

-
薄壳山核桃(Carya illinoensis (Wangenh.) K. Koch)属胡桃科(Juglandaceae)山核桃属(Carya),又称碧根果、长山核桃、美国山核桃,自然分布于美国和墨西哥北部,为一种多年生高大落叶乔木。现以美国为产业中心,分为东南、中南、西南和北部四大商业化产区,涵盖全美24个州[1]。薄壳山核桃是典型的果材兼用型树种。其坚果产量高、壳薄个大、营养丰富,是大众颇为喜爱的保健食品;种仁含油量高达70%以上,不饱和脂肪酸超97%,优于茶油、花生油等,是世界重要的木本油料植物[1-3]。薄壳山核桃因其世代周期长、树形优美、树势挺拔,也是重要的景观绿化树种;其木材结构细密、坚固强韧、纹理美观,可用于军工、建筑以及制作高档家具和工艺品,属于珍贵用材树种[1-3]。
我国引种薄壳山核桃已有百年历史,前期主要引进种子及苗木,后期开始引进系列良种,并进行自主选育与栽培推广。据不完全统计,近年来江苏、浙江、安徽、四川、云南等省份陆续引进和保存的薄壳山核桃品种超过100个[4],相继审(认)定的良种超过40个,包括美国引进与自主选育的品种[5]。但薄壳山核桃品种选育多利用种子繁殖,后代变异大且童期较长,目前栽培推广的大多仍是从美国引进的品种。由于薄壳山核桃品种引进均为种条,形态相似难以区分,且前期引种历史较长、引种程序不规范,各地间相互频繁引种难免造成品种间相互混杂混淆,甚至直接改换国外品种名称,造成同物异名或同名异物的现象经常发生,这也给薄壳山核桃品种种质评价、遗传改良以及面上推广工作带来极大的不便。因此,对薄壳山核桃品种进行准确的评价与鉴定,不仅有助于保护林木植物新品种权、保障所有权人的利益,而且能从根本上保证我国薄壳山核桃产业的商业化发展。
分子标记技术不受外界环境及人为因素的影响,可直接反映不同个体间DNA水平上的差异,实验过程简便快捷、结果稳定可靠,已广泛应用于林木遗传学及基因组学的各个领域。在薄壳山核桃亲缘关系分析与品种鉴定方面,前期已有RAPD[6-8]、ISSR[9]、SRAP[10]、SSR[11-12]、SNP[13]、DNA条形码[14]和叶绿体基因组[15-16]等众多标记的研究。与显性标记相比,SSR标记在基因组中各个区域广泛分布、数量极大、变异丰富,且为共显性遗传,是目前使用最广泛的一种分子标记[17],尤其是结合荧光标记高通量基因分型技术,已成功应用于杨树[18]、柳树[19]、栎树[20]、刺槐[21]、含笑[22]、桂花[23]等多个木本植物指纹图谱的构建研究。国内薄壳山核桃分子生物学的研究起步较晚,在指纹图谱构建方面尚未见相关报道。鉴于此,本研究选取薄壳山核桃及其近缘种开发的SSR标记,经筛选后利用荧光标记并对25个美国引进品种进行基因分型,用以构建指纹图谱并分析其亲缘关系,旨在建立一种快速、稳定、可靠的薄壳山核桃基因分型体系,为薄壳山核桃品种改良工作提供有益参考,同时也为薄壳山核桃遗传资源的鉴定与品种保护提供理论依据。
-
本研究所用25个薄壳山核桃品种均从美国引进,定植于江苏省句容市南京林业大学薄壳山核桃种质资源圃内(表1)。采集所有品种幼嫩叶片,利用试剂盒(天根生化科技有限公司,DP305)按说明书步骤提取叶片总DNA。分别使用0.8%的琼脂糖凝胶电泳和分光光度计检测总DNA的完整性、纯度与浓度。
表 1 试验材料
Table 1. Experimental materials
编号
Code品种
Variety遗传背景
Genetic background雌雄异熟类型
Dichogamy type1 ‘Cape Fear’ ‘Schley’ 自由授粉子代 Open pollination progeny of ‘Schley’ 雄先型 Protandrous 2 ‘Choctaw’ ‘Success’ × ‘Mahan’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Mahan’ 雌先型 Protogynous 3 ‘Desirable’ ‘Russell’ × ‘Success’ 杂交子代 Hybrid progeny of ‘Russell’ × ‘Success’ 雄先型 Protandrous 4 ‘Elliott’ 实生苗 Seedling 雌先型 Protogynous 5 ‘Forkert’ ‘Success’ × ‘Schley’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Schley’ 雌先型 Protogynous 6 ‘Gloria Grande’ ‘Stuart’ 子代 Seedling of ‘Stuart’ 雌先型 Protogynous 7 ‘Hirschi’ 实生苗 Seedling 雄先型 Protandrous 8 ‘Jackson’ ‘Success’ × ‘Schley’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Schley’ 雄先型 Protandrous 9 ‘Kanza’ ‘Major’ × ‘Shoshoni’ 杂交子代 Hybrid progeny of ‘Major’ × ‘Shoshoni’ 雌先型 Protogynous 10 ‘Kiowa’ ‘Mahan’ × ‘Desirable’ 杂交子代 Hybrid progeny of ‘Mahan’ × ‘Desirable’ 雌先型 Protogynous 11 ‘Mahan’ ‘Schley’ 实生苗 Seedling of ‘Schley’ 雌先型 Protogynous 12 ‘Major’ 实生苗 Seedling 雄先型 Protandrous 13 ‘Mandan’ ‘BW-1’ × ‘Osage’ 杂交子代 Hybrid progeny of ‘BW-1’ × ‘Osage’ 雌先型 Protogynous 14 ‘Mohawk’ ‘Success’ × ‘Mahan’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Mahan’ 雌先型 Protogynous 15 ‘Nacono’ ‘Cheyenne’ × ‘Sioux’ 杂交子代 Hybrid progeny of ‘Cheyenne’ × ‘Sioux’ 雌先型 Protogynous 16 ‘Navaho’ ‘Apalachee’ × ‘Wichita’ 杂交子代 Hybrid progeny of ‘Apalachee’ × ‘Wichita’ 雄先型 Protandrous 17 ‘Oconee’ ‘Schley’ × ‘Barton’ 杂交子代 Hybrid progeny of ‘Schley’ × ‘Barton’ 雄先型 Protandrous 18 ‘Pawnee’ ‘Mohawk’ × ‘Starking Hardy Giant’ 杂交子代 Hybrid progeny of ‘Mohawk’ × ‘Starking Hardy Giant’ 雄先型 Protandrous 19 ‘Schley’ 实生苗 Seedling 雌先型 Protogynous 20 ‘Shoshoni’ ‘Odom’ × ‘Evers’ 杂交子代 Hybrid progeny of ‘Odom’ × ‘Evers’ 雌先型 Protogynous 21 ‘Sumner’ ‘Schley’ 实生苗 Seedling of ‘Schley’ 雌先型 Protogynous 22 ‘Surprize’ 实生苗 Seedling 雄先型 Protandrous 23 ‘Tejas’ ‘Mahan’ × ‘Risien #1’ 杂交子代 Hybrid progeny of ‘Mahan’ × ‘Risien #1’ 雌先型 Protogynous 24 ‘Western’ ‘Longfellow’ 实生苗 Seedling of ‘Longfellow’ 雄先型 Protandrous 25 ‘Wichita’ ‘Halbert’ × ‘Mahan’ 杂交子代 Hybrid progeny of ‘Halbert’ × ‘Mahan’ 雌先型 Protogynous -
从前期已发表的薄壳山核桃及其近缘种SSR标记中挑选54对通用性较高的引物,并随机选择16个品种为模板进行引物初筛。经高分辨率琼脂糖凝胶电泳检测,最终确定10对SSR标记用于后续分析(表2)。用荧光基团标记所有引物前项5′端,交由生物公司进行合成。
表 2 10对SSR引物信息
Table 2. Details of 10 SSR primer pairs
编号
Code位点
Locus前项引物
Forward primer后项引物
Reverse primer基序
Motif参考文献
Reference1 PM-CIN4 GGCATCAGAGAAGGCTCCT CTCACCCGTCTCTAGGGCTA (CTT)12 [11] 2 PM-GA31 TGAACTCCAAAAGCCTCCTCTC GTATTTGTATTTTTTCCTTGAGCTTTCTC (CT)12 [11] 3 PM-GA38 AAAAGTTTTAGGGTTGTTTGCTCTCT GTAAAGCCTACAACCTACAACAGTCTATG (CT)12 [11] 4 PM-GA41 TCTTCAGAAAAAACCCTTACCTCTCT GAAAAATATAAACTCCCATACTACCCACAT (CT)9 [11] 5 Cc19 AAACCTTGGCATAGTCATTTGAGA GCTTTGTCAACTTTGTTTTGGGTGT (TC)8 [24] 6 Cc4 GCTCCAAGCGAAAGTCAAGT TCATAAACCAACGCCAAAGA (TC)6 [24] 7 WGA70 TGTAATTGGGGAATGTTGCA TGGGAGACACAATGATCGAA (GA)14 [25] 8 BFU-Jr19 CAGAAAGCACGTGAACCAGA AACACTCACGCACCTTCATTT (TA)10 [26] 9 BFU-Jr82 TTGCCTCTCTATAGGCTTTTTAGC TGAGTGTCTGGACAGCAAGG (CT)7 [26] 10 Zm26 TTCTTCGTCCATACCCACC ACCCCATTAGCGACCTTTA (TC)22 [27] 采用10 μL PCR反应体系进行引物初筛,包含1 μL DNA模板,各0.4 μL前后项引物,5 μL PCR Mix(上海生工Taq PCR Mix 预混液,2 ×,含蓝染料),3.2 μL ddH2O。PCR扩增程序按以下步骤进行:首先94℃预变性4 min;然后94℃变性30 s,56℃退火30 s,72℃延伸1 min,循环35次;最后72℃延伸10 min。
-
基因分型PCR反应为25 μL体系,包含1 μL DNA模板,各1 μL前后项引物,1 μL dNTP,2.5 μL Taq Buffer(含MgCl2),0.5 μL Taq酶,18 μL ddH2O。PCR扩增程序按以下步骤进行:首先95℃预变性3 min;其次94℃变性30 s,60℃退火30 s,72℃延伸30 s,循环10次;随后94℃变性30 s,55℃退火30 s,72℃延伸30 s,循环35次;最终72℃延伸8 min。PCR产物中加入HiDi与LIZ 500内标,经98℃变性5 min后迅速移至冰上进行冷却,随后放置于ABI 3730 XL全自动DNA测序仪内进行电泳检测。
-
利用测序仪自带的Genemapper软件统计各位点基因分型数据;使用MSA软件计算各位点等位基因数(A)和多态信息含量(PIC)等数据;使用NTSYS-pc软件中Qualitative data模块计算两两个体间遗传相似系数(GS):GS = 2Nij /(Ni + Nj),式中:Nij为i和j共有的谱带数,Ni和Nj为i和j个体的谱带数,以clustering程序中SHAN进行非加权组平均法(UPGMA)聚类。
-
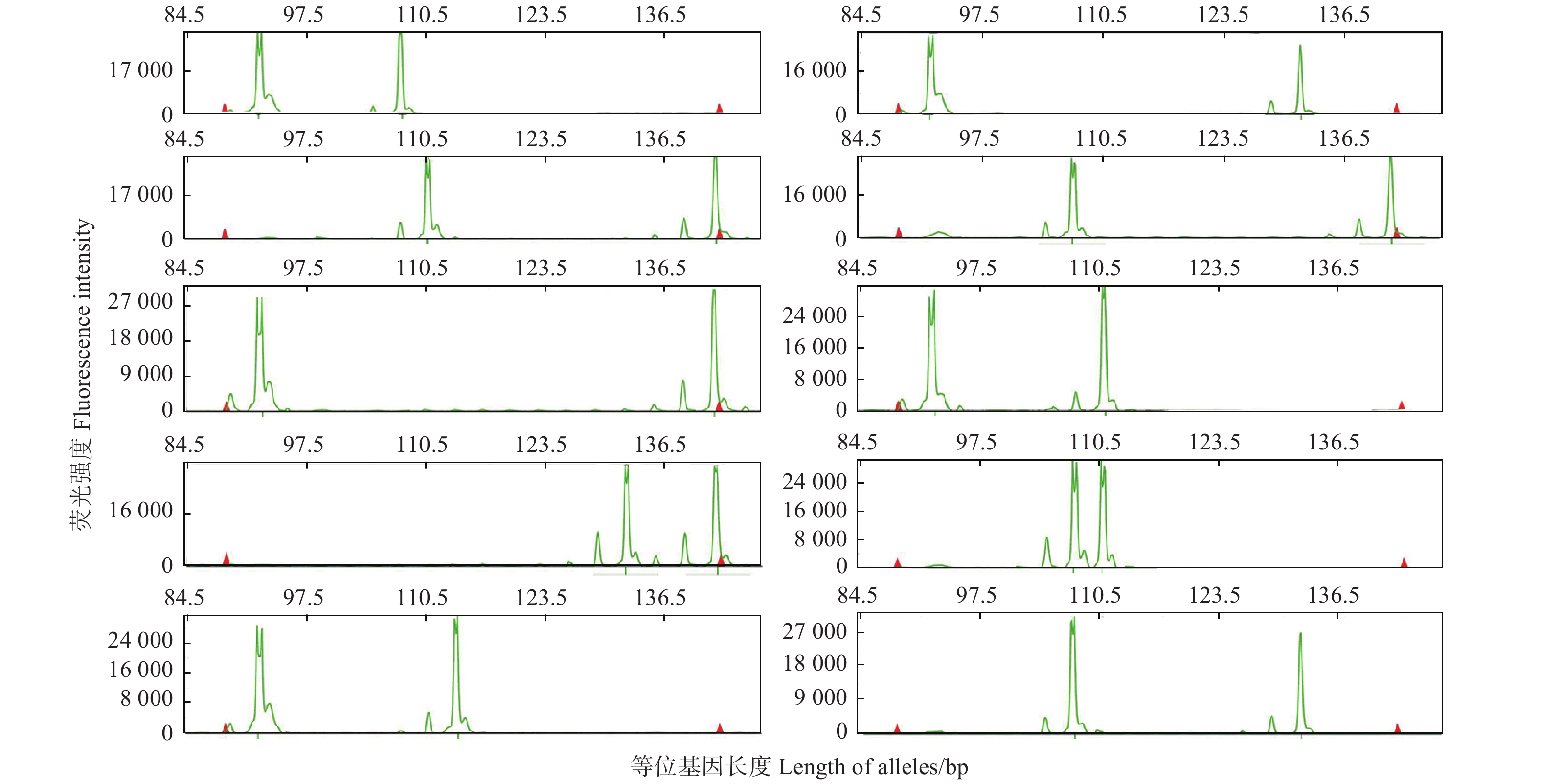
以25个薄壳山核桃品种为模板,检验10对荧光SSR标记多态性情况。表3表明 :10对SSR标记共扩增出68条等位片段,平均6.8个,其中,位点BFU-Jr19扩增的等位基因数最少(3个),位点Cc19扩增的等位基因数最多(12个)。10个SSR标记的多态信息含量(PIC)的变化范围为0.291 0~0.843 5,平均0.588 3,其中,位点Cc19的PIC值最高,位点BFU-Jr82的PIC值最低。图1为位点PM-CIN4在部分薄壳山核桃品种中的基因分型结果。
表 3 10对SSR标记多态性检测
Table 3. Polymorphism detection of the 10 pairs of SSR markers
位点
Locus等位基因数
No. of allele等位基因长度
Length of allele/bp多态信息含量
PICPM-CIN4 9 89~143 0.763 2 PM-GA31 10 84~112 0.733 4 PM-GA38 6 82~98 0.681 5 PM-GA41 4 82~88 0.437 3 Cc19 12 86~120 0.843 5 Cc4 6 185~197 0.467 7 WGA70 6 173~195 0.644 3 BFU-Jr19 3 272~290 0.343 0 BFU-Jr82 5 271~289 0.291 0 Zm26 7 201~219 0.678 5 合计 Total 68 平均 Average 6.8 0.588 3 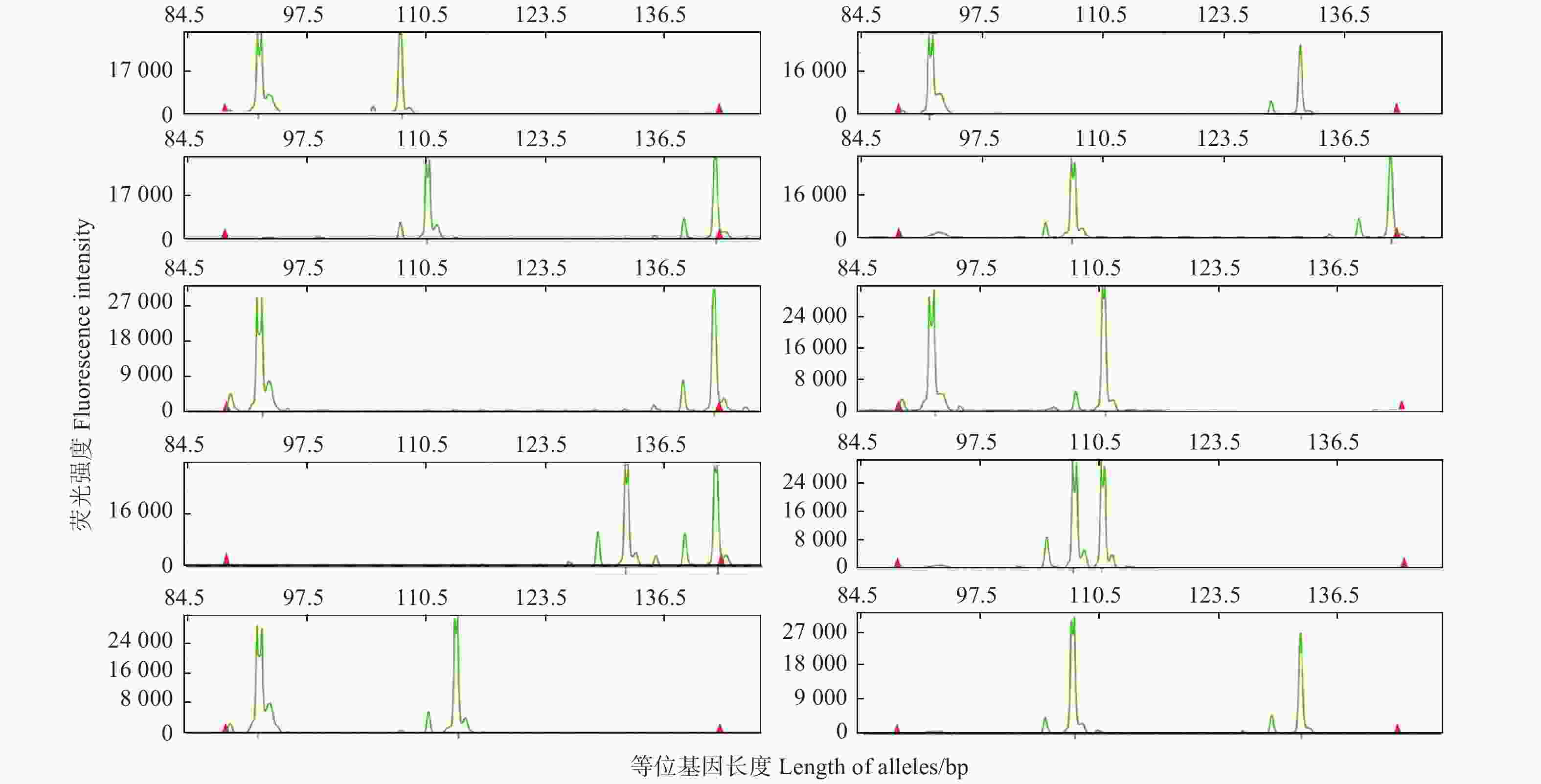
图 1 位点PM-CIN4在薄壳山核桃部分品种中的基因分型
Figure 1. Genotyping by locus PM-CIN4 for partial varieties of Pecan.
-
对10个SSR位点扩增的等位片段做进一步统计分析,经不同标记间相互组合,最终确认4对核心引物的组合可用于25个薄壳山核桃品种指纹图谱的构建。表4表明:标记Cc19可以区分11个品种,标记PM-GA31能区分10个品种,标记PM-CIN4能区分6个品种。通过引物Cc19、PM-GA31与PM-CIN4的组合可区分21个薄壳山核桃品种,加入标记PM-GA41后,4对引物组合可完全区分薄壳山核桃的25个品种。
表 4 25个薄壳山核桃品种的指纹图谱
Table 4. Fingerprint of 25 varieties of Pecan
品种
Varieties核心引物 Core primers PM-CIN4 PM-GA31 PM-GA41 Cc19 ‘Cape Fear’ 107/143 102 84/86 118 ‘Choctaw’ 131/143 98/100 84/86 118 ‘Desirable’ 131/143 98 84/86 108/116 ‘Elliott’ 110/143 94/102 84/86 98/118 ‘Forkert’ 107/143 102 84/86 96/98 ‘Gloria Grande’ 107 94 82/88 88 ‘Hirschi’ 104/107 94 84/86 94/96 ‘Jackson’ 92/107 94 84/86 96/98 ‘Kanza’ 107/143 94 84/86 96/118 ‘Kiowa’ 104/107 94 86 94/96 ‘Mahan’ 107/143 94 84/86 98 ‘Major’ 110/143 84/86 84/86 88/96 ‘Mandan’ 89/131 84/88 86/88 92/98 ‘Mohawk’ 92/107 94 84/86 94/98 ‘Nacono’ 92/113 94 84/86 98/118 ‘Navaho’ 107/131 94 86 96/120 ‘Oconee’ 107/143 102 84/86 116/118 ‘Pawnee’ 110/143 88 84/86 114/116 ‘Schley’ 92/107 92/94 84/86 94/98 ‘Shoshoni’ 92/107 86/88 86 86/116 ‘Sumner’ 131/143 96/98 86 108/116 ‘Surprize’ 107/128 84 84/86 96/100 ‘Tejas’ 92/143 88 86 94/96 ‘Western’ 92/107 94 86 94/98 ‘Wichita’ 110/143 112 84/86 94 -
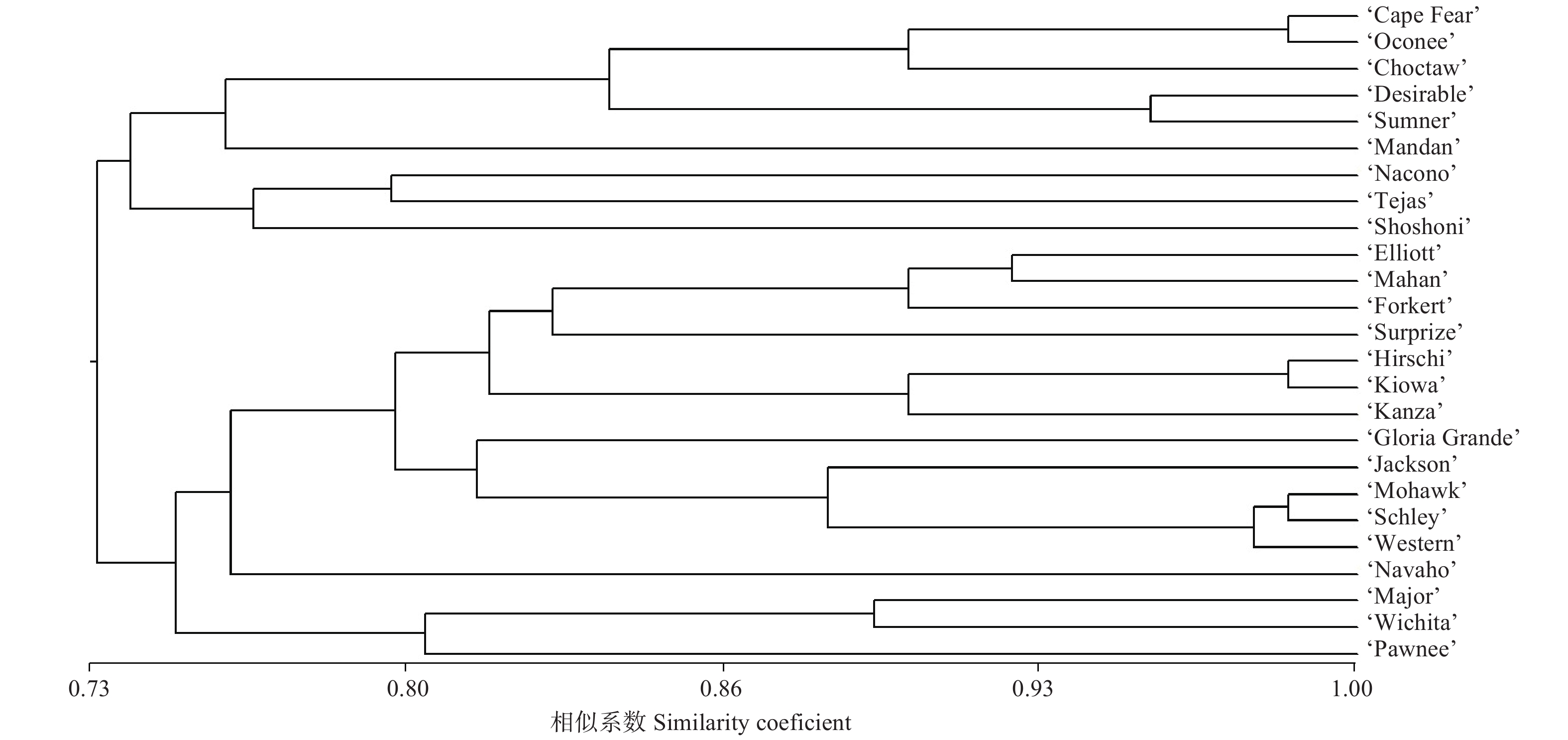
计算各品种间遗传相似系数,并按UPGMA法进行聚类(图2)。25个薄壳山核桃品种间遗传相似系数在0.62~0.99之间,并聚类成2个大的类群。
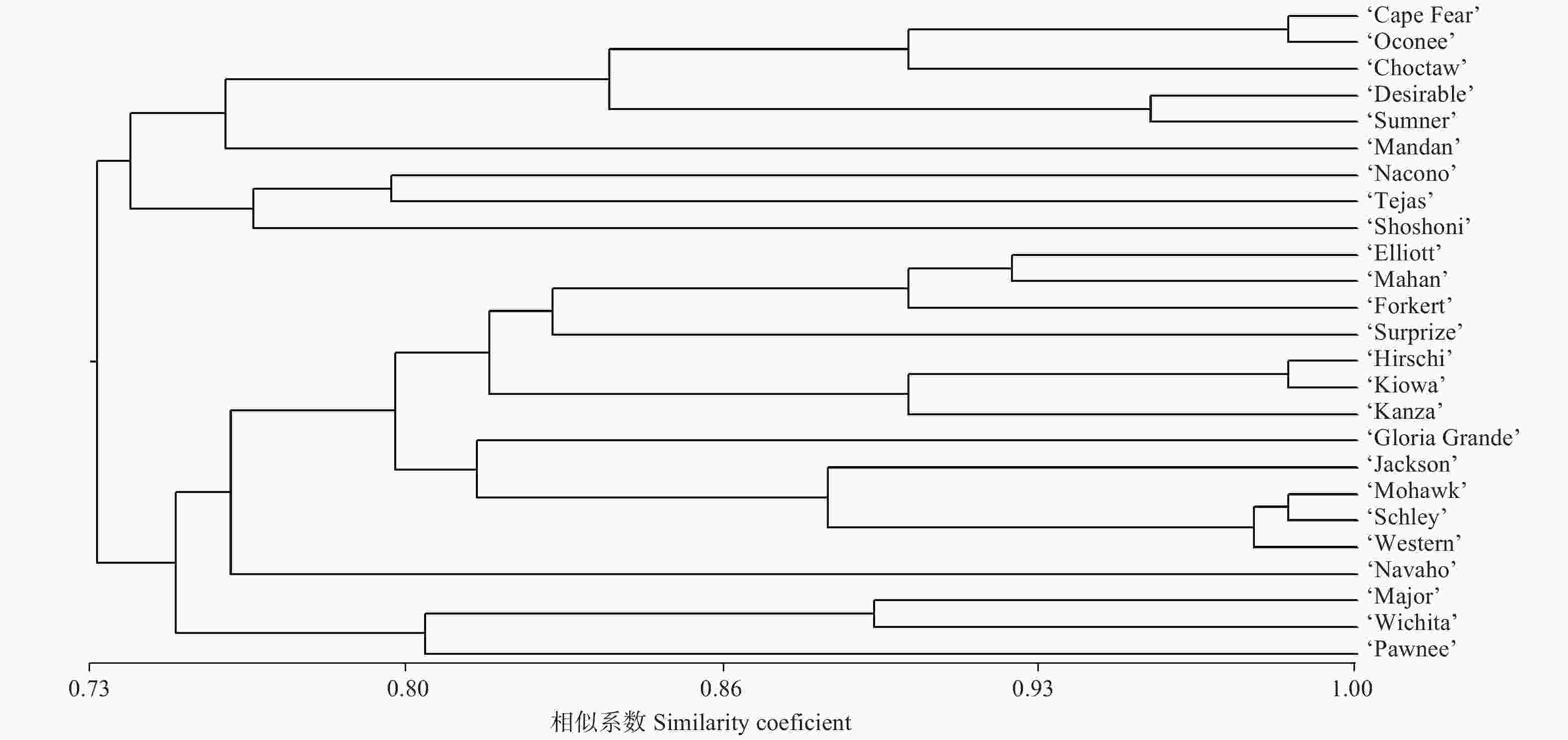
图 2 25个薄壳山核桃品种的聚类
Figure 2. Dendrogram of 25 varieties of Pecan
第一个大的类群包含9个薄壳山核桃品种,其中,‘Schley’自由授粉子代‘Cape Fear’与‘Schley’杂交子代‘Oconee’聚成一个小的分枝,并与品种‘Choctaw’聚在一起,而‘Choctaw’的一个亲本‘Mahan’也是‘Schley’实生子代。其他品种如‘Mandan’、‘Nacono’、‘Tejas’和‘Shoshoni’因杂交亲本差异较大,各品种间的亲缘关系也相对较远。第二个大的类群包含16个薄壳山核桃品种,其中,来源于实生苗的品种‘Hirschi’与‘Kiowa’亲缘关系较近;品种‘Mohawk’、‘Schley’、‘Western’和‘Jackson’聚类成一个小的分枝,其中‘Mohawk’一个亲本为‘Mahan’,而‘Mahan’也是‘Schley’实生子代;‘Western’又名‘Western Schley’,虽然来源于实生苗选育,但其性状与‘Schley’极为相似;而‘Jackson’为杂交选育,其中的一个亲本即为‘Schley’。其余几个品种如‘Navaho’、‘Wichita’和‘Pawnee’虽然亲缘关系相对较远,但追溯其亲本来源,均与品种‘Schley’相关。
-
薄壳山核桃为雌雄同株,一般通过控制授粉进行杂交,再收取种子进行子代测定,进而选育新的品种。从表1可以看出,本研究选取的25个薄壳山核桃品种中,有一半的品种直接与‘Schley’相关,或是通过‘Schley’的子代进一步杂交选育的品种,可见薄壳山核桃品种间的遗传基础较狭窄,品种间的形态极为相似,很难通过表型性状进行准确的鉴定,而基于分子标记技术的指纹图谱无疑是进行快速鉴定的重要技术手段。同时,基于林木新品种权申请与保护的需要,也可提交分子标记指纹图谱作为辅助证据[28]。SSR为共显性标记,基因分型时能区分纯合子与杂合子,且SSR标记变异程度高,不但能够区分亲缘关系较近的种,甚至能够鉴定同一个杂交组合中的不同个体,如在核桃中就有过类似报道[29]。此外,如图1所示,基于荧光SSR标记联合高通量测序仪的基因分型技术,相比传统的PAGE胶更加直观、快速、准确,特别是针对差异较小的等位片段有着更高的分辨率[30]。
SSR标记广泛分布于植物基因组的各个区域,通常包含EST-SSR、基因组SSR(genomic SSR)和叶绿体SSR(cpSSR)3种类型。EST-SSR来源于基因组转录区域,其序列相比基因组SSR更加保守,种间通用性较高但多态性较低[31]。有研究表明:EST-SSR标记不仅可应用于种间及属间[32-33],甚至在亚科间与科间均表现出较高的通用性[34-35]。本研究中有2个EST-SSR标记(Cc19、Cc14)来源于山核桃(C. cathayensis,胡桃科山核桃属),3个EST-SSR标记(BFU-Jr19、BFU-Jr82、Zm26)来源于核桃(Juglans regia,胡桃科核桃属),可见EST-SSR标记在同属不同种间、甚至属间均表现出较好的通用性用性。此外,Li等[24]的研究也发现:从山核桃中开发的311个标记有63.02%可以用于山核桃属的其他树种,如薄壳山核桃、大别山山核桃(C. dabieshanensis)和湖南山核桃(C. hunanensis)等。从核桃中开发的EST-SSR标记在核桃属(Juglans)、山核桃属(Carya)以及喙核桃属(Annamocarya)不同树种中通用性均较好[26]。
本研究选取的10个SSR标记中,有5个标记为基因组SSR标记(PM-CIN4、PM-GA31、PM-GA38、PM-GA41、WGA70),5个为EST-SSR标记(Cc19、Cc4、BFU-Jr19、BFU-Jr82、Zm26)。从表3中PIC数据可以看出,尽管EST-SSR标记Cc19的值最大,但总体而言,基因组SSR标记的多态性相比较于EST-SSR标记要高。通常认为,高度多态位点的PIC值一般大于0.5,中度多态位点的PIC值一般在0.25~0.5之间,低度多态位点的PIC值一般小于0.25[36]。本研究中,有6个标记PIC值大于0.5,4个标记PIC值在0.25~0.50之间,总体标记的多态性较高。
聚类分析中,25个薄壳山核桃品种聚成了2个大的分枝,各个品种之间的聚类与遗传背景之间没有明显的对应关系。虽然一些品种从遗传背景看,亲缘关系应该较近,如‘Forkert’和‘Jackson’均为‘Success’与‘Schley’的杂交子代,‘Choctaw’与‘Mohawk’均为‘Success’与‘Mahan’的杂交子代,但聚类时并没有聚在一起。又如‘Cape Fear’、‘Mahan’与‘Sumner’,均为‘Schley’子代,尽管只有一个亲本不一样,但彼此间的亲缘关系相差较远。类似的情况在Conner等[6]以及Grauke等[11]的研究中也有过报道。推测可能是由于薄壳山核桃为多年生高大乔木,世代周期较长,遗传背景复杂且高度杂合,杂交子代遗传分化较严重造成的。此外,标记的数量、类型以及不同的聚类方法均会对聚类结果产生一定的影响。因此,在后期的研究中,一方面要加入各个品种的杂交亲本,另一方面要挑选覆盖全基因组范围、一定数量、效率更高的标记,才能最大程度揭示薄壳山核桃品种间亲缘关系的真实水平。
-
薄壳山核桃品种间遗传基础较为狭窄,外观形态极为相似,仅从表型性状难以进行准确区分。本研究基于荧光SSR标记开展了25个美国薄壳山核桃品种亲缘关系分析与指纹图谱构建研究,筛选出的10对SSR引物多态性较高,建立的基因分型体系稳定且可靠,可为薄壳山核桃种质鉴定及品种保护提供有效方法,也可为薄壳山核桃遗传资源的引进及杂交育种亲本的选配提供科学依据。
薄壳山核桃品种亲缘关系分析与指纹图谱构建
Genetic Relationship Analysis and Fingerprint Construction of Carya illinoensis Varieties
-
摘要:
目的 基于荧光SSR标记结合高通量毛细管电泳技术,建立一种快速、高效、稳定、准确的薄壳山核桃基因分型体系,用以分析各品种间亲缘关系并构建指纹图谱,旨在为薄壳山核桃品种鉴定与新品种保护提供理论依据,也为薄壳山核桃种质资源评价、品种选育与推广提供有益参考。 方法 选取薄壳山核桃及其近缘种54对SSR引物进行初筛,最终确定10对标记合成荧光引物用于后续分析。基于毛细管电泳技术对25个薄壳山核桃品种进行基因分型,利用软件统计位点数据并计算各个位点的等位基因数(A)和多态信息含量(PIC)。利用SSR标记间的相互组合构建薄壳山核桃不同品种的指纹图谱。通过等位片段的转化对薄壳山核桃品种进行聚类,并分析其亲缘关系。 结果 10对荧光SSR标记共扩增出68条等位片段,平均为6.8个;位点Cc19最多,有12个等位基因,位点BFU-Jr19最少,有3个等位基因。多态信息含量(PIC)变化范围为0.2910~0.8435(平均为0.5883)。优选的4对核心引物Cc19、PM-GA31、PM-CIN4和PM-GA41组合能完全区分25个薄壳山核桃品种。25个品种遗传相似系数在0.62~0.99之间,并聚类成2个大的类群,部分亲缘关系较近的品种聚类在一起,部分品种聚类不能与遗传背景完全对应。 结论 与传统的显性标记及聚丙烯酰胺凝胶电泳分型技术相比,荧光SSR标记结合毛细管电泳技术构建薄壳山核桃品种指纹图谱切实可行,通量大且速度快,结果稳定可靠,通过标记组合可有效的对不同品种进行区分。在品种亲缘关系分析中,建议增加杂交亲本数量,并在全基因组范围内选取一定数量效率更高的标记,以便最大程度地揭示薄壳山核桃品种间亲缘关系的真实水平。 Abstract:Objective Since the morphological appearances among pecan (Carya illinoensis) varieties are extremely similar due to the narrow genetic basis of the crossing parents, making it difficult to precisely identify by phenotypic characters only, this study aims at elucidating the genetic relationships and constructing fingerprint among pecan varieties, and establishing a rapid, efficient, stable, and accurate genotyping system for pecan based on fluorescent SSR markers combining high throughput capillary electrophoresis technology. Method A total of 54 SSR primers were selected from pecan and related species for preliminary screening and ten of them were labeled by fluorophore for further analysis. 25 pecan varieties were genotyped by capillary electrophoresis, and the number of allele (A) and polymorphic information content (PIC) of each locus was scored and calculated by software. The fingerprint of pecan varieties was constructed using different combination of SSR markers and the genetic relationships among varieties were also investigated through cluster analysis. Result A total of 68 alleles were detected by ten pairs of SSR markers with an average of 6.8 alleles. The most (12 alleles) were screened at the Cc19 locus and the least (3 alleles) were obtained at the BFU-Jr locus. The polymorphic information content varied with locus from 0.2910 to 0.8435 (mean 0.5883). With the four optimal pairs of core primers, all of the 25 pecan varieties could be completely distinguished by the primer combinations of Cc19, PM-GA31, PM-CIN4, and PM-GA41. The cluster analysis demonstrated that the similarity coefficient of the 25 pecan varieties varied from 0.62 to 0.99. Two main clades were formed, in which some related varieties could be clustered together and some of them could not be completely corresponding with genetic background. Conclusion Compared with traditional dominant markers and polyacrylamide gel electrophoresis, the genotyping technology consisting of fluorescence-labeled SSR primers and capillary electrophoresis is quite practical with high throughput, fast speed, and stable and reliable result. The different varieties can be efficiently identified by the combination of core markers. In order to deeply reveal the real architecture of genetic relationship among pecan varieties, it is suggested that more crossing parents should be added and a certain number of more efficient markers covered the whole genome need to be selected during the analysis of genetic relationship. -
Key words:
- Carya illinoensis
- / fluorescent markers
- / SSR
- / genetic relationship
- / fingerprint
-
图 1 位点PM-CIN4在薄壳山核桃部分品种中的基因分型
Figure 1. Genotyping by locus PM-CIN4 for partial varieties of Pecan.
表 1 试验材料
Table 1. Experimental materials
编号
Code品种
Variety遗传背景
Genetic background雌雄异熟类型
Dichogamy type1 ‘Cape Fear’ ‘Schley’ 自由授粉子代 Open pollination progeny of ‘Schley’ 雄先型 Protandrous 2 ‘Choctaw’ ‘Success’ × ‘Mahan’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Mahan’ 雌先型 Protogynous 3 ‘Desirable’ ‘Russell’ × ‘Success’ 杂交子代 Hybrid progeny of ‘Russell’ × ‘Success’ 雄先型 Protandrous 4 ‘Elliott’ 实生苗 Seedling 雌先型 Protogynous 5 ‘Forkert’ ‘Success’ × ‘Schley’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Schley’ 雌先型 Protogynous 6 ‘Gloria Grande’ ‘Stuart’ 子代 Seedling of ‘Stuart’ 雌先型 Protogynous 7 ‘Hirschi’ 实生苗 Seedling 雄先型 Protandrous 8 ‘Jackson’ ‘Success’ × ‘Schley’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Schley’ 雄先型 Protandrous 9 ‘Kanza’ ‘Major’ × ‘Shoshoni’ 杂交子代 Hybrid progeny of ‘Major’ × ‘Shoshoni’ 雌先型 Protogynous 10 ‘Kiowa’ ‘Mahan’ × ‘Desirable’ 杂交子代 Hybrid progeny of ‘Mahan’ × ‘Desirable’ 雌先型 Protogynous 11 ‘Mahan’ ‘Schley’ 实生苗 Seedling of ‘Schley’ 雌先型 Protogynous 12 ‘Major’ 实生苗 Seedling 雄先型 Protandrous 13 ‘Mandan’ ‘BW-1’ × ‘Osage’ 杂交子代 Hybrid progeny of ‘BW-1’ × ‘Osage’ 雌先型 Protogynous 14 ‘Mohawk’ ‘Success’ × ‘Mahan’ 杂交子代 Hybrid progeny of ‘Success’ × ‘Mahan’ 雌先型 Protogynous 15 ‘Nacono’ ‘Cheyenne’ × ‘Sioux’ 杂交子代 Hybrid progeny of ‘Cheyenne’ × ‘Sioux’ 雌先型 Protogynous 16 ‘Navaho’ ‘Apalachee’ × ‘Wichita’ 杂交子代 Hybrid progeny of ‘Apalachee’ × ‘Wichita’ 雄先型 Protandrous 17 ‘Oconee’ ‘Schley’ × ‘Barton’ 杂交子代 Hybrid progeny of ‘Schley’ × ‘Barton’ 雄先型 Protandrous 18 ‘Pawnee’ ‘Mohawk’ × ‘Starking Hardy Giant’ 杂交子代 Hybrid progeny of ‘Mohawk’ × ‘Starking Hardy Giant’ 雄先型 Protandrous 19 ‘Schley’ 实生苗 Seedling 雌先型 Protogynous 20 ‘Shoshoni’ ‘Odom’ × ‘Evers’ 杂交子代 Hybrid progeny of ‘Odom’ × ‘Evers’ 雌先型 Protogynous 21 ‘Sumner’ ‘Schley’ 实生苗 Seedling of ‘Schley’ 雌先型 Protogynous 22 ‘Surprize’ 实生苗 Seedling 雄先型 Protandrous 23 ‘Tejas’ ‘Mahan’ × ‘Risien #1’ 杂交子代 Hybrid progeny of ‘Mahan’ × ‘Risien #1’ 雌先型 Protogynous 24 ‘Western’ ‘Longfellow’ 实生苗 Seedling of ‘Longfellow’ 雄先型 Protandrous 25 ‘Wichita’ ‘Halbert’ × ‘Mahan’ 杂交子代 Hybrid progeny of ‘Halbert’ × ‘Mahan’ 雌先型 Protogynous  下载: 导出CSV
下载: 导出CSV
表 2 10对SSR引物信息
Table 2. Details of 10 SSR primer pairs
编号
Code位点
Locus前项引物
Forward primer后项引物
Reverse primer基序
Motif参考文献
Reference1 PM-CIN4 GGCATCAGAGAAGGCTCCT CTCACCCGTCTCTAGGGCTA (CTT)12 [11] 2 PM-GA31 TGAACTCCAAAAGCCTCCTCTC GTATTTGTATTTTTTCCTTGAGCTTTCTC (CT)12 [11] 3 PM-GA38 AAAAGTTTTAGGGTTGTTTGCTCTCT GTAAAGCCTACAACCTACAACAGTCTATG (CT)12 [11] 4 PM-GA41 TCTTCAGAAAAAACCCTTACCTCTCT GAAAAATATAAACTCCCATACTACCCACAT (CT)9 [11] 5 Cc19 AAACCTTGGCATAGTCATTTGAGA GCTTTGTCAACTTTGTTTTGGGTGT (TC)8 [24] 6 Cc4 GCTCCAAGCGAAAGTCAAGT TCATAAACCAACGCCAAAGA (TC)6 [24] 7 WGA70 TGTAATTGGGGAATGTTGCA TGGGAGACACAATGATCGAA (GA)14 [25] 8 BFU-Jr19 CAGAAAGCACGTGAACCAGA AACACTCACGCACCTTCATTT (TA)10 [26] 9 BFU-Jr82 TTGCCTCTCTATAGGCTTTTTAGC TGAGTGTCTGGACAGCAAGG (CT)7 [26] 10 Zm26 TTCTTCGTCCATACCCACC ACCCCATTAGCGACCTTTA (TC)22 [27]
下载: 导出CSV
表 3 10对SSR标记多态性检测
Table 3. Polymorphism detection of the 10 pairs of SSR markers
位点
Locus等位基因数
No. of allele等位基因长度
Length of allele/bp多态信息含量
PICPM-CIN4 9 89~143 0.763 2 PM-GA31 10 84~112 0.733 4 PM-GA38 6 82~98 0.681 5 PM-GA41 4 82~88 0.437 3 Cc19 12 86~120 0.843 5 Cc4 6 185~197 0.467 7 WGA70 6 173~195 0.644 3 BFU-Jr19 3 272~290 0.343 0 BFU-Jr82 5 271~289 0.291 0 Zm26 7 201~219 0.678 5 合计 Total 68 平均 Average 6.8 0.588 3
下载: 导出CSV
表 4 25个薄壳山核桃品种的指纹图谱
Table 4. Fingerprint of 25 varieties of Pecan
品种
Varieties核心引物 Core primers PM-CIN4 PM-GA31 PM-GA41 Cc19 ‘Cape Fear’ 107/143 102 84/86 118 ‘Choctaw’ 131/143 98/100 84/86 118 ‘Desirable’ 131/143 98 84/86 108/116 ‘Elliott’ 110/143 94/102 84/86 98/118 ‘Forkert’ 107/143 102 84/86 96/98 ‘Gloria Grande’ 107 94 82/88 88 ‘Hirschi’ 104/107 94 84/86 94/96 ‘Jackson’ 92/107 94 84/86 96/98 ‘Kanza’ 107/143 94 84/86 96/118 ‘Kiowa’ 104/107 94 86 94/96 ‘Mahan’ 107/143 94 84/86 98 ‘Major’ 110/143 84/86 84/86 88/96 ‘Mandan’ 89/131 84/88 86/88 92/98 ‘Mohawk’ 92/107 94 84/86 94/98 ‘Nacono’ 92/113 94 84/86 98/118 ‘Navaho’ 107/131 94 86 96/120 ‘Oconee’ 107/143 102 84/86 116/118 ‘Pawnee’ 110/143 88 84/86 114/116 ‘Schley’ 92/107 92/94 84/86 94/98 ‘Shoshoni’ 92/107 86/88 86 86/116 ‘Sumner’ 131/143 96/98 86 108/116 ‘Surprize’ 107/128 84 84/86 96/100 ‘Tejas’ 92/143 88 86 94/96 ‘Western’ 92/107 94 86 94/98 ‘Wichita’ 110/143 112 84/86 94
下载: 导出CSV
-
[1] 彭方仁, 李永荣, 郝明灼, 等. 我国薄壳山核桃生产现状与产业化发展策略[J]. 林业科技开发, 2012, 26(4):1-4. [2] 张日清, 吕芳德, 何 方. 美国山核桃及其在我国的适应性研究[J]. 江苏林业科技, 2001, 28(4):45-47. doi: 10.3969/j.issn.1001-7380.2001.04.018 [3] 贾晓东, 王 涛, 张计育, 等. 美国山核桃的研究进展[J]. 中国农学通报, 2012, 28(4):74-78. doi: 10.3969/j.issn.1000-6850.2012.04.015 [4] 张日清, 李 江, 吕芳德. 我国引种美国山核桃历程及资源现状研究[J]. 经济林研究, 2003, 21(4):107-109. doi: 10.3969/j.issn.1003-8981.2003.04.036 [5] 张普娟, 鲍 瑾, 刘 鹏, 等. 近年我国长山核桃审(认)定情况进展[J]. 江苏农业科学, 2016, 44(8):216-219. [6] Conner P, Wood B. Identification of Pecan cultivars and their genetic relatedness as determined by randomly amplied polymorphic DNA analysis[J]. Journal of the American Society for Horticultural Science, 2001, 126: 474-480. doi: 10.21273/JASHS.126.4.474 [7] 张日清, 何 方, 吕德芳, 等. 美国山核桃群体遗传多样性的RAPD分析[J]. 经济林研究, 2001, 19(2):1-6. doi: 10.3969/j.issn.1003-8981.2001.02.001 [8] 张日清, 吕德芳, 谭晓风, 等. 美国山核桃主要栽培品种的RAPD鉴定[J]. 经济林研究, 2004, 22(4):1-5. doi: 10.3969/j.issn.1003-8981.2004.04.001 [9] 李 晖, 张 瑞, 彭方仁, 等. 美国山核桃种质资源遗传多样性ISSR分析[J]. 南京林业大学学报:自然科学版, 2015, 39(4):7-12. [10] 刘广勤, 王鹏良, 周蓓蓓, 等. 薄壳山核桃SRAP标记体系的优化和遗传多样性分析[J]. 江苏农业学报, 2010, 26(5):1037-1042. doi: 10.3969/j.issn.1000-4440.2010.05.025 [11] Grauke L J, Iqbal M J, Reddy A S, et al. Developing microsatellite DNA markers in Pecan[J]. Journal of the American Society for Horticultural Science, 2003, 128(3): 374-380. doi: 10.21273/JASHS.128.3.0374 [12] Zhang C, Yao X, Ren H, et al. Characterization and development of genomic SSRs in Pecan (Carya illinoinensis)[J]. Forests, 2020, 11(1): 61. doi: 10.3390/f11010061 [13] Nolan B, Grauke L J, Klein P. Genotyping by sequencing (GBS) and SNP marker analysis of diverse accessions of Pecan (Carya illinoinensis)[J]. Tree Genetics & Genomes, 2019, 15(1): 8. [14] 陈亚辉, 朱海军, 生静雅, 等. DNA条形码序列对不同品种美国山核桃的鉴定[J]. 江苏农业学报, 2013, 29(6):1445-1450. doi: 10.3969/j.issn.1000-4440.2013.06.042 [15] Mo Z, Lou W, Chen Y, et al. The Chloroplast genome of Carya illinoinensis: Genome structure, adaptive evolution, and phylogenetic analysis[J]. Forests, 2020, 11(2): 207. doi: 10.3390/f11020207 [16] Wang X, Rhein H S, Jenkins J, et al. Chloroplast genome sequences of Carya illinoinensis from two distinct geographic populations[J]. Tree Genetics & Genomes, 2020, 16(4): 48. [17] Kalia R K, Rai M K, Kalia S, et al. Microsatellite markers: an overview of the recent progress in plants[J]. Euphytica, 2011, 177(3): 309-334. doi: 10.1007/s10681-010-0286-9 [18] 毛秀红, 朱士利, 李善文, 等. 基于荧光SSR标记的毛白杨核心种质构建[J]. 北京林业大学学报, 2020, 42(7):40-47. doi: 10.12171/j.1000-1522.20190413 [19] 郑纪伟, 教忠意, 王保松, 等. 柳树新品种指纹图谱构建[J]. 江苏林业科技, 2020, 47(2):1-5, 56. doi: 10.3969/j.issn.1001-7380.2020.02.001 [20] 郭 斌. 栎属近缘种指纹图谱构建及遗传结构[J]. 北京林业大学学报, 2018, 40(4):10-18. [21] 毛秀红, 郑勇奇, 孙百友, 等. 基于SSR的刺槐无性系遗传多样性分析和指纹图谱构建[J]. 林业科学, 2017, 53(10):80-89. doi: 10.11707/j.1001-7488.20171009 [22] 郑纪伟, 教忠意, 窦全琴, 等. 利用荧光SSR标记构建含笑种质指纹图谱[J]. 分子植物育种, 2018, 16(14):4705-4714. [23] 段一凡, 王贤荣, 梁丽丽, 等. 桂花品种SSR荧光指纹图谱的构建[J]. 南京林业大学学报:自然科学版, 2014, 38(S):1-6. [24] Li J, Zeng Y, Shen D, et al. Development of SSR markers in Hickory (Carya cathayensis Sarg.) and their transferability to other species of Carya[J]. Current Genomics, 2014, 15(5): 357-379. doi: 10.2174/138920291505141106103734 [25] Woeste K, Burns R, Rhodes O, et al. Thirty polymorphic nuclear microsatellite loci from black walnut[J]. The Journal of Heredity, 2002, 93(1): 58-60. doi: 10.1093/jhered/93.1.58 [26] 张智勇. 核桃EST-SSR引物开发及其在喙核桃保育遗传学中的应用[D]. 北京: 北京林业大学, 2013. [27] 齐建勋, 王克建, 吴春林, 等. 核桃EST-SSR标记的开发[J]. 农业生物技术学报, 2009, 17(5):872-876. doi: 10.3969/j.issn.1674-7968.2009.05.022 [28] 王 琼, 郑勇奇, 周建仁. 分子标记在林业植物新品种鉴别中的应用及前景[J]. 林业科学, 2008, 44(6):180-182. doi: 10.3321/j.issn:1001-7488.2008.06.032 [29] Pollegioni P, Woeste K, Mugnozza G S, et al. Retrospective identification of hybridogenic walnut plants by SSR fingerprinting and parentage analysis[J]. Molecular Breeding, 2009, 24(4): 321-335. doi: 10.1007/s11032-009-9294-7 [30] Liesebach H, Schneck V, Ewald E. Clonal fingerprinting in the genus Populus L. by nuclear microsatellite loci regarding differences between sections, species and hybrids[J]. Tree Genetics Genomes, 2010, 6(2): 259-269. doi: 10.1007/s11295-009-0246-5 [31] Varshney R K, Graner A, Sorrells M E. Genic microsatellite markers in plants: features and applications[J]. Trends in Biotechnology, 2005, 23(1): 48-55. doi: 10.1016/j.tibtech.2004.11.005 [32] Gasic K, Han Y, Kertbundit S, et al. Characteristics and transferability of new apple EST-derived SSRs to other Rosaceae species[J]. Molecular Breeding, 2009, 23: 397-411. doi: 10.1007/s11032-008-9243-x [33] Yadav H K, Ranjan A, Asif M H, et al. EST derived SSR markers in Jatropha curcas L.: development, characterization, polymorphism, and transferability across the species/genera[J]. Tree Genetics & Genomes, 2011, 7(1): 207-219. [34] Zorrilla-Fontanesi Y, Cabeza A, Torres A M, et al. Development and bin mapping of strawberry genic-SSRs in diploid Fragaria and their transferability across the Rosoideae subfamily[J]. Molecular Breeding, 2011, 27(2): 137-156. doi: 10.1007/s11032-010-9417-1 [35] Savadi S B, Fakrudin B, Nadaf H L, et al. Transferability of sorghum genic microsatellite markers to peanut[J]. American Journal of Plant Science, 2012, 3(9): 1169-1180. doi: 10.4236/ajps.2012.39142 [36] Botstein D, White R L, Skolnick M, et al. Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J]. American Journal of Human Genetics, 1980, 32(3): 314-324. -

 点击查看大图
点击查看大图
图(2) / 表(4)
计量
- 文章访问数: 5176
- HTML全文浏览量: 3391
- PDF下载量: 71
- 被引次数: 0
