




-
光是植物进行光合作用和形态建成的直接影响因子,也是影响森林群落演替的主要驱动力[1]。林内光环境影响植物的生长、存活及分布,且在时间和空间上均存在较高异质性[2]。研究表明,光照影响植物的光合荧光等生理过程,进而影响植物生长发育[3-4]。然而,不同植物对光照需求存在差异,随着光照条件变化,不同耐阴性植物叶片光合特性对光环境的响应也不尽相同。在低光环境中,喜阳植物光合速率降低,而耐阴性植物仍能维持较高的光合速率。因此,明确不同植物在变化光环境下光合生理特征对植物高效培育具有重要意义。
氮和磷是植物生长所必需的大量元素,在维持生态系统物质循环和人工林生产力方面具有重要作用。氮和磷在参与蛋白质合成、细胞分裂、新生组织发育和能量转化方面有特殊功能,直接影响植物光合作用、生长发育和初级生产力的形成[5-6]。因此,植物体内氮、磷的含量及其相对比率会影响植物的光合速率和生长。研究表明,植物氮、磷元素含量与土壤养分显著相关[7],因此,土壤养分有效性影响植物氮、磷含量,从而影响植物光合速率。此外,不同生态型植物光合生理特性对氮磷养分供应的响应也存在差异。因此,研究氮磷供应比对不同植物光合生理特性的影响具有重要意义。亚热带地区氮沉降加剧,且土壤普遍缺磷,导致氮磷比例失调加剧[8],进而影响植物生长发育。适宜的氮磷供应比不仅可以改善植物的养分需求,也可节约资源成本。总之,光照和氮磷供应比显著影响植物的光合生理特性和生长发育,且不同生态型树种对光照和氮磷供应比的响应也不尽相同,但相关研究大多集中于单一因素对植物生长的影响,自然生态系统中植物生长受多种环境因素共同影响。因此,开展光照和氮磷供应比交互作用对植物光合生理特性影响的研究有助于深入认识不同植物的生态适应性。
麻栎(Quercus acutissima Carruth.)、闽楠(Phoebe bournei(Hemsl.)Yang)和木荷(Schima superba Gardn. et Champ.)广泛分布于我国亚热带地区,具有较高的生态和经济价值[9-11],其中,麻栎为典型的阳生落叶阔叶树种,而木荷和闽楠为偏阴生的常绿阔叶树种,这些树种需光性和需肥性存在一定差异;同时这3个树种也是亚热带地区营造针阔混交林的优良伴生树种。我国亚热带地区以松、杉等针叶人工林为主,现有的人工纯林普遍存在林分结构简单、生态效益不高等问题。研究表明,选择合适的树种进行混交对提高人工林生产力尤其重要。鉴于此,明确不同生态型树种对光照和氮磷供应比的响应对改善林分结构树种选择和配置意义重大。因此,选择这3个典型树种开展不同光照和施肥处理下光合生理特性与生长关系的研究,以期为亚热带地区人工混交林树种选择和配置提供理论参考。
-
本试验在杭州亚热带林业研究所虎山试验基地(30°05′ N,119°57′ E)实施。选择亚热带地区常见的麻栎、闽楠和木荷2年生实生苗为研究对象。2020年3月中旬在温室大棚内布设盆栽(花盆35 × 25 × 15 cm)实验。盆栽基质为试验地周边灌木林地采集的红黄壤(pH 4.87,有机质2.86 g·kg−1,全氮0.24 g·kg−1,全磷0.11 g·kg−1),同时添加蛭石,土壤:蛭石=4:1。施用的氮肥为硝酸铵分析纯(NH4NO3),磷肥为磷酸二氢钾分析纯(KH2PO3,其中P2O5含量约为51%)。 基于现有的一些研究结果,植物N:P<10时,植物倾向于氮限制,而N:P>20时,倾向于磷限制,二者之间不受氮磷限制[12-14];为此本研究设置5、15、和45这3个梯度来分别表征较低、中等和较高的氮磷比,分析不同氮磷供应比对植物生长影响,并将不施肥处理作为对照(CK)。同时,结合前期观测中等大小的林窗(70~100 m2)有利于林木生长,因此,本研究设置全光照和遮阴(45%全光照,采用遮阴度为50%的黑色尼龙网遮盖)来研究不同植物生理响应特征。试验采用双因素(光照和施肥)随机区组设计,光照和施肥共8个处理(LCK(全光照不施肥)、LNP5(全光照氮磷施肥比为5)、LNP15(全光照氮磷施肥比为15)、LNP45(全光照氮磷施肥比为45)、SCK(遮阴不施肥)、SNP5(遮阴氮磷施肥比为5)、SNP15(遮阴氮磷施肥比为15)、SNP45(遮阴氮磷施肥比为45)),每个处理设置6次重复,每个树种48盆苗木,3个树种共计144盆苗木。2020年4月初进行不同氮磷供应比施肥试验,根据土壤养分特征和3个树种苗木养分需求[15-16],每个处理速效态氮肥施加量为3.5 g,基本保证氮素供应处于相对丰富水平,同时添加适量磷肥,保证3个氮磷比处理水平:5(N:P=5)、15(N:P=15)和45(N:P=45)。试验期间对所有苗木进行统一管理。
-
2020年8月中旬,选择晴朗无风的天气,在上午9:00—11:00挑选冠层中上部健康成熟的新生叶,采用LI-6400便携式光合仪(LI-COR,美国)测定光响应曲线。选择红蓝光源6 cm2的标准叶室模式,叶室温度控制在25 ℃,CO2浓度控制400 μmol·mol−1。光照强度梯度设置为1 800、1 500、1 200、1 000、800、500、300、100、80、50、30、10、0 μmol·m−2·s−1,当光强梯度改变时,设置最小等待时间为120 s,每株测定1片叶子,每个处理随机挑选3株进行测定。
-
在光合作用测定的同时,使用PAM-2500便携式脉冲调制荧光仪(WALZ,德国)进行叶片叶绿素荧光特性的测定。挑选受光条件相似、长势趋于一致且健康的相同叶位处的叶片,测定前先利用暗适应叶夹对所测叶片进行30 min充分暗适应,经照射检测光测得初始荧光(F0),最大荧光(Fm)在强度为1 000 μmol·m−2·s−1的饱和脉冲下测得,脉冲时间为0.8 s,可根据公式计算PSII的最大光合量子产量(FV/Fm=(Fm−F0)/Fm),每个处理组随机选取3株植株进行检测,每株测定3片叶子。其余参数:PSⅡ实际光合量子产量Y(II)、调节性能量耗散的量子产量Y(NPQ)、非调节性能量耗散的量子产量Y(NO)、电子传递速率(ETR)和光化学猝灭系数(qP)均由仪器测定后得到。测定过程中活化光光强为1 000 μmol·m−2·s−1,共测定180 s。ETR=Y(II)× PAR × 0.5 × 0.84,其中0.84是植物的经验性吸光系数,0.5是假设吸收的光能被PSⅠ和PSⅡ平均分配[17]。
-
光合荧光参数测定完成后,每个处理随机选择3株苗木,选取受光一致,生长健康的成熟叶用于光合色素含量测定,采用乙醇丙酮提取法[18]测定3个树种的光合色素含量,包括叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)。
-
分别用卷尺和游标卡尺测定每株苗木苗高和地径。苗高(RGR-H)和地径(RGR-B)相对增长率计算公式分别为:RGR-H=(㏑H1−㏑H0)/(t1−t0),RGR-B=(㏑B1−㏑B0)/(t1−t0),式中H0和B0分别是施肥前的苗高和地径,H1和B1分别是施肥处理7个月后的苗高和地径,t0为2020年4月1日,t1为2020年10月25日,t1−t0为施肥前后时间间隔。
-
光照和氮磷供应比对不同树种的光合生理参数影响进行双因素方差分析(Two-Way ANOVA),同一光照和不同氮磷供应比下不同树种的光合生理性状的差异采用单因素方差分析(One-Way ANOVA)和LSD多重比较方法分析。生长率与光合生理特性参数的相关性采用主成分分析(PCA)。光响应曲线的拟合采用叶子飘的光合分析软件直角双曲线模型拟合[19]。本研究的数据统计分析均在SPSS22中进行,图表均由Origin绘制,表中数据为平均值±标准差(Mean±SD)。
-
经双因素方差分析(附表1),光照、施肥及其交互作用极显著影响麻栎光饱和点(LSP)和最大净光合速率(Pnmax);施肥显著或极显著影响闽楠LSP、Pnmax、光补偿点(LCP)和暗呼吸速率(Rd);光照、施肥及其交互作用显著或极显著影响木荷Pnmax、LCP及Rd;其中遮阴显著降低了麻栎的LSP和Pnmax,增加了木荷的Pnmax,而对闽楠Pnmax没有显著影响,这表明3个树种对光照和施肥处理的光合响应具有较大差异。
树种 Species 因素 Factor df Chla Chlb Chla/b Chla + b Car Y(II) Y(NPQ) Y(NO) qP 麻栎
Q. acutissima光照
light1 801.43** 504.28** 19.79** 753.11** 738.86** 4.09 395.89** 82.95** 20.54** 施肥
Fertilization3 604.60** 296.19** 0.18 530.90** 595.70** 60.38** 81.48** 19.31** 74.28** 光照 × 施肥
Light × Fertilization7 86.22** 62.88** 4.45** 84.50** 73.85** 16.05** 86.85** 1.42** 24.21** 闽楠
P. bournei光照
light1 2 024.56** 1 872.50** 9.50** 2 140.89** 434.38** 2.69 1 176.90** 1 272.00** 108.80** 施肥
Fertilization3 137.61** 104.56** 0.15 138.15** 104.19** 25.20** 302.16** 230.45** 19.38** 光照 × 施肥
Light × Fertilization7 265.88** 217.90** 0.50 272.24** 136.60** 286.11** 670.71** 81.58** 262.40** 木荷
S. superba光照
light1 1 758.04** 1 871.52** 252.79** 1 869.68** 1 148.82** 1.31 616.44** 460.04** 29.57** 施肥
Fertilization3 888.07** 858.84** 143.42** 920.57** 731.98** 54.97** 384.15** 54.61** 28.62** 光照 × 施肥
Light × Fertilization7 275.53** 224.51** 158.86** 272.60** 252.05** 128.83** 483.14** 85.67** 132.52** 树种 Species 因素 Factor df ETR Fv/Fm AQY LSP Pnmax LCP Rd RGR-H RGR-B 麻栎
Q. acutissima光照
light1 4.09 9.50** 0.05 40.69** 41.01** 3.30 2.96 11.56** 4.07 施肥
Fertilization3 60.38** 2.43 1.77 12.05** 54.04** 1.69 1.27 11.84** 9.16** 光照 × 施肥
Light × Fertilization7 16.05** 0.58 11.21** 9.31** 17.13** 3.75* 2.92 0.92 0.75 闽楠
P. bournei光照
light1 2.69 5.42* 9.65** 4.43 2.67 21.31** 31.83** 3.70 4.22 施肥
Fertilization3 25.20** 2.66 0.33 6.91** 4.98* 4.00* 5.31** 57.28** 2.54 光照 × 施肥
Light × Fertilization7 286.11** 6.74** 3.07 3.76* 10.19** 1.81 3.17 8.68** 2.97 木荷
S. superba光照
light1 1.31 8.42** 2.81 1.92 6.35* 32.83** 53.22** 1.30 12.85** 施肥
Fertilization3 54.97** 4.25* 14.70** 6.04** 15.53** 12.79** 10.49** 5.26** 2.14 光照 × 施肥
Light × Fertilization7 128.83** 3.52* 2.05 1.41 5.69** 10.91** 5.69** 0.96 4.50* 注:*和**分别表示5%、1%水平显著差异
Notes:* and ** represent significant differences in levels of 5% and 1%Table 1. Schedule 1 Effects of light and N:P supply ratio on photosynthetic parameters of three tree species (F)
由表1可知:在2种光照处理下施肥对麻栎和闽楠表观量子效率(AQY)及Rd影响较小,但施肥显著降低了木荷AQY(LNP45除外,P<0.05),而显著增加了Rd(LNP5除外,P<0.05),说明施肥显著降低了木荷的光量子效率,增强了呼吸消耗。此外,在2种光照条件下施肥均显著增加麻栎Pnmax(SNP15除外,P<0.05),但对木荷Pnmax影响较小(Pnmax仅在SNP15时显著高于CK),而闽楠则在全光照处理下Pnmax显著高于CK(P<0.05),表明施肥对不同植物潜在光合能力的影响存在差异。在2种光照处理下,施肥均增加了闽楠和木荷的LSP和LCP,说明施肥增强了闽楠和木荷对光的利用能力。在全光照处理下,施肥显著降低了麻栎LSP(P<0.05),增加了LCP(P<0.05),说明施肥削弱了麻栎对强光的利用能力;在遮阴处理下,不同氮磷供应比对麻栎LSP的影响不同,而LCP均有不同程度的降低,说明施肥提高了麻栎对弱光的利用能力。
树种
Species处理
Treatments表观量子效率
AQY/(μmol·m−2·s−1)光饱和点
LSP/(μmol·m−2·s−1)最大净光合速率
Pnmax/(μmol·m−2·s−1)光补偿点
LCP/(μmol·m−2·s−1)暗呼吸速率
Rd/(μmol·m−2·s−1)麻栎
Q. acutissimaSCK 0.05±0.021 ab 835.60±110.61 c 3.71±0.86 d 12.46±5.94 ab 0.51±0.12 b SNP5 0.07±0.004 a 1 086.45±56.28 bc 8.58±0.71 b 8.56±2.28 b 0.54±0.14 b SNP15 0.04±0.007 b 637.30±62.05 d 5.24±0.34 cd 12.40±0.12 ab 0.45±0.08 b SNP45 0.06±0.004 ab 998.73±160.67 bc 6.36±1.41 c 8.49±4.81 b 0.46±0.28 b LCK 0.07±0.001 a 1 705.95±14.11 a 4.51±0.19 d 8.58±2.87 b 0.52±0.17 b LNP5 0.02±0.011 b 1 283.79±248.59 b 8.01±0.69 b 16.56±1.37 a 0.41±0.20 b LNP15 0.07±0.017 a 987.25±183.61 c 7.45±0.52 bc 14.43±1.47 a 0.88±0.22 a LNP45 0.06±0.007 a 1 060.61±143.55 bc 11.50±0.19 a 11.46±0.25 ab 0.65±0.08 ab 闽楠
P. bourneiSCK 0.06±0.008 b 722.67±142.06 b 4.45±0.63 bc 5.69±3.96 b 0.35±0.26 c SNP5 0.07±0.007 ab 973.74±56.86 b 5.78±0.56 ab 9.25±1.59 b 0.66±0.11 bc SNP15 0.06±0.013 b 826.74±254.65 b 3.58±0.22 c 7.61±1.99 b 0.46±0.20 c SNP45 0.07±0.001 b 899.31±177.79 b 5.13±0.81 b 5.87±1.78 b 0.40±0.11 c LCK 0.09±0.014 ab 666.40±165.22 c 3.47±0.86 c 7.52±0.99 b 0.56±0.11 c LNP5 0.07±0.005 ab 1 117.74±191.49 ab 5.29±1.24 b 12.22±0.61 ab 0.83±0.05 bc LNP15 0.09±0.006 a 1 336.65±178.85 a 7.08±0.36 a 14.78±3.83 a 1.15±0.19 a LNP45 0.07±0.009 ab 868.75±41.58 b 5.19±1.02 b 12.99±3.14 ab 0.84±0.17 b 木荷
S. superbaSCK 0.07±0.006 a 495.97±2.51 d 5.32±0.36 b 4.07±2.52 d 0.28±0.19 d SNP5 0.04±0.010 c 541.64±139.45 d 4.48±0.49 b 17.96±4.94 bc 0.65±0.01 c SNP15 0.06±0.004 b 1 663.25±161.59 b 7.48±0.31 a 10.97±3.16 c 0.60±0.12 c SNP45 0.05±0.000 b 1 415.29±1 027.62 a 4.39±0.02 b 13.36±0.00 c 0.63±0.00 c LCK 0.08±0.005 a 593.42±123.60 d 4.47±0.48 b 12.89±1.84 c 0.84±0.07 c LNP5 0.06±0.005 b 621.44±119.77 d 4.64±0.42 b 15.43±6.47 bc 0.77±0.33 c LNP15 0.05±0.006 b 986.34±260.10 c 5.44±1.22 b 35.60±2.91 a 1.59±0.21 a LNP45 0.07±0.006 a 797.94±288.29 cd 4.71±0.45 b 20.59±6.55 b 1.23±0.23 b 注:不同小写字母表示不同处理间差异显著(P<0.05)。SCK, SNP5, SNP15 和SNP45分别表示遮阴下不施肥、氮磷比为5,15和45的处理;LCK,LNP5,LNP15和LNP45分别表示全光照下不施肥、氮磷比为5,15和45的处理。下表同 Notes: Different low letters indicate significant difference between different treatments (P<0.05). SCK, SNP5, SNP15 and SNP45 represent treatments with no fertilization under shading and N:P ratios of 5, 15 and 45; LCK, LNP5, LNP15 and LNP45 represent treatments without fertilization under full light and N:P ratios of 5, 15 and 45. The same as in the following table Table 1. Changes of different light and fertilization in photosynthetic parameters of three tree species.
-
经双因素方差分析(附表1),光照、施肥及其交互作用均极显著影响麻栎、闽楠和木荷Y(NPQ)、Y(NO)及qP,而Y(II)和ETR受施肥极显著影响;其中遮阴显著降低了麻栎、闽楠及木荷Y(NPQ)和qP,显著增加了Y(NO),表明遮阴降低了植物用于光化学反应的能量和非光化学耗散的能量,表现出植物对光合机构的自我保护机制。
由表2可知:在全光照处理下,与不施肥处理CK相比,施肥显著增加了闽楠和木荷Y(II)、qP(闽楠LNP45除外)及ETR( P <0.05),而麻栎 Y (II)、 qP 及ETR随氮磷供应比增加呈现出增—减—增的趋势。可见,总体上施肥均增加了植物用于光化学反应的能量和植物PSII反应中心的电子流动速率。此外,施肥显著降低了麻栎、闽楠及木荷的Y(NPQ),显著增加了Y(NO)(P<0.05),说明施肥降低了植物用于跨膜质子梯度相关的非辐射热耗散,增加了非调节性能量耗散的量子产量。
树种
Species处理
treatmentPSⅡ的实际光合
量子产量 Y(II)调节性能量耗散的
量子产量 Y (NPQ)非调节性能量耗散的
量子产量 Y (NO)光化学猝灭
系数 qP电子传递速率
ETRPSⅡ的最大光合
量子产量 Fv/Fm麻栎
Q. acutissimaSCK 0.26±0.024 b 0.32±0.017 e 0.42±0.059 b 0.43±0.061 b 107.10±17.46 b 0.70±0.038 b SNP5 0.30±0.012 a 0.30±0.012 f 0.39±0.010 bc 0.48±0.018 a 127.00±8.57 a 0.74±0.014 ab SNP15 0.13±0.004 e 0.38±0.004 c 0.48±0.004 a 0.21±0.010 d 55.44±2.62 e 0.72±0.020 b SNP45 0.22±0.003 c 0.33±0.000 e 0.45±0.004 ab 0.35±0.008 c 91.98±1.92 c 0.71±0.019 b LCK 0.21±0.003 cd 0.46±0.003 a 0.32±0.003 d 0.37±0.010 c 90.02±2.31 cd 0.75±0.009 ab LNP5 0.28±0.007 ab 0.38±0.007 c 0.33±0.005 c 0.47±0.020 ab 115.64±4.83 ab 0.76±0.011 a LNP15 0.19±0.003 d 0.40±0.002 b 0.41±0.003 b 0.33±0.009 c 77.84±1.89 d 0.74±0.022 ab LNP45 0.29±0.004 a 0.34±0.004 d 0.36±0.004 c 0.49±0.012 a 122.64±2.94 a 0.73±0.011 ab 闽楠
P. bourneiSCK 0.17±0.000 e 0.37±0.000 c 0.46±0.002 a 0.28±0.002 f 70.42±0.24 e 0.72±0.003 ab SNP5 0.28±0.017 b 0.33±0.010 d 0.39±0.010 d 0.47±0.019 b 117.60±6.94 b 0.74±0.011 a SNP15 0.18±0.006 d 0.40±0.003 b 0.42±0.008 b 0.31±0.012 e 77.28±2.62 a 0.73±0.012 a SNP45 0.25±0.012 c 0.32±0.006 e 0.42±0.006 b 0.41±0.020 c 106.82±5.05 c 0.71±0.020 ab LCK 0.14±0.005 f 0.56±0.002 a 0.30±0.003 g 0.34±0.013 d 59.64±2.10 f 0.69±0.009 b LNP5 0.25±0.006 c 0.39±0.003 b 0.35±0.003 e 0.48±0.011 b 106.68±2.34 c 0.70±0.019 b LNP15 0.32±0.010 a 0.37±0.007 c 0.32±0.003 f 0.56±0.016 a 133.00±3.99 a 0.73±0.013 a LNP45 0.20±0.008 d 0.40±0.003 b 0.40±0.005 c 0.35±0.014 d 83.02±3.15 d 0.73±0.010 a 木荷
S. superbaSCK 0.23±0.038 c 0.35±0.011 e 0.42±0.032 b 0.39±0.059 c 95.20±15.85 c 0.72±0.012 c SNP5 0.16±0.003 d 0.37±0.000 d 0.47±0.003 a 0.24±0.005 d 65.24±1.06 d 0.75±0.009 b SNP15 0.27±0.007 b 0.35±0.018 e 0.38±0.011 c 0.45±0.011 b 114.52±2.80 b 0.75±0.009 b SNP45 0.26±0.008 bc 0.37±0.004 d 0.37±0.004 cd 0.39±0.013 c 107.94±3.36 bc 0.74±0.014 b LCK 0.10±0.003 e 0.70±0.002 a 0.20±0.002 f 0.26±0.009 d 41.44±1.47 e 0.75±0.017 b LNP5 0.24±0.003 c 0.44±0.002 b 0.32±0.002 e 0.42±0.006 bc 99.54±1.26 c 0.75±0.004 ab LNP15 0.36±0.015 a 0.31±0.018 f 0.33±0.003 e 0.61±0.017 a 151.06±6.09 a 0.75±0.017 b LNP45 0.24±0.009 c 0.40±0.004 c 0.36±0.004 d 0.39±0.014 c 102.76±3.57 c 0.78±0.014 a Table 2. Effect of different light and N:P supply ratio on chlorophyll fluorescence parameters of the three species
在遮阴处理下,与CK相比,施肥显著增加了闽楠Y(II)、qP及ETR(P<0.05),麻栎Y(II)、qP和ETR随氮磷供应比增大呈先增加后降低趋势(仅氮磷供应比为5时增加),而木荷则呈先降低后增加的趋势(仅氮磷供应比为5时降低),表明氮磷供应比对不同植物的光能利用效率和电子传递活性的影响存在差异。此外,氮磷供应比为15显著增加了麻栎和闽楠Y(NPQ),但氮磷供应比为15对木荷Y(NPQ)无显著影响。另外,麻栎和闽楠的Y(NO)随氮磷供应比增加呈先降低后增加的趋势(仅氮磷供应比为5时降低),而木荷的Y(NO)随氮磷供应比增加呈先增加后降低的趋势(仅氮磷供应比为5时增加)。这说明不同植物对光能利用和光合机构保护调节机制存在差异。
-
经双因素方差分析(附表1),光照、施肥及其交互作用均显著影响麻栎、闽楠和木荷Chla、Chlb、Chla + b、Car;麻栎和闽楠Chla/b不受施肥显著影响,而木荷Chla/b受光照、施肥及其交互作用的极显著影响;其中遮阴显著增加了麻栎、闽楠和木荷Chla、Chlb、Chla + b及Car(P<0.05),3个树种的Chla/b值总体上均为遮阴处理小于全光照处理,表明遮阴提高了3个树种对光能的捕获能力。
由表3可知:在2种光照处理下,施肥显著增加了麻栎和木荷Chla、Chlb、Chla+b及Car(P<0.05)。在全光照处理下,麻栎和木荷Chla、Chlb、Chla+b及Car在N:P为45时最高;在遮阴处理下麻栎Chla、Chlb、Chla+b及Car在N:P为15时最高,而木荷在N:P为5时最高。在全光照处理下,施肥显著增加了闽楠Chla、Chlb、Chla+b和Car(P<0.05);但在遮阴处理下,在氮磷供应比为45时闽楠Chla、Chlb和Chla+b与对照没有显著差异(P>0.05),而N:P为5和15时显著低于对照处理(P<0.05)。由此可知,2种光照条件下施肥均有利于提高麻栎和木荷的光合色素含量,增加植物对光能的捕获能力;而闽楠仅全光照条件下施肥提高了光合色素含量。
树种
Species处理
Treatments叶绿素 a
Chla/
(mg.g−1)叶绿素 b
Chlb/
(mg.g−1)叶绿素 a/b
Chla/b叶绿素 a + b
Chla + b/
(mg.g−1)类胡萝卜素
Car/
(mg.g−1)麻栎
Q. acutissimaSCK 3.188±0.002 f 1.10±0.045 f 2.90±0.120 b 4.29±0.043 f 0.60±0.016 f SNP5 7.944±0.414 b 2.89±0.270 b 2.76±0.115 b 10.83±0.684 b 1.35±0.048 b SNP15 9.371±0.250 a 3.37±0.004 a 2.78±0.078 b 12.74±0.246 a 1.63±0.052 a SNP45 6.720±0.031 c 2.31±0.033 c 2.91±0.055 b 9.03±0.002 c 1.16±0.021 c LCK 1.617±0.048 g 0.55±0.018 g 2.96±0.182 ab 2.16±0.030 g 0.33±0.018 g LNP5 4.705±0.114 e 1.53±0.040 e 3.07±0.006 ab 6.24±0.155 e 0.91±0.024 e LNP15 5.025±0.153 e 1.62±0.021 e 3.10±0.055 a 6.65±0.174 e 0.90±0.034 e LNP45 5.979±0.304 d 2.05±0.113 d 2.91±0.012 b 8.03±0.417 d 1.01±0.048 d 闽楠
P. bourneiSCK 5.97±0.206 a 2.08±0.034 a 2.87±0.053 b 8.06±0.240 a 1.08±0.076 b SNP5 4.85±0.144 c 1.67±0.051 c 2.91±0.004 ab 6.52±0.195 c 0.86±0.020 de SNP15 5.40±0.185 b 1.85±0.036 b 2.92±0.043 ab 7.25±0.221 b 1.02±0.053 c SNP45 6.12±0.075a 2.16±0.066 a 2.84±0.052 b 8.27±0.141 a 1.16±0.011 a LCK 1.22±0.081 e 0.40±0.063 e 3.06±0.284 a 1.62±0.145 e 0.28±0.002 f LNP5 3.84±0.024 d 1.29±0.030 d 2.98±0.049 ab 5.14±0.054 d 0.83±0.003 e LNP15 4.05±0.126 d 1.34±0.041 d 3.02±0.001 ab 5.39±0.168 d 0.89±0.030 d LNP45 3.88±0.058 d 1.28±0.056 d 3.03±0.086 ab 5.16±0.113 d 0.89±0.022 d 木荷
S. superbaSCK 3.69±0.102 ef 1.27±0.010 d 2.90±0.103 c 4.96±0.092 e 0.61±0.021 e SNP5 6.85±0.032 a 2.39±0.010 a 2.87±0.001 c 9.24±0.042 a 1.14±0.006 a SNP15 6.18±0.256 b 2.03±0.081 b 3.05±0.004 b 8.21±0.337 b 1.05±0.055 b SNP45 4.72±0.071 d 1.69±0.018 c 2.80±0.013 d 6.41±0.089 d 0.75±0.015 d LCK 1.50±0.033 g 0.38±0.007 e 3.94±0.060 a 1.88±0.040 f 0.17±0.009 f LNP5 3.85±0.088 e 1.28±0.055 d 3.00±0.061 bc 5.13±0.144 e 0.66±0.018 e LNP15 3.63±0.152 f 1.24±0.054 d 2.92±0.004 c 4.88±0.206 e 0.63±0.027 e LNP45 5.08±0.051 c 1.68±0.036 c 3.02±0.035 b 6.77±0.088 c 0.87±0.003 c Table 3. Effects of light and N:P supply ratio on photosynthetic pigments
-
经双因素方差分析,施肥极显著影响麻栎、闽楠、木荷苗高相对增长率以及麻栎地径相对增长率,光照显著影响麻栎苗高相对增长率和木荷地径相对增长率(附表1);其中遮阴显著增加了麻栎苗高相对增长率和木荷地径相对增长率,这可能是植物逃避低光环境的一种生态适应策略。
在全光照处理中,施肥显著增加了3个树种苗高相对增长率(图1A~C,P<0.05)和麻栎地径相对增长率(图1D,P<0.05),而对闽楠和木荷地径相对增长率影响较小(除闽楠氮磷供应比45外),说明施肥对麻栎的促生效应强于闽楠和木荷;在遮阴处理中,施肥显著增加闽楠苗高相对增长率(图1B,P<0.05),但对闽楠地径、木荷苗高和木荷地径相对增长率无显著影响(除木荷氮磷供应比15外),另外,氮磷供应比为5和15 显著增加麻栎苗高相对增长率(图1A,P<0.05),氮磷供应比为5和45显著增加麻栎地径相对增长率(图1D,P<0.05),表明施肥对木荷的促生效应相对较弱。
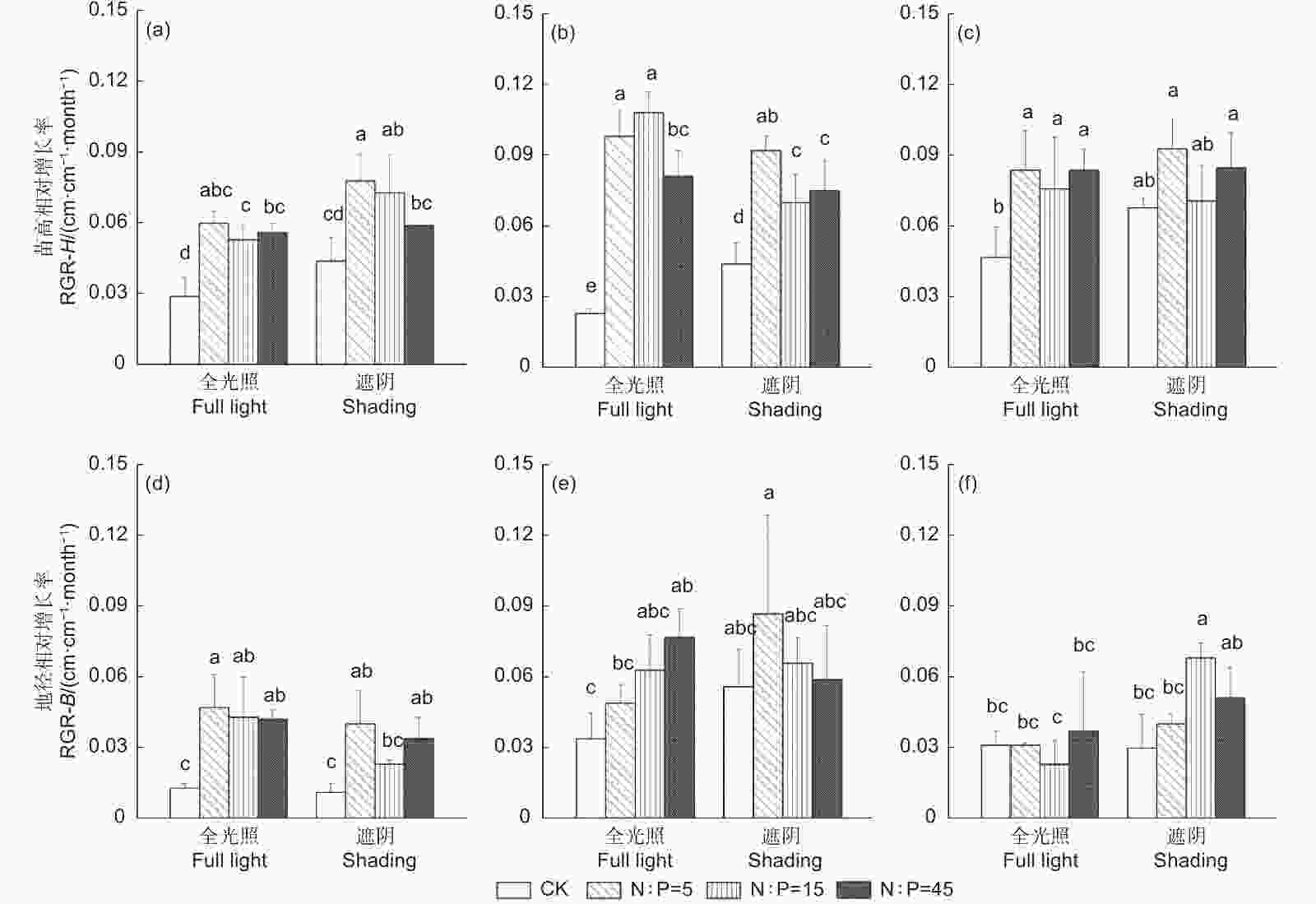
Figure 1. Relative growth rate of seedling height and ground diameter of three tree species
-
主成分分析前2轴分别解释了麻栎(图2A)、闽楠(图2B)和木荷(图2C)光合生理特征变异的74.58%、84.03%和83.51%。麻栎苗高相对增长率与Chla、Car相关性紧密,而地径相对增长率与Pnmax相关性更为紧密,说明光合色素含量可能是影响麻栎苗高生长的关键光合生理指标,而地径生长受潜在光合能力影响。闽楠苗高和地径相对增长率均与Y(II)、ETR和Pnmax相关性紧密,表明叶绿素荧光参数对闽楠生长的影响更加明显。木荷苗高相对增长率与Chla、Chlb相关性紧密,地径相对增长率与Pnmax和LSP相关性更为紧密,且主要分布于遮阴处理,说明光合色素含量及其光合光响应参数可能是影响木荷生长的重要指标。
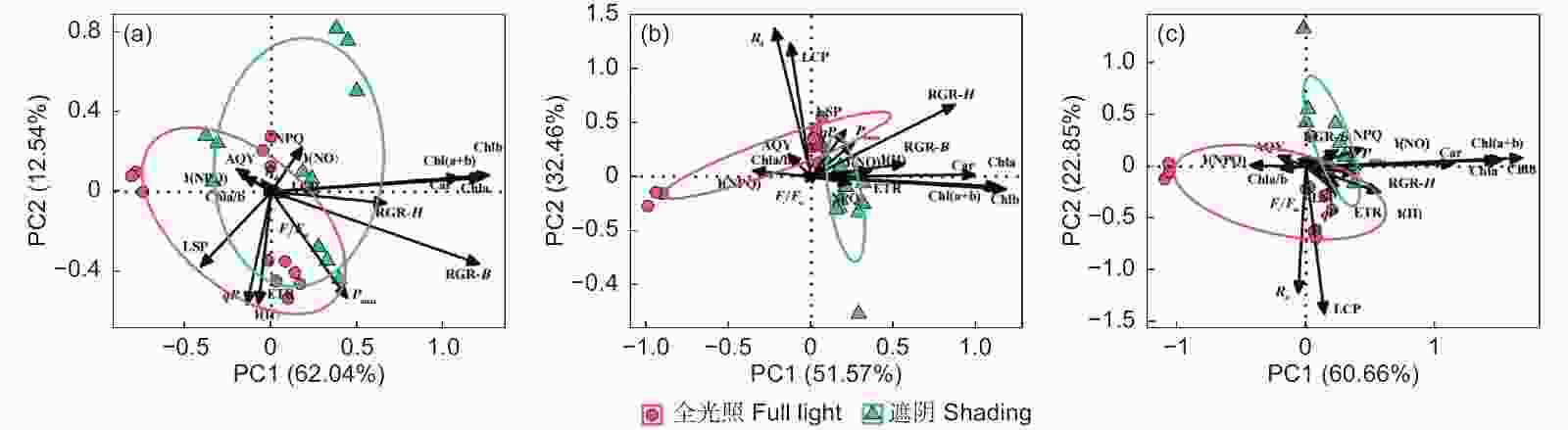
Figure 2. Principal component analysis of growth rate and photosynthetic characteristics
-
本研究中,麻栎、闽楠和木荷对不同光照的光合响应特征存在较大差异。遮阴降低了麻栎的Pnmax,增加了木荷的Pnmax,而闽楠Pnmax没有显著变化。麻栎是典型的喜阳树种,光照强度显著降低是导致其光合能力下降的主要原因[1];而木荷偏阴生,幼龄期耐阴性较强,在中等光照条件下光合能力增加,表现出极强的后期演替树种特点[1]。闽楠光合能力较稳定,这可能主要与闽楠属中性植物具有较宽的光生态幅有关。整体而言,施肥增加了3个树种的Pnmax,其中,麻栎增幅最高、闽楠次之、木荷较低。因此,贫瘠土壤增加氮磷养分供应,可以促进植物光合代谢。但施肥对不同树种光合速率的影响存在差异,这可能与植物的需肥性有关。麻栎为速生先锋树种,需肥性较强,对氮磷养分供应的敏感性较高;而木荷属于后期演替树种,前期对养分供应不敏感;闽楠则处于中间类型。此外,在全光照处理下,施肥显著增加了麻栎和闽楠Pnmax,但对木荷Pnmax无显著影响,其中,麻栎Pnmax在氮磷供应比为45时最高,闽楠Pnmax在氮磷供应比为15时最高。这说明光照充足环境下,麻栎对氮肥需求远高于磷肥,氮磷均衡施肥则促进了闽楠光合潜力的提升。然而,木荷光合能力对氮磷施肥的响应不敏感,可能受木荷自身的喜阴特性有关,即施肥对木荷的影响依赖于特定的光照环境。本研究发现,在遮阴处理下,氮磷供应比为15显著增加了木荷Pnmax,而氮磷供应比为5显著增加了麻栎Pnmax,说明麻栎在弱光环境中需要更高比例的磷肥供应来维持较高的光合能力。与全光照下木荷Pnmax的变化趋势类似,施肥对遮阴环境下闽楠Pnmax无显著影响。上述结果表明,氮磷供应比对3个树种光合能力的提高均表现出一定的光依赖性,但常绿阔叶树种(木荷和闽楠)对施肥的响应受光照条件的调控弱于落叶阔叶树种麻栎。由此可知,不同植物在不同光照条件下适宜的氮磷供应比对提高植物光合能力具有重要意义,但相关机制还需要进一步研究。 LSP和LCP的高低直接反映植物叶片对强光和弱光的利用能力[20]。本研究中,遮阴显著降低了麻栎LSP,但对闽楠和木荷LSP影响较小。说明遮阴降低了麻栎对光的利用能力,而对闽楠和木荷利用光合能力影响较小。此外,在2种光照处理下,施肥均增加了闽楠和木荷LSP和LCP,尤其是LSP增幅较大,表明施肥有助于提高闽楠和木荷光能利用率。
在全光照处理中,施肥显著增加了闽楠和木荷的Y(II)和ETR,表明施肥提高了闽楠和木荷的电子传递活性和光能转化效率,从而促进了PSII的活性[21];麻栎Y(II)、qP和ETR在氮磷供应比为45时显著高于CK,说明高氮磷供应比有利于提高麻栎的光合利用效率,这可能主要与氮添加增加植物叶片氮含量,植物叶片氮含量显著影响植物光合速率有关[22]。同时,植物光系统活性可能与氮磷供应比大小有关,在一定阈值范围内,氮磷供应比施肥有利于植物光系统活性的提高[23]。在遮阴处理中,麻栎和闽楠Y(II)、qP与ETR均在氮磷供应比为5最高,木荷Y(II)、qP和ETR在氮磷供应比为15时最高,说明麻栎和闽楠在较低土壤氮磷比时PSII的活性较强,木荷在中等氮磷比时PSII的活性较强,这可能是植物响应氮磷养分限制保护PSII免受伤害的一种保护机制[24]。同时,这些光合荧光参数对光照和施肥的响应特性进一步证实了上述Pnmax研究结果。Fv/Fm表示PSII反应中心光合原初反应能量转化效率,植物在不受胁迫影响的情况下,Fv/Fm一般介于0.75~0.85[25]。本试验中,闽楠所有处理Fv/Fm均低于0.75,与施福军等[26]的研究结论一致。这可能是因为植物幼苗的光合机构还处在发育过程中,对环境因子变化的响应还不够稳定所致[27]。也可能是植物发生的一种可逆减量调节,即植物为了保护光合机构而产生的一种保护措施[28]。
光合色素对植物的光合作用具有重要作用。本研究中,与全光照处理相比,遮阴显著增加了麻栎、闽楠和木荷Chla、Chlb、Chla + b及Car,但降低了Chla/b。这可能是植物适应弱光环境的生存策略,即在遮阴环境下植物会增加光合色素含量(尤其Chlb),进而增强植物对光能的捕获效率[29]。而光合色素含量的升高很可能与遮阴降低光氧化伤害,叶绿体内基粒片层垛叠程度变高相关[30]。研究表明,氮磷养分影响植物的光合作用,氮磷养分的缺乏均会降低植物的光合速率[31]。在2种光照处理下,氮磷供应比施肥均显著增加了麻栎、木荷和闽楠(除遮阴处理外)的Chla、Chlb、Chla + b及Car,说明氮磷养分对麻栎、木荷和闽楠光合作用的影响较一致,即通过增加光合色素含量来增强植物叶片对光能捕获,进而增强植物光合速率。但在遮阴处理中,与不施肥处理相比,闽楠的光合色素变化随氮磷供应比增加呈先降低后增加的趋势,这与Pnmax变化趋势不一致,可能与光合作用相关的酶及其辅酶有关[32]。因此,在未来的研究中需要进一步研究生理生化指标来揭示不同树种光合能力的差异。
遮阴显著增加了麻栎苗高相对增长率和木荷地径相对增长率,而对闽楠生长影响较小。Poorter[33]研究表明,遮阴会增加植物高生长,以使植物有利于获得更多的光能进行光合作用。此外,杨莹[34]研究表明,喜阳树种具有更高的生长潜力,而耐阴性较强树种拥有更高的低光碳同化率和碳捕获能力,这也部分支持了本研究中麻栎(喜阳植物)和木荷(阴性植物)研究结果。遮阴提高了木荷生长,可能与遮阴促进光合作用(平均Pnmax比全光照增加12%),降低光补偿点和增加叶绿素含量获得更多光能有关。薛黎[35]研究也发现,30%和50%遮光率对闽楠株高和地径生长无显著影响。这可能主要归因于闽楠为中性树种,对一定范围的光强具有相对稳定的适应性。综合看,在2种光照处理下,氮磷供应有利于促进植物的生长。这种促进作用很可能主要归因于施肥增加植物氮、磷含量,进而促进植物光合色素的合成[23]。但施肥对林木促生效应的影响与光照环境和树种有关。在全光照处理中,施肥对麻栎的促生效应最强;在遮阴处理中,施肥对木荷的促生效应最弱。这与上述施肥对3个树种Pnmax的影响类似。本研究中,麻栎苗高相对增长率与Chla、Car相关性紧密,而地径相对增长率与Pnmax相关性更紧密,说明麻栎苗高生长主要与叶片对光能的捕获相关,这可能是因为光合色素含量直接影响植物对光能的捕获,从而促进了植物生长。而地径生长主要与潜在光合能力有关,这很可能主要归因于光合电子传递耦联形成的跨膜质子梯度驱动叶绿体ATP合酶生成ATP[36]。闽楠苗高和地径相对增长率均与Y(II)和ETR相关性紧密,表明闽楠生长可能主要取决于PSII的实际光合效率及电子传递速率[21]。木荷苗高相对增长率与Chla、Chlb相关性紧密,地径相对增长率与Pnmax和LSP相关性更紧密,且主要分布于遮阴处理,说明木荷生长与光合利用效率和潜在光合能力相关。这可能是木荷生长对光照的响应比施肥更敏感的原因。
-
麻栎、闽楠和木荷光合特性及生长受光照和氮磷供应比协同影响。遮阴降低了麻栎Pnmax,增加了木荷Pnmax,但对闽楠Pnmax无显著影响;同时氮磷供应比对3个树种光合能力的提高均表现出一定的“光”依赖性,其中,常绿阔叶树种(木荷和闽楠)对施肥的响应受光照条件的调控弱于落叶阔叶树种麻栎。
Effects of Light and N:P Ratio on Photosynthetic Characteristics in Seedlings of Three Typical Tree Species
- Received Date: 2022-04-28
- Accepted Date: 2022-06-26
- Available Online: 2023-02-20
Abstract:
 DownLoad:
DownLoad: