



-
磷是植物生长所必需的营养元素之一,参与植物体内众多的生理生化过程,在植物生长发育过程中发挥重要作用[1-2]。光合作用是植物固定无机碳为有机碳的重要生理化学过程,是植物进行碳吸收的基本动力。土壤磷养分在植物进行光合作用过程中扮演重要角色[3]。如,赵燕等[4]发现随着基质磷浓度增加,毛白杨(Populus tomentosaa Carr.)幼苗的净光合速率会先升高后下降。在热带和亚热带森林,磷限制植物生长现象普遍存在[5],且随着工业发展和森林的演替,森林的缺磷程度不断加剧[6]。不同森林植被生长对土壤磷养分含量的响应也存在差异[7-12]。夏威夷多型铁心木(Metrosideros polymorpha Gaudich.)林分群落植被的生长对磷添加表现出积极响应[7],但土壤磷的可利用性对热带地区森林树木生长呈现抑制或不显著作用[8-10]。此外,在亚热带区域相同人工林树种对磷添加的生长与光合响应研究结果也存在一定争议[11-12]。因此,进一步探究磷浓度对树木光合作用和生长的影响十分必要。
马尾松(Pinus massoniana Lamb.)喜光、耐旱、耐贫瘠,是亚热带地区广泛种植的先锋树种[11-14]。全国马尾松林土壤养分调查结果表明马尾松林分土壤的全磷(0.320 mg·kg−1)和有效磷(2.247 mg·kg−1)平均含量较低,处于“极缺”水平[13]。Huang等[14]也发现在主要分布区马尾松生长与土壤有效磷含量显著正相关,说明在亚热带区域土壤磷限制了马尾松的生长[15]。此外,Yu等[16]预测亚热带地区的土壤全磷含量随时间演替仍有下降趋势。因此,马尾松林分生长可能面临日益加剧的磷限制的问题。
目前,部分学者也通过控制试验开展关于马尾松生长对磷浓度响应方面的研究,但研究结果存在差异[11-12,17]。如黄盛怡等[11]和庞丽等[17]发现马尾松幼苗生长随磷增加而显著加快,但也有研究表明在不同浓度磷添加条件下幼苗高生长并不存在显著的差异[12,17]。除此之外,目前大多数关于磷胁迫相关研究中培养基质磷添加含量普遍处于较高水平(对照组设置为8~32 mg·kg−1)[11,13,17],内容更多局限于比较植株对高磷和低磷两个浓度梯度的生长生理响应差异[11,17],中间不同的低磷浓度对植物生长和光合造成的影响差异往往被忽略。同时,现有磷控制试验培养时间相对较短[17-18],缺少对整个生长季的研究[19]。
因此,本研究以两年生马尾松幼苗为研究对象,以全国马尾松林分表层土土壤有效磷含量为依据[13],培养基质设置7个磷浓度梯度,探究不同磷浓度下马尾松幼苗在生长季的不同时间生长参数和光合参数的差异,为马尾松林经营管理和更精准的生长模拟预测提供试验依据,也有助于更好地揭示耐瘠薄树种对低磷环境的适应机制。
-
本研究以贵州省都匀市马鞍山林场种子园的两年生马尾松良种幼苗为试验材料。2020年12月将马尾松幼苗移栽至湖北秭归三峡库区森林生态系统国家定位观测研究站(110°54′ E,30°53′ N,海拔296 m)进行练苗。2021年5月初将个体大小一致、长势均匀的幼苗移栽到花盆(高17 cm,直径16.5 cm)中进行养分控制培养,每盆一株。培养基质是以石英砂:蛭石:珍珠岩 = 7:2:1的混合基质。移栽前,培养基质用3%稀盐酸进行酸洗,浸泡1周去除杂质后倒入营养液平衡离子[18]。所有幼苗植株置于定位站露天培养苗床上进行试验培养及后期观测。
-
以全国马尾松林表层土(0~20 cm)土壤有效磷含量(2.25 mg·kg−1)为参照[13],设置无磷(0AP,0 mg·kg−1)、1/8倍磷(1/8AP,0.291 mg·kg−1)、1/4倍磷(1/4AP,0.581 mg·kg−1)、1/2倍磷(1/2AP,1.162 mg·kg−1)、对照组(CK,2.325 mg·kg−1)、2倍磷(2AP,4.650 mg·kg−1)和4倍磷(4AP,9.299 mg·kg−1)等共7个磷浓度处理组,每个处理组预备200株幼苗。根据基质质量(约3 kg·盆-1),计算每盆植株的养分需求量,为避免高浓度离子毒害作用,在栽植后的1个月内,分5次将营养液均匀浇入盆中,每间隔7 d添加一次,来降低介质和植物组织中营养物质浓度的波动;同时为保持离子的有效性,将大量元素和微量元素分别添加。不同浓度的营养液中大量元素用量详见表1,微量元素营养液由改良的Hoagland营养液[20]提供,其中Fe3+、Zn2+、Mn2+、Cu2+、Mo6+、B4+的含量分别为7.149 mg·kg−1、0.131 mg·kg−1、0.110 mg·kg−1、0.032 mg·kg−1、0.048 mg·kg−1和0.270 mg·kg−1;营养液pH值在5.3~6.0之间。由于石英砂基质保水性差,需每日日落后用雾化喷灌装置进行灌溉降温并提供充足的水分,同时用遮荫网进行适当遮荫来减少基质水分蒸发。养分控制试验从2021年5月初开始,到2021年11月末结束,生长周期试验持续7个月。
处理梯度
Treatment gradient四水硝酸钙
Ca(NO3)2·4H2O/(g·kg−1)磷酸二氢钾
KH2PO4/(g·kg−1)硫酸镁
MgSO4/(g·kg−1)氯化钾KCl/(g·kg−1) 无磷 0AP 0.945 0 0.120 0.253 1/8倍磷 1/8AP 0.945 0.001 0.120 0.253 1/4倍磷 1/4AP 0.945 0.002 0.120 0.253 1/2倍磷 1/2AP 0.945 0.003 0.120 0.252 对照组 CK 0.945 0.006 0.120 0.250 2倍磷 2AP 0.945 0.012 0.120 0.247 4倍磷 4AP 0.945 0.024 0.120 0.240 Table 1. Contents of macroelements in different phosphorus content groups
-
在5月初养分处理前,用钢直尺(精度0.1 cm)测量所有幼苗初始株高值(cm);养分处理后分别在7月、9月和11月初再次测量所有待测植株的株高(cm)。两次连续测量的马尾松幼苗株高之差记为幼苗株高在此期间的相对生长增量(Relative height increment,RHI)[21]:
式中H表示测试月幼苗的株高,n为测量月份(n = 7,9,11)。
将不同磷浓度处理下幼苗株高数据与生长时间(月)分别进行线性拟合,发现方程R2的范围在0.935~0.979(表2),表明拟合方程的斜率能够表征马尾松幼苗高生长速率的大小[21]。因此,本研究中将该斜率记为马尾松幼苗的相对生长速率(Relative growth rate, RGR)。
处理梯度 Treatment gradient 公式 Formula R2 P 无磷 0AP H=1.857 t + 53.736 0.954 0.023* 1/8倍磷 1/8AP H=1.984 t + 56.636 0.979 0.011* 1/4倍磷 1/4AP H=1.812 t + 52.450 0.947 0.027* 1/2倍磷 1/2AP H=1.910 t + 57.275 0.935 0.012* 对照组 CK H=1.913 t + 55.265 0.948 0.026* 2倍磷 2AP H=2.059 t + 56.390 0.979 0.011* 4倍磷 4AP H=2.349 t + 50.555 0.956 0.022* 注:P<0.05,*;P<0.01,**;P<0.001,***。下同
Notes: P<0.05, *; P<0.01, **; P<0.001, ***. The same were as belowTable 2. The parameters of linear regression model between growing time and height of P. massoniana seedlings
-
用Li-6400XT便携式光合作用测量系统(Li-Cor,Lincoln,USA)测定针叶气体交换参数。选择大小相似,生长势良好的不同处理植株各4株,分别在5月、6月、7月、8月、9月、10月的连续晴朗的上午9:00—11:30,选择植株中上部的成熟叶测定气体交换参数。测定过程中设置叶室温度为25 ℃,光合有效辐射为1 000 μmol·m−2·s−1,气体流速500 μmol·s−1。测量前对马尾松针叶进行光诱导,待数值稳定后记录数据。
-
所有数据的数理统计分析和绘图均由SPSS v23.0和R4.1.2软件(
http://www.r-project.org/ )完成。本研究所有数据通过正态分布(Shapiro-Wilk检验)和方差齐次性检验(Bartlett检验),满足数据进一步分析条件。利用R语言 “lme4”程序包完成线性混合效应模型(Linear mixed-effects model)的分析,其中将重复作为随机因子,分别分析生长时间、磷浓度及二者间的交互作用对马尾松幼苗的高生长参数。
RHI和光合参数指标(净光合速率(Net photosynthetic rate,Pn)、气孔导度(Conductance to H2O,Gs)、胞间CO2浓度(Intercellular CO2 concentration,Ci)和蒸腾速率(Transpiration rate,Tr))的影响。用单因素方差分析(One-way ANOVA)和LSD检验方法比较不同生长时间和磷浓度梯度下高生长和光合参数指标的差异。用学生检验(Student's t test)和简单线性回归方法确定基质磷浓度与幼苗相对生长速率的关系。通过相关分析和偏相关分析(Partial correlation analysis)描述生长时间、磷浓度及其交互作用与高生长和光合参数之间的相关性。
-
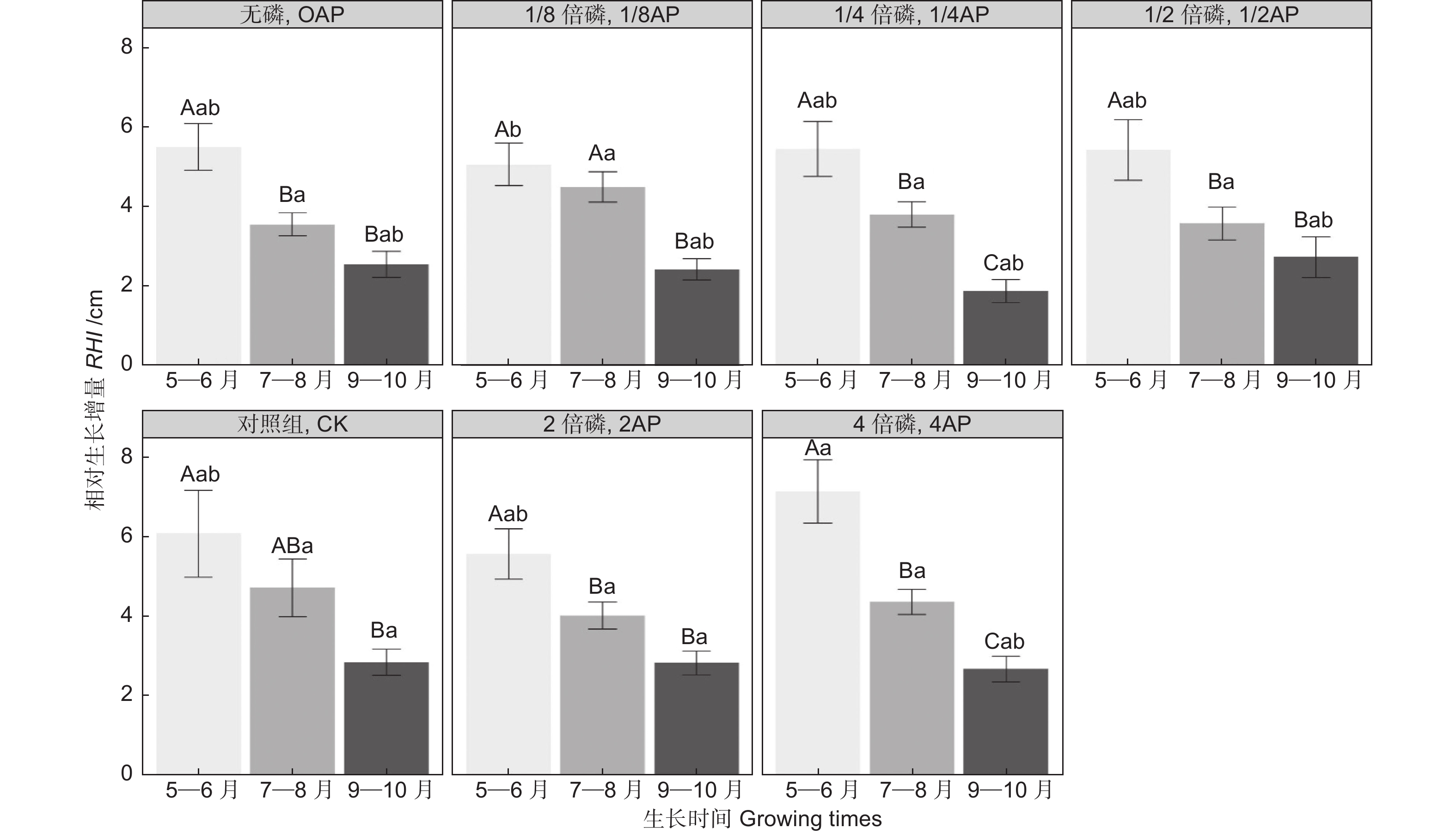
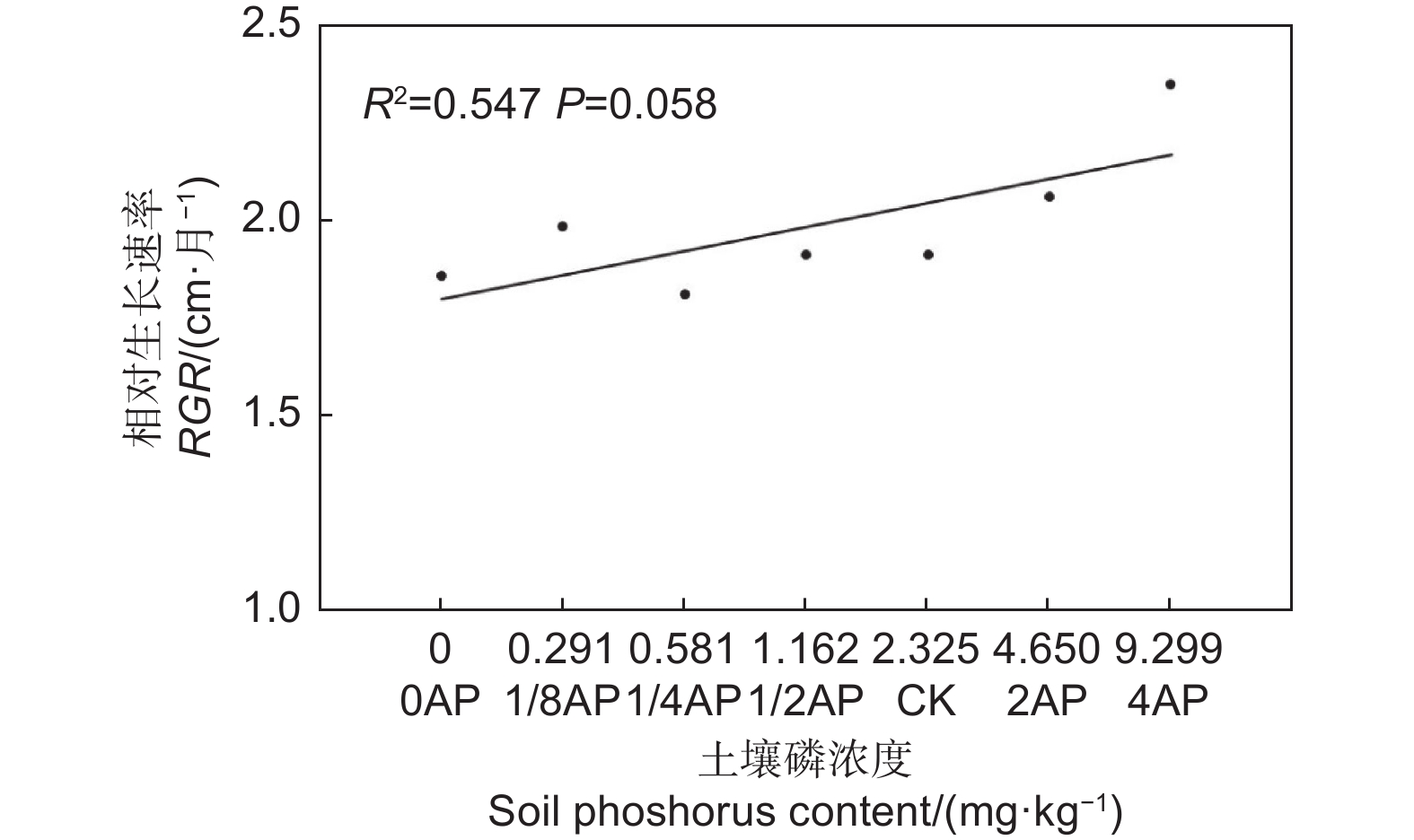
生长时间和磷浓度对马尾松幼苗的RHI影响显著(表3,P<0.01)。如图1所示,在不同的磷浓度条件下,幼苗RHI均随生长时间的延长而减小, 且RHI5~6显著高于RHI9~10(P<0.05)。同一生长时间段内,仅4AP生长条件下的RHI5~6显著高于1/8AP(图1,P<0.05)。此外,马尾松幼苗的RGR受基质磷浓度影响显著(t-Value
= 29.113,P = 0.000),且RGR随基质磷浓度的升高而线性增高(图2)。 因子
Indicator磷浓度
Phosphorus content生长时间
Growing time磷浓度×时间
Phosphorus content × Growing timeF-value P F-value P F-value P 相对生长增量 RHI 2.810 0.004** 293.052 0.000*** 0.755 0.796 净光合速率 Pn 24.167 0.000*** 1 746.995 0.000*** 59.456 0.000*** 气孔导度 Gs 127.570 0.000*** 10 287.470 0.000*** 104.970 0.000*** 胞间CO2浓度 Ci 20.274 0.000*** 2 345.131 0.000*** 32.512 0.000*** 蒸腾速率 Tr 184.030 0.000*** 12 552.540 0.000*** 210.810 0.000*** Table 3. Effects of growing time, phosphorus content, and their interactions on relative height increment and photosynthetic parameters of P.massoniana seedlings
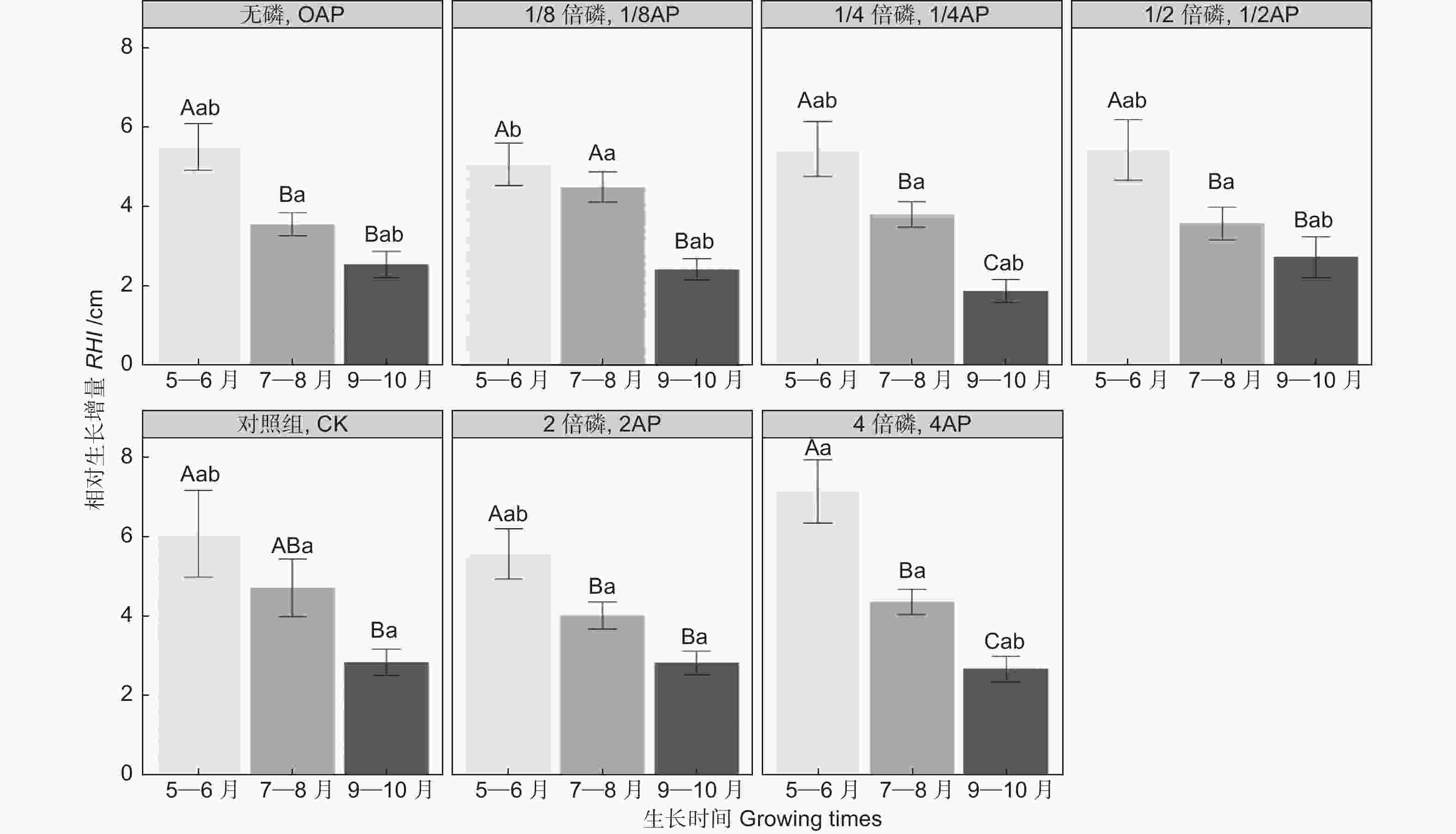
Figure 1. Variation of relative height increment in P. massoniana seedlings in different soil phosphorus contents during different growing times

Figure 2. Bivariate relationship between soil phosphorus contents and relative growth rate of P. massoniana seedlings
-
磷浓度、生长时间以及二者间的交互作用对马尾松幼苗的Pn、Gs、Ci和Tr等光合参数均有显著影响(表3,P<0.01)。如表4所示,在相同磷浓度条件下,Pn在不同生长时间段内存在显著差异(P<0.05),除CK处理,Pn均在8月达到最大值;在4AP条件下,Pn在7月、8月和9月显著高于10月。Gs、Ci、Tr与Pn表现出相似的时间变化趋势,即随生长时间先增长后下降(表4);相同磷浓度条件下,在不同生长时间的Gs、Ci、Tr差异显著(P<0.05),7—9月值最高(表4)。在8月和9月,Ci、Tr在4AP条件下显著高于低磷条件(0AP、1/8AP和1/4AP);在5月和6月,低磷条件下的Ci、Tr显著高于高磷处理组(2AP和4AP)(表4,P<0.05)。
光合参数
Parameter5月
May6月
June7月
July8月
Augest9月
September10月
October无磷 0AP 净光合速率 Pn 0.489 ± 0.045 Fd 3.052 ± 0.136 Ea 8.591 ± 0.126 Ca 11.460 ± 0.507 Acd 9.719 ± 0.183 Ba 3.925 ± 0.118 Dc 气孔导度 Gs 0.009 ± 0.001 Dc 0.023 ± 0.001 Dab 0.101 ± 0.002 Bd 0.602 ± 0.012 Aab 0.568 ± 0.011 Ab 0.043 ± 0.001 Cc 胞间CO2浓度 Ci 288.745 ± 12.107 Ba 182.986 ± 2.900 Dc 253.569 ± 3.205 Cc 374.556 ± 2.321 Aab 371.886 ± 1.118 Abc 253.302 ± 4.506 Cb 蒸腾速率 Tr 0.342 ± 0.015 Dd 0.521 ± 0.030 Ca 1.054 ± 0.008 Bd 3.846 ± 0.081 Ab 3.794 ± 0.062 Ac 0.531 ± 0.011 Cd 1/8倍磷 1/8AP 净光合速率 Pn 3.169 ± 0.134 Ea 3.007 ± 0.165 Da 6.926 ± 0.094 Cc 13.088 ± 0.432 Abc 8.496 ± 0.183 Bb 3.145 ± 0.138 Dd 气孔导度 Gs 0.030 ± 0.002 Da 0.016 ± 0.001 Dd 0.133 ± 0.003 Bc 0.625 ± 0.013 Aa 0.553 ± 0.011 Ab 0.100 ± 0.003 Ca 胞间CO2浓度 Ci 212.857 ± 1.979 Db 247.100 ± 3.639 Ca 304.003 ± 3.066 Bb 378.667 ± 2.968 Aab 370.981 ± 1.034 Ac 299.400 ± 5.281 Ba 蒸腾速率 Tr 1.280 ± 0.058 Ba 0.309 ± 0.011 Dcd 1.297 ± 0.028 Bc 4.012 ± 0.088 Aab 3.841 ± 0.073 Abc 1.031 ± 0.021 Ca 1/4倍磷 1/4AP 净光合速率 Pn 2.529 ± 0.044 Eb 2.279 ± 0.122 Eb 3.843 ± 0.256 De 15.133 ± 0.707 Aa 10.240 ± 0.196 Ba 5.265 ± 0.147 Cab 气孔导度 Gs 0.017 ± 0.000 Cb 0.021 ± 0.001 Cbc 0.060 ± 0.004 Bf 0.468 ± 0.010 Ac 0.402 ± 0.012 Ad 0.053 ± 0.001 Bb 胞间CO2浓度 Ci 150.802 ± 2.065 Dc 239.876 ± 4.615 Cab 255.639 ± 8.275 Bc 381.289 ± 4.029 Aa 354.678 ± 3.568 Ad 232.541 ± 4.578 Ccd 蒸腾速率 Tr 0.798 ± 0.008 Bb 0.470 ± 0.021 Ca 0.616 ± 0.040 Bf 3.356 ± 0.079 Ac 2.910 ± 0.064 Ae 0.717 ± 0.021 Bc 1/2倍磷 1/2AP 净光合速率 Pn 2.730 ± 0.079 Dab 3.007 ± 0.165 Da 6.222 ± 0.219 Cd 10.628 ± 0.948 Ad 8.496 ± 0.183 Bb 3.145 ± 0.138 Dd 气孔导度 Gs 0.018 ± 0.000 Db 0.020 ± 0.001 Dbc 0.070 ± 0.002 Be 0.584 ± 0.010 Aab 0.549 ± 0.009 Ab 0.032 ± 0.001 Cd 胞间CO2浓度 Ci 165.346 ± 4.382 Cc 168.897 ± 4.208 Cd 250.187 ± 6.273 Bc 383.211 ± 2.987 Aa 376.275 ± 1.025 Aab 240.331 ± 4.496 Bbc 蒸腾速率 Tr 0.701 ± 0.032 Bb 0.384 ± 0.022 Cb 0.637 ± 0.020 Bf 4.113 ± 0.102 Aa 3.972 ± 0.063 Aab 0.468 ± 0.0110 Ce 对照组 CK 净光合速率 Pn 1.459 ± 0.011 Dc 2.139 ± 0.148 Db 8.328 ± 0.506 Aab 6.386 ± 0.469 Be 8.635 ± 0.323 Ab 5.184 ± 0.167 Cab 气孔导度 Gs 0.011 ± 0.000 Dc 0.019 ± 0.001 Dcd 0.077 ± 0.004 Be 0.389 ± 0.008 Ad 0.461 ± 0.010 Ac 0.053 ± 0.002 Cb 胞间CO2浓度 Ci 160.194 ± 5.135 Dc 245.621 ± 4.995 Ba 230.055 ± 3.825 Cd 359.778 ± 1.039 Ac 373.345 ± 1.501 Abc 224.969 ± 5.089 Cd 蒸腾速率 Tr 0.535 ± 0.011 Cc 0.343 ± 0.021 Cbc 0.773 ± 0.040 Be 3.098 ± 0.065 Ad 3.406 ± 0.054 Ad 0.769 ± 0.019 Bb 2倍磷 2AP 净光合速率 Pn 1.503 ± 0.064 Ec 1.472 ± 0.088 Ed 7.984 ± 0.108 Cb 13.460 ± 0.452 Aab 8.571 ± 0.170 Bb 4.842 ± 0.143 Db 气孔导度 Gs 0.012 ± 0.000 Dc 0.016 ± 0.001 Dd 0.242 ± 0.004 Bb 0.665 ± 0.012 Aa 0.605 ± 0.011 Aa 0.050 ± 0.001 Cb 胞间CO2浓度 Ci 157.584 ± 3.788 Dc 232.621 ± 3.992 Cb 334.049 ± 1.665 Ba 387.692 ± 4.015 Aa 379.243 ± 0.766 Aa 234.204 ± 4.228 Ccd 蒸腾速率 Tr 0.452 ± 0.008 CDcd 0.210 ± 0.009 De 2.447 ± 0.024 Bb 4.258 ± 0.069 Aa 3.921 ± 0.055Aabc 0.750 ± 0.015 Cbc 4倍磷 4 4AP 净光合速率 Pn 1.226 ± 0.011 Bc 2.029 ± 0.116 Bbc 8.555 ± 0.122 Aa 8.015 ± 0.662 Ae 7.864 ± 0.135 Ac 2.660 ± 0.104 Be 气孔导度 Gs 0.008 ± 0.000 Cc 0.026 ± 0.002 Ca 0.272 ± 0.003 Ba 0.550 ± 0.010 Ab 0.546 ± 0.010 Ab 0.025 ± 0.001 Ce 胞间CO2浓度 Ci 148.373 ± 2.852 Dc 228.389 ± 5.704 Cb 334.975 ± 1.021 Ba 367.730 ± 2.592Ab 379.254 ± 0.764 Aa 223.245 ± 5.370 Cd 蒸腾速率 Tr 0.402 ± 0.003CDcd 0.272 ± 0.017 Dd 3.056 ± 0.008 Ba 4.012 ± 0.088 Aab 4.058 ± 0.058 Aa 0.412 ± 0.012 Cf 注:表中数据为平均值 ± 标准误差。同一行数值不同大写字母表示相同磷浓度不同生长时间马尾松幼苗光合参数在0.05 水平上差异显著,同列不同小写字母表示相同生长时间不同磷浓度幼苗光合参数在0.05 水平上差异显著。
Notes: All data are expressed as mean ± se. For photosynthetic parameters of P. massoniana, different capital letters in same rows meant significant difference among different growing times in the same phosphorus content, and different lowercase letters in same columns demoted significant difference among different phosphorus contents in the same growing time at the 0.05 level.Table 4. Comparison on photosynthetic parameters of P. massoniana seedlings in different growing times and phosphorus contents
-
如表5所示,Pn和Ci与RHI显著正相关(P<0.05);生长时间与RHI显著正相关(P<0.01),与Pn、Gs、Ci也有表现出一定的正相关关系(0.05<P<0.1)。当控制磷浓度时,马尾松幼苗的RHI与Pn和Ci以及生长时间之间显著正相关(P<0.05);生长时间与Ci显著正相关。当控制幼苗生长时间或同时控制磷浓度和生长时间时,RHI和光合参数与磷浓度之间无显著相关性,RHI与光合参数的相关性也不显著。
控制变量
Controlled variable变量
Variable净光合速率
Pn气孔导度
Gs胞间CO2浓度
Ci蒸腾速率
Tr相对生长量
RHI生长时间
Growing
time磷浓度
Phosphorus
content无控制变量
No controlled variables净光合速率 Pn 1 0.783** 0.787** 0.783** 0.426* 0.387 −0.013 气孔导度 Gs 0.783** 1 0.885** 0.979** 0.338 0.338 0.09 胞间 CO2浓度 Ci 0.787** 0.885** 1 0.874** 0.453* 0.421* −0.045 蒸腾速率 Tr 0.783** 0.979** 0.874** 1 0.282 0.261 0.135 相对生长增量 RHI 0.426* 0.338 0.453* 0.282 1 0.906** 0.131 生长时间 Growing time 0.387 0.338 0.421* 0.261 0.906** 1 0.061 磷浓度 Phosphorus content −0.013 0.09 −0.045 0.135 0.131 0.061 1 磷浓度
Phosphorus content净光合速率 Pn 1 0.793*** 0.788*** 0.802*** 0.428* 0.389 气孔导度 Gs 0.793*** 1 0.890*** 0.981*** 0.338 0.338 胞间 CO2浓度 Ci 0.788*** 0.890*** 1 0.885*** 0.453* 0.421* 蒸腾速率 Tr 0.802*** 0.981*** 0.885*** 1 0.283 0.262 相对生长增量 RHI 0.428* 0.338 0.453* 0.283 1 0.906*** 生长时间
Growing time0.389 0.338 0.421* 0.262 0.906*** 1 生长时间
Growing time净光合速率 Pn 1 0.751*** 0.746*** 0.766*** 0.192 −0.066 气孔导度 Gs 0.751*** 1 0.871*** 0.981*** 0.081 0.095 胞间 CO2浓度 Ci 0.746*** 0.871*** 1 0.873*** 0.187 −0.012 蒸腾速率 Tr 0.766*** 0.981*** 0.873*** 1 0.112 0.152 相对生长增量 RHI 0.192 0.081 0.187 0.112 1 0.012 磷浓度 Phosphorus content −0.066 0.095 −0.012 0.152 0.012 1 磷浓度和生长时间
Phosphorus content and
growing time净光合速率 Pn 1 0.763*** 0.747*** 0.787*** 0.194 气孔导度 Gs 0.763*** 1 0.876*** 0.982*** 0.08 胞间 CO2浓度 Ci 0.747*** 0.876*** 1 0.885*** 0.187 蒸腾速率 Tr 0.787*** 0.982*** 0.885*** 1 0.111 相对生长增量 RHI 0.194 0.08 0.187 0.111 1 Table 5. Correlation coefficients between relative height increment and photosynthetic parameters of P. massoniana seedlings in different phosphorus contents
-
磷与光合作用之间的关系可以用某些光合指标建立线性或对数函数来表示[22-26],但在不同研究中磷与光合作用的关系并未达成统一[27]。例如,Prior等[28]对热带稀树草原的红穗花(Erythrophleum chlorostachys (F.Muell.) Baill.)、金合欢(Acacia auriculiformis Benth.)和澳洲蓝柏(Callitris intratropica R.T.Baker & H.G.Sm.)等植被的光合作用研究表明光合作用受到磷的制约,而Smith等[29]对全球201个站点不同植物光合作用的研究发现光合作用与土壤养分含量无关。在本研究中,马尾松幼苗的Pn、Gs、Ci和Tr均受到不同磷浓度的显著影响(表3,P<0.001)。在低磷条件(0AP、1/8AP和1/4AP)下,Pn在7—9月迅速提高,最大Pn显著大于高磷处理(表4,P<0.05);而高磷条件(1/2AP、CK、2AP和4AP)下,Pn从6月开始迅速提高,最大Pn持续时间更长。高磷处理(2AP、4AP)的Tr和Ci从6月开始迅速升高,7—9月达到最大值,且该生长时间段的Tr和Ci均显著大于低磷处理(0AP、1/8AP和1/4AP)(表4,P<0.05)。Pn、Ci和Tr表现出的这种时间变化可能是马尾松幼苗为适应低磷环境产生的光合适应对策:缩短最大光合作用持续时间,提高光合作用强度,进而减少磷在高强度光合中转换产生的耗能和伤害,提高光合作用磷利用效率[30]。这结果一定程度上符合最低消耗理论(The least-cost hypothesis)[31-32],即当蒸散能力和羧化能力达到平衡时,光合作用就能以总的最低消耗来维持植物的正常活动。Gs在不同磷浓度条件下,均在8—9月达到最高值,与其他生长时间的Gs差异显著(表4);但相同生长时间不同处理间Gs差异不显著,表明气孔调节的蒸散和气体进出过程达到对应磷浓度条件下的平衡状态。在低磷条件下,马尾松幼苗可能采取集中高强度的产能方式减少长期高强度转换带来的能量损失,产生了对低磷环境的适应[2,26];当磷含量相对充足时,植株能够获取更多的可利用磷,保证自身长时间有效进行光合作用,促进生长。
-
光合作用生产的化合物为植物生长提供能量,在幼苗期,高生长是植物生长的重要表现形式[12]。简尊吉等[13]对马尾松主要分布区土壤养分进行调查,马尾松林严重缺磷(0.02~78.65 mg·kg−1)。在整个苗期,马尾松幼苗株高相对生长速率受基质磷浓度影响显著,且随磷浓度增加相对生长速率线性增加(表2,图2,P<0.05),表明在0~9.299 mg·kg−1 有效磷浓度范围内,磷限制马尾松幼苗的生长。这一结果与Huang等[15]一致,符合并验证了生长在热带亚热带高度风化土壤地区的森林受到磷限制的假说[2,5,7,33]。
然而,在生长季的不同生长时间段,不同磷浓度处理条件下幼苗高生长对磷的响应存在差异(图1,P<0.01)。我们发现5—6月是高的迅速生长阶段,高磷处理组幼苗的RHI显著大于低磷处理组,7—8月高生长减缓,到9—10月,高磷处理组的RHI9-10与低磷组无显著差异(图1)。这表明磷对马尾松幼苗高生长的促进作用具有时效性,随时间的延长以及生长季的发展,效果减弱。马尾松幼苗株高RGR随磷浓度增加而增加(图2),这主要是由高磷处理组幼苗在5—6月高生长量最大导致。黄盛怡等[11]对马尾松容器幼苗进行磷添加发现高生长差异显著,但也有研究表明不同浓度磷添加导致的马尾松高生长差异不显著[12]。本研究结果与前者一致。
-
光合作用是植物合成促进自身生长所需要的碳水化合物的重要生化过程[3]。本研究发现RHI与Pn和Ci正相关(表5,P<0.05)。胞间CO2是植物进行光合作用的直接原料,Ci越大,光合过程的原料供给越充足;Pn越高,就能产生更多的碳水化合物,促进株高生长[34]。在Crous等[3]对细叶桉(Eucalyptus tereticornis Smith)进行的为期3年的施肥试验中发现施磷显著促进了干的径向生长,Jiang等[35]的研究也表明施磷增强了植物的光合作用。但光合作用的增强并不一定伴随着生物量的增加,磷的增加还可能会增强呼吸作用,增加了消耗[35];在低磷条件下,光合作用提高并不一定导致生产力的提高[36-37]。本研究中磷浓度显著影响了马尾松幼苗的光合产能过程,但在生长表现上没有显著差异(表5)。这可能是因为在低磷条件下,光合作用生产的有机物可能优先分配到根系增加吸收面积[17]或者分配到茎干作为储能物质[3]。本研究试验控制时间为7个月,较短的观测时间也可能是不同磷浓度条件下幼苗生长没有表现出显著差异的原因。
-
本研究以两年生马尾松幼苗为对象,设置不同磷浓度梯度,探究马尾松幼苗高生长和光合特征对磷的响应。结果表明,马尾松幼苗的光合参数受到不同磷浓度、生长时间及其交互作用的显著影响,低磷处理的马尾松幼苗的Pn在整个生长季表现先增高后下降趋势。不同磷浓度对马尾松幼苗株高的相对生长增量影响显著,株高相对生长速率随磷浓度升高而提高。Pn和Ci与RHI显著正相关。马尾松幼苗在低磷环境下通过调节光合作用和高生长动态变化,对低磷环境产生了一定适应机制。
Variations on the Height Growth and Photosynthetic Characteristics of Pinus massoniana Lamb. Seedlings along a Soil Phosphorus Content Gradient
- Received Date: 2022-05-16
- Accepted Date: 2022-07-27
- Available Online: 2023-04-20
Abstract:
 DownLoad:
DownLoad: