared in the third day, which was 1.7 m/d; while the peak of the average diffusion rate of females occurred on 12th day, at 2.5 m/d. With the prolonging of the release time, the diffusivity of male and female insects increased gradually. According to the population amount estimation equation, the estimated population amount of A. mali was 2,253, the female adult was estimated at 1,262, the estimated male adult was 989. Conclusion A. mali in the forest has a certain natural diffusion ability up to 35 meters. the female is more diffusive than the male, the population estimation of A. mali in forestland can provide a reference for the dynamic prediction and control strategy. -
Key words:
- Agrilus mali
- / marking-recovery
- / dispersal behavior
- / population assessment
-
图 1 苹小吉丁在不同方向上的扩散数量
Figure 1. The number of diffusions in different directions of Agrilus mali Matsumura
图 3 不同性别苹小吉丁成虫扩散速度
Figure 3. Different sex Agrilus mali Matsumura adult diffusion rate
图 4 不同释放时间下苹小吉丁扩散率
Figure 4. The diffusivity of different release time inAgrilus mali Matsumura
图 6 苹小吉丁不同距离回捕数量
Figure 6. The number of catches from different distances of Agrilus mali Matsumura
表 1 苹小吉丁扩散距离拟合方程
Table 1. The diffusion distance fitting equation of Agrilus mali Matsumura
模型
Model回归方程
The regression equationR2 多项式 Polynomial y = 0.053 8x2−2.8595x+37.286 0.8452 线性 Linear y = −0.7071x+21.143 0.5893 指数 Index y = 30.448e−0.106x 0.9296  下载: 导出CSV
下载: 导出CSV
表 2 果园边缘防护林未标记成虫诱捕数量
Table 2. The number of insect traps in the orchard edge shelterbelt of not marked
方向 Direction 回捕数量 Number of recovered 合计 Total 3 d 6 d 9 d 12 d ♀ ♂ ♀ ♂ ♀ ♂ ♀ ♂ 东 East 1 3 0 2 1 0 0 0 7 南 South 0 1 4 2 1 0 1 0 9 西 West 0 1 0 3 0 0 0 0 4 北 North 1 0 2 1 2 3 0 1 10
下载: 导出CSV
表 3 苹小吉丁虫回收数量及种群估计
Table 3. The recovery quantity and population estimation of Agrilus mali Matsumura
释放虫数
Release insect
count累计回收虫数
Cumulative number of recovered估计数量
Estimated
number标记
Tag未标记
Untag雌虫
Female247 27 111 1 262 雄虫
Male191 22 92 989 合计
Combined438 49 203 2 253
下载: 导出CSV
-
[1] 李利平, 海 鹰, 安尼瓦尔·买买提, 等. 新疆伊犁地区野果林的群落特征及保护[J]. 干旱区研究, 2011, 28(1):60-66. [2] 成克武, 周晓芳, 臧润国, 等. 新疆野苹果资源保护对策探讨[J]. 干旱区研究, 2008(6):760-765. [3] 张艳敏, 冯 涛, 张春雨, 等. 新疆野苹果研究进展[J]. 园艺学报, 2009, 36(3):447-452. doi: 10.3321/j.issn:0513-353X.2009.03.022 [4] 宋益学. 新疆野苹果的管理现状和保护措施[J]. 新疆林业, 2006,(6):34-35. doi: 10.3969/j.issn.1005-3522.2006.06.024 [5] 阎雄飞, 张彦龙, 刘永华, 等. 苹果小吉丁虫在陕西省潜在危害性风险分析和管理对策[J]. 植物检疫, 2017, 31(3):46-52. [6] 李孟楼, 张正青. 苹果小吉丁虫的生物学及其生活史讨论[J]. 西北林学院学报, 2017, 32(4):139-146. [7] 刘忠权, 陈卫民, 许 正, 等. 新疆天山西部野苹果林分布与苹果小吉丁虫危害现状研究[J]. 北方园艺, 2014(17):121-124. [8] 季 英, 季 荣, 黄人鑫. 外来入侵种——苹果小吉丁虫及其在新疆的危害[J]. 新疆农业科学, 2004,(1):31-33. doi: 10.3969/j.issn.1001-4330.2004.01.009 [9] 崔晓宁, 刘德广, 刘爱华. 苹果小吉丁虫综合防控研究进展[J]. 植物保护, 2015, 41(2):16-23. doi: 10.3969/j.issn.0529-1542.2015.02.003 [10] 刘爱华. 新疆野果林苹果小吉丁与优势天敌生物学、生态学研究[D]. 乌鲁木齐: 新疆农业大学, 2010. [11] 何海明, 银建民, 伊力哈木, 等. 伊犁地区果树发生苹果小吉丁虫[J]. 新疆农业科技, 1999,(6):18. [12] 温俊宝, 李友常, 夏乃斌, 等. 杨树光肩星天牛成虫扩散格局的研究[J]. 生态学报, 1998, 18(3):47-55. [13] Russell R C, Webb C E, Williams C R, et al. Mark–release–recapture study to measure dispersal of the mosquito Aedes aegypti in Cairns, Queensland, Australia[J]. Medical and Veterinary Entomology, 2005, 19: 451-457. doi: 10.1111/j.1365-2915.2005.00589.x [14] Botero-Garces N, Isaacs R. Movement of the grape berry moth, Endopiza viteana: displacement distance and direction[J]. Physiological Entomology, 2004, 29(5): 443-452. doi: 10.1111/j.0307-6962.2004.00415.x [15] 李 晶. 马铃薯甲虫种群空间格局与扩散研究[D]. 乌鲁木齐: 新疆农业大学, 2015. [16] 李国宏, 高瑞桐, Michael T. Smith, 等. 光肩星天牛种群扩散规律的研究[J]. 林业科学研究, 2010, 23(5):678-684. [17] 卡德艳·卡德尔, 彭 彬, 马志龙, 等. 苹小吉丁对不同单色光及波长的趋性研究[J/OL]. 林业科学研究, 2020,33(1):113-122.https://doi.org/10.13275/j.cnki.lykxyj.2020.01.015. [18] 卡德艳·卡德尔, 彭 彬, 马志龙, 等. 寄主植物挥发物与粘虫板对林间苹小吉丁引诱效果评价[J]. 新疆农业科学, 2019, 56(9):1720-1732. [19] 李 超, 彭 赫, 程登发, 等. 马铃薯甲虫成虫田间扩散规律[J]. 生态学报, 2014, 34(2):359-366. [20] 唐 杰. “模拟动物种群密度调查的标记重捕法”实践的一些改进[J]. 中学生物学, 2014, 30(3):42-43. [21] 艾怡雯. 螺旋粉虱起飞和扩散的影响因子研究[D]. 海口: 海南大学, 2011. [22] 周福才, 王 勇, 李传明, 等. 寄主种类、距离和种群密度对烟粉虱扩散的影响[J]. 生态学报, 2007,(11):4913-4918. doi: 10.3321/j.issn:1000-0933.2007.11.063 [23] 王 杰, 李岳诚, 张大治. 基于标记重捕法的沙蒿金叶甲种群扩散研究[J]. 环境昆虫学报, 2016, 38(5):912-917. [24] 江幸福, 罗礼智, 李克斌, 等. 温度对甜菜夜蛾飞行能力的影响[J]. 昆虫学报, 2002, 45(2):275-278. doi: 10.3321/j.issn:0454-6296.2002.02.021 [25] 郑作涛. 二点委夜蛾飞行与生殖的互作[D]. 北京: 中国农业科学院, 2014. [26] 王 辉, 许志春, Baode Wang, 等. 花曲柳窄吉丁潜在飞行能力初步研究[J]. 北京林业大学学报, 2015, 37(1):92-97. [27] Lu Y, Wu K, Guo Y. Flight Potential of Lygus lucorum (Meyer-Dur) (Heteroptera: Miridae)[J]. Environmental Entomology, 2007, 36(5): 1007-1013. [28] Reisen W K, Lothrop H D, Lothrop B. Factors Influencing the Outcome of Mark-Release-Recapture Studies with Culex tarsalis (Diptera: Culicidae)[J]. Journal of Medical Entomology, 2003, 40(6): 820-829. doi: 10.1603/0022-2585-40.6.820 [29] Cronin J T, Hyland K, Abrahamson W G. The pattern, rate, and range of within-patch movement of a stem-galling fly[J]. Ecological Entomology, 2010, 26(1): 16-24. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4120
- HTML全文浏览量: 2208
- PDF下载量: 41
- 被引次数: 0



