



-
气候变化导致全球水分循环的改变,重塑了大部分地区的土壤-植被-大气连续体,增加了干旱发生的频率和持续时间[1]。目前,干旱已成为多数地区植物主要遭受的环境胁迫[2],而研究干旱对植物的作用机理,制定高效节水的灌溉策略成为当前人们的重要议题。
辽东栎(Quercus liaotungensis Koidz.)是我国华北中低山地带针阔叶混交林的建群树种[3]。随着气候变化加剧,土壤干旱成为辽东栎,尤其是辽东栎幼树生长的主要限制因素之一[4]。光合作用对干旱胁迫有明显的响应,其可以引发气孔限制(SL)或非气孔限制(NSL),亦或使2种限制的叠加出现,植物表现出多种干旱响应,例如:在光合系统的结构方面,气孔[5]、叶片表型特征[6]及叶绿体等超微结构[7]发生变化;在光合系统与环境的物质交换方面,气体交换参数[8]和叶片含水量发生变化[9];在光合系统的能量流动方面,用于抗氧化防御和非光化学猝灭的能量改变,光能利用效率随之变动[10]。目前,干旱胁迫对辽东栎幼树的表型生理特性、叶片气体交换、水分利用效率、木质部液流的影响已有研究报道[11-13],但对光合系统的影响机制仍然不清晰。
由于植物光合系统各组分对土壤干旱的敏感性不同[14],可以利用敏感性指标判断干旱程度。目前已有通过监测植物水势、叶绿素含量[15]、叶片反射光谱[16],叶片颜色信息[17]、叶温[18]、叶倾角[19]和叶片叶绿素荧光[20]判断干旱胁迫程度的研究。现有研究证实,叶倾角、叶片叶绿素荧光和光合气体交换参数均与植株含水量关联密切[21-23],但干旱胁迫下叶倾角与光合系统各组分之间的关联尚不清晰。鉴于此,本研究采用盆栽渐进式干旱胁迫法,以2年生辽东栎幼树为试材,研究干旱胁迫对辽东栎光合作用的结构、物质交换、能量流动等方面的影响机制,筛选干旱敏感性指标,为辽东栎高效灌溉提供理论依据。
-
辽东栎种子于2018年采自山西省临汾市三交村(35°57′53.64″ N,112°4′35.76″ E,海拔1 197 m),在山西省林业科学与草原研究院阳曲试验基地(38.0981° N,112.7346° E,海拔961 m)进行播种和培育。播种容器高25 cm、上径36 cm、下径31 cm,容积26.6 L,pH 6.5~7.0。培养土为壤土,水分特征按照《森林土壤水分-物理性质的测定》(LY/T1215-1999)测定并计算,毛管孔隙度45.7% ± 8.4%,非毛管孔隙度4.8% ± 1.4%,总孔隙度50.5% ± 7.1%,土壤饱和含水量29.9% ± 3.7%。2021年1月选择15株生长状况良好、长势一致(苗高20 ± 5 cm,地径4.0 ± 0.5 mm)的幼树,带盆转移到人工气候室[光照强度350 μmol·m−2·s−1,光照时间10 h·d−1,昼夜环境温度(28 ± 2)°C/(22 ± 2)°C,空气湿度(50 ± 10)%],进行1个月的环境适应,期间采用与田间培养时相同的除草和浇水等管理措施。
-
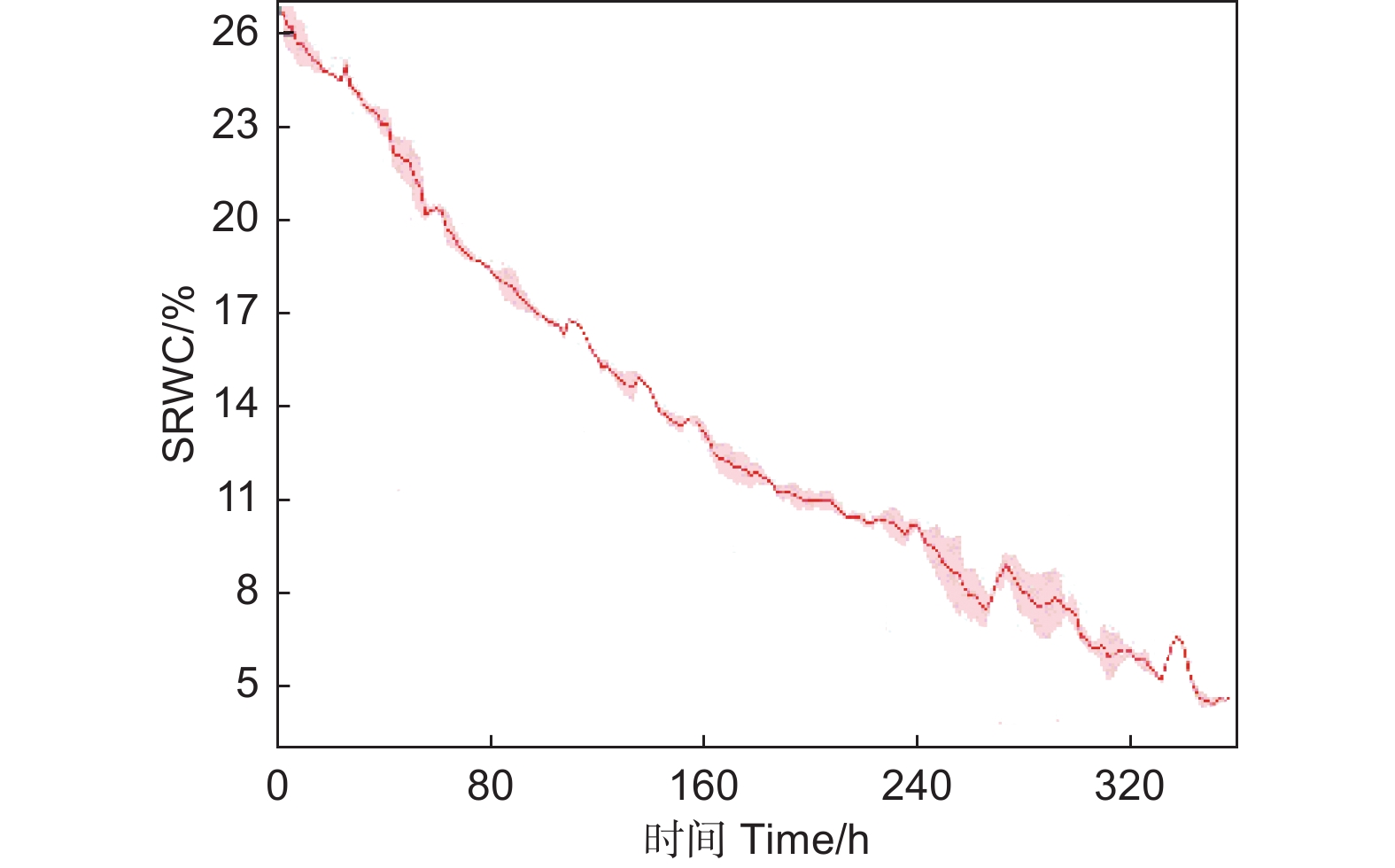
试验在人工气候室内进行,环境参数不变。采用盆栽渐进式干旱胁迫法进行干旱处理,即试验开始前48 h,将15株辽东栎幼树充分灌水,达到土壤最大持水量,然后停止浇水,进行自然干旱处理。随机选取5株幼树,在试验期间每2 h利用与电脑相连的土壤湿度传感器(ST-TR-EC,Sciento,中国)自动记录1次土壤相对含水率(SRWC)。根据预试验结果,当平均SRWC低于5%时,叶片永久萎蔫,此时试验停止(图1)。
Figure 1. Changes of soil relative water content (SRWC) in rooting zone under progressive drought conditions
-
试验期间,在每株上的半木质化枝上选择1片成熟、完整、无遮挡的叶片,于每日10:00,采用数码相机(H805,Forsafe,中国)在固定位置、镜头高度与叶柄基部相同的角度拍照。试验期间保持拍照植株的位置和角度不变动。获得的图像采用ImageJ 1.53c(National Institutes of Health,美国)测量叶倾角,共计30个重复。测量步骤如下(图2):

Figure 2. Schematic diagram of leaf droop angle measurement
(1)在试验开始时的照片中,将通过叶柄基部的水平线确定为基线;
(2)在其余照片中,测量叶柄基部至叶尖的连线与基线之间的夹角,即叶倾角;
当叶柄基部至叶尖的连线位于基线上部时,叶倾角定义为正值;位于基线下部时,叶倾角定义为负值。
-
每日9:00开始测定快速叶绿素荧光指标,每株选取1片成熟、完整、无遮挡的叶片,暗适应15 min,使用Mini调制式叶绿素荧光仪(FluorPen FP110,Photon Systems Instruments,捷克)测量叶肉部分。测量脉冲光强度900 μmol·m−2·s−1(持续30 µs),饱和光强度2 000 μmol·m−2·s−1,光化光强度300 μmol·m−2·s−1。根据获取的叶绿素荧光诱导动力学曲线计算:暗适应后照光150 μs和300 μs时的相对荧光强度VL和VK、光系统Ⅱ(PSⅡ)单位面积有活性反应中心数量RCO/CS、单位反应中心吸收的能量ABS/RC、单位反应中心捕获的用于还原QA的能量TRO/RC,单位反应中心用于将电子从Q− A传递到PQ的能量ETO/RC、单位反应中心用于将电子从Q− A传递到PSI的能量REO/RC、单位反应中心以热能形式耗散的能量DIO/RC、叶片性能指数PIABS。计算方法参考文献[24-25]。
快速叶绿素荧光指标测定完成后,选取同一叶片继续黑暗适应15 min后,采用前述Mini调制式叶绿素荧光仪,变更测量位置,测定叶肉部分的稳态叶绿素荧光,仪器参数设置不变。按照仪器内置的NPQ2程序(光周期200 s,每隔20 s测量1次,共10次;暗周期390 s,每隔60 s测量1次,共7次)自动计算出最大量子产额QYmax和非光化学猝灭系数NPQ。
-
叶绿素荧光测定后,在同一叶片采用光合仪(LCpro-T,ADC,英国)测定气体交换参数。仪器参数为:白色光源,光强300 μmol·m−2·s−1,开放气路,空气流速200 μmol·s−1,环境CO2浓度443 ± 12 μmol·mol−1,叶片温度25 ± 0.2℃。测量开始后,每隔30 s记录1次数据,将20 min内的最大值记为光合速率Pn、蒸腾速率E、气孔导度gs和胞间CO2浓度Ci。水分利用效率WUE由Pn与E的比值计算得出。
-
依据各项指标测定时的SRWC,将数据分为8个分组:
SRWC≥24.5%(标记为SRWC-26%),
24.5%>SRWC≥21.5%(标记为SRWC-23%),
21.5%>SRWC≥18.5%(标记为SRWC-20%),
18.5%>SRWC≥15.5%(标记为SRWC-17%),
15.5%>SRWC≥12.5%(标记为SRWC-14%),
12.5%>SRWC≥9.5%(标记为SRWC-11%),
9.5%>SRWC≥6.5%(标记为SRWC-8%),
SRWC<6.5%(标记为SRWC-5%)。
采用方差分析和数据平滑统计指标间的差异和变化趋势,数据平滑方式为LOESS,平滑阈值0.7。以SRWC 26%分组作为对照(CK)。使用R语言4.0.2(R Core Team,2020)和Origin(Origin Lab Corporation,2019)进行数据分析和绘图。数据分析前利用R语言的MASS和car程序包对数据进行正态性和方差齐性检验,必要时进行数据转换以满足数据分布的正态性要求,对不满足方差齐性的指标采用R语言的userfriendlyscience程序包进行Games-Howell方差分析。
-
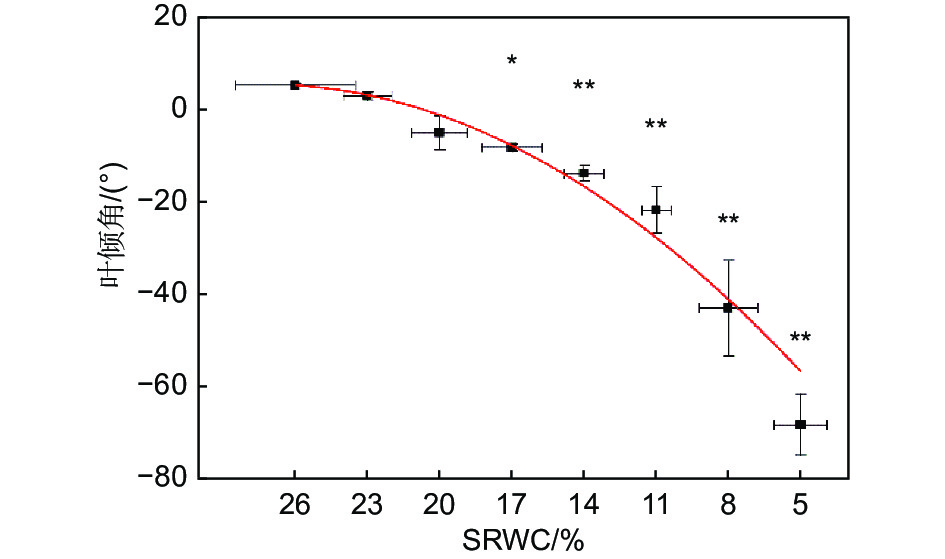
随着SRWC下降,叶倾角逐渐减小(R2=0.97,P<0.001)。当SRWC下降至17%时,叶倾角与对照相比显著减小(从5.2° ± 0.4°降低至−8.4° ± 0.7°),当SRWC降至5%时,叶倾角达到最小值(−68.5° ± 6.7°)(P<0.01)(见图3)。
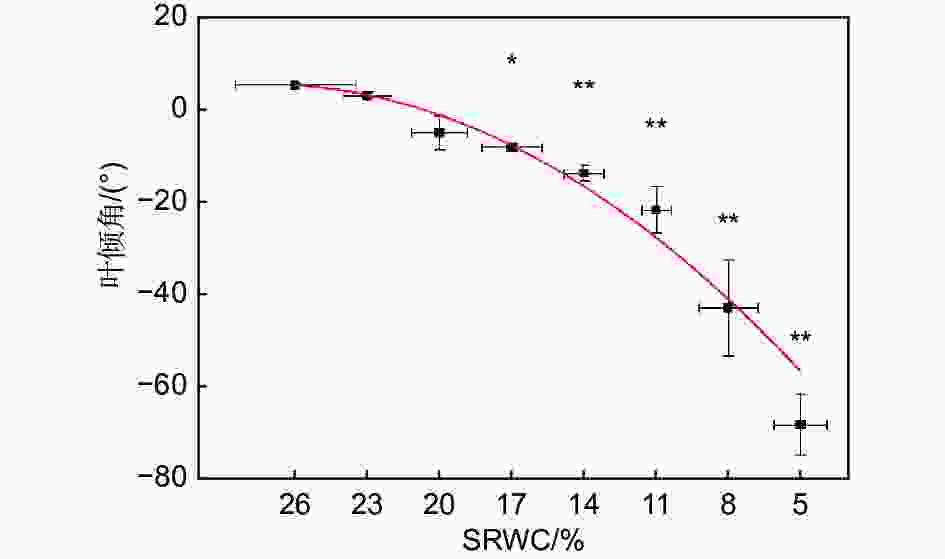
Figure 3. Effects of progressive drought on leaf droop angle
-
随着SRWC下降,VK、VL、ABS/RC、DIO/RC和TRO/RC均呈增加趋势,其中ABS/RC在SRWC-5%时显著增加到CK的116.7%,表明随着干旱程度加剧,放氧复合体和PSII反应中心内的电子转移逐渐受到干扰,PSⅡ单位面积反应中心吸收、耗散和捕获的光能逐渐增加。RC/CSO、ETO/RC、REO/RC和PIABS均随SRWC下降而下降,当SRWC为5%时,RC/CSO和PIABS较CK分别显著减少了20.2%和36.6%,稳态叶绿素荧光指标QYmax随着SRWC降低而上升,而NPQ随之先增后降,均与CK无显著差异(图4)。

Figure 4. Effects of progressive drought on chlorophyll fluorescence parameters
-
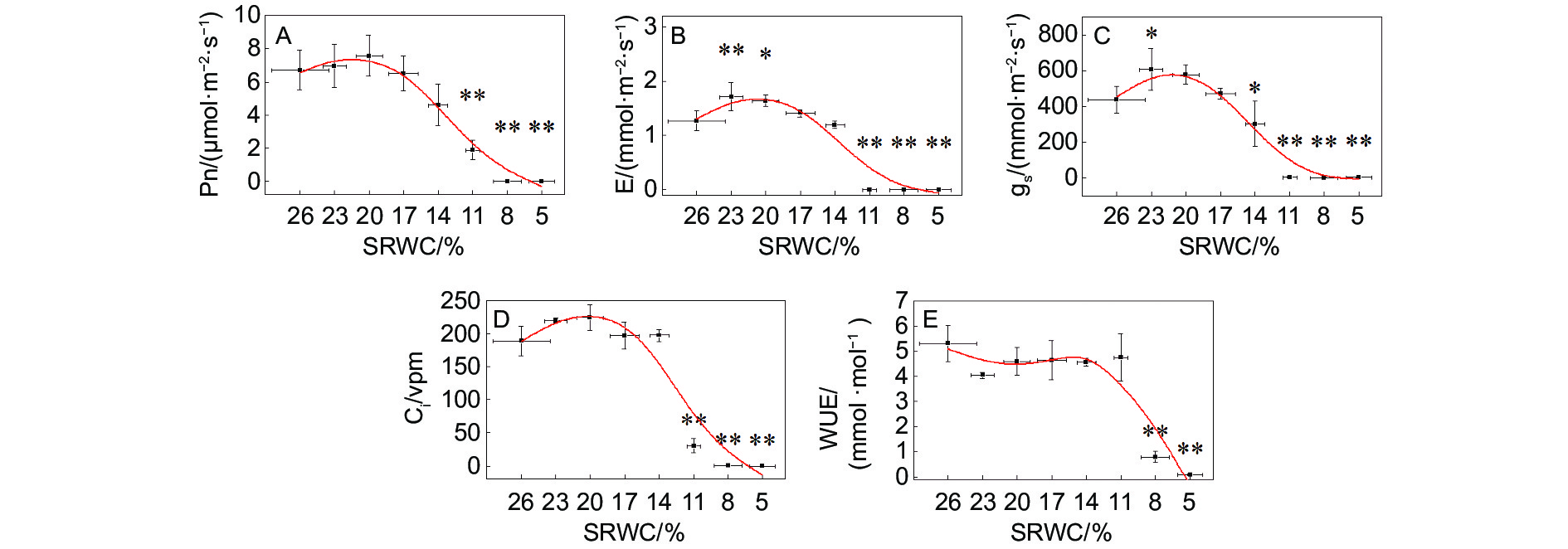
随着SRWC降低,Pn、E、gs和Ci先升高,后逐渐降低。E和gs在SRWC-23%时分别升高至CK的134.6%(P<0.01)和138.6%(P<0.05)。gs从SRWC-14%开始显著降低,Pn、E和Ci从SRWC-11%时开始显著降低。WUE随SRWC降低而减少,SRWC-8%时极显著降低至CK的15.0%(图5)。

Figure 5. Response of gas exchange parameters to progressive drought
-
本研究发现渐进式干旱过程中,叶倾角在SRWC为17%时显著减小,至5%达到最小值;叶绿素荧光参数中PSII有活性的反应中心数量、PSII与PSI之间的能量传递和PSII光化学活性逐渐下降,但最大量子产额与非光化学猝灭所受影响较小;气体交换参数在SRWC低于20%时开始受到影响,低于11%时影响显著。
-
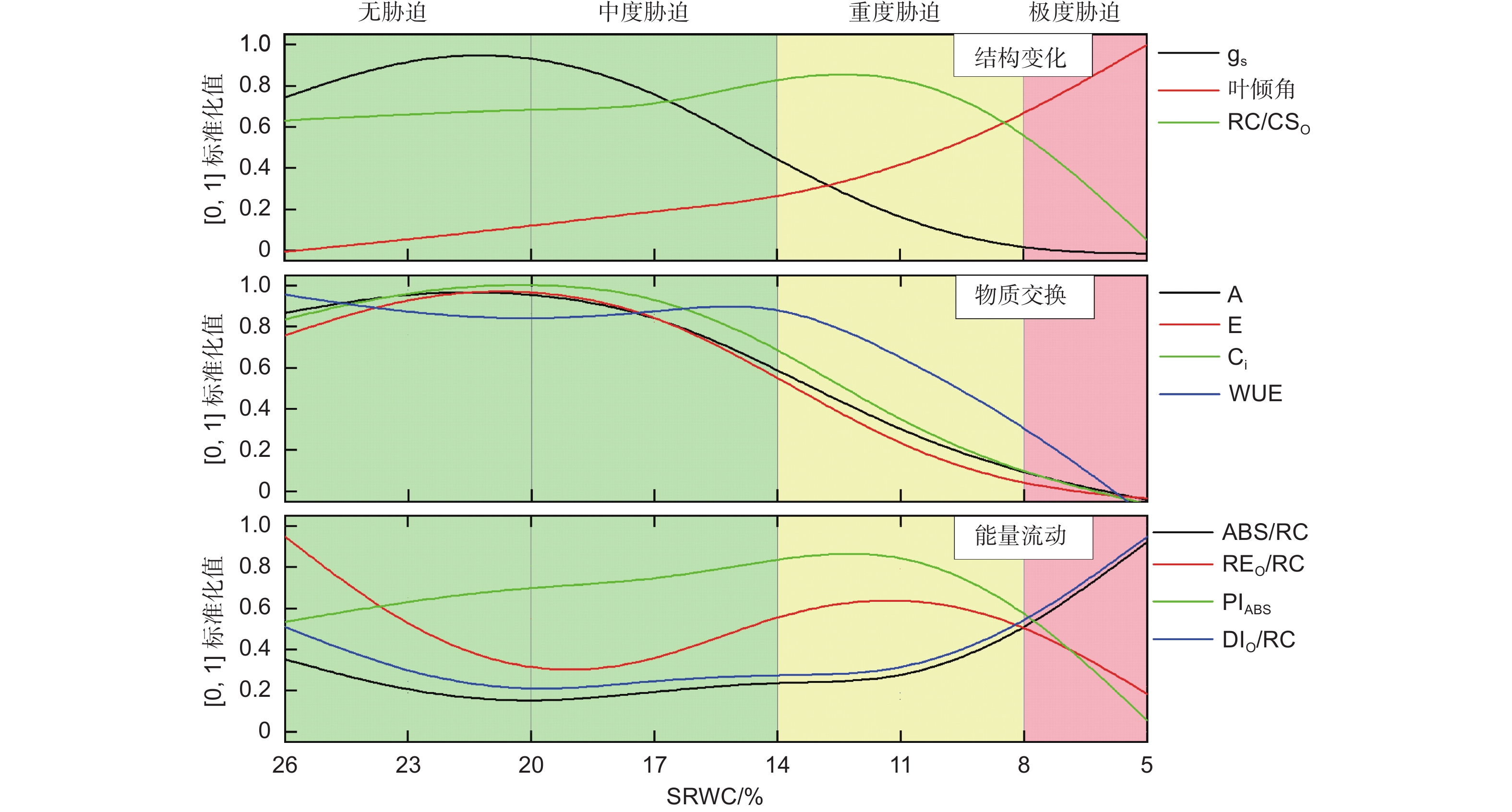
将不同土壤含水量对应的光合作用参数进行[0,1]标准化(图6),从光合机构的结构变化、物质交换、能量流动3方面分析辽东栎幼树叶片光合系统对干旱的响应,发现其存在明显的4个阶段:第1阶段(26%>SRWC>20%),无胁迫发生,A、E、Ci、gs持续上升;第2阶段(20%>SRWC>14%),中度干旱胁迫,A、E、Ci、gs持续下降,WUE基本保持稳定;第3阶段(14%>SRWC>8%),重度干旱胁迫,WUE、PIABS、REO/RC开始大幅度下降。第4阶段(SRWC<8%),极度干旱胁迫,A、E、Ci、gs趋近于0。其响应机制如下所述。
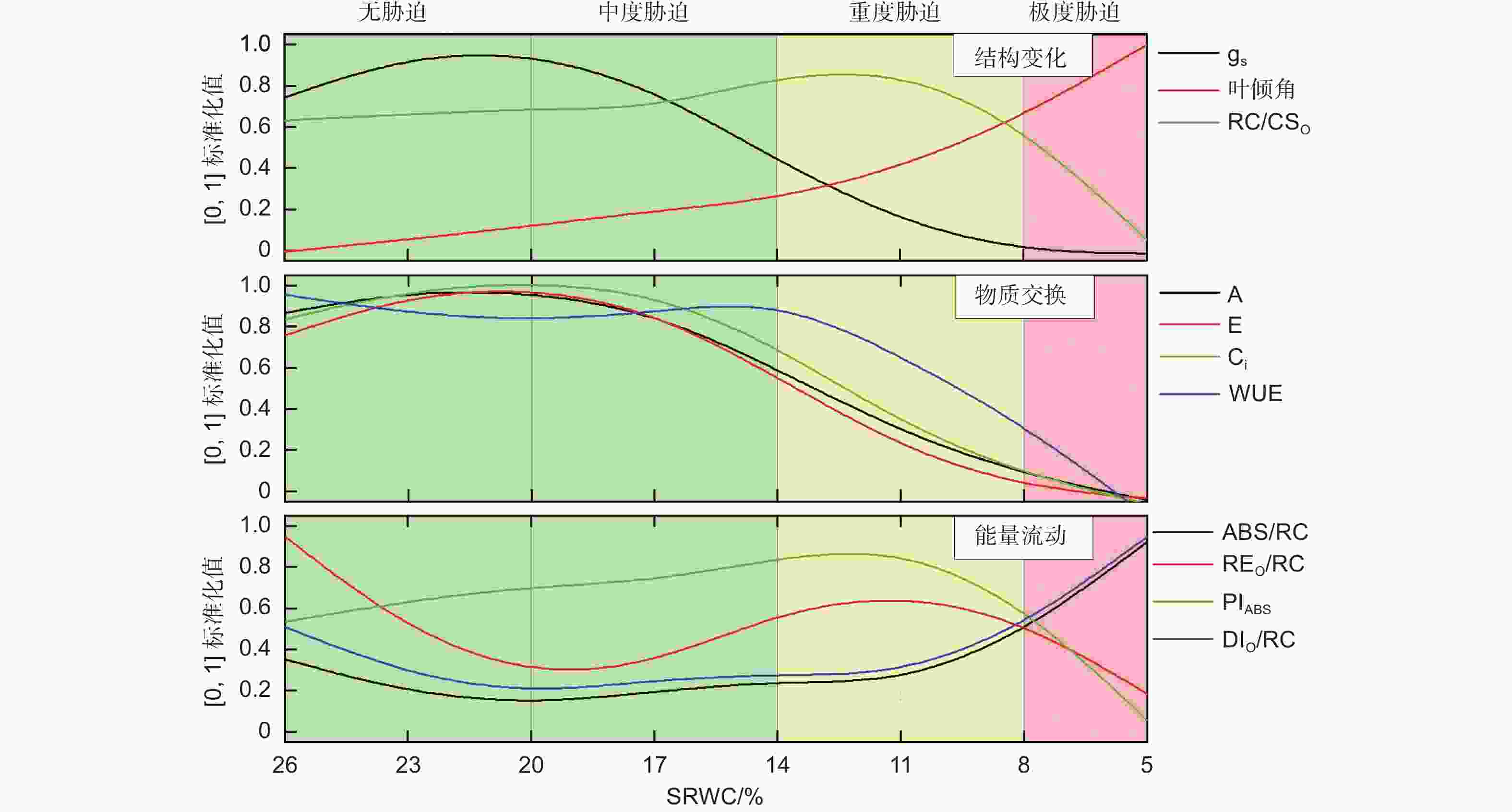
Figure 6. Stress-levels scheme and irrigation schedule of Q. liaotungensis seedling cultivation
通常认为,干旱胁迫会影响气孔开放程度、气体交换和PSⅡ运行效率[26-28]。无干旱胁迫时,SRWC下降导致了辽东栎幼树气孔导度(gs)和PSII光化学活性(PIABS)上升,叶倾角和PSⅡ单位面积有活性反应中心数量(RC/CSO)略有升高,虽然PSⅡ单位面积反应中心吸收(ABS/RC)和耗散的光能(DIO/RC)下降,但叶片光合能力仍然维持较高水平,表明此阶段尚未发生干旱胁迫。此阶段还观察到,单位反应中心用于将电子从Q− A传递到PSⅠ的能量(REO/RC)下降。已有研究表明,干旱胁迫会导致PSI受体侧的电子传递和量子产额下降[28]。鉴于此,REO/RC降低可能源于该参数对土壤含水量降低相对敏感。当感应到土壤含水量下降后,PSII立即通过降低向PSⅠ传输的电子效率,以避免未及时固定的电子与氧分子结合形成活性氧(ROS)[29]。
中度干旱胁迫时,辽东栎幼树关闭的气孔增多,但光合机构的能量流动未受明显影响,仅阻碍了CO2(Pn和Ci)和水(E)的物质交换,因此WUE仍维持较高水平。这一阶段的典型表现是gs显著下降和PSII的正常运转,这可能因为气孔限制被启动的SRWC阈值较非气孔限制高。该现象印证了前人关于气孔导度对干旱发生的敏感程度比叶绿素荧光参数高的论断[30]。此阶段还观察到WUE逐渐增加,类似现象已有报道[31],可能由于作为一种相对温和的干旱胁迫反应,自然状态下会频繁发生气孔关闭,WUE对此已经脱敏。为了确保有充足的电子在PSII和PSI之间传递能量,所以WUE不降反增。
重度干旱胁迫时,尽管PSII的能量吸收(ABS/RC)和热耗散(DIO/RC)开始增加,但在能量传输方面,PSII光化学活性(PIABS)及传递到PSI受体侧的能量(REO/RC)开始下降;在物质交换方面,水和CO2的流通进一步受到抑制;在结构方面,RC/CSO、叶倾角、gs急剧减少。这一阶段主要的变化是光合机构结构和能量流通的改变,源于非气孔降低了植物的光合能力。具体而言:首先,叶绿素荧光参数变化明显,表明PSII结构遭到破坏[32],因为重度干旱导致PSII捕获的光能与CO2还原利用的光能失衡,多余的电子与氧分子结合形成ROS,极易造成类囊体膜上的蛋白复合物受损;其次,WUE开始下降,这一现象可能与抗旱能力的种间差异有关;第三,叶片萎蔫逐渐加速,主要由于水份缺乏致使细胞膨胀压力下降。
极度干旱胁迫时,结构方面,叶片迅速下垂,RC/CSO和气孔导度趋近于零;在物质交换方面,叶片与环境间的H2O和CO2流动逐渐停止;在能量方面,PSII吸收和耗散的能量增加,而能量传递却迅速下降至0(PIABS和REO/RC)。此阶段叶绿素荧光参数的变化最为明显,反映出极度干旱胁迫对叶片光合机构造成了严重损伤,导致PSII受体和供体侧的电子供应失衡。在此阶段还观察到ABS/RC急速上升,Manaa等人也发现了类似现象[33],这可能是植物的胁迫应激反应,缓解PSII反应中心的大量关闭和电子转运速率的大幅下降。
在水分紧缺的情况下,植株幼苗的耗水量减少,可以维持较高的WUE[34]。本研究发现,SRWC介于14%至26%之间时,WUE、光合作用的固碳能力(Pn)和PSII的光利用效率(PIABS)均没有显著变化,表明中度干旱对辽东栎幼树的水分利用效率和生产力没有明显影响。当缺少灌溉水时,适度减少用水量仍然能获得较高的辽东栎幼树生长量。
-
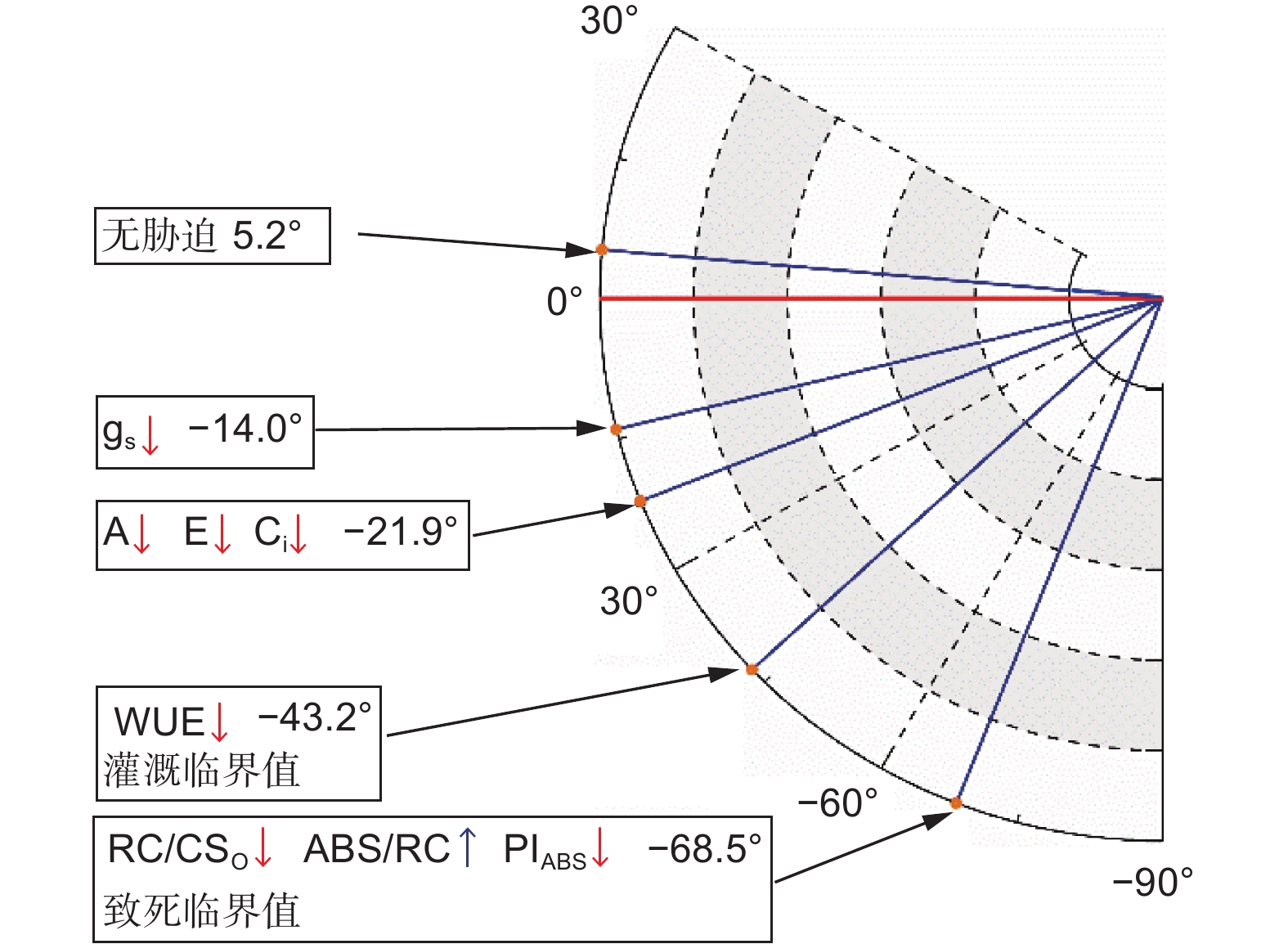
辽东栎幼树叶倾角对土壤干旱的响应存在阶段性差异,将叶倾角与光合参数的变化趋势匹配(图7),可发现:当叶倾角为−14.0° ± 1.7°时,气孔闭合量增加;当叶倾角为−43.2° ± 10.3°时,WUE显著降低,此时需要灌溉;当叶倾角为−68.5° ± 6.7°时,光合机构受到严重破坏,此时必须立即灌溉。通过叶倾角检测干旱胁迫程度的方法具有观测便捷、成本较低的优势,但需要考虑拍照时风速对叶倾角的干扰。

Figure 7. Leaf droop angle prediction model for the effects of progressive drought on photosynthesis of Q. liaotungensis
评估干旱胁迫程度的常用方法为监测SRWC,但测量程序繁琐费时。利用植物生理指标检测干旱胁迫具有测量速度快和结果精准的优势。已有研究将ABS/RC[35]、DIO/RC[36]、RC/CSO和PIABS[37]做为检测植物干旱胁迫程度的指标。我们的研究发现辽东栎幼树叶片的这些参数在重度和极端干旱胁迫下变化明显,可作为严重干旱胁迫发生的预警指标。
-
综上所述,辽东栎幼树叶片遭受渐进式干旱胁迫时,光合系统的气体交换变化最显著,其次是叶倾角,最后是PSII的电子传递和能量耗散。干旱胁迫下光合系统的响应可划分为4个阶段:(1)无干旱胁迫,叶片结构稳定,物质交换正常;(2)中度干旱胁迫,叶倾角降低、气孔的结构开始闭合,水和CO2的物质交换逐渐减少,但尚能高效的利用土壤水分;(3)重度干旱胁迫,结构变化持续,物质交换和能量流动受阻,土壤水分利用效率降低;(4)极度干旱胁迫,光合系统的结构变化、物质交换和能量流通均趋于停止。据此,本研究筛选出灌溉的预警指标—叶倾角,以及叶片的致死预警指标—RCO/CS、ABS/RC、DIO/RC、PIABS。这一结论可为制定辽东栎育苗的高效灌溉方案提供理论依据和数据支持。
Effects and Sensitivity of Progressive Drought on Photosynthetic Characteristics of Quercus liaotungensis Seedlings
- Received Date: 2022-05-05
- Accepted Date: 2022-08-09
- Available Online: 2023-06-20
Abstract:


 DownLoad:
DownLoad: