



-
植物表型是物种各种形态特征的组合,是基因与环境共同调控的结果[1],表型变异是植物适应不同环境条件的表现,因此,其在适应环境与进化上具有重要意义[2]。遗传多样性是群体间和群体内个体间遗传变异的综合[3],表型多样性主要研究群体在不同生境条件下的表型变异规律及分布格局,是遗传多样性的重要组成部分[4]。植物经过长期的自然选择与环境压力,其生长发育水平往往会发生不可逆的变化,进而引起形态特征的变异产生新的表型[5]。通过表型性状多样性的研究可以了解植物对生态环境的响应机制和遗传变异规律[6-7],尤其在物种种质资源的收集、保存、评价与利用研究等方面极为重要,保持丰富的变异对于天然群体十分有利。
大别山山核桃(Carya dabieshanensis M. C. Liu et Z. J. Li)是胡桃科(Juglandaceae)山核桃属(Carya Nutt.)的多年生落叶乔木,是一种重要木本油料树种,同时兼具较高的经济和生态价值[8-9],主要分布于我国安徽、湖北、河南三省交界的大别山山区(29~31° N,115~116° E,海拔600~900 m)。大别山山核桃是刘茂春等[10]在进行植物调查时首次在金寨县燕子河镇发现,认为大别山山核桃是另一个不同于山核桃(C. cathayensis Sarg.)的新种,Youjun, et al [11]也支持此观点,并通过基因组测序得出大别山山核桃与山核桃的亲缘关系最近。与山核桃相比,大别山山核桃具有籽大、壳薄、出仁率和出油率较高的优点[12-15],其果仁中所含的丰富的不饱和脂肪酸、维生素、氨基酸等营养成分对于健脑、抗肿瘤和预防心脑血管疾病具有重要作用[16],有很好的开发利用价值。
目前,对大别山山核桃的研究主要涉及到果实营养品质、繁殖栽培方法、生产开发以及新品种选育等方面[17-20],而大别山山核桃种群遗传多样性的调查是开展各项研究的基础,近年来对其调查范围及取样数量也不断增多。王正加等[21]选取3个天然群体利用RAPD分子标记技术对大别山山核桃的种群遗传多样性进行调查,发现其种群内有较丰富的变异;夏国华等[15]选取了9个种群对其表型及脂肪酸组分进行分析,认为坚果表型性状和脂肪酸组分遗传变异丰富;庞赛[22]选取13个种群,对坚果经济性状和营养物质含量进行分析,认为大别山山核桃具有较好的遗传多态性,存在着丰富的变异。基于前人对大别山山核桃的研究,本文选取了19个天然种群的大别山山核桃,旨在更系统全面的揭示大别山山核桃表型性状与地理分布的关系及其变异规律,为后续的种质资源收集、优选、改良提供理论支持,为大别山山核桃的开发利用奠定基础。
HTML
-
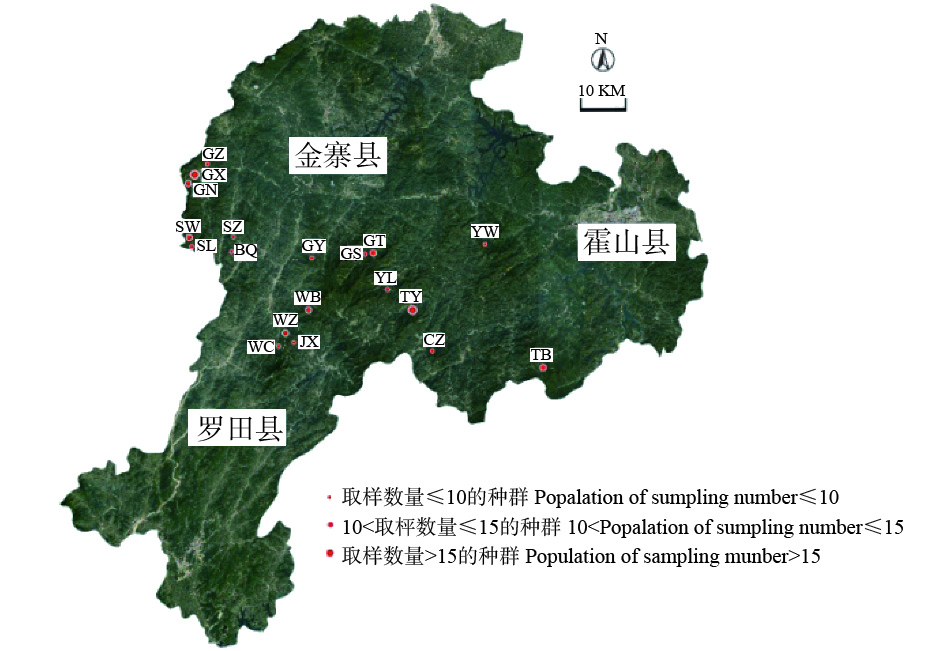
在大别山山核桃自然成熟期间,于2017年9月上中旬在其主要分布区进行取样,各取样点分别位于安徽省六安市金寨县吴家店镇、天堂寨镇、斑竹园镇、燕子河镇、古碑镇、关庙乡、长岭乡、果子园镇、沙河乡和霍山县太阳乡及湖北省罗田县九资河镇,共选取198个单株。根据各种群的大小确定适量的采样植株数,选取树龄、长势相近的健康植株,单株间距50 m以上,对其拍照并用全球卫星系统定位各单株地理位置及海拔,各种群地理位置信息见表1,对应的种群分布见图1。
种群名称
Population names地理位置
Geographical location纬度(N)
Latitude经度(E)
Longitude海拔/m
Elevation取样数量/株
Sampling number吴包WB 安徽省金寨县吴家店镇包畈村 31°14' 115°38' 712 15 吴蔡WC 安徽省金寨县吴家店镇蔡河村张湾组 31°09' 115°35' 687 4 吴竹WZ 安徽省金寨县吴家店镇竹根河村登山组 31°10' 115°36' 739 15 天渔TY 安徽省金寨县天堂寨镇渔潭村 31°15' 115°50' 685 18 斑漆BQ 安徽省金寨县斑竹园镇漆家店村 31°20' 115°28' 761 5 燕龙YL 安徽省金寨县燕子河镇龙马岩前村 31°16' 115°49' 707 5 燕万YW 安徽省金寨县燕子河镇万畈村王家寨组 31°21' 116°02' 620 6 古太GT 安徽省金寨县古碑镇太古村王湾组 31°19' 115°47' 887 15 古宋GS 安徽省金寨县古碑镇宋河村新民组 31°19' 115°46' 624 10 关南GN 安徽省金寨县关庙乡仙桃村南泥沟组 31°28' 115°22' 652 15 关仙GX 安徽省金寨县关庙乡仙桃村老林沟组 31°29' 115°23' 528 25 关周GZ 安徽省金寨县关庙乡仙桃村周院组 31°30' 115°25' 737 5 长撞CZ 安徽省金寨县长岭乡撞畈村 31°8' 115°55' 733 6 果姚GY 安徽省金寨县果子园镇姚冲村 31°19' 115°39' 706 8 沙吴SW 安徽省金寨县沙河乡祝畈村吴冲组 31°21' 115°23' 850 11 沙卢SL 安徽省金寨县沙河乡祝畈村 31°20' 115°23' 804 5 沙枣SZ 安徽省金寨县沙河乡枣林村柏树冲组 31°22' 115°28' 474 5 太白TB 安徽省霍山县太阳乡金竹坪村白马尖组 31°4.2' 115°60' 875 15 九小JX 湖北省罗田县九资河镇小寨林场 31°10' 115°36' 905 10 总计Total 198 Table 1. The situation of geographical location information and sampling in C. dabieshanensis
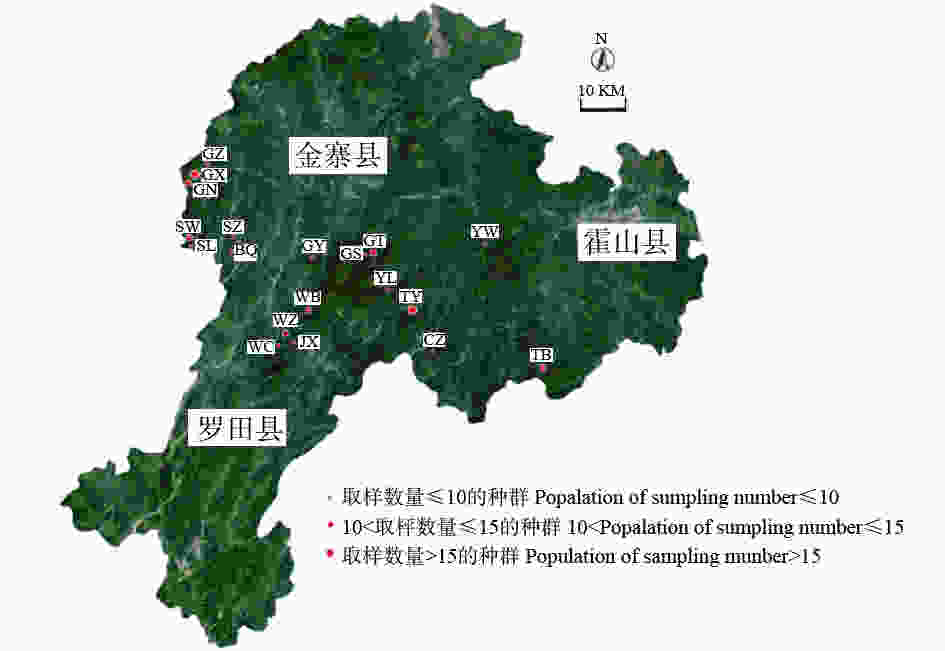
Figure 1. Map of natural population distribution of C.dabieshanensis
分别取果实和叶片两部分样品,每个单株取中下部树冠外围的果实1.5~3.0 kg,称鲜质量后放置于编织网中,保持通风,以防发霉;在植株的4个方向均选取中下部枝条,取各枝条中部成熟的完整复叶共12片,带回室内进行表型性状测定。
-
每单株随机选取30个大小均匀无病害的果实进行表型测定,果实鲜质量与坚果鲜质量均采用10粒一组用电子天平称总质量求平均值的方法得出。果实纵径、果实横径、坚果纵径、坚果横径用游标卡尺(精度0.01 mm)进行测量;蒲壳厚度即外果皮厚度,取山核桃中部两棱之间的外果皮,测定其切面的厚度;敲碎坚果取坚果壳及隔膜,选择其厚度均匀的部分测量果壳厚度、隔膜厚度。果形指数采用以下公式计算:果形指数=坚果纵径/坚果横径。
叶片形态中顶生小叶叶长、宽,侧生小叶叶长、宽,使用直尺测量(精度0.01 cm)。顶生小叶叶形比=顶生小叶叶长/顶生小叶叶宽。侧生小叶叶形比=侧生小叶叶长/侧生小叶叶宽。每个单株随机选取3片大小相似的完整复叶,顶生小叶选取顶部叶片测定,侧生小叶选取第一组右侧的叶片测定。
-
应用单因素方差分析,嵌套设计方差分析等计算各表型性状在种群间和种群内的变异规律及特征;应用相关性分析揭示各表型性状间的相关性,相关系数使用pearson系数。在方差分析的基础上对各表型性状进行多重比较,种群内各表型性状的变异程度为平均值±标准差,离散程度用变异系数(CV)表示。
CV =(标准差/平均值) ×100%
表型分化系数VST=δ2t/s/(δ2t/s + δ2s)× 100%
式中:δ2t/s、δ2s分别为群体间、群体内的方差。
以上数据分析在Spss22.0软件上完成,方差分量的计算在Minitab 17.0软件中完成,非加权配对算数平均法的聚类分析在R3.4.3软件中完成,其他数据处理在Excel2016软件完成。
1.1. 样品采集
1.2. 测定方法
1.3. 数据处理
-
大别山山核桃16个表型性状的方差分析见表2。经F值检验,果实与叶片性状在种群间和种群内差异极显著(P<0.01),说明大别山山核桃在种群间和种群内变异丰富。各表型性状的平均值、标准偏差和多重比较结果(表3)表明:大部分种群间存在显著差异,其中,沙枣(SZ)种群的果实鲜质量、果实横径、坚果横径最大,外果皮、坚果壳最厚;吴包(WB)种群的果形指数最大(1.31),果实形态为椭圆形;古宋(GS)种群果形指数最小(1.07),果实形态为圆形,其坚果纵径最小。吴蔡(WC)种群的坚果壳厚度与隔膜厚度均最小,沙吴(SW)种群的果实鲜质量、果实横径最小,蒲壳也最薄。沙卢(SL)种群的坚果鲜质量、坚果纵径最大,其顶生小叶叶长、叶宽,侧生小叶叶长、叶宽均最小,沙枣(SZ)种群顶、侧生小叶叶形比最小,关周(GZ)种群顶、侧生小叶叶形比最大。
性状
Phenotypic trait均值±标准偏差
Mean ± SD均方
Mean squareF值
F value种群间
Among populations种群内
Within population随机误差
Random errors种群间
Among populations种群内
Within populationFFW/g 15.204±3.437 258.308 9.684 1.454 26.776** 6.662** FHD/mm 30.546±2.388 127.900 4.969 0.454 25.742** 10.934** FVD/mm 31.569±3.512 349.180 4.486 0.494 77.843** 9.088** PT/mm 5.032±1.076 30.494 0.590 0.073 51.704** 8.112** FNW/g 5.426±0.904 17.468 0.635 0.136 27.613** 4.657** NHD/mm 20.646±1.061 16.878 1.569 0.205 10.758** 7.651** NVD/mm 24.371±1.979 103.068 1.977 0.263 52.140** 7.518** NSI 1.183±0.833 0.187 0.003 0.001 74.803** 3.460** ST/mm 1.001±0.117 0.344 0.009 0.001 39.621** 9.843** SMT/mm 0.595±0.109 0.297 0.007 0.001 40.597** 7.879** TLL/mm 16.875±3.029 42.406 11.170 6.549 3.797** 1.705** TLW/mm 4.847±0.967 4.485 1.105 0.673 4.059** 1.643** TLSR 3.526±0.471 1.430 0.212 0.163 6.763** 1.299** LLL/mm 16.703±2.842 25.753 10.860 5.915 2.371** 1.836** LLW/mm 4.606±0.869 3.269 0.930 0.546 3.516** 1.702** LLSR 3.667±0.461 1.126 0.255 0.146 4.422** 1.748** 注:**,极显著相关,P<0.01。FFW:果实鲜质量;FHD:果实横径;FVD:果实纵径;PT:蒲壳厚度;FNW:坚果鲜质量;NHD:坚果横径;NVD:坚果纵径;NSI:果形指数;ST:坚果壳厚度;SMT:隔膜厚度;TLL:顶生小叶叶长;TLW:顶生小叶叶宽;TLSR:顶生小叶叶形比;LLL:侧生小叶叶长;LLW:侧生小叶叶宽;LLSR:侧生小叶叶形比。下同。
Notes: **, Highly significant correlation, P<0.01. FFW: Fresh fruit weight. FHD: Fruit horizontal diameter. FVD: Fruit vertical diamete; PT: Pericarp thickness; FNW: Fresh nut weight; NHD: Nut horizontal diameter; NVD: Nut vertical diameter; NSI: Nut shape index; ST: Shell thickness; SMT: Shell membrane thickness; TLL: Terminal leaflet length; TLW: Terminal leaflet width; TLSR: Terminal leaflet shape ratio; LLL: Lateral leaflet length; LLW: Lateral leaflet width; LLSR: Lateral leaflet shape ratio. The same below.Table 2. Analysis of variance of phenotypic traits among and within populations in C. dabieshanensis
种群
Population表型性状 Phenotypic traits FFW/g FHD/mm FVD/mm PT/mm FNW/g NHD/mm NVD/mm NSI WB 17.89±2.36bcde 31.52±1.47cdef 36.44±1.84a 5.16±0.59efg 6.18±0.48ab 20.11±0.73def 26.33±0.92ab 1.31±0.04a WZ 19.16±1.27ab 33.09±0.83bc 36.53±0.96a 6.47±0.37abc 5.30±0.36cde 20.32±0.44def 25.59±0.59abc 1.26±0.02bc WC 18.13±1.51bcd 32.40±1.35bcd 35.35±1.76ab 6.18±0.41abcd 5.40±0.28bcde 19.93±0.39ef 25.38±0.47bcd 1.28±0.02ab TY 14.21±1.32fg 30.11±1.19efgh 29.12±1.01gh 4.93±0.45fgh 5.03±0.41cdef 20.90±0.53abcde 23.74±0.70efg 1.14±0.02fgh BQ 18.61±2.10abc 34.01±1.22ab 30.36±0.95fg 6.76±0.49ab 5.68±0.46bc 21.24±0.68abcd 22.89±0.66fg 1.08±0.03i YL 17.00±1.99bcdef 31.85±2.20bcde 32.14±1.56cdef 5.63±0.66def 5..54±0.49bcd 20.63±0.89bcdef 25.61±0.59abc 1.24±0.04bc YW 18.52±1.70abc 33.05±1.21bc 31.43±0.76ef 6.19±0.25abcd 5.44±0.58bcde 21.24±0.85abcd 25.02±0.90bcde 1.18±0.01ef GT 12.02±0.93gh 28.84±0.66ghi 26.36±0.56i 4.35±0.25hi 4.68±0.28ef 19.86±0.44ef 21.26±0.42h 1.07±0.02i GS 12.19±1.06gh 29.02±0.68ghi 26.84±0.61i 4.26±0.37hi 4.74±0.22def 19.86±0.42ef 21.17±0.49h 1.07±0.02i GN 11.91±1.95gh 28.04±1.57hi 30.20±1.49fg 4.16±0.36hi 5.12±0.64cdef 20.60±0.83cdef 24.41±0.84cde 1.19±0.03def GX 13.97±2.13fg 29.29±1.62fghi 32.57±1.48cde 3.91±0.63i 6.66±0.57a 21.88±0.85ab 26.43±0.91ab 1.21±0.03cde GZ 14.79±1.36efg 29.88±0.73efgh 33.79±0.92bc 5.14±0.50efg 5.04±0.18cdef 19.47±0.82f 24.14±1.53def 1.24±0.04bcd CZ 15.41±1.69cdef 30.02±0.74efgh 31.60±0.72def 4.82±0.37gh 5.39±0.36bcde 20.41±0.75def 25.01±0.46bcde 1.23±0.04bcd GY 19.79±1.89ab 33.12±1.26bc 36.99±1.00a 6.02±0.30bcd 6.15±0.64ab 21.01±0.75abcde 26.27±1.50ab 1.25±0.04bc SL 14.97±1.04defg 30.38±0.81defg 33.68±0.75bcd 3.91±0.37i 6.81±0.50a 21.79±0.37ab 26.99±0.69a 1.24±0.01bc SW 10.40±1.42h 27.06±1.14i 27.67±1.13hi 3.61±0.32i 4.89±0.41cdef 19.80±0.60ef 22.46±0.49gh 1.14±0.02fg SZ 21.71±3.35a 35.47±1.59a 31.65±0.93def 6.83±0.50a 6.18±0.40ab 21.93±0.65a 23.97±0.69def 1.10±0.01ghi TB 15.44±2.43cdef 31.36±1.54cdef 29.03±1.12gh 5.72±0. 41cde 4.67±0.62ef 20.59±1.13cdef 22.33±0.96gh 1.09±0.03hi JX 14.62±1.33fg 30.27±0.99defgh 33.33±1.28bcde 5.63±0.23def 4.35±0.53f 20.07±0.69def 24.62±0.57cde 1.23±0.03bcde 种群
Population表型性状 Phenotypic traits ST/mm SMT/mm TLL/mm TLW/mm TLSR LLL/mm LLW/mm LLSR WB 0.83±0.03k 0.56±0.03ghi 17.50±2.78ab 5.02±0.83abcd 3.51±0.42bcde 17.14±2.60abc 4.78±0.74abcd 3.61±0.41bcd WZ 0.98±0.06fg 0.74±0.07b 15.24±2.62bc 4.18±0.76cde 3.68±0.39abc 15.51±2.69bc 4.02±0.77de 3.89±0.40ab WC 0.81±0.03k 0.46±0.04l 18.48±3.81a 5.63±0.89a 3.29±0.41cde 18.27±3.10ab 5.22±0.95a 3.52±0.30bcd TY 1.00±0.05efg 0.53±0.03hij 16.44±2.79ab 4.99±0.86abcd 3.31±0.34cde 16.34±2.54abc 4.74±0.76abcde 3.47±0.36bcd BQ 1.12±0.07b 0.62±0.05ef 16.16±2.27abc 5.21±1.00ab 3.15±0.39de 16.31±1.92abc 5.12±0.83ab 3.23±0.37d YL 1.09±0.05bc 0.76±0.06b 19.29±4.04a 5.15±1.37abc 3.87±0.75ab 17.11±3.03abc 4.59±1.05abcde 3.78±0.36abc YW 1.00±0.05efg 0.48±0.04kl 17.69±2.90ab 5.43±0.91ab 3.28±0.33cde 17.23±2.66ab 4.93±0.94abcd 3.57±0.69bcd GT 1.06±0.06cd 0.49±0.07jkl 17.48±2.79ab 4.64±0.85abcde 3.80±0.36ab 16.34±2.78abc 4.24±0.76bcde 3.88±0.51ab GS 0.91±0.03ij 0.48±0.03kl 17.13±2.58ab 4.69±0.92abcde 3.71±0.50abc 15.91±2.27abc 4.38±0.81abcde 3.68±0.40abcd GN 1.01±0.05def 0.60±0.03efg 18.13±2.83ab 5.07±0.77abc 3.59±0.42bcd 17.92±2.75ab 4.89±0.80abcd 3.69±0.40abcd GX 1.05±0.04cde 0.57±0.05fgh 16.65±3.51ab 4.88±1.08abcd 3.45±0.48bcde 17.18±3.42ab 4.74±0.97abcde 3.65±0.34bcd GZ 0.86±0.08k 0.50±0.04ijkl 16.40±2.12ab 4.02±0.56de 4.11±0.46a 16.98±2.38abc 4.13±0.48cde 4.14±0.51a CZ 1.22±0.06a 0.88±0.06a 18.02±3.12ab 5.51±1.09ab 3.31±0.36cde 17.52±3.08ab 5.00±0.93abc 3.54±0.40bcd GY 0.95±0.07ghi 0.58±0.04fgh 19.02±2.76a 5.19±0.99abc 3.71±0.39abc 18.94±2.69a 5.02±0.83abc 3.81±0.41bcd SL 0.97±0.03fgh 0.51±0.03ijk 13.08±2.62c 3.84±0.99e 3.48±0.51bcde 14.08±3.05c 3.85±1.02e 3.72±0.44abc SW 0.98±0.11fg 0.71±0.13bc 15.07±2.59bc 4.79±0.94abcde 3.18±0.44de 15.40±2.53bc 4.70±0.81abcde 3.31±0.41cd SZ 1.26±0.05a 0.64±0.04de 16.38±2.04ab 5.45±1.14ab 3.07±0.39e 16.57±2.21abc 5.18±0.86a 3.22±0.28d TB 1.14±0.08b 0.67±0.06cd 16.94±2.29ab 4.88±0.74abcd 3.51±0.39bcde 16.92±2.26abc 4.54±0.69abcde 3.75±0.33abc JX 0.91±0.04jk 0.57±0.03fgh 16.43±2.42ab 4.59±0.69bcde 3.60±0.37bcd 16.81±3.03abc 4.43±0.73abcde 3.82±0.56ab 注:种群缩写见表1,表型性状缩写见表2注。不同小写字母表示显著性差异(P <0.05)。
Notes:See Table1 for abbreviation of population, and see Table2 notes for abbreviation of phenotypic trait. different lowercase letters indicate significant difference (P <0.05).Table 3. Average value, standard deviation and multiple comparison of phenotypic characters of pecan fruit in C. dabieshanensis
-
19个大别山山核桃种群的表型性状变异范围为1.64%~26.59%,平均变异系数为9.63%(表4)。不同种群间同一表型性状的变异范围不同,同一种群内的表型变异也因不同性状而有差别。
% 表型性状
Phenotypic
traits种群 Populations 种群内
Within
population种群间
Among
populationsWB WZ WC TY BQ YL YW GT GS GN GX GZ CZ GY SL SW SZ TB JX FFW 13.69 7.26 8.30 8.70 12.09 11.27 10.77 9.81 9.85 17.42 16.05 10.43 11.63 9.90 8.36 10.77 16.24 16.03 10.33 11.52 22.44 FHD 4.89 2.79 4.58 4.26 3.69 6.65 4.03 2.79 2.95 5.92 5.75 2.46 3.00 4.03 3.08 4.49 4.98 5.00 3.80 4.17 7.77 FVD 5.26 2.89 6.46 3.84 3.52 4.62 3.11 2.70 2.53 5.19 4.84 2.88 2.75 2.98 3.97 4.60 3.99 4.17 4.16 3.92 11.08 PT 11.49 6.19 8.64 10.09 7.28 12.10 5.68 6.51 9.18 9.58 17.01 9.72 11.04 6.01 10.18 9.87 8.52 8.57 5.47 9.11 21.31 FNW 9.15 6.52 7.69 7.51 9.20 9.63 12.21 9.28 6.43 13.08 9.72 3.86 7.43 11.52 8.23 9.29 7.79 13.62 13.38 9.24 16.71 NHD 4.18 2.50 3.13 2.90 3.36 4.85 4.21 2.67 2.42 4.13 4.26 4.95 4.42 3.80 2.23 3.58 3.17 5.48 4.09 3.70 5.14 NVD 3.78 2.62 2.95 3.16 2.82 3.32 3.44 2.54 2.52 3.68 3.84 7.08 3.00 5.81 3.24 2.65 3.12 4.34 2.99 3.52 8.16 NSI 3.81 2.40 3.49 2.38 2.82 4.42 2.37 2.72 2.20 2.66 2.75 3.35 3.68 3.66 2.65 3.03 1.64 3.01 3.60 2.98 7.02 ST 3.43 5.81 3.62 4.78 5.85 4.17 4.53 5.78 3.76 5.22 3.52 9.52 4.66 7.70 3.10 11.33 4.01 7.42 4.69 5.42 11.57 SMT 6.14 9.09 9.63 4.98 8.43 7.59 7.91 13.71 6.61 5.26 8.22 8.64 7.10 6.08 4.94 18.83 5.51 8.30 5.13 8.01 18.31 TLL 15.91 17.20 20.63 16.97 14.03 20.95 16.36 15.95 15.07 15.59 21.08 12.94 17.34 14.51 20.06 17.21 12.46 13.52 14.72 16.45 17.95 TLW 16.51 18.18 15.74 17.22 19.18 26.52 16.81 18.27 19.65 15.17 22.23 13.86 19.83 19.03 25.73 19.53 20.98 15.08 14.99 18.66 19.94 TLSR 12.05 10.55 12.57 10.13 12.52 19.29 9.91 9.55 13.55 11.58 13.90 11.22 10.99 10.53 14.63 13.92 12.84 11.25 10.32 12.17 13.35 LLL 15.14 17.34 16.97 15.57 11.78 17.69 15.42 17.02 14.26 15.33 19.92 14.01 17.58 14.18 21.68 16.41 13.33 13.33 18.00 16.05 17.01 LLW 15.42 19.04 18.24 16.13 16.29 22.94 18.98 17.91 18.46 16.35 20.39 11.75 18.57 16.57 26.59 17.21 16.61 15.27 16.40 17.85 18.87 LLSR 11.33 10.41 8.45 10.25 11.57 9.50 19.21 13.18 10.93 10.94 9.19 12.35 11.28 10.73 11.80 12.50 8.76 8.74 14.56 11.35 12.57 均值Mean 9.51 8.80 9.44 8.68 9.03 11.59 9.68 9.40 8.77 9.82 11.42 8.69 9.64 9.19 10.65 10.95 9.00 9.57 9.16 9.63 14.33 注:种群缩写见表1,表型性状缩写见表2注。
Notes: See Table 1 for abbreviation of population, and see Table 2 notes for abbreviation of phenotypic trait.Table 4. Average coefficient of variation of phenotypic characters in natural populations of pecan in C. dabieshanensis
在所有表型性状中,以顶生小叶叶宽的平均变异系数最高(18.66%),其次是侧生小叶叶宽(17.85%),果形指数的平均变异系数最小(2.98%)。16种表型性状的变异情况不同,但总体上叶片表型的变异程度高于果实表型。对于不同的种群,平均变异系数最大的是YL种群(11.59%),其次为GX种群(11.42%),而TY种群的平均变异系数最小(8.68%),这表明YL、GX种群的表型变异更丰富。综合比较果实表型与叶片表型在种群内和种群间的平均变异系数,种群内的平均变异系数均小于种群间。分开来看,果实性状在种群内的变异系数(6.16%)小于叶片性状的变异系数(15.42%),在种群间的变异系数(12.95%)也小于叶片表型的变异系数(16.62%)。表明大别山山核桃的叶片性状较果实性状其变异程度更大,果实与叶片性状在种群间的变异均大于种群内。
-
果实与叶片性状的相关性分析(表5)表明:性状间达极显著相关的有60对,达显著相关的有11对。在果实表型性状中,果实鲜质量,果实纵径,果实横径这3个性状与其他果实表型性状基本上都存在极显著正相关,其中,果实横径与果实鲜质量、蒲壳厚度,果形指数与果实纵径、坚果纵径这6组性状的相关性极高,pearson相关系数均在0.75以上。这表明,果实横径的增加会引起果实鲜质量和蒲壳厚度的增加;而果实纵径与坚果纵径的变化也与果形指数的大小紧密相关。对于叶片的表型性状,所有叶片性状相互间均存在显著或极显著相关,其中,顶生小叶叶长与顶生小叶叶宽、侧生小叶叶长,顶生小叶叶宽与侧生小叶叶长、侧生小叶叶宽,侧生小叶叶长与侧生小叶叶宽这5组性状的相关性极高,pearson相关系数系数均大于0.7。在果实表型与叶片表型相关的区域,果实鲜质量、果实横径、蒲壳厚度均与顶生小叶叶长、叶宽,侧生小叶叶长这3个性状存在极显著正相关,但相关系数较低。
FFW FHD FVD PT FNW NHD NVD NSI ST SMT TLL TLW TLSR LLL LLW LLSR FFW 1 FHD 0.860** 1 FVD 0.652** 0.591** 1 PT 0.753** 0.861** 0.474** 1 FNW 0.471** 0.316** 0.490** −0.033 1 NHD 0.261** 0.330** 0.219** 0.059 0.581** 1 NVD 0.403** 0.326** 0.788** 0.116** 0.654** 0.536** 1 NSI 0.285** 0.145** 0.765** 0.102* 0.330** −0.117** 0.773** 1 ST 0.106* 0.174** −0.277** 0.114** 0.100* 0.384** −0.120** −0.422** 1 SMT 0.152** 0.139** 0.157** 0.217** −0.059 −0.030 0.086* 0.133** 0.439** 1 TLL 0.168** 0.161** 0.110* 0.149** 0.034 0.047 0.015 −0.026 −0.026 −0.036 1 TLW 0.115** 0.120** 0.052 0.114** 0.032 0.093* 0.008 −0.064 −0.008 −0.061 0.755** 1 TLSR 0.060 0.042 0.078 0.038 0.002 −0.081 0.011 0.069 −0.021 0.041 0.205** −0.471** 1 LLL 0.152** 0.147** 0.106* 0.170** −0.028 0.004 0.002 −0.002 −0.045 −0.006 0.853** 0.706** 0.090* 1 LLW 0.077 0.086* 0.070 0.096* 0.004 0.039 0.028 0.002 −0.073 −0.057 0.667** 0.791** −0.279** 0.753** 1 LLSR 0.088* 0.062 0.030 0.081 −0.043 −0.067 −0.048 −0.005 0.063 0.073 0.148** −0.222** 0.541** 0.217** −0.462** 1 注:**,P<0.01; *,P<0.05。
Notes: **,P<0.01; *,P<0.05.Table 5. Correlation analysis of fruit and leaf characters of pecan in C. dabieshanensis
-
大别山山核桃果实与叶片性状的表型分化系数和方差分量(表6)表明:表型分化系数为25.23%~89.43%,平均为66.25%,即种群内的表型分化系数为33.75%。这表明种群间的遗传分化远大于种群内,大别山山核桃表型变异的主要来源为种群间。比较果实性状与叶片性状的平均表型分化系数,果实性状(VST=78.26%)>叶片(VST=46.25%),即果实性状在种群间的分化大于叶片,叶片性状的变异主要来源于种群内。除坚果横径外,果实性状中的其他性状表型分化系数均大于70%,这表明果实性状以群体间的变异为主要变异。
表型性状
Phenotypic traits方差分量
Variance component方差分量百分比/%
Percentage of variance component/%表型分化系数VST/%
Phenotypic differentiation
coefficient/%种群间
Among populations种群内
Within population随机误差
Random error种群间
Among populations种群内
Within populationFFW 8.161 2.748 1.454 66.01 22.23 74.81 FHD 4.017 1.505 0.454 67.22 25.18 72.75 FVD 11.262 1.331 0.494 86.06 10.17 89.43 PT 0.977 0.172 0.073 79.95 14.10 85.03 FNW 0.551 0.166 0.136 64.54 19.49 76.85 NHD 0.500 0.455 0.205 43.13 39.19 52.36 NVD 3.303 0.571 0.263 79.84 13.81 85.26 NSI 0.006 0.001 0.001 82.09 8.07 85.71 ST 0.011 0.003 0.001 75.97 17.93 78.57 SMT 0.009 0.002 0.001 75.45 17.13 81.82 TLL 1.167 1.540 6.549 1.08 16.64 43.11 TLW 0.126 0.144 0.673 13.39 15.28 46.67 TLSR 0.046 0.016 0.163 20.27 7.22 74.19 LLL 0.556 1.648 5.915 6.85 20.30 25.23 LLW 0.087 0.128 0.546 11.47 16.78 40.47 LLSR 0.033 0.036 0.146 15.17 16.93 47.83 均值Mean 1.926 0.654 1.067 49.28 17.53 66.25 Table 6. Variance components of phenotypic traits and differentiation coefficient among populations in C. dabieshanensis
-
基于前期对于叶片表型的分析,由于其变异系数大,性状不稳定易受环境影响,而顶生小叶叶形比、侧生小叶叶形比这2个性状相对于其他叶片性状变异较小,因此,在进行UPGMA聚类分析时,只应用了以上2个叶片性状及所有果实表型进行计算。结果(图2)表明:在距离系数为0.06时,19个种群可聚为2类,其中,SZ、BQ种群单独聚为一类,其余种群聚为一类。可能由于BQ、SZ 2个种群的经度同为115°28′ E,海拔高差300多米,距离较近有利于花粉传播,从而产生基因交流使其单独聚为一类。当距离系数为0.04时,可聚为4类,SZ、BQ种群为第Ⅰ类;GZ、JX、WB、GY、WZ、WC这6个种群为第Ⅱ类,除GZ种群外,其余5个种群经度均为115°35′~115°39′ E,且纬度也较接近;GX、SL、GN、SW 4个种群聚为第Ⅲ类,仅GN种群经度为115°22′ E,其余3个种群经度均为115°23′ E;GT、GS、TY、YL、CZ、YW、TB 7个种群聚为第Ⅳ类,它们的经度为115°46′~116°10′ E,属于19个种群中的高经度种群。对应实际的地理位置分布,UPGMA聚类基本与经度的变化一致,这可能与局部降水条件有关,但部分地理位置较近的种群并没有聚在一起。
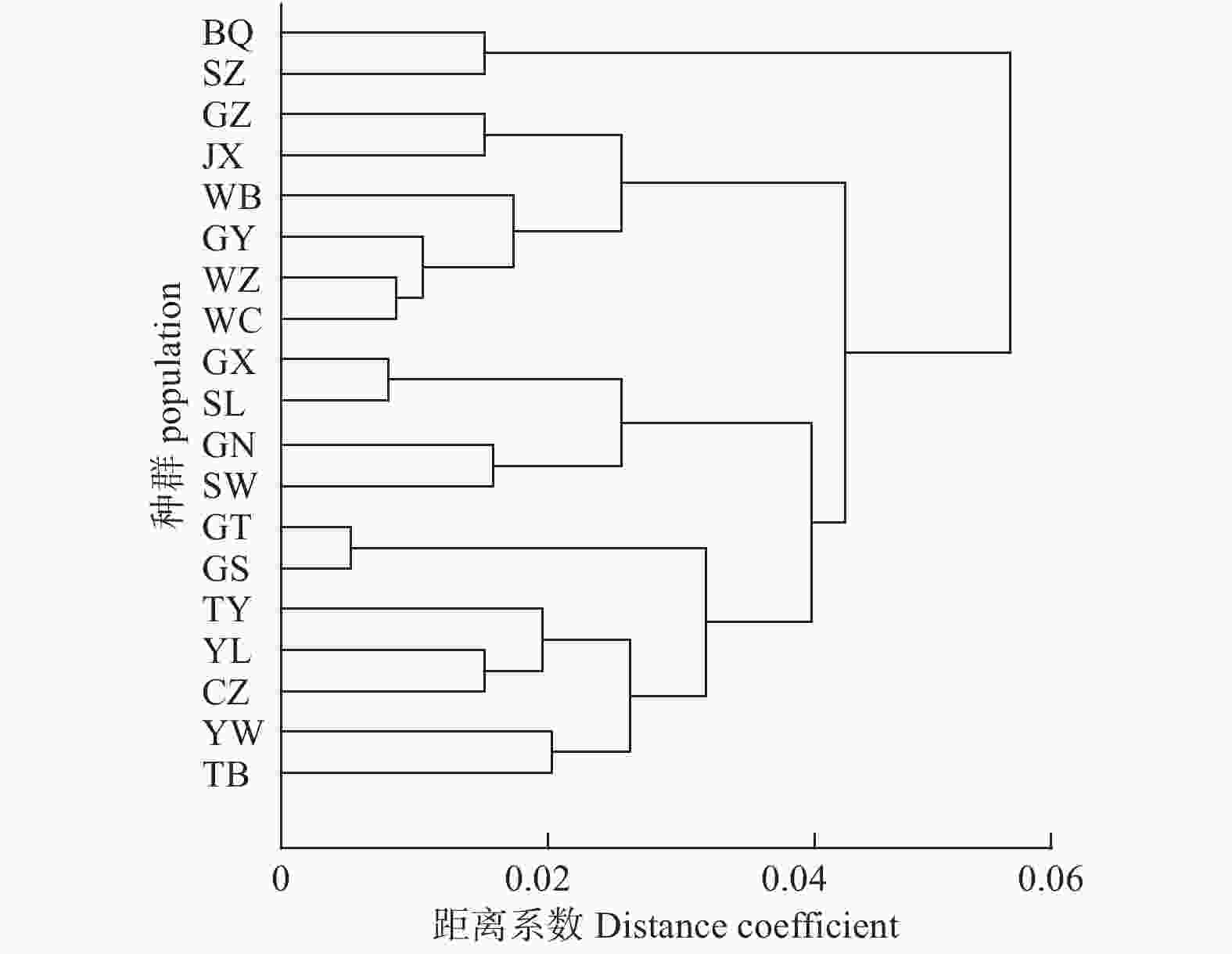
Figure 2. UPGMA cluster map of phenotypic traits in C. dabieshanensis with different geographical populations
2.1. 果实与叶片性状在种群间和种群内的变异特征
2.2. 大别山山核桃表型性状的变异情况
2.3. 大别山山核桃果实及叶片性状间的相关性
2.4. 大别山山核桃种群间的表型分化
2.5. 大别山山核桃种群聚类分析

-
通过对19个天然种群的果实与叶片表型性状的调查,发现大别山山核桃在种群间和种群内均存在极显著差异,表明大别山山核桃天然种群具有丰富的表型变异,此结果与郭传友[23],庞赛[22]对大别山山核桃天然群体的调查结果一致。其他物种如思茅松[24],青梅[25],香椿[3],云南松[26],油松[27]等在种群间和种群内也表现为极显著差异,具有很好的育种潜力,可为后期的遗传品质改良工作奠定重要基础。
表型分化系数表明了物种对于环境的适应情况,其值的大小与物种能够适应的范围成正比[24]。大别山山核桃16个表型性状的平均分化系数VST=66.25%,高于岷江柏VST=43.4%[28]、长柄扁桃VST=45.90%[29]、白花树VST=59.08%[30]等物种,与山苍子VST=60.19%[31]、无患子VST=62.21%[32]、内蒙古山杏VST=73.03%[33]接近,而远低于滇西北野生牡丹VST=98.51%[34],表明大别山山核桃天然种群分化程度较高,对环境的适应能力较强,种群间变异是表型变异的主要来源。
从地理位置的分布看,大别山山核桃天然种群的分布范围较狭窄,经纬度之差1度,而海拔落差约500 m,各种群分布于不同的山脉,产生诸多不同的局部小气候,且花粉传播也受到山脉的阻隔,这可能是引起种群间差异的原因之一。从繁殖系统看,大别山山核桃是雌雄同株异花植物,风媒异花传粉[35],但花期经常会遭受连绵阴雨,影响花粉传播,减少种群间的交流。另外,山核桃果实无休眠期,在温度湿度合适的情况下即可萌发,甚至在雨水较多的年份会出现果实未完全成熟却已出芽萌发的状况,使得种群内部幼苗增多,种子传播距离也缩短,种群间交流减少,逐步形成分化[36]。
-
种群间的变异反映了地理与生殖隔离上的变异,植物在种群间保持较大的变异也有在进化上的积极意义[34]。大别山山核桃果实与叶片性状在种群间变异均大于种群内变异,即果实与叶片性状均以种群间变异为主。相比于叶片表型,果实表型性状相对稳定,而叶片性状具有更丰富的表型多样性,此结果与山苍子[31]、浙江楠[37]的调查结果一致。这可能与叶片的功能有关,叶片属于营养器官,通过光合作用为植物提供养分,易受到环境因素的影响,变异性较强;而果实属于生殖器官,主要受自身遗传因素控制,性状相对稳定,这也保证了物种遗传上的相似性和稳定性[38]。
果实性状的变异程度因性状不同而差异较大,大别山山核桃果实鲜质量、坚果鲜质量与蒲壳厚度在所有果实表型中均呈现较大的变异,而果实横、纵径和坚果横、纵径的变异程度较小,这与翟大才等[39]对皖南山核桃表型性状的调查结果一致。两地果实性状的变异系数相比,除蒲壳厚度和坚果鲜质量2个性状以外,皖南山核桃其他果实性状的变异系数均大于大别山山核桃的;与安徽地区华东野核桃[40]和不同无性系的美国山核桃[41]对应的果实性状相比,大别山山核桃的果实表型变异系数均小于华东野核桃和美国山核桃,即大别山山核桃的果实性状变异程度更低,更不易受环境的影响,在遗传上表现也较稳定,对植物分类有重要作用[38]。种群变异系数可以反映种群的多样性丰富度,大别山山核桃19个种群的变异程度相似,平均值为9.63%,其中,YL、GX、SL、SW 4个种群的平均变异系数均大于10%,表型多样性较丰富,由于YL种群分布范围较小,其他3个种群的部分性状表现优良,因此,在进行种质资源选育与利用时,可重点考虑GX、SL、SW种群。
-
大别山山核桃作为优良的油料作物,富含优质脂肪酸及人体必需氨基酸,营养价值高,近年来逐步得到市场认可,经济价值也逐步提高。蒲壳厚度、坚果壳厚度及隔膜厚度这些性状是评定果实经济价值的重要指标,蒲壳厚度越小,出籽率越高;坚果壳厚度和隔膜厚度越小,则出仁率越高,经济效益更好。皖南山核桃果实表型与营养成分的相关性分析表明,坚果壳厚度与蛋白质、脂肪、油酸和亚油酸的含量呈显著正相关,因此,坚果壳较厚的山核桃可能营养价值更高[39]。蒲壳厚度与果实鲜质量、果实纵径、果实横径呈极显著正相关,这表明果实越大,其质量越大,则蒲壳越厚,而且扁圆形果实的蒲壳会更厚。坚果壳厚度与坚果横径、隔膜厚度呈极显著正相关,与果实纵径、坚果纵径、果形指数呈极显著负相关,即坚果横径越大,坚果壳越厚,而坚果纵径越大,果形细长的果实其坚果壳越薄,这在优良种质资源选育中有重要意义。
从经济效益的角度看,WB、WC 2个种群的坚果纵径、果形指数都较大,坚果壳、隔膜厚度也较薄,坚果呈椭圆形,可在这2个种群中选择薄壳椭圆形大果的大别山山核桃;GX、SL种群坚果鲜质量较大,而蒲壳较薄,可在其中选择籽大壳薄、出籽率高的优良种质;SW种群蒲壳最薄,果实鲜质量、果实横径在所有种群中最小,坚果壳、隔膜也较薄,可选择小果薄壳出仁率高的大别山山核桃。
3.1. 大别山山核桃不同种群的表型变异来源
3.2. 大别山山核桃果实与叶片表型的变异规律
3.3. 大别山山核桃的重要性状及经济效益
-
大别山山核桃天然种群的果实与叶片性状在种群间和种群内均存在极显著差异,其表型多样性非常丰富,变异主要来自于种群间。果实表型相对于叶片表型其性状更加稳定,其中果形指数的变异程度最小,性状最稳定。WB、WC、GX、SL、SW种群在经济价值方面表现良好,GX、SL、SW 3个种群表型多样性丰富且性状优良,可重点关注。在进行种质资源选育时,可从种群间和种群内两个层次展开,在增加取样种群密度的同时也应增大每个种群的取样范围,以选择出最优、最适的种质。大别山山核桃天然种群因生态环境、遗传因子、栽培管理情况的不同,产生了丰富的表型变异,本文对大别山山核桃的表型变异特征、来源及规律进行分析,为大别山山核桃种质资源的改良及新品种选育奠定了理论基础,对于其种质资源的保护、评价和利用有重要意义。
 DownLoad:
DownLoad: