



-
温度是限制植物分布和生长发育的主要环境因子之一[1]。虽然全球变暖是大趋势,但近年极端高、低温频发,一次短时极限低温即致一些喜温植物死亡[2]。温度低于14 ℃时,热起源喜温植物易遭受寒害影响,8 ~ 12 ℃是其寒害临界低温[3]。光合作用是植物物质和能量生产的主要来源,因此,光合低温响应和适应的生理生态机制一直是植物寒害研究中的核心问题[4]。热起源的C4植物普遍比中温的C3植物更易受寒害危害[5]。喜温植物光合作用的光能转化分配、电子传递、光合磷酸化及碳同化进程各环节均易受寒害胁迫[6-7]。
红树林植物是生长在亚热带、热带河流入海口或海岸潮间带的木本植物群落[8]。低温对红树植物光合作用的影响已有零星报道。如低温降低红树植物秋茄(Kandelia obovata Sheue)和白骨壤(Avicennia marina (Forsk.) Vierh.)叶片光系统反应中心的活性和电子传递速率[9],降低秋茄叶片光合气体交换能力[10]。虽然红树植物主要分布在热带和亚热带的热区,但纬向范围跨度大,从北半球的日本九州本岛海岸(31° 34′ N)至南半球的澳大利亚Westonport海湾(38° 45' S),间断分布着不同种类的红树植物[11-12]。本研究假设不同纬度地区的红树植物遭受寒害时,它们光合作用受害程度与种源地温度条件密切关联,种源地最冷月温度越低,其幼苗适应低温的能力越强,受寒害程度越低。为验证该假设,以典型的广布种红树科植物木榄(Bruguiera gymnorrhiza (L.) Savigny)为材料,分别从日本冲绳、深圳福田、福建云霄3地收集其成熟果实,在我国典型亚热带地区同质园(广西大学校园内)培养成苗,监测比较它们在寒害低温胁迫前后的光合作用光反应和碳反应特性的差别。研究结果将为喜温植物光合低温寒害生理生态研究提供新资料,同时为红树植物的生态保育、生态恢复和生态规划提供理论参考。
-
于2016年6—9月,从日本冲绳(26°39'19.3″ N,128°00'39.0″ E)、深圳福田(22°31'29.0" N,114°00'34.4" E)、福建云霄(23°51'32.3" N,117°30'23.1" E)3地收集木榄成熟果实,经消毒后播种于半径约为10 cm、高约30 cm的聚乙烯塑料袋中(表1)。每袋装3 kg干燥的天然红树林淤泥,播种子1粒,出苗后培养于广西大学林学院温室大棚内(同质园)(22°50'44″ N,108°17'20″ E)。树苗培育期间,每周浇施1次0.5 mol·L−1 Hoagland’s的培养液和0.2 mol·L−1的NaCl溶液,每天定时顶喷适量淡水,使袋子内土壤始终保持湿润。
种源地
Region材料编号
Material number纬度(N)
Latitude经度(E)
longitude最冷月平均温度
Average temperature of
the coldest month/℃最热月平均温度
Average temperature of
the hottest month/℃福田Futian FT 22°31'29.0" 114°00'34.4" 14.1 28.2 云霄Yunxiao YX 23°51'32.3" 117°30'23.1" 13.3 28.2 冲绳Okinawa UR 26°39'19.3" 128°00'39.0" 16.1 27.9 注:气象数据来源于 http://www.worldclim.org.cn 。
Note: Meteorological data fromhttp://www.worldclim.org.cn .Table 1. Geographic locations and climatic characteristics of Bruguiera gymnorrhiza
-
于2022年3月,选取生长旺盛、长势均匀的3类木榄苗各3株,从温室大棚移至室外处理3 d,于第4天测定叶绿素荧光特性和光合气体交换特性。然后将这些树苗移入多级智能编程光照培养箱(宁波江南仪器厂)培养15 d。箱内温度条件设置为20 ℃ / 20 ℃(昼/夜);光强为200 μmol·m−2·s−1,光周期为12 h / 12 h(昼/夜),相对湿度保持在60%左右。第16 天将温度从20 ℃/20 ℃逐渐降至10 ℃/10 ℃,于此温度下进行低温处理。低温处理期的第4天,再次测定叶绿素荧光特性和光合气体交换特性。
-
选取植株枝顶倒数第2叶,用Dual-PAM-100测量系统(Walz,德国)于凌晨5:00测定受暗适应处理叶的光系统Ⅱ(PSⅡ)最大光能转换效率(Fv/Fm)和光系统Ⅰ(PSI)最大光能转换效率(Pm)。上午7:00用1 000 μmol·m−2·s−1光化光测定经30 min暗适应的叶片荧光动力学曲线,达到稳态后记录PSI有效光化学量子产额(Y(I))、PSI供体侧限制(Y(ND))、PSI受体侧限制(Y(NA))、PSII有效光化学量子产额(Y(II))、调节性能量耗散(Y(NPQ))和非调节性能量耗散(Y(NO))等参数。
-
测完叶绿素荧光特性的叶片立即使用LI-6800光合系统仪(Li-Cor,美国)测定光合气体交换参数。测定条件中流速为500 µmol·m−2·s−1,用小钢瓶控制CO2浓度为400 μmol·mol−1,设置相对湿度为60%,叶室温度为10 ℃。先在1 500 µmol·m−2·s−1光强下诱导,到达稳态后记录净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、叶外CO2 浓度(Co)等指标参数。计算瞬时水分利用效率(WUE)(
$ WUE=Pn/Tr $ )、内禀水分利用效率(WUEi)($ WUEi=Pn/Gs $ )和气孔限制值(LS=1-Ci/Co)。 -
使用Microsoft Excel 2021软件进行数据整理。使用SPSS 26.0软件判断数据属于正态分布,满足方差齐次要求,在此基础上作温度、种源地双因素的方差分析和多重比较,用Duncan检验法进行显著性检验(P < 0.05),采用皮尔森法作相关性分析。采用Origin 2021作图。
-
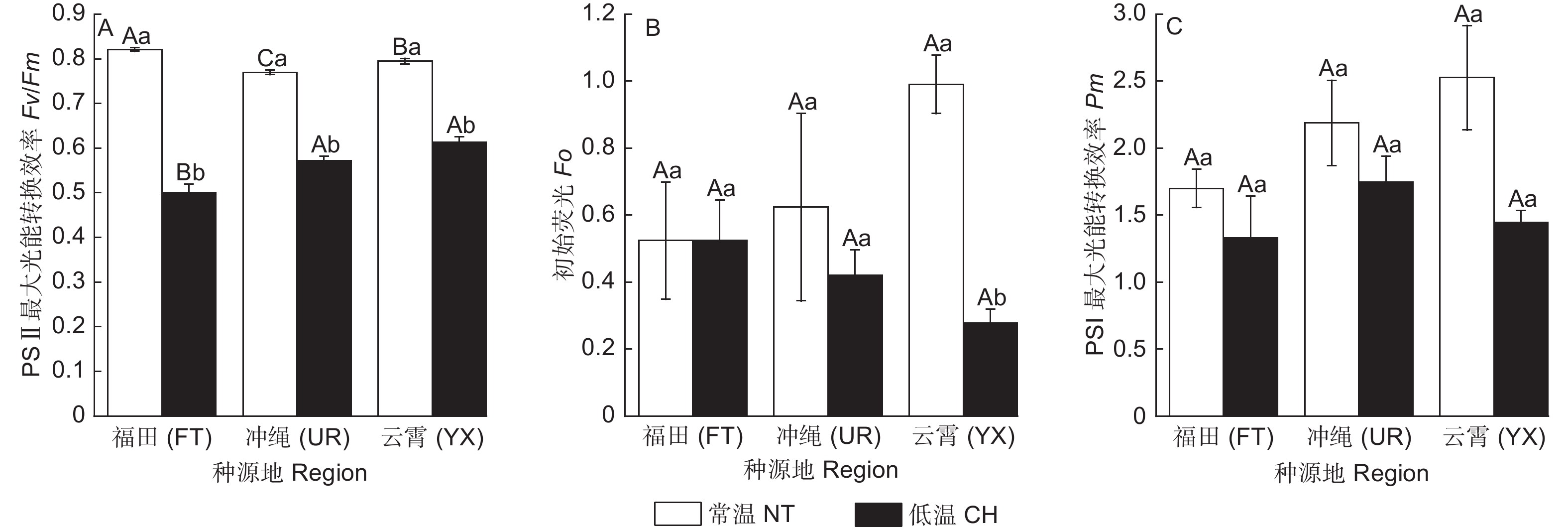
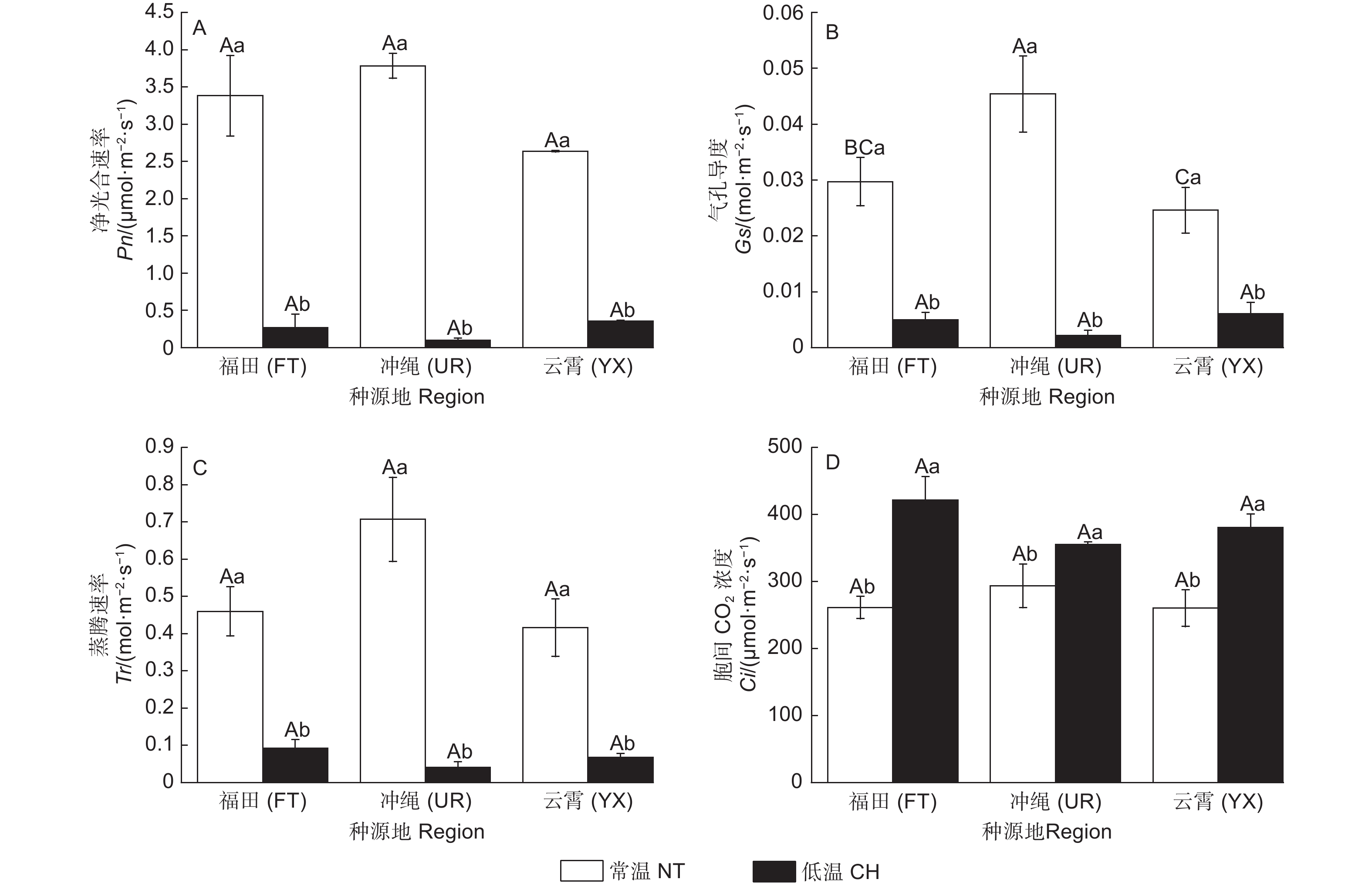
和常温对照相比,低温处理后3类木榄(福田(FT)、冲绳(UR)、云霄(YX))Fv/Fm值分别降低了39.04%、25.69%、22.83%,其中,FT的Fv/Fm值降幅最大(图1A);YX的Fo显著下降,其他2类无显著差异(图1B);FT、UR、YX的Pm值分别降低了21.80%、20.19%、42.84%,但无显著变化(图1 C)。
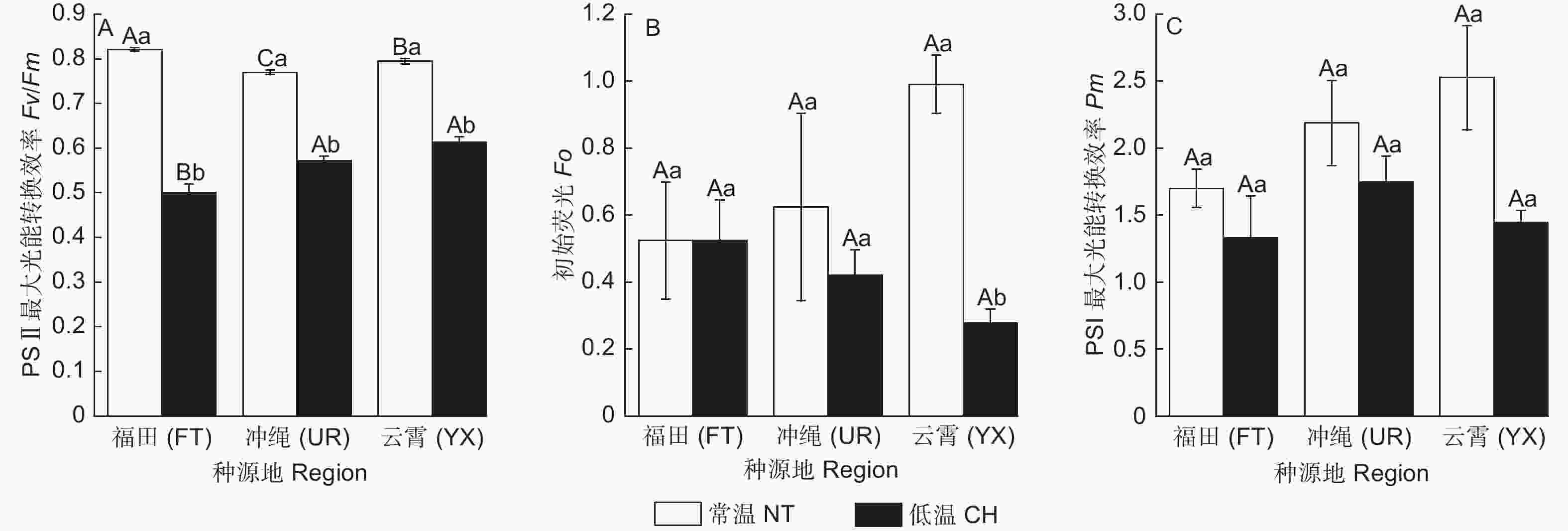
Figure 1. Chlorophyll fluorescence characteristic parameters of different regions of Bruguiera gymnorrhiza at normal temperature and chill temperature treatment
-
图2表明:经低温后,3类木榄的Y(NO)占比均有升高;FT的Y(II)占比升高,而其他2类木榄的Y(II)均降低;YX的Y(NPQ)占比升高,而其他2类木榄的Y(NPQ)均降低。3类木榄的Y(I)占比均降低;FT的Y(ND)占比降低,其他2类木榄的Y(ND)均升高;FT的Y(NA)占比升高,其他2类木榄的Y(NA)均降低。
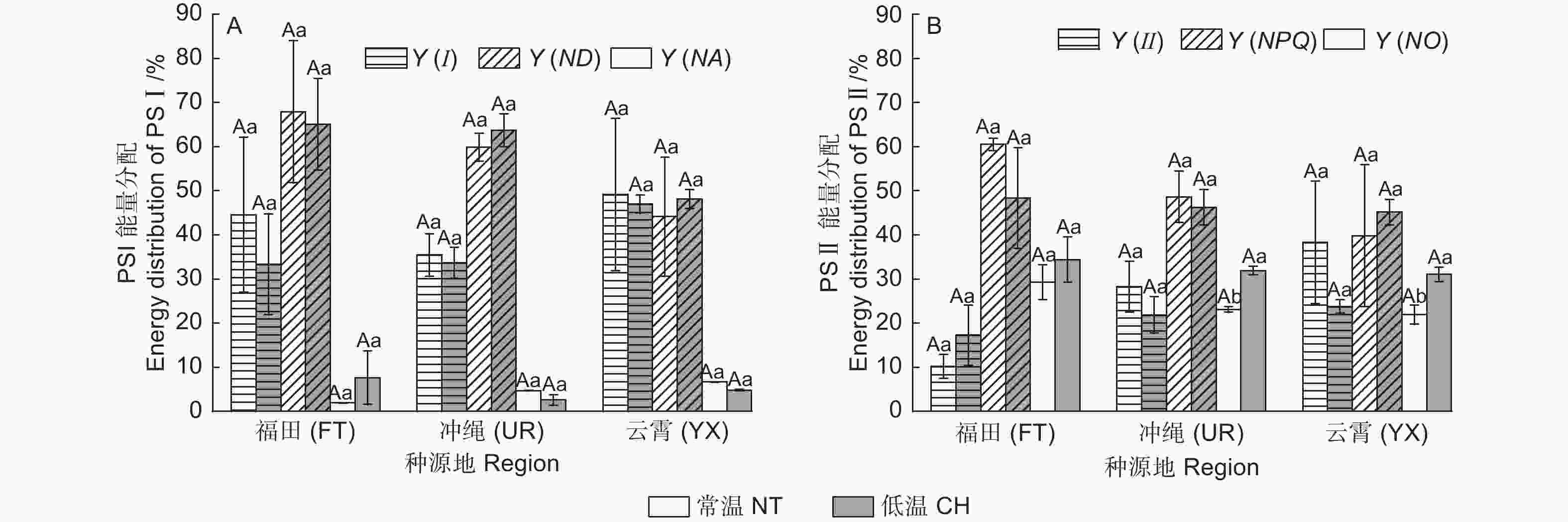
Figure 2. Energy distribution of normal temperature and chill temperature treatment of different regions of B. gymnorrhiza
-
常温下UR的Pn值最高,为3.784 µmol·m−2·s−1。经低温后,FT、UR、YX的Pn值分别下降了92.12%、97.50%、86.44%(图3A)。
Figure 3. Photosynthetic gas exchange characteristic parameters of different regions of B. gymnorrhiza were treated under normal and chill temperature
-
Gs的表现和Pn类似,常温中UR的Gs值(0.045 4 mol·m−2·s−1)显著高于其它2类木榄。经低温处理后,FT、UR、YX的Gs值分别降低了83.12%、95.17%、75.02%(图3B)。
-
常温下3类木榄的Tr无显著差异,经低温后,FT、UR、YX的Tr值分别降低了79.99%、94.16%、83.89%,其中,UR的Tr值低至0.041 mmol·m−2·s−1,气孔蒸腾近乎停止(图3C)。
-
常温时,3类木榄的Ci值无显著差异。经低温后Ci均显著上升,FT、UR、YX分别上升60.97%、24.1%、48.62%(图3D)。
-
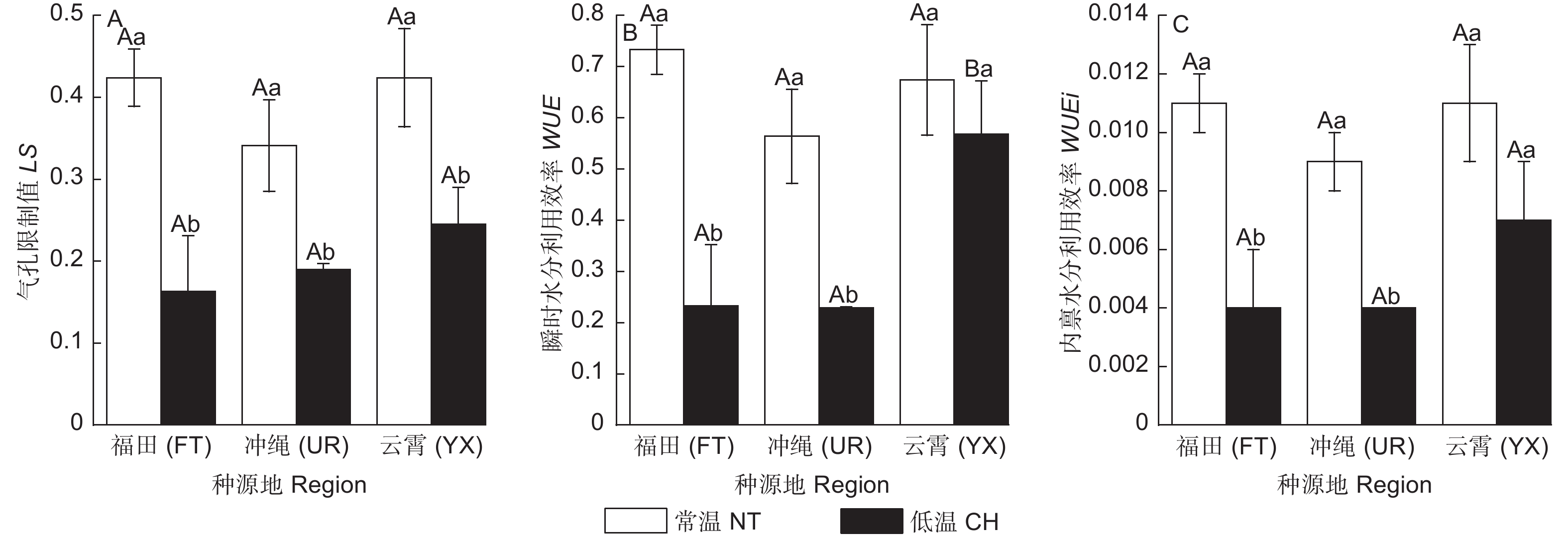
常温下3类木榄的LS值无显著差异,经低温后,FT、UR、YX的LS分别显著降低63.55%、41.88%、42.45%(图4A)。

Figure 4. LS and water use efficiency of different regions of B. gymnorrhiza under normal and chill temperature
-
常温下3类木榄的瞬时水分利用效率(WUE)无显著差异;经低温后,FT和UR的WUE分别降低了68.23%、59.38%,而YX降幅较小(图4B);内禀水分利用效率(WUEi)变化趋势和WUE相似,经低温后,FT、UR的WUEi分别显著降低了62.54%、49.06%,而YX无显著变化(图4C)。
-
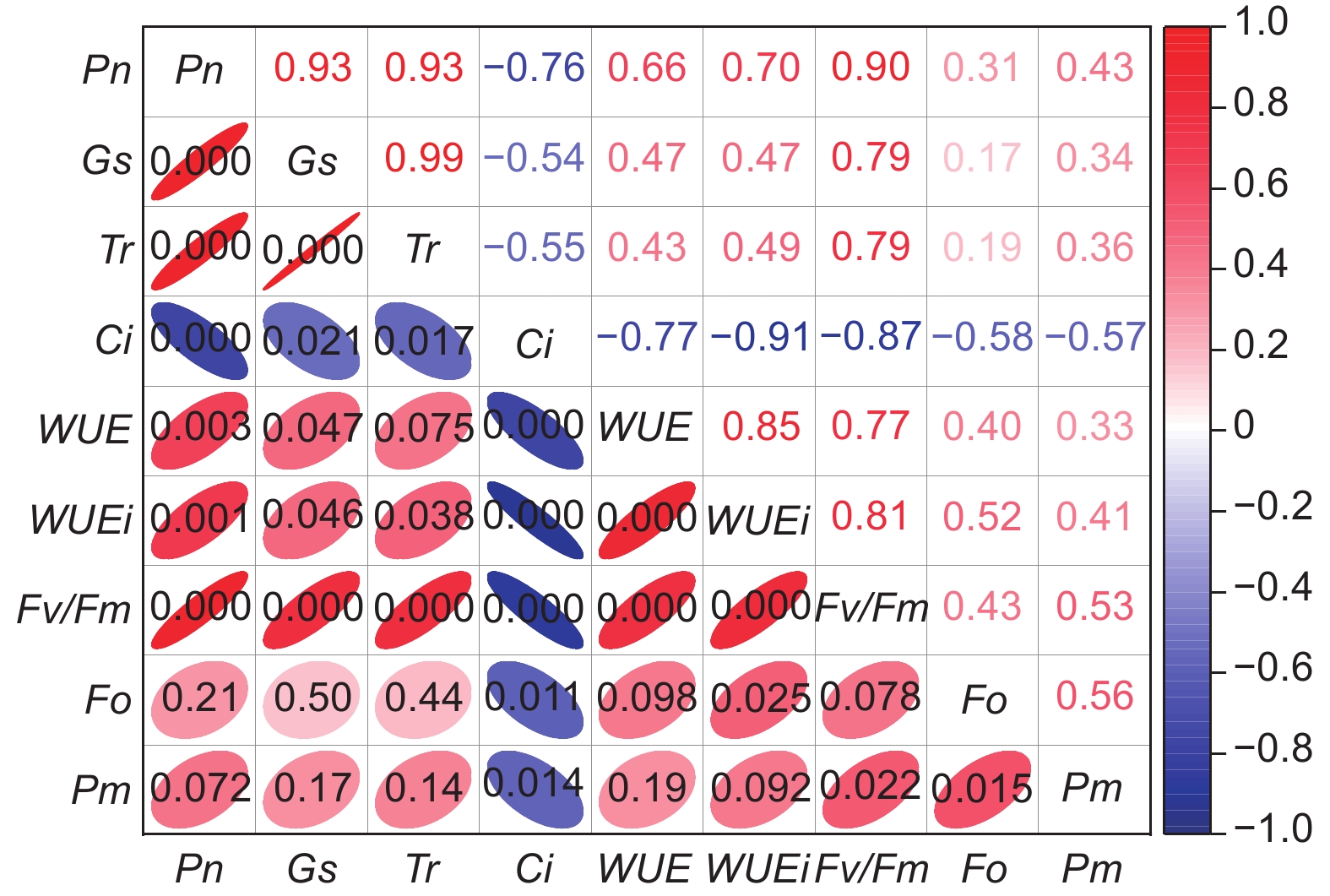
图5表明:经低温后,3类木榄的Fv/Fm和Pm间呈显著正相关;Ci与Pn、Gs、Tr间呈显著负相关;Pn与Fv/Fm、Gs与Fv/Fm及Tr与Fv/Fm间呈显著正相关,Ci与Fv/Fm、Pm间呈显著负相关。

Figure 5. Correlation between the photosynthetic gas exchange characteristics of B. gymnorrhiza and chlorophyll fluorescence characteristics before and after chill stress
-
图6A表明:最冷月平均温度分别与3类木榄的Pn呈显著负相关;图6B表明:最热月平均温度与最冷月平均温度差值与木榄的Pn差值呈显著负相关。
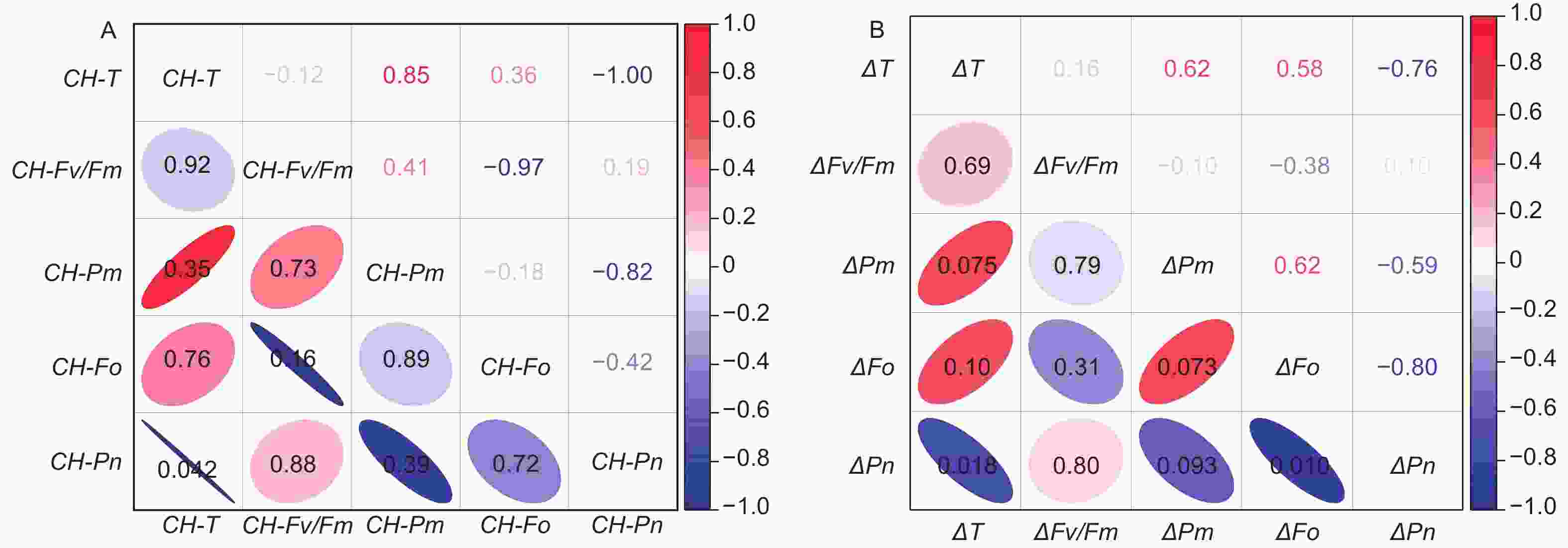
Figure 6. Correlation between the temperature of the B. gymnorrhiza regions and photosynthetic parameters before and after chilling stress
-
本研究发现,即使只遭受到短时寒害冲击,3类木榄凌晨Pm值降低,Fv/Fm值和上午Pn值显著降低,表现出明显的低温光抑制(图1、3)。叶绿素荧光特性通常作为植物进行光合作用生理过程的内在探头[13],可以快速反映植物对低温的响应。有研究认为,Fv/Fm不仅能够反映出植物的潜在光合能力,还是体现植物遭外界胁迫程度的可靠指标[14],本研究结果支持这一观点。综合2个光系统的监测结果,短时低温对3类木榄的光合机构有冲击,但没有形成毁灭性伤害,尤其是PSI低温前后始终维持相对稳定。Tjus等[15]研究发现,在4 ℃温度条件下,PSII的主要电子受体极度变少,电子从PSII转移到PSI过程受阻,产生对PSII的光抑制作用,可视为保护PSI中心避免遭受光损伤的机理机制。郭菊兰等[16]研究表明,严酷低温对红树植物秋茄幼苗叶片的光合机构造成较严重的损伤,可能是因为位于PSII反应中心的D1蛋白受损,从而导致其在低温环境条件下的修复被限制,D1蛋白的降解途径受到阻碍,从而导致新D1蛋白的合成被阻止[17]。本研究进一步比较不同种源地木榄光合作用对低温的响应,发现木榄遭受短时低温冲击后,其碳同化能力与种源地最冷月平均温及最热月平均温和最冷月平均温的差值线性相关(图6),显然,红树植物的光合机构受害程度与低温的胁迫程度有关,且直接受种源地温度条件的影响,该研究结果支持前文的假设。由于研究地点较少,该假说是否适合所有地点的红树植物有待进一步验证。
-
本研究发现,根据Kramer等[18]研究模型,经寒害处理后3类木榄PSI通过光化学途径耗能比例均下降,而UR、YX通过热耗散耗能的比例则增加(图2),表明低温通过影响光系统活性改变了光能的分配。低温作用下,木榄叶片吸收的光能用于光化学反应占比降低的直接后果是易在光系统内形成过剩光能。热耗散途径没能消耗掉所有过剩光能,叶绿体内存留的过剩光能引发了低温光抑制,表现为凌晨Pm的降低(图1)。需要注意的是,FT叶片的Y(NA)较常温增加了2倍,UR和YX叶片的则只下降了1倍,显然FT的PSI受害最重。这些研究结果还表明,3类木榄PSII反应中心对低温的敏感度有差异,对能量的微调也有不同,造成这种结果的原因究竟是结构的差异, 如PSII捕光蛋白或中心蛋白构象和组成的变化[19],还是生理代谢活动因素,如叶黄素循环、光呼吸耗散光能[20]等的差异引起,尚需进一步研究。
经寒害处理后,3类木榄PSII均表现出明显的低温光抑制,但PSII的光能分配则较PSI相对更复杂。FT叶片的Y(II)占比经寒害处理后不降反升,表现出相对较强的抗寒能力,YX叶片的Y(II)比例则大幅下降,主要通过增加Y(NO)的占比维持能量平衡,显然不同于PSI的表现[21]。有观点认为,PSI较PSII更稳定[22],可能原因是,从光合机构电子传递体的排布看,PSII在前,而PSI在后,PSII对PSI有保护作用[23];PSII破坏后易修复,而PSI破坏后则难修复[24]。这些结果表明,木榄光系统可以通过多种机制灵活应对寒害胁迫。
-
本研究结果显示,3类木榄经寒害后碳同化能力均显著下降(图3、4)。逆境胁迫下,植物光合叶片碳同化下降既有气孔因素,也有非气孔因素的影响[25-26]。依据Farquhar等[27]研究理论,当植物的Pn、Gs值下降,而Ci值升高时,碳同化速率下降主要受非气孔因素的影响,相反则是气孔因素起主导作用。本研究中,寒害条件下3类木榄叶片碳同化速率的下降均是非气孔因素起主导作用。有报道指出,低温环境条件下,碳同化速率的非气孔限制主要是由RuBP活性下降或者RuBP合成速率降低所致[28],Rubisco活性减弱,导致CO2与RuBP的结合被限制,进而对光合作用产生影响[28]。此外,非气孔限制条件下,ATP合酶活性也会减弱,通过光合磷酸化合成ATP的途径受阻[29],反馈限制细胞固定CO2,从而导致CO2在细胞间积累。
寒害处理后,随着Pn值的降低,3类木榄叶片的Gs、Tr值也直线下降,进一步影响叶片的水分利用效率(图3)。水分利用效率是衡量植物应对逆境胁迫能力的重要生理生态指标[30-31]。本研究中,经寒害低温处理后,FT和UR水分利用效率显著下降,而YX的表现相对稳定,这主要与其Pn值和Tr值的降幅较小有关,该结果进一步证实了YX光合作用有相对较强的耐低温寒害胁迫能力。Lin等[32]研究结果表明,低温胁迫期间,白骨壤幼苗的Pn和Gs间存在显著正相关。有研究指出,低温胁迫进程中,Pn与Gs的同步变化,有助于植物叶肉细胞以相同的方式保持细胞气孔导度、叶肉细胞导度,用以维持相对稳定的叶肉细胞间水分和CO2分压[33],进而维持相对稳定的水分利用效率。
-
综合分析3类木榄的光、碳反应参数,二者间相关性明显(图5)。植物叶绿体中叶绿素分子吸收的光能通过ETR途径以ATP和NADPH方式储藏,这二者随后用于CO2的同化过程。在低温寒害胁迫下,木榄的PSII反应中心受到光抑制或者光损伤时,叶绿体内光化学反应速率下降,电子传递受到受阻或改变方式,使得ATP和NADPH的转化效率降低,无法为碳同化提供充足的同化力,进一步使得CO2的同化效率下降[34];另一方面,低温胁迫下,非气孔因素进一步削弱碳同化活性,助长了碳同化效率的下降[28],而碳同化效率的大幅下降反馈到叶绿体光合反应中心,引发对ATP和NADPH需求量降低,最终导致光反应能力的下降。
-
本研究认为,3类木榄光合作用的光、碳反应阶段均对低温敏感。总体看,三者光反应阶段的差异较碳反应阶段更明显,尤其PSII响应策略表现更多样。FT叶片PSI较其他二者受寒更明显,而YX叶片PSII受寒害相对较重。非气孔因素是低温限制3类木榄碳反应的主要方式,综合碳、水调节结果,YX碳反应受低温冲击最小,这与其种源地的气温条件密切关联。研究结果为喜温植物的光合低温响应、适应生理生态机制研究增加了新资料,为红树植物生态修复和国家红树生境建设提供了理论参考。
Response of Photosynthesis to Chill Temperature in Different Regions of Bruguiera gymnorrhiza
- Received Date: 2022-09-17
- Accepted Date: 2022-12-03
- Available Online: 2023-04-20
Abstract:




 DownLoad:
DownLoad: