


-
人工林是我国森林资源的重要组成部分,是重要的商品林种类[1]。第八次森林资源连续清查的结果显示,国内森林面积为20768.73万hm2,其中,人工林面积为6933.38万hm2,占33.38%[2]。人工林主要表现为大规模、大面积且单一树种的集约化经营,林下植被通常被人为清除[3-4]。因而,人工纯林生态系统通常呈现乔木物种单一、林下植被物种稀少、林分结构相对简单等特征,由此带来的生物多样性减少、水土流失、生态系统稳定性降低等一系列生态问题,对人工林的可持续经营以及生态环境保护极其不利[3, 5-6]。
林下植被是人工林生态系统的重要结构组分,对提高生物多样性、生态系统稳定性、森林生产力及促进养分循环等具有重要影响[7-8],如王瑞华等[9]研究表明,林下植被能有效增加森林生物多样性、生物量及养分供应水平。林下植被对土壤生物学过程也有较好的促进作用,如林下植被有利于促进硝化作用,进而影响土壤氮元素的供给[10];Rivalland等[11]发现,林下植被对森林水分平衡与碳储量产生显著的影响。然而,人工林的组成树种与其林下植被之间存在复杂的相互作用关系,如徐馨等[12]发现,不同林龄木麻黄(Casuarina equisetifolia Forst.)人工林林下植物物种数随林龄增加而显著增加;庞圣江等[13]研究西南桦(Betula alnoides Buch.-Ham. ex D. Don)与红锥(Castanopsis hystrix Miq.)、灰木莲(Magnolia sumatrana var. glauca (Blume) Figlar & Noot.)、杉木(Cunninghamia lanceolata (Lamb.) Hook.)及马尾松(Pinus massoniana Lamb.)等树种的混种人工林生态系统的稳定性,揭示不同林分结构对林下植被的动态特征及其稳定性的影响。这些研究都表明,人工林的林分结构与林下植被相互之间的作用对于人工林的经营十分重要。
植物叶片功能性状是植物适应环境所表现出的叶片水平上的特征参数,对阐明植物对环境变化的适应具有特殊意义[14]。比叶质量是指单位叶片面积的干质量,反应植物对光的截获与干物质量投入的比例关系,能够较好地反应植被更新、群落及生态系统的演替规律[15]。在叶片的光合作用中,光合色素是参与光的吸收、传递以及引起原初光化学反应的重要载体[16];而非结构性碳水化合物包含可溶性糖及淀粉,是植物生长代谢过程中的重要能量储存方式[17]。研究表明,这些植物叶片功能性状间存在着密切的联系,共同反应植物对环境变化的适应性及可塑性,同时这些性状又极易受到土壤养分供应状况的影响,因此,在人工林林下植被生长及更新中不可忽视[14-15]。
近年来,人工林由以木材产品为主的经营方式向以发挥生态效应为主的公益林或碳汇林的经营方式逐渐转变,更加突出人工林的综合生态功能服务效应,因此,构建结构完整及功能完善的人工林生态系统对生态环境建设具有重要的实践意义[18-19]。林下植被是人工林的重要组成部分,其生长的优劣对人工林生态系统功能的发挥具有重要的意义[20]。随着人工纯林逐渐转变为成熟林或过熟林,其林下植被的更新及群落构建已成为人工林经营中的重要议题。因此,研究人工林不同林下植被的生长状况及其受林冠层环境的影响有利于人工林林下植被的选择与优化,以期为未来人工林生态系统的重建提供理论参考。
马占相思人工林是华南地区的重要人工林类型之一[21],与黧蒴(Castanopsis fissa (Champ. ex Benth.) Rehd. et Wils.)、荷木(Schima superba Gardn. et Champ.)、马尾松、杉木等人工林相比,马占相思林分在林分生长及固碳功能上有较大的优势[22]。本文探讨广东鹤山恢复34年的马占相思人工林4种林下灌木叶片功能性状(包含比叶质量、氮磷元素含量、光合色素含量及非结构性碳水化合物等)及其根围土壤的养分特征,探讨叶片不同功能性状之间的内在联系,试图从叶片功能性状的角度,阐明华南地区人工林林下植被的结构功能以及对人工林管理的影响,同时为华南地区主要人工林林下植被更新的树种选择提供科学数据。
-
本研究样地设置在广东鹤山森林生态系统国家野外科学观测研究站内,简称鹤山站。该区域属南亚热带粤中丘陵地区,112°54′ E,22°41′ N,平均海拔80 m。该地区年均气温23℃,年降水量约1 400~1 700 mm,5—9月为雨季(降雨量约占全年75.8%),年蒸发量约1 600 mm,年平均 ≥ 10℃的有效积温为7 597.2℃。鹤山站的土壤类型为赤红壤(砖红壤),其所代表的区域范围包括粤中、闽南和桂东南。该区域的顶极植被群落是亚热带季风常绿阔叶林,由于过往人类活动过度频繁导致该区域植被退化、水土流失、生物多样性锐减等严重的生态环境问题。自1984年起,鹤山站建立起多种人工林类型,主要包括桉树林、马尾松林、荷木林、马占相思林等[23]。
-
马占相思人工林于1984年在广东鹤山退化荒坡上建立,栽种规格为2.5 m × 2.5 m,样地总面积为1 hm2,至2018年时已有34年的恢复年限,已接近成熟人工林的群落状态。本试验选择马占相思人工林林下个体数较多的4种灌木开展试验,包括九节、梅叶冬青、三叉苦和桃金娘,4种灌木的具体信息见表1。
表 1 马占相思人工林林下4种灌木生长特性
Table 1. Growth characteristics of four understory shrubs in Acacia mangium plantation
种名
Species Name科
Family地径
Ground diameter/cm株高
Height/cm冠幅
Canopy/(cm × cm)密度
Density/(株·hm−2)盖度
Coverage/%九节Psychotria rubra 茜草科 Rubiaceae 4.04 ± 0.74 207.6 ± 28.9 197.8 × 174.6 309 10.5 梅叶冬青 Ilex asprella 冬青科 Aquifoliaceae 4.28 ± 0.28 292.8 ± 39.2 327.1 × 353.8 252 29.1 三叉苦 Evodia lepta 芸香科 Rutaceae 3.60 ± 0.67 255.3 ± 34.4 173.7 × 188.4 271 8.8 桃金娘 Rhodomyrtus tomentosa 桃金娘科 Myrtaceae 2.38 ± 0.47 203.0 ± 19.8 129.1 × 150.4 509 9.9 在2018年7月,每种灌木选择3~5个生长状况相对一致的个体,测定每种灌木的株高、地径、冠幅等,并在每个个体采集30片健康、完全展开的成熟叶片,并调查其种群密度与盖度,同时在根部附近使用土钻(内径为6 cm)采集土样,在每个灌木个体的根围附近(距离主根30~50 cm)采集3~5钻混合样,采样深度为0~10 cm土层。所有样品放入封口薄膜袋标记保存并带回实验室备用。
-
新鲜叶片带回实验室清洗干净并晾干,去除多余水分,部分新鲜叶片使用叶面积仪测定叶面积,经烘干后称质量再计算比叶质量(g·m−2);另取部分新鲜叶片,用打孔器(直径7 mm)在叶片上打圆片,经80%(体积比)丙酮中浸泡72 h,至完全褪色,提取液用紫外-可见分光光度计测定3种波长,分别为OD663、OD645和OD470,并通过以下公式计算叶绿素和类胡萝卜素含量:
叶绿素a Chl a/(mg·L−1)=12.7×OD663−2.69×OD645
叶绿素b Chl b/(mg·L−1)=22.9×OD645−4.68×OD663
类胡萝卜素Car/(mg·L−1)=(1000×OD470−3.27×Chl a−104 × Chl b)/229
叶片样品经烘干粉碎后测定N、P含量,计算N∶P。叶片全N含量采用半开氏法测定,全P含量采用酸溶-钼锑抗比色法测定。
叶片非结构性碳水化合物(NSC)使用蒽酮比色法测定,借鉴Buysse等[24]的测定方法。植物样品中的可溶性糖用无水乙醇法提取,提取液可测定葡萄糖含量,同时用氢氧化钾溶液处理后测定蔗糖含量,残渣用高氯酸提取后测定淀粉含量。分别以不含提取液的混合液作空白对照,绘制相应的0、10、20、30、40、50 μg·mL−1的标准曲线,用蒽酮比色法测定620 nm波长下的吸光值,再根据葡萄糖的标准曲线分别计算出可溶性糖和淀粉的含量,其中,NSC为可溶性总糖与淀粉的总和。
土壤样品带回实验室后均在4℃条件下冷藏,采样后48 h内浸提土壤无机氮(铵态氮和硝态氮)。样品挑去根系残体后过2 mm筛,称取10 g鲜土,加入50 mL 2 mol·L−1 KCl溶液浸提,震荡1 h后过滤储存备用。铵态氮采用靛酚蓝比色法测定,硝态氮采用紫外分光双波长测定。土壤含水量用烘干法测定,即称取10 g左右鲜土,105℃烘干48 h至恒定质量。所有结果均折算成单位干土质量。
土壤样品经自然风干后磨碎过0.25 mm筛后测定总有机质、总氮和总磷等。土壤pH用水土比(2.5∶1)测定;土壤总碳用浓硫酸-重铬酸钾外加热法测定,土壤总有机质(SOM)为土壤总碳乘以转换系数1.72[23];总氮(TN)用半微量凯氏定氮法测定;土壤总磷(TP)用微量凯氏消煮法消煮后,再用钼锑抗比色法测定;土壤有效磷(AP)以Bray-2溶液浸提后用钼锑抗比色法测定。
-
采用单因素方差分析(One-way-ANOVA)和最小显著差异法(LSD)评估不同林下灌木叶片比叶质量、氮磷元素含量、光合色素、非结构性碳水化合物及土壤化学特性的差异;同时采用Pearson相关分析4种林下灌木叶片各种指标间的相关性。本文的统计分析均在SPSS 22.0(SPSS, Chicago, USA)中进行,统计图形均在Excel 2013(Microsoft Crop., Redmond, WA, USA)中绘制。
-
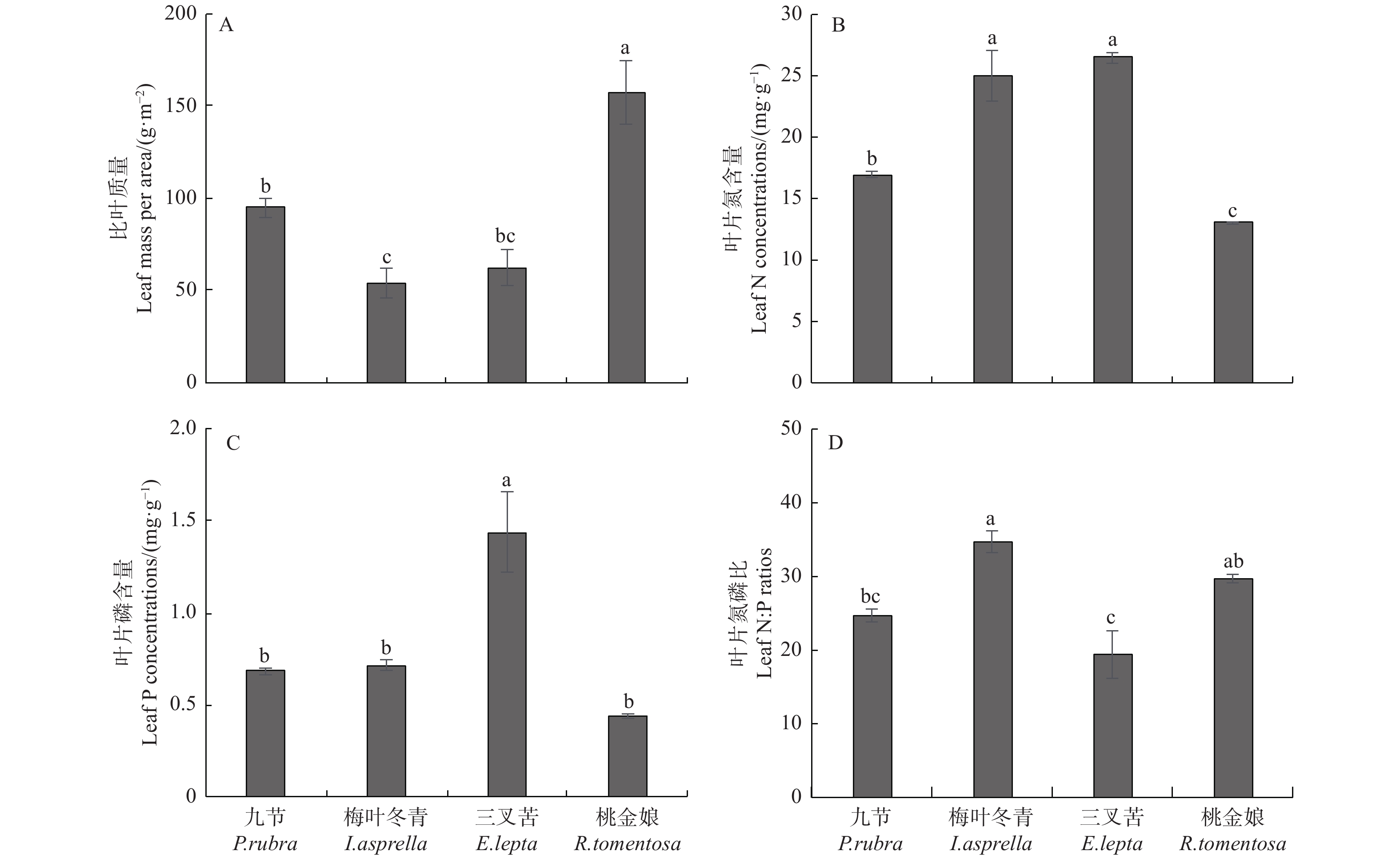
本研究中,桃金娘的比叶质量为157.51 g·m−2,显著高于其它3种灌木,且约为梅叶冬青比叶质量的3倍,九节的比叶质量也显著高于梅叶冬青的(图1A)。梅叶冬青和三叉苦叶片的氮含量分别为24.93、26.43 mg·g−1,二者均显著高于九节和桃金娘叶片的,且九节的叶片氮含量也显著高于桃金娘叶片的(图1B)。三叉苦叶片的磷含量最高,为1.44 mg·g−1,显著高于其它3种灌木叶片的磷含量,但后三者叶片的磷含量之间无显著差异(图1C)。梅叶冬青和桃金娘叶片氮磷比分别为35和30,二者之间无显著差异,但二者叶片的氮磷比均显著高于三叉苦叶片的,同时梅叶冬青叶片的氮磷比也显著高于九节叶片的(图1D),三叉苦叶片的氮磷比最低,为20。

图 1 马占相思人工林林下4种灌木叶片比叶质量、氮磷含量及氮磷比
Figure 1. Leaf mass per area (LMA), N and P concentrations, and N∶P ratios of four understory species in Acacia mangium plantation
-
马占相思人工林林下4种灌木叶片光合色素含量呈显著的物种差异,梅叶冬青叶片的叶绿素a(Chl a)、叶绿素b(Chl b)和叶绿素(a+b)分别为1.89、0.85、2.74 mg·g−1,均显著高于九节和桃金娘的叶绿素含量,4种灌木叶片叶绿素含量(a、b及a+b)大小顺序为:梅叶冬青 > 三叉苦 > 九节 > 桃金娘(图2);而不同灌木间叶片类胡萝卜素差异与叶绿素不同,九节叶片的类胡萝卜素含量为0.37 mg·g−1,均显著高于其它3种植物的(图2C)。桃金娘叶片的Chl a:Chl b的比值显著高于梅叶冬青,但与九节和三叉苦的比值无显著差异(图2E)。

图 2 马占相思人工林林下4种灌木叶片光合色素含量
Figure 2. Leaf photosynthetic pigments of four understory species in Acacia mangium plantation
-
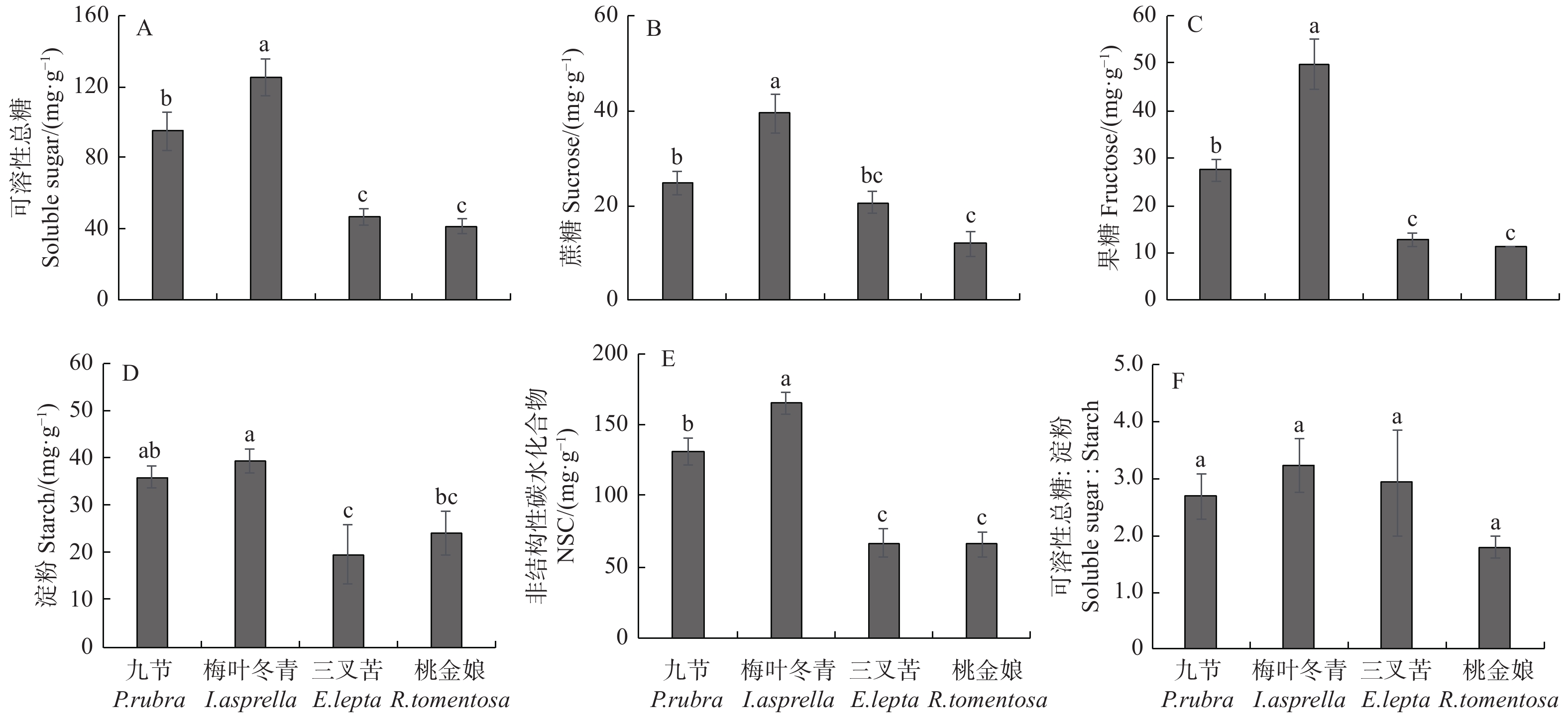
梅叶冬青叶片可溶性总糖最高(125.52 mg·g−1),显著高于其它3种灌木,分别是三叉苦和桃金娘叶片可溶性总糖含量的2.7和3.0倍(图3A)。梅叶冬青叶片的蔗糖和果糖浓度也显著高于其它3种灌木,大小顺序表现为:梅叶冬青 > 九节 > 三叉苦 > 桃金娘,其中,梅叶冬青叶片的蔗糖含量分别约为九节、三叉苦和桃金娘的1.6、1.9、3.3倍,果糖含量分别约为1.8、3.9、4.4倍(图3B、C)。梅叶冬青叶片的淀粉含量最高(39.41 mg·g−1),显著高于桃金娘和三叉苦叶片的(图3D)。梅叶冬青叶片非结构性碳水化合物(NSC)含量最高(164.93 mg·g−1),显著高于其它3个物种(图3E)。本研究中,4种灌木叶片可溶性总糖与淀粉比值无显著的种间差异(图3F)。

图 3 马占相思人工林林下4种灌木叶片非结构性碳水化合物含量
Figure 3. Leaf non-structural carbohydrates of four understory species in Acacia mangium plantation
-
九节和三叉苦根围土壤的pH值均显著高于桃金娘根围土的pH值,且桃金娘根围土壤pH值仅为3.38,表现出较强的酸性;除pH外,不同灌木根围土壤理化特性差异不大(表2)。马占相思人工林不同灌木根围土壤的有机质变化范围为9.12%~9.99 %,而桃金娘根围土壤的总氮和硝态氮含量最高。4种灌木根围土壤总磷含量变化范围为0.78~0.95 mg·g−1,而梅叶冬青根围土壤有效磷含量最低(表2)。
表 2 马占相思人工林林下4种灌木根围土壤理化特性
Table 2. Soil physiochemical characteristics of four understory species in Acacia mangium plantation
参数
Variables九节
P. rubra梅叶冬青
I. asprella三叉苦
E. lepta桃金娘
R. tomentosapH值 3.57 ± 0.02a 3.46 ± 0.09ab 3.59 ± 0.09a 3.38 ± 0.02b 有机质 SOM/% 9.99 ± 0.53a 9.49 ± 0.44a 9.98 ± 3.70a 9.12 ± 4.71a 总氮 TN/(g·kg−1) 3.96 ± 0.37a 3.78 ± 0.29a 3.83 ± 1.21a 5.42 ± 1.01a 总磷 TP/(g·kg−1) 0.95 ± 0.11a 0.78 ± 0.05a 0.91 ± 0.07a 0.88 ± 0.12a 有效磷 AP/(mg·kg−1) 7.64 ± 1.87a 4.96 ± 0.72a 6.92 ± 2.61a 6.27 ± 2.56a 硝态氮 NO3−/(mg·kg−1) 1.25 ± 0.56a 0.18 ± 0.06a 1.06 ± 0.69a 1.48 ± 0.16a 铵态氮 NH4+/(mg·kg−1) 0.24 ± 0.05a 0.56 ± 0.26a 0.39 ± 0.13a 0.17 ± 0.02a 无机氮 IN/(mg·kg−1) 1.49 ± 0.52a 0.74 ± 0.21a 1.45 ± 0.57a 1.65 ± 0.18a 注:表中不同小写字母表示不同物种间的显著差异(p < 0.05)。
Note: The different lower letters represent the significant difference among four species (p < 0.05). -
林下灌木叶片比叶质量与Chl a、Chl b、Chl(a+b)、氮含量、磷含量以及蔗糖含量呈显著负相关,而叶片氮含量与Chl a、Chl b和Chl(a+b)呈显著或极显著正相关;叶片氮磷比仅与果糖和淀粉含量呈显著正相关(表3)。
表 3 马占相思人工林林下4种灌木叶片氮磷含量与光合色素、非结构性碳水化合物的关系
Table 3. Relationships among leaf N and P concentrations, photosynthetic pigments and non-structural carbohydrates of four understory species in Acacia mangium plantation
参数
Variables比叶质量
Leaf mass per area/(g·m−2)叶片氮含量
Leaf N concentration/(mg·g−1)叶片磷含量
Leaf P concentration/(mg·g−1)叶片氮磷比
Leaf N:P ratios比叶质量 Leaf mass per area/(g·m−2) 1 −0.860** −0.629* 0.153 叶绿素a Chl a/(mg·g−1) −0.589* 0.736** 0.240 0.345 叶绿素b Chl b/(mg·g−1) −0.586* 0.619* −0.034 0.559 类胡萝卜素 Car/(mg·g−1) 0.114 −0.393 −0.454 0.168 总叶绿素 Chl (a+b)/(mg·g−1) −0.617* 0.715** 0.129 0.453 可溶性总糖 Soluble sugar/(mg·g−1) −0.446 0.233 −0.260 0.518 蔗糖 Sucrose/(mg·g−1) −0.639* 0.484 −0.025 0.418 果糖 Fructose/(mg·g−1) −0.500 0.293 −0.234 0.577* 淀粉 Starch/(mg·g−1) −0.137 0.045 −0.505 0.664* 注:**和*表示显著相关(p < 0.001和p < 0.05)。
Notes: ** and * indicate significantly correlated at p < 0.001 and p < 0.05, respectively. -
植物的生理特性是对环境适应的基础。研究表明,叶片的形态结构与植物适应环境的策略有关[25]。比叶质量与叶绿素含量、光合作用和植物的发育等方面密切相关[26],受到光照强弱、水分与养分条件影响[27]。有研究表明,比叶质量与光合能力呈负相关[28]。本研究发现,4种灌木的比叶质量与叶绿素a和b均呈显著负相关,与前述研究结果一致。本研究中,4种灌木的比叶质量表现出显著的种间差异,且桃金娘的比叶质量在4种灌木中最高,这有可能导致桃金娘在林下荫蔽环境中的光合能力比较低,说明桃金娘不能很好适应林分密度较大的人工林。实际上,桃金娘更倾向于分布在光照充足的环境中。相反,九节、梅叶冬青和三叉苦的比叶质量较小,可能是其更适应林下荫蔽环境的原因之一[15,29]。
在光合作用中,叶片光合色素在光能吸收、传递光能以及光能转化化学能中起重要作用。研究表明,叶绿素b分子具有捕获光能和传递光能的作用,少数特殊状态的叶绿素a分子有将光能转化为化学能的作用[30]。叶绿素含量高低是衡量植物光合作用强弱的重要指标,同时叶绿素含量易受植物体内氮素供应的影响[31]。本研究发现,4种灌木叶片氮含量与叶绿素a、b及总叶绿素均呈显著正相关,表明植物体内氮素供应能力越高,其光合色素含量越高。其中,叶绿素(a+b)反映了叶绿素含量多少,叶绿素a/b反应了光能利用效率。本研究中,叶绿素含量存在显著的种间差异,梅叶冬青和三叉苦叶绿素含量高于九节和桃金娘,表明梅叶冬青和三叉苦在林下环境中具有更好的光能吸收与转化能力,能更好地适应林下环境,这与前述梅叶冬青和三叉苦比叶质量较小而光合能力较强的结论相一致。
非结构性碳水化合物(NSC)是植物光合同化作用的主要产物之一,反应了植物在不同环境下的生存策略[32]。NSC主要以可溶性糖的形式溶于植物体内的液体中进行运输。可溶性糖是植物光合产物的主要存在形式,也是植物能直接利用并获取活动能量的物质,其浓度体现碳利用的大小[33],包含单糖、大部分二糖和多聚果糖,在植物生长发育中各组织的可溶性糖含量会发生周期性变化,极易受到环境因子的影响[34]。植物体内可溶性糖和淀粉含量高低可以反应植物光合作用强弱和营养贮藏状况。通常来说,耐荫树种贮存更高的NSC来增强自身与其它林下物种的光利用与竞争能力[35]。本研究中,4种灌木叶片NSC存在显著的种间差异,其中,梅叶冬青叶片中可溶性总糖、蔗糖、果糖和淀粉含量一般高于其它3个物种,表明梅叶冬青在马占相思林下有较好的耐荫蔽环境的能力。研究表明,可溶性糖是植物的直接功能物质,而淀粉是植物的长期储能物质,通常作为可溶性糖的一种中间转化形式[36-37]。本研究中,梅叶冬青叶片储存了较高的可溶性糖和淀粉,是其较好地适应林下荫蔽环境的原因之一。然而,本研究发现,人工林林下灌木叶片的NSC含量的变化规律,与季风常绿阔叶林林下层植物NSC的研究结果并不一致[37]。刘万德等研究指出,之所以季风常绿阔叶林即顶极群落的林下层植物NSC含量较低,是由于顶极群落中绝大多数的林下物种都为耐阴性植物,这些物种间耐阴性的个体差异并不能够完全导致它们体内的NSC产生差异,也反应出它们在光合作用及碳利用策略上的差异[37]。一般研究认为,耐阴树种比向阳树种具有更高的NSC含量[38-39],这进一步证明梅叶冬青叶片维持较高的NSC含量是其较好地适应了林分年龄较大的人工林林下荫蔽的环境基础。本研究中,桃金娘叶片NSC含量最低,导致其更倾向在人工林早期阶段开阔林地光照充足的环境生长。
结合以上几项指标,本研究中4种灌木对人工林林下荫蔽环境的适应能力大小为:梅叶冬青 > 九节 > 三叉苦 > 桃金娘。因此,在华南地区成熟人工林林下植被更新时,梅叶冬青可以作为首先考虑选择的物种,而桃金娘则可以作为人工林定植早期的可选择的灌木种类。
-
本研究发现,4种灌木根围土壤的pH值均小于4.0,且九节和三叉苦根围土壤pH值均显著高于桃金娘根围土壤pH值,表明4种灌木生长的土壤酸性较强;土壤有效氮和磷的含量相对较低,表明马占相思人工林林下灌木的生长可能受到氮磷养分的限制。氮和磷是植物生长发育所必需的营养元素,植物叶片氮磷含量及氮磷比可以反应出土壤的氮磷养分有效性状况[40-41]。据统计,全国植物叶片氮含量变化范围为11.8~28.6 g·kg−1及磷含量变化范围为0.46~2.45 g·kg−1[42]。本研究中,马占相思人工林林下4种灌木叶片全氮含量变化范围为13.02~26.43 g·kg−1、全磷含量为0.44~1.44 g·kg−1,与二者全国范围内植物叶片氮磷含量的数据基本符合。研究表明,温带森林生态系统中氮素通常成为限制植物生长的因子,而磷素更容易成为热带森林植物生长的限制因子[43]。研究表明,当植物叶片氮磷比小于14时,其生长极易受到氮的限制,而当氮磷比大于16时,其生长则极易受到磷的限制[44-45]。本研究中,马占相思纯林土壤有效磷含量为4.96~7.64 mg·kg−1,表明该人工林林地土壤磷有效性较低,植物生长受到土壤低磷的长期影响。有研究表明,同在该地区开展的人工林恢复试验表明,人工林恢复8年后,其林下土壤有效磷含量仅为3.30~4.07 mg·kg−1[46],这比较接近华南地区季风常绿阔叶林林下土壤有效磷含量的水平[47]。另外,从4种灌木叶片氮磷比看,梅叶冬青叶片的氮磷比最大(35),三叉苦叶片的氮磷比最小(20),均远远超过16,表明马占相思纯林林下4种灌木的生长均受到强烈的磷限制[48]。本研究发现,虽然4种灌木根围土壤的理化性质没有显著的差异,但由于灌木本身的生物学属性差异,它们受到氮和磷的限制表现出显著的种间差异(图1),尤其是从4种灌木叶片氮磷比的差异可以很好地验证这种种间差异。另外,本研究发现,叶片氮磷比与果糖和淀粉含量呈正相关,说明该地区的土壤氮磷供应水平可能调控着林下灌木的主要能量储存状态及其对环境的适应性。
因此,由土壤磷有效性较低导致人工林林下灌木生长受到的磷限制,可能对林下灌木生态功能的实现产生一定的抑制作用。这在人工林经营管理中需引起注意,尤其是从以木材生产为主的人工林经营方式向碳汇林或公益林经营方式的大规模转变时,需在一定程度上对大面积人工林进行生态系统养分调控与管理。
-
本文比较了广东鹤山马占相思人工林林下4种林下灌木叶片功能性状的差异。在4种林下灌木中,梅叶冬青叶片有较小的比叶质量、较高的氮含量、叶绿素含量及非结构性碳水化合物含量,这有助于其更好地适应林下荫蔽的环境,优于九节和三叉苦,且从这几个指标的结果看,桃金娘更适合生长在光环境较好的林下区域。本研究中4种灌木对人工林林下荫蔽环境适应能力的大小顺序为,梅叶冬青 > 九节 > 三叉苦 > 桃金娘。因此,在华南地区人工林定植的早期阶段,林分光照充足,引入桃金娘能更好地建立起稳定的植物群落,但随着乔木林分的生长,林内环境逐渐荫蔽,梅叶冬青、九节和三叉苦则能更好地适应人工林林下荫蔽环境,可考虑作为华南人工林林下植物更新时选择的可利用灌木树种。
马占相思人工林不同林下灌木叶片功能性状的差异
Variations of Leaf Functional Traits of Some Understory Species in Acacia mangium Plantation
-
摘要:
目的 探究马占相思(Acacia mangium Willd.)人工林不同林下灌木叶片功能性状的差异,为华南地区成熟人工林林下植被更新中物种选择提供科学参考。 方法 本研究以广东鹤山地区造林恢复34年的马占相思人工林林下4种灌木(九节(Psychotria rubra (Lour.) Poir.)、梅叶冬青(Ilex asprella (Hook. et Arn.) Champ. ex Benth var.as)、三叉苦(Evodia lepta (Spreng.) Merr.)和桃金娘(Rhodomyrtus tomentosa (Ait.) Hassk.))为研究对象,探讨4种灌木叶片功能性状的差异,试图阐明不同灌木对人工林林下环境适应的生理机制。 结果 (1)4种灌木中,桃金娘的比叶质量最大,为157.51 g·m−2,显著高于其它3个物种,梅叶冬青的比叶质量最小;三叉苦叶片的氮和磷含量最高,但氮磷比却最低;(2)梅叶冬青叶片叶绿素a和b含量都显著高于其它物种,但其类胡萝卜素含量却显著低于九节,而高于三叉苦和桃金娘;(3)梅叶冬青叶片可溶性总糖和非结构性碳水化合物(NSC)显著高于其它3个物种,淀粉含量显著高于三叉苦和桃金娘,说明维持较高的可溶性糖和淀粉有利于梅叶冬青适应林下的荫蔽环境。 结论 综合以上的各项指标,在4种灌木中,梅叶冬青较低的比叶质量可以维持较强的光合能力,且其较高的叶片非结构性碳水化合物含量也能为其适应林下荫蔽环境提供稳定的能量供应。因此,梅叶冬青可以作为华南地区成熟人工林林下植被更新的选择物种。 Abstract:Objective To study the variation of leaf functional traits of different understory shrubs in Acacia mangium Willd. plantation, so as to provide references for species selection for regeneration of understory vegetation in mature plantation in southern China. Method In this paper, four understory shrubs, including Psychotria rubra (Lour.) Poir., Ilex asprella (Hook. et Arn.) Champ. ex Benth var.as, Evodia lepta (Spreng.), and Rhodomyrtus tomentosa (Ait.) Hassk. in a thirty-four-year reforested A. mangium plantation in Heshan City, Guangdong Province, were used to study the variation of leaf functional traits and try to clarify the adapting mechanisms of understory species to light environment in plantation. Result (1) The leaf mass per area (LMA) of R. tomentosa was the highest, with the value of 157.51 g·m-2, which was significantly higher than those of the other species, and the LMA of I. asprella was the smallest; the leaf N and P concentrations of E. lepta was the highest, but with the lowest N: P ratios; (2) Both the Chl a and Chl b contents in I. asprella leaves were significantly larger than those of the other species, and the leaf carotinoid content in I. asprella was lower than that of P. rubra, but higher than that of E. lepta and R. tomentosa; (3) The leaf soluble sugar and non-structural carbohydrates (NSC) of I. asprella was significantly higher than those of the other species, and the leaf starch concentration of I. asprella was also significantly larger than those of E. lepta and R. tomentosa, indicating that I. asprella can well adapt to the shade environment via storing relatively higher soluble sugar and starch. Conclusion Among the four understory plants, the lower LMA of I. asprella can be helpful to maintain the strong photosynthetic capacity. Also, the higher NSC in leaf of I. asprella can provide stable energy for adapting shade environment. Therefore, I. asprella can be a selective species for regeneration of understory in mature plantation in southern China. -
Key words:
- understory
- / photosynthesis pigment
- / non-structural carbohydrates
- / plantations
- / southern China
-
图 1 马占相思人工林林下4种灌木叶片比叶质量、氮磷含量及氮磷比
Figure 1. Leaf mass per area (LMA), N and P concentrations, and N∶P ratios of four understory species in Acacia mangium plantation
图 2 马占相思人工林林下4种灌木叶片光合色素含量
Figure 2. Leaf photosynthetic pigments of four understory species in Acacia mangium plantation
图 3 马占相思人工林林下4种灌木叶片非结构性碳水化合物含量
Figure 3. Leaf non-structural carbohydrates of four understory species in Acacia mangium plantation
表 1 马占相思人工林林下4种灌木生长特性
Table 1. Growth characteristics of four understory shrubs in Acacia mangium plantation
种名
Species Name科
Family地径
Ground diameter/cm株高
Height/cm冠幅
Canopy/(cm × cm)密度
Density/(株·hm−2)盖度
Coverage/%九节Psychotria rubra 茜草科 Rubiaceae 4.04 ± 0.74 207.6 ± 28.9 197.8 × 174.6 309 10.5 梅叶冬青 Ilex asprella 冬青科 Aquifoliaceae 4.28 ± 0.28 292.8 ± 39.2 327.1 × 353.8 252 29.1 三叉苦 Evodia lepta 芸香科 Rutaceae 3.60 ± 0.67 255.3 ± 34.4 173.7 × 188.4 271 8.8 桃金娘 Rhodomyrtus tomentosa 桃金娘科 Myrtaceae 2.38 ± 0.47 203.0 ± 19.8 129.1 × 150.4 509 9.9  下载: 导出CSV
下载: 导出CSV
表 2 马占相思人工林林下4种灌木根围土壤理化特性
Table 2. Soil physiochemical characteristics of four understory species in Acacia mangium plantation
参数
Variables九节
P. rubra梅叶冬青
I. asprella三叉苦
E. lepta桃金娘
R. tomentosapH值 3.57 ± 0.02a 3.46 ± 0.09ab 3.59 ± 0.09a 3.38 ± 0.02b 有机质 SOM/% 9.99 ± 0.53a 9.49 ± 0.44a 9.98 ± 3.70a 9.12 ± 4.71a 总氮 TN/(g·kg−1) 3.96 ± 0.37a 3.78 ± 0.29a 3.83 ± 1.21a 5.42 ± 1.01a 总磷 TP/(g·kg−1) 0.95 ± 0.11a 0.78 ± 0.05a 0.91 ± 0.07a 0.88 ± 0.12a 有效磷 AP/(mg·kg−1) 7.64 ± 1.87a 4.96 ± 0.72a 6.92 ± 2.61a 6.27 ± 2.56a 硝态氮 NO3−/(mg·kg−1) 1.25 ± 0.56a 0.18 ± 0.06a 1.06 ± 0.69a 1.48 ± 0.16a 铵态氮 NH4+/(mg·kg−1) 0.24 ± 0.05a 0.56 ± 0.26a 0.39 ± 0.13a 0.17 ± 0.02a 无机氮 IN/(mg·kg−1) 1.49 ± 0.52a 0.74 ± 0.21a 1.45 ± 0.57a 1.65 ± 0.18a 注:表中不同小写字母表示不同物种间的显著差异(p < 0.05)。
Note: The different lower letters represent the significant difference among four species (p < 0.05).
下载: 导出CSV
表 3 马占相思人工林林下4种灌木叶片氮磷含量与光合色素、非结构性碳水化合物的关系
Table 3. Relationships among leaf N and P concentrations, photosynthetic pigments and non-structural carbohydrates of four understory species in Acacia mangium plantation
参数
Variables比叶质量
Leaf mass per area/(g·m−2)叶片氮含量
Leaf N concentration/(mg·g−1)叶片磷含量
Leaf P concentration/(mg·g−1)叶片氮磷比
Leaf N:P ratios比叶质量 Leaf mass per area/(g·m−2) 1 −0.860** −0.629* 0.153 叶绿素a Chl a/(mg·g−1) −0.589* 0.736** 0.240 0.345 叶绿素b Chl b/(mg·g−1) −0.586* 0.619* −0.034 0.559 类胡萝卜素 Car/(mg·g−1) 0.114 −0.393 −0.454 0.168 总叶绿素 Chl (a+b)/(mg·g−1) −0.617* 0.715** 0.129 0.453 可溶性总糖 Soluble sugar/(mg·g−1) −0.446 0.233 −0.260 0.518 蔗糖 Sucrose/(mg·g−1) −0.639* 0.484 −0.025 0.418 果糖 Fructose/(mg·g−1) −0.500 0.293 −0.234 0.577* 淀粉 Starch/(mg·g−1) −0.137 0.045 −0.505 0.664* 注:**和*表示显著相关(p < 0.001和p < 0.05)。
Notes: ** and * indicate significantly correlated at p < 0.001 and p < 0.05, respectively.
下载: 导出CSV
-
[1] 盛炜彤. 关于我国人工林长期生产力的保持[J]. 林业科学研究, 2018, 31(1):1-14. [2] 许传德. 从连续八次森林资源清查数据看我国森林经营[J]. 林业经济, 2013, 37(4):8-11, 36. [3] 童 冉, 周本智, 姜丽娜, 等. 我国杉木人工林可持续经营面临的问题及发展策略——基于全国分布区的调查[J]. 世界林业研究, 2019, 32(2):90-96. [4] 王志超, 杜阿朋, 陈少雄. 我国桉树人工林现状及可持续经营对策研究[J]. 桉树科技, 2012, 29(4):58-62. doi: 10.3969/j.issn.1674-3172.2012.04.012 [5] Rédei T, Csecserits A, Lhotsky B, et al. Plantation forests cannot support the richness of forest specialist plants in the forest-steppe zone[J]. Forest Ecology and Management, 2020, 461: 117964. doi: 10.1016/j.foreco.2020.117964 [6] Tavares A, Beiroz W, Fialho A, et al. Eucalyptus plantations as hybrid ecosystems: Implications for species conservation in the Brazilian Atlantic forest[J]. Forest Ecology and Management, 2019, 433: 131-139. doi: 10.1016/j.foreco.2018.10.063 [7] Wu J, Liu Z, Chen D, et al. Understory plants can make substantial contributions to soil respiration: Evidence from two subtropical plantations[J]. Soil Biology and Biochemistry, 2011, 43(11): 2355-2357. doi: 10.1016/j.soilbio.2011.07.011 [8] Cole E, Newton M, Bailey J D. Understory vegetation dynamics 15 years post-thinning in 50-year-old Douglas-fir and Douglas-fir/western hemlock stands in western Oregon, USA[J]. Forest Ecology and Management, 2017, 384: 358-370. doi: 10.1016/j.foreco.2016.11.003 [9] 王瑞华, 葛晓敏, 唐罗忠. 林下植被多样性、生物量及养分作用研究进展[J]. 世界林业研究, 2014, 27(1):43-48. [10] 李光敏, 陈伏生, 徐志文, 等. 间伐和林下植被剔除对毛竹林土壤氮矿化速率及其温度敏感性的影响[J]. 生态学报, 2019, 39(11):4106-4115. [11] Rivalland V, Calvet J C, Berbigier P, et al. Transpiration and CO2 fluxes of a pine forest: modelling the undergrowth effect[J]. Annales Geophysicae, 2005, 23(2): 994-1000. [12] 徐 馨, 王法明, 邹 碧, 等. 不同林龄木麻黄人工林生物多样性与土壤养分状况研究[J]. 生态环境学报, 2013, 22(9):1514-1522. doi: 10.3969/j.issn.1674-5906.2013.09.009 [13] 庞圣江, 张 培, 杨保国, 等. 广西大青山西南桦人工林林下植物多样性与稳定性[J]. 中南林业科技大学学报, 2018, 38(2):103-107, 113. [14] Heilmeier, Hermann. Functional traits explaining plant responses to past and future climate changes[J]. Flora, 2019, 254: 1-11. doi: 10.1016/j.flora.2019.04.004 [15] Poorter, Lourens. Leaf traits show different relationships with shade tolerance in moist versus dry tropical forests[J]. New Phytologist, 2009, 181(4): 890-900. doi: 10.1111/j.1469-8137.2008.02715.x [16] Cendrero M M P, Moran M S, Papuga S A, et al. Plant chlorophyll fluorescence: active and passive measurements at canopy and leaf scales with different nitrogen treatments[J]. Journal of Experimental Botany, 2015, 67(1): 275-286. [17] Michael C D, Anna S, Mariah S C, et al. Nonstructural carbon in woody plants[J]. Annual Review of Plant Biology, 2014, 65(1): 667-687. doi: 10.1146/annurev-arplant-050213-040054 [18] 孙美美, 宋变兰, 时伟宇, 等. 黄土丘陵区刺槐、辽东栎林生态系统碳汇功能特征[J]. 水土保持研究, 2020, 27(2):55-61. [19] 李 娟, 白彦锋, 彭 阳, 等. 湖南会同县杉木人工林管理碳汇的核算研究[J]. 林业科学研究, 2017, 30(3):436-443. [20] 李 萌, 陈永康, 徐浩成, 等. 不同间伐强度对南亚热带杉木人工林林下植物功能群的影响[J]. 生态学报, 2020, 40(14):1-9. [21] 王 锋. 海拔和造林密度对马占相思人工林生长的影响[J]. 广东林业科技, 2014, 30(1):42-45. [22] 林 雯, 李聪颖, 周 平. 广州城市森林六种典型林分碳积累研究[J]. 生态科学, 2019, 38(6):74-80. [23] Wang F, Li Z, Xia H, et al. Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China[J]. Soil Science and Plant Nutrition, 2010, 56(2): 297-306. doi: 10.1111/j.1747-0765.2010.00454.x [24] Buysse J, Merckx R. An improved colorimetic method to quantify sugar content of plant tissue[J]. Journal of Experiment Botany, 1993, 44: 1627-1629. [25] Díaz S, Lavorel S, de Bello F, et al. Incorporating plant functional diversity effects in ecosystem service assessments[J]. Proceedings of the National Academy of Science of the United States of America, 2007, 104(52): 20684-20689. doi: 10.1073/pnas.0704716104 [26] Ülo N. Components of leaf dry mass per area-thickness and density – alter leaf photosynthetic capacity in reverse directions in woody plants[J]. New Phytologist, 1999, 144(1): 35-47. doi: 10.1046/j.1469-8137.1999.00466.x [27] Ali A M, Darvishzadeh R, Skidmore A K, et al. Specific leaf area estimation from leaf and canopy reflectance through optimization and validation of vegetation indices[J]. Agricultural and Forest Meteorology, 2017, 236: 162-174. doi: 10.1016/j.agrformet.2017.01.015 [28] 刘西军, 陈 静, 徐小牛, 等. 桂花叶片SPAD、叶绿素含量和比叶重特征[J]. 安徽农业大学学报, 2013, 40(1):51-54. [29] 韦宏宇. 广西崇左市凤凰山林场尾叶桉和马尾松林下植被多样性特征分析[J]. 安徽农业科学, 2020, 48(2):127-129, 132. doi: 10.3969/j.issn.0517-6611.2020.02.034 [30] Bojović B, Stojanović J. Chlorophyll and carotenoid content in wheat cultivars as a function of mineral nutrition[J]. Archives of Biological Sciences, 2005, 57(4): 283-290. doi: 10.2298/ABS0504283B [31] Baker R N. Chlorophyll fluorescence: a probe of photosynthesis in vivo[J]. Annual Review of Plant Biology, 2008, 59(1): 89-113. doi: 10.1146/annurev.arplant.59.032607.092759 [32] 赵 镭, 杨海波, 王达力, 等. 浙江天童常见种幼苗的光合特性及非结构性碳水化合物储存[J]. 华东师范大学学报:自然科学版, 2011(4):35-44. [33] 于丽敏, 王传宽, 王兴昌. 三种温带树种非结构性碳水化合物的分配[J]. 植物生态学报, 2011, 35(12):1245-1255. [34] Davidson A, Keller F, Turgeon R. Phloem loading, plant growth form, and climate[J]. Protoplasma, 2011, 248(1): 153-163. doi: 10.1007/s00709-010-0240-7 [35] 杨 芳, 王振孟, 朱大海, 等. 常绿阔叶林林下6种木本植物叶片非结构性碳水化合物的动态特征[J]. 应用与环境生物学报, 2019, 25(5):1075-1083. [36] Hoch G, Richter A, Korner C. Non-structural carbon compounds in temperate forest trees[J]. Plant, Cell & Environment, 2003, 26(7): 1067-1081. [37] 刘万德, 苏建荣, 李帅锋, 等. 云南普洱季风常绿阔叶林不同林层非结构性碳水化合物特征[J]. 应用生态学报, 2018, 29(3):775-782. [38] Poorter L, Kitajima K. Carbohydrate storage and light requirements of tropical moist and dry forest tree species[J]. Ecology, 2007, 88(4): 1000-1011. doi: 10.1890/06-0984 [39] Myers J A, Kitajima K. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest[J]. Journal of Ecology, 2007, 95(2): 383-395. doi: 10.1111/j.1365-2745.2006.01207.x [40] Tanner E V J, Vitousek P M, Cuevas E. Experimental investigation of nutrient limitation of forest growth on wet tropical mountains[J]. Ecology, 1998, 79(1): 10-22. doi: 10.1890/0012-9658(1998)079[0010:EIONLO]2.0.CO;2 [41] Güsewell S. N: P ratios in terrestrial plants: variation and functional significance[J]. New Phytologist, 2004, 164(2): 243-266. doi: 10.1111/j.1469-8137.2004.01192.x [42] Han W, Fang J, Guo D, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2): 377-385. doi: 10.1111/j.1469-8137.2005.01530.x [43] Finzi AC, Canham C D. Sapling growth in response to light and nitrogen availability in a southern New England forest[J]. Forest Ecology and Management, 2000, 131(1): 153-165. [44] Gusewell S, Koerselman W, Verhoeven J T A. Biomass N: P ratios as indicators of nutrient limitation for plant populations in wetlands[J]. Ecological Applications, 2003, 13(2): 372-384. doi: 10.1890/1051-0761(2003)013[0372:BNRAIO]2.0.CO;2 [45] Davidson E A, Reis de Carvalho C J, Vieira I C G, et al. Nitrogen and phosphorus limitation of biomass growth in a tropical secondary forest[J]. Ecological Applications, 2004, 14(4): 150-163. [46] Mo Q, Li Z, Zhu W, et al. Reforestation in southern China: revisiting soil N mineralization and nitrification after 8 years restoration[J]. Scientific Reports, 2016, 6: 19770. doi: 10.1038/srep19770 [47] Liu L, Zhang T, Gilliam F S, et al. Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest[J]. PLoS ONE, 2013, 8(4): e61188. doi: 10.1371/journal.pone.0061188 [48] Li Y, Niu S, Yu G. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: a meta-analysis[J]. Global Change Biology, 2016, 22(2): 934-943. doi: 10.1111/gcb.13125 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5676
- HTML全文浏览量: 3494
- PDF下载量: 52
- 被引次数: 0
