



-
SQUAMOSA PROMOTER-BINDING PROTEIN-LIKE (SPL) 是植物中广泛存在且特有的一类转录因子,具有高度保守的SBP结构域,该结构域包含2个Zn2+结合位点和1个核定位信号序列[1-2]。拟南芥(Arabidopsis thaliana (L.) Heynh.)17个SPL家族成员中,11个基因是miR156的靶基因[3]。SPL基因家族成员的编码区或3′非编码区(UTR)中miR156的识别位点非常保守[4]。系统发育分析表明,这些SPL基因分为5组,分别为AtSPL3(AtSPL3、AtSPL4和AtSPL5)、AtSPL9(AtSPL9和AtSPL15)、AtSPL2(AtSPL2、AtSPL10和AtSPL11)、AtSPL6和AtSPL13[5-7]。根据其在拟南芥中的功能,上述SPL基因划分为三组:(1)AtSPL9、AtSPL15、AtSPL2、AtSPL10、AtSPL11和AtSPL13与营养和生殖阶段的转变相关,其中,AtSPL9、AtSPL15和AtSPL13起着最重要的作用;(2)AtSPL3、AtSPL4和AtSPL5促进花分生组织特性的转变;(3)AtSPL6影响某些生理过程[8]。众多研究表明,miR156/SPL调控系统影响植物的多种生物学过程,包括营养和生殖阶段的转变、叶毛发育、分蘖/分枝、果实成熟、胁迫反应、胚胎发育和花青素生物合成等[8-20]。拟南芥DICER-LIKE1(DCL1)突变体的八细胞胚胎时期,miR156的2个靶基因AtSPL10、AtSPL11上调最高[21]。过表达csi-miR156a或单独敲除2个靶基因CsSPL3和CsSPL14,可以显著提高柑橘愈伤组织的体细胞胚发生能力[22]。上述研究表明,miR156/SPL调控系统可能参与植物的胚胎发育。
落叶松(Larix spp.)体细胞胚体系的建立为现代分子育种技术进行遗传改良奠定了基础,也是研究植物胚胎发育及细胞全能性的理想实验材料。针对落叶松体细胞胚再生体系中,诸如胚性细胞诱导困难、体细胞胚同步化发育、生根率低等问题,选取落叶松原胚ESMs(embryonal-suspensor mass)、单胚和子叶胚材料,利用基因芯片、高通量测序和qRT-PCR等技术,通过预测与分析鉴定了许多已知和未知的MicroRNAs及其靶基因[23-26],根据其表达模式研究[25, 27-28],证明了miRNAs作为调控因子,参与了落叶松体细胞胚的发育过程,其中,发现miR156表达次高峰出现在早期子叶胚,最高峰在后期子叶胚[26]。另外,还发现miR156在同步胚中的表达远高于非同步胚[23]。上述研究表明,miR156的表达调控可能在落叶松体细胞胚发生过程中起重要作用。
为了阐明miR156/SPL调控系统在落叶松体细胞胚发育中的作用,笔者克隆了2个SPL基因的cDNA序列,将其命名为LaSPL2和LaSPL3。通过生物信息学分析及亚细胞定位研究,结合其在落叶松体细胞胚发育中的表达模式,揭示miR156如何调节靶向SPLs基因的表达,进而调控体细胞胚的发育进程。
HTML
-
以日本落叶松(Larix kaempferi (Lamb.) Carr)胚性细胞系S287为试验材料,于黑暗条件下22 ± 2℃培养,每3周继代1次。挑取一部分继代培养3周的材料并迅速放入液氮冷冻,用于总RNA提取和基因克隆;将另一部分ESMs分为两组,一组转到成熟培养基,并在诱导后0、2、5、7、10、14、21、28、35、42 d取材,共取10个时期;另一组转到ABA缺失的成熟培养基,分别在0、2、5 d取材,置于−80℃冰箱备用。
-
落叶松各组织总RNA,参照TaKaRa的Fruit-mate™ for RNA Purification说明书提取。取1 μg总RNA,按照TransScript® II All-in-One First-Strand cDNA Synthesis SuperMix for PCR反转录试剂盒操作说明书合成cDNA。研究表明,火炬松(Pinus taeda Linn.)TC68758和TC75581可能是miR156的靶基因[29]。根据TC68758序列设计引物LaSPL2-F/LaSPL2-R,从落叶松中扩增到947 bp的同源序列;将TC75581作为种子序列,搜索到1条落叶松同源序列(JR171955)。根据上述获得的2条落叶松同源序列,设计4组引物:3′RLaSPL2-1/3′ RLaSPL2-2、5′ RLaSPL2-1/5′ RLaSPL2-2、3′ RLaSPL3-1/3′ RLaSPL3-2、5′ RLaSPL3-1/5′ RLaSPL3-2(表1),用于RACE克隆LaSPL2和LaSPL3的全长cDNA序列(参照Clontech公司的SMARTer RACE 5′ /3′ Kit说明书进行)。
引物名称
Primer name序列 (5′–3′)
Sequence (5′–3′)用途
ApplicationLaSPL2-F CTCACTGGCAATAAACCC LaSPL2 PCR扩增 LaSPL2-R AAGACCTCAACAACTGCG PCR amplificationfor LaSPL2 3′ RLaSPL2-1 GTGGCATTGGAAACTTTGGAGGAC LaSPL2 的3′ Race 扩增 3′ RLaSPL2-2 CGCAGGTCCTCAGAAACACAAAG 3′ Race of LaSPL2 5′ RLaSPL2-1 GAGGATTGGGAGCAGATTGTCAGT LaSPL2 的5′ Race 扩增 5′ RLaSPL2-2 GAGAGCACACCCAGAGTCCGAGA 5′ Race of LaSPL2 3′ RLaSPL3-1 CTCCAGCAACGCTTCTGTCAGCA LaSPL3 的3′ Race 扩增 3′ RLaSPL3-2 CCCTCTGATTCACCAGTTGACAG 3′ Race of LaSPL3 5′ RLaSPL3-1 CTGCTACATTGCTGACAGAAGCGT LaSPL3 的5′ Race 扩增 5′ RLaSPL3-2 CAAACCCTGTGCCTCCTGTAGTA 5′ Race of LaSPL3 SLaSPL2-F CCCAAGCTTATGCTGTTTTCAGGCAACACC LaSPL2基因ORF扩增 SLaSPL2-R GGACTAGTCAGCAATTGATGAGTGTCATA LaSPL2 ORF amplification SLaSPL3-F CCCAAGCTTATGGACGAAGTCGAAGTCAAG LaSPL3基因ORF扩增 SLaSPL3-R GGACTAGTATGATTCTTTATTTCCTTGC LaSPL3 ORF amplification qLaSPL2-F GCTCCCAATCCTCTACTAATCT LaSPL2荧光定量引物 qLaSPL2-R CAGCCTGAGAACCTCTGAA Real-time PCR for LaSPL2 qLaSPL3-F TGACAAGATGACAATGGTGC LaSPL3荧光定量引物 qLaSPL3-R TGAAGAGAGATGAACTCAGGG Real-time PCR for LaSPL3 Table 1. Oligonucleotide primers used for PCR in the study
-
利用NCBI ORF Finder (
https://www.ncbi.nlm.nih.gov/orffinder/ ),搜索LaSPL2和LaSPL3的开放阅读框(ORF),并分析其氨基酸序列及UTR,鉴定miR156的识别位点;用ClustalX 2.1(https://clustalx.software.informer.com/2.1/ ),将推导的LaSPL2和LaSPL3氨基酸序列与拟南芥AtSPLs比对;利用EXPASY-PROSITE(http://prosite.expasy.org/ )对其特征序列进行预测;通过MEGAX,采用邻接法构建系统发育树。 -
通过EXPASY-PSORT(
http://psort.hgc.jp/ )预测LaSPL2和LaSPL3亚细胞定位情况。根据LaSPL2和LaSPL3 cDNA序列,设计引物并添加限制性酶切位点(表1),用于ORF的PCR扩增。用T4连接酶连接目的片段和pSuper1300-GFP。对阳性克隆鉴定并测序以确保序列的正确性。将融合表达载体构建完成后转入农杆菌GV3101,挑取阳性菌到LB培养基,其中含有25 μg·mL−1利福平(Rif)、40 μg·mL−1卡那霉素(Kan)、30 μg·mL−1庆大霉素(Gen),28℃振荡培养,最佳OD600为0.6~0.8;然后在4℃,6 000 g的条件下将菌液离心10 min,用转化液10 mmol·L−1氯化镁(MgCl2),10 mmol·L−12-(N-吗啡啉)-乙基磺酸(MES),pH 5.7,200 μmol·L−1乙酰丁香酮(AS)重悬菌体,室温条件下放置3 h。用小型注射器将重悬菌液注入烟草叶片,置于培养箱内培养2 d。取部分烟草叶片,置于1 μg·mL−14′,6-二脒基-2-苯基吲哚(DAPI)溶液中浸泡,分别于405 nm和488 nm观察DAPI染色和GFP融合蛋白的表达情况。 -
根据LaSPL2和LaSPL3序列比对结果,设计2对引物qLaSPL2-F/qLaSPL2-R、qLaSPL3-F/qLaSPL3-R(表1)。通过PCR扩增,将LaSPL2和LaSPL3的目的条带进行测序并验证其准确性。利用CFX96荧光定量PCR仪检测LaSPL2和LaSPL3在落叶松体细胞胚不同发育阶段的表达水平,使用的试剂盒为TaKaRa SYBR Premix Ex TaqTM kit,内参基因为LaEF1A1(GenBank accession: JR153706)[26]。
1.1. 材料
1.2. LaSPL2和LaSPL3全长cDNA克隆
1.3. LaSPL2和LaSPL3生物信息学分析
1.4. LaSPL2和LaSPL3亚细胞定位研究
1.5. 落叶松体细胞胚发育过程中LaSPL2和LaSPL3的表达模式
-
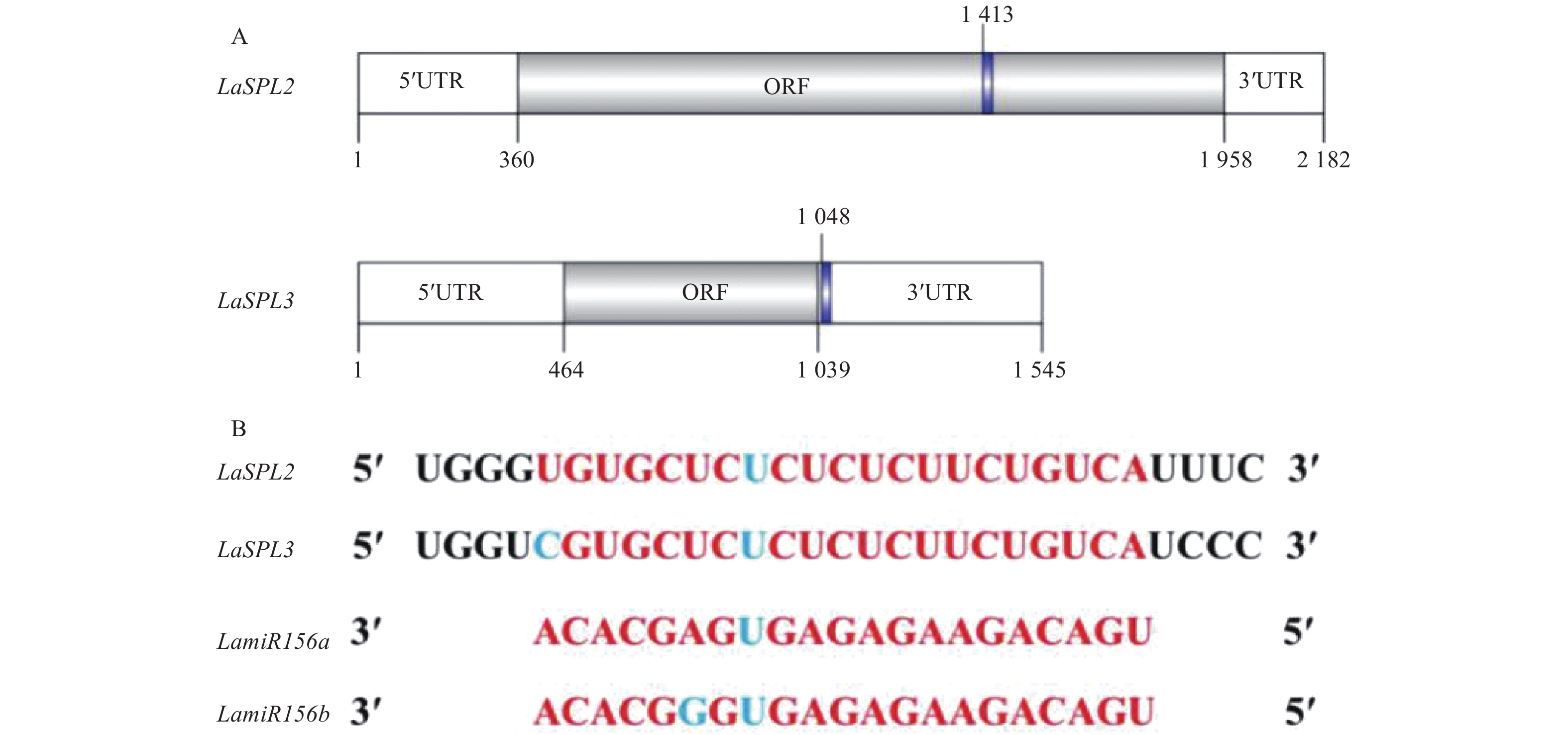
火炬松中预测了2条miR156的靶基因序列TC68758和TC75581[29]。根据TC68758序列设计引物LaSPL2-F/LaSPL2-R,进行RT-PCR扩增,从落叶松中扩增到947 bp的同源序列,并在落叶松转录组中搜索到TC75581的同源序列JR171955。4组RACE扩增的引物是以上述2条落叶松的同源序列为模板,经过2轮巢式PCR,分别获得了287、1268、365和755 bp。经序列组装后获得了2条全长cDNA序列,并分别命名为LaSPL2 (GenBank Accession No.: MN315910)和LaSPL3 (GenBank Accession No.: MN315911)。经NCBI ORF finder搜索发现,LaSPL2和LaSPL3分别编码532个和191个氨基酸,预测蛋白分子量分别为57 213.77、21 726.37 Da,等电点分别为8.90、9.33。进一步分析发现,LaSPL2和LaSPL3 mRNA中均存在miR156识别位点(图1),其中,LaSPL2的识别位点位于编码区,而LaSPL3位于3′UTR区域,表明其可能受miR156的调控。
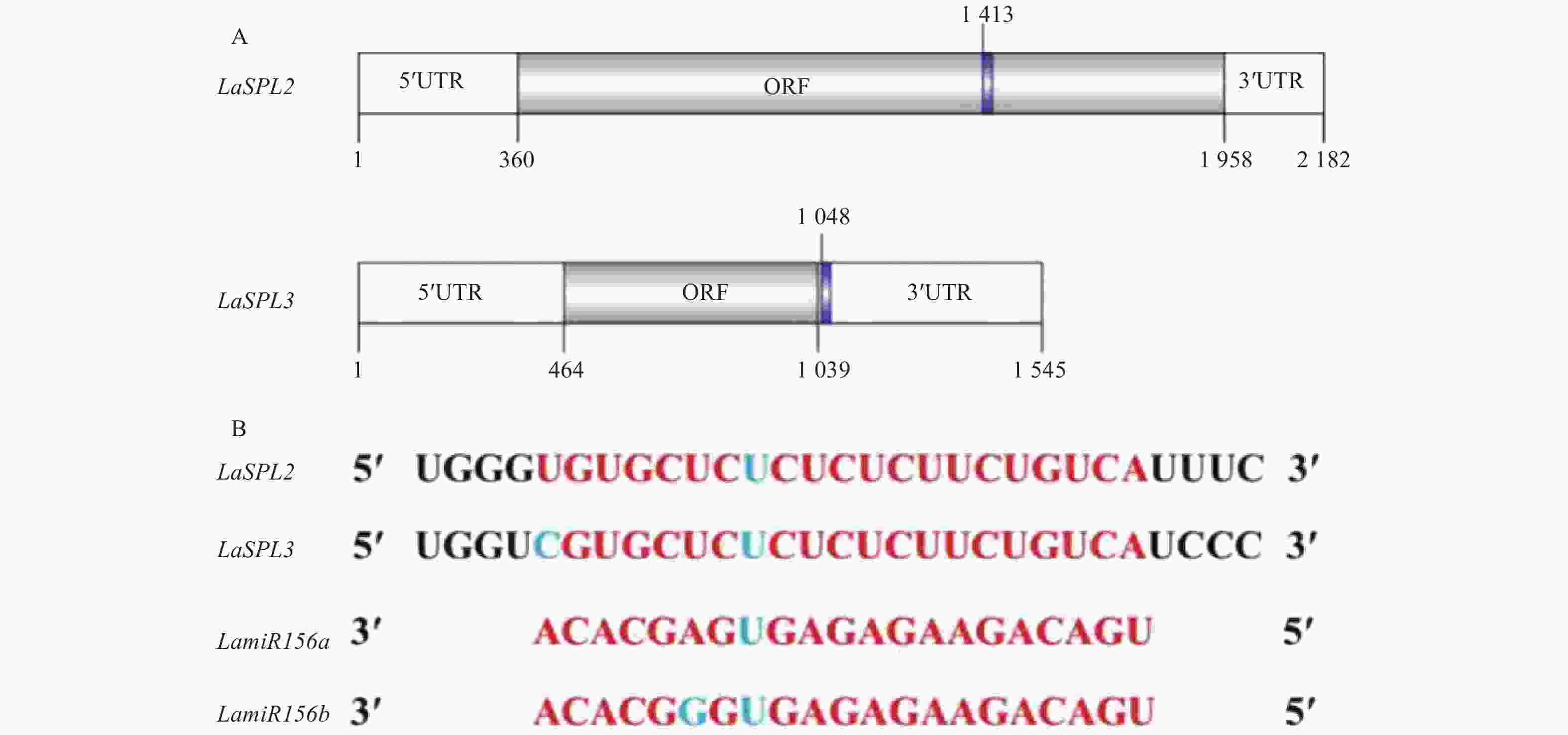
Figure 1. Predicted miR156 position in LaSPL2 and LaSPL3 (A) and alignment of miR156 and LaSPL2 and LaSPL3 transcript sequences (B)
从UniProt数据库搜集拟南芥AtSPLs家族成员的蛋白序列,并将其与推导的蛋白序列进行比对。结果表明,LaSPL2和LaSPL3具有非常保守的SBP结构域(ZF_SBP,Zinc finger SBP-type profile)(图2)。通过EXPASY-PROSITE搜索,氨基酸序列124-201[LaSPL2,V(124)PRCQVEGCKTELTTAKDYHRRHKVCELHSKSPKVIVNGIEQRFCQQCSRFHILSEFDEGKRSCRRRLAGHNERRRKP(201)]、65-142[LaSPL3,A(65)PSCQVEKCAADLADAKEYYRRHRVCEQHSKARIVLVLGLQQRFCQQCSRFHELEEFDEAKRSCRRRLAGHNERRRKT(142)],与SBP结构域的典型特征序列(C-x4-C-x16-C-x2-[HC]-x15-C-x2-C-x3-H-x11-C)相匹配,证明其具有SPL家族蛋白的特征序列。
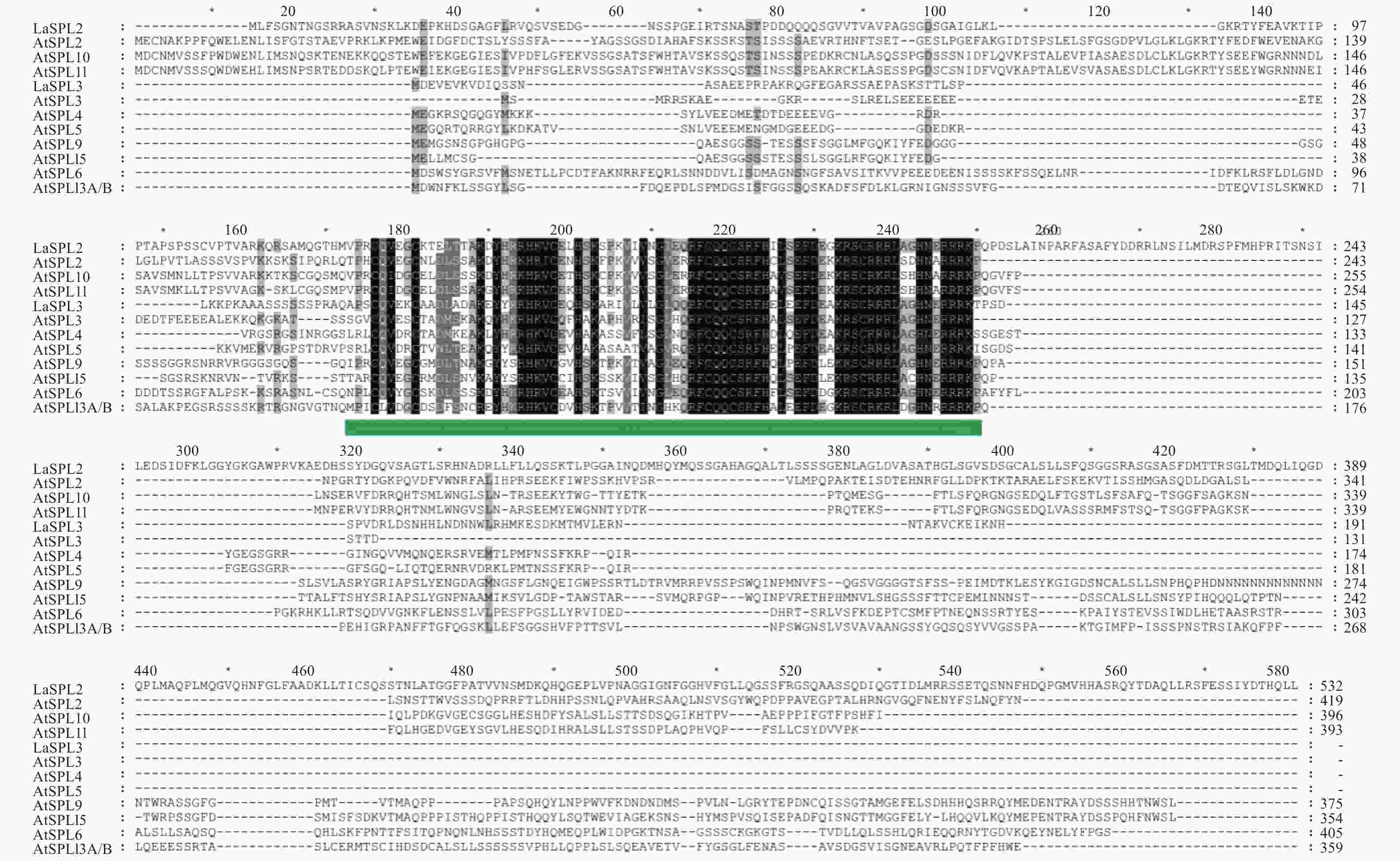
Figure 2. ClustalX 2.1 alignment of the deduced protein sequence encoded by the Japanese larch (Larix leptolepis) LaSPL2 and LaSPL3 and the proteins encoded by the members of the Arabidopsis thaliana SPL family regulated by miR156.
利用MEGAX,构建了SPL蛋白家族的系统发育树(图3)。结果表明,LaSPL2和LaSPL3分别与拟南芥的AtSPL2/10/11、AtSPL3/4/5最相似。拟南芥中AtSPL2、AtSPL10、和AtSPL11 与营养和生殖阶段的转变相关,而AtSPL3、AtSPL4和AtSPL5与花分生组织的转变相关 [8]。因此在基因的结构和功能上,推测LaSPL2和LaSPL3分别与AtSPL2和AtSPL3具有相似性。
Figure 3. Phylogenetic analysis of LaSPL2 and LaSPL3 and the Arabidopsis thalianaSPL family members.
-
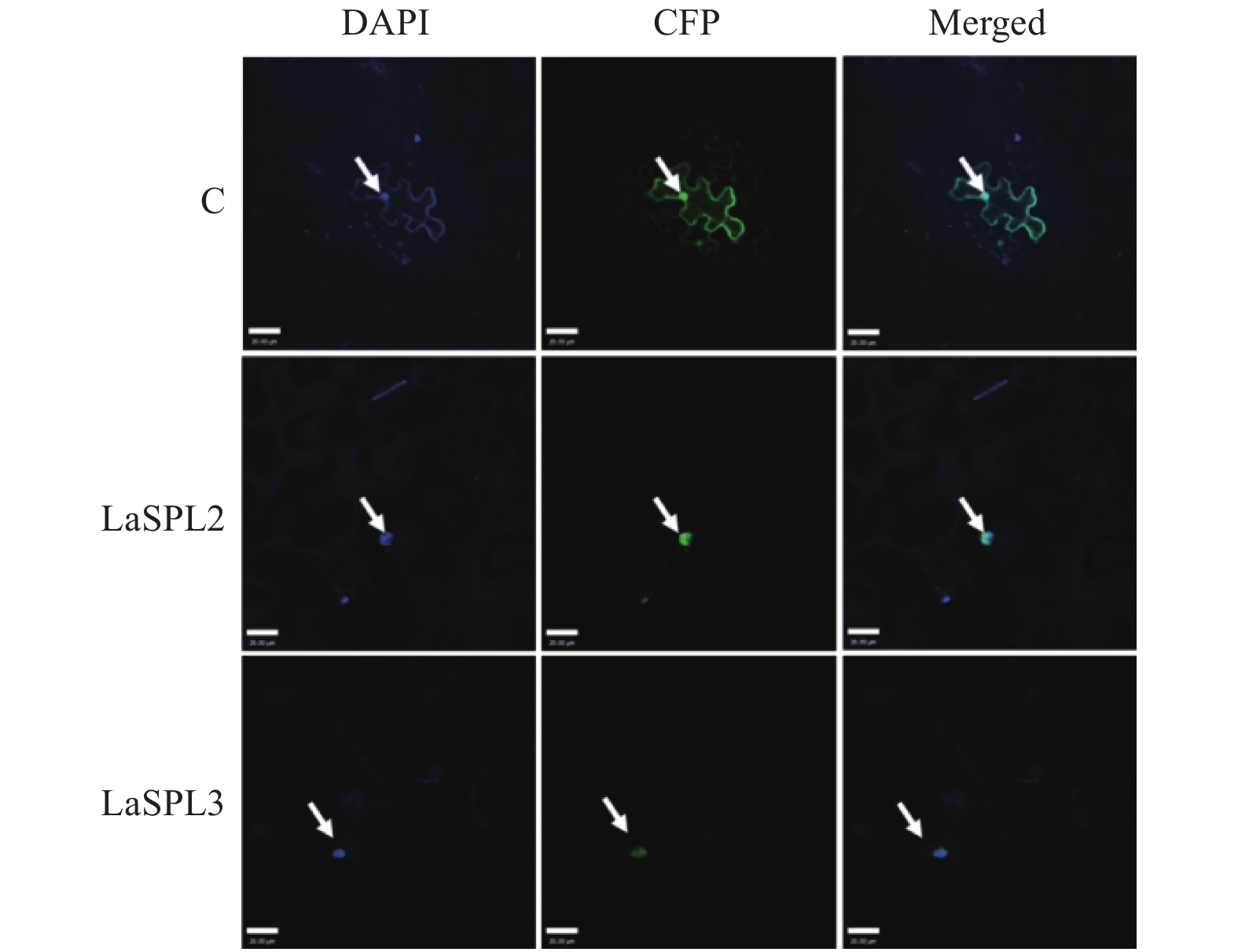
前人研究表明,拟南芥AtSPLs定位于细胞核[30]。ExPASy-PSORT预测LaSPL2和LaSPL3定位于细胞核。为了明确其在细胞中发挥功能的具体部位,引物在LaSPL2和LaSPL3 ORF两端设计,并添加Hind III和Spe I酶切位点(表1),用于扩增其ORF序列;用T4连接酶将PCR产物和pSuper1300-GFP连接,构建pSuper1300-LaSPL2-GFP和pSuper1300-LaSPL3-GFP融合表达载体。将序列正确的表达载体,转入农杆菌GV3101,空质粒pSuper1300-GFP作为阴性对照(C)。用注射器将农杆菌重悬液注入烟草叶片,将其置于培养箱内继续培养2 d。取部分烟草叶片,置于1 μg·mL−1 DAPI溶液中浸泡,用激光共聚焦扫描显微镜在488 nm观察GFP的绿色荧光信号,并在405 nm下观察DAPI染细胞核的蓝色荧光信号。LaSPL2-GFP和LaSPL3-GFP仅在细胞核内观察到有绿色荧光信号,而对照则呈现弥散的荧光信号(图4)。表明LaSPL2和LaSPL3是核定位蛋白,与其作为转录因子的功能相一致。
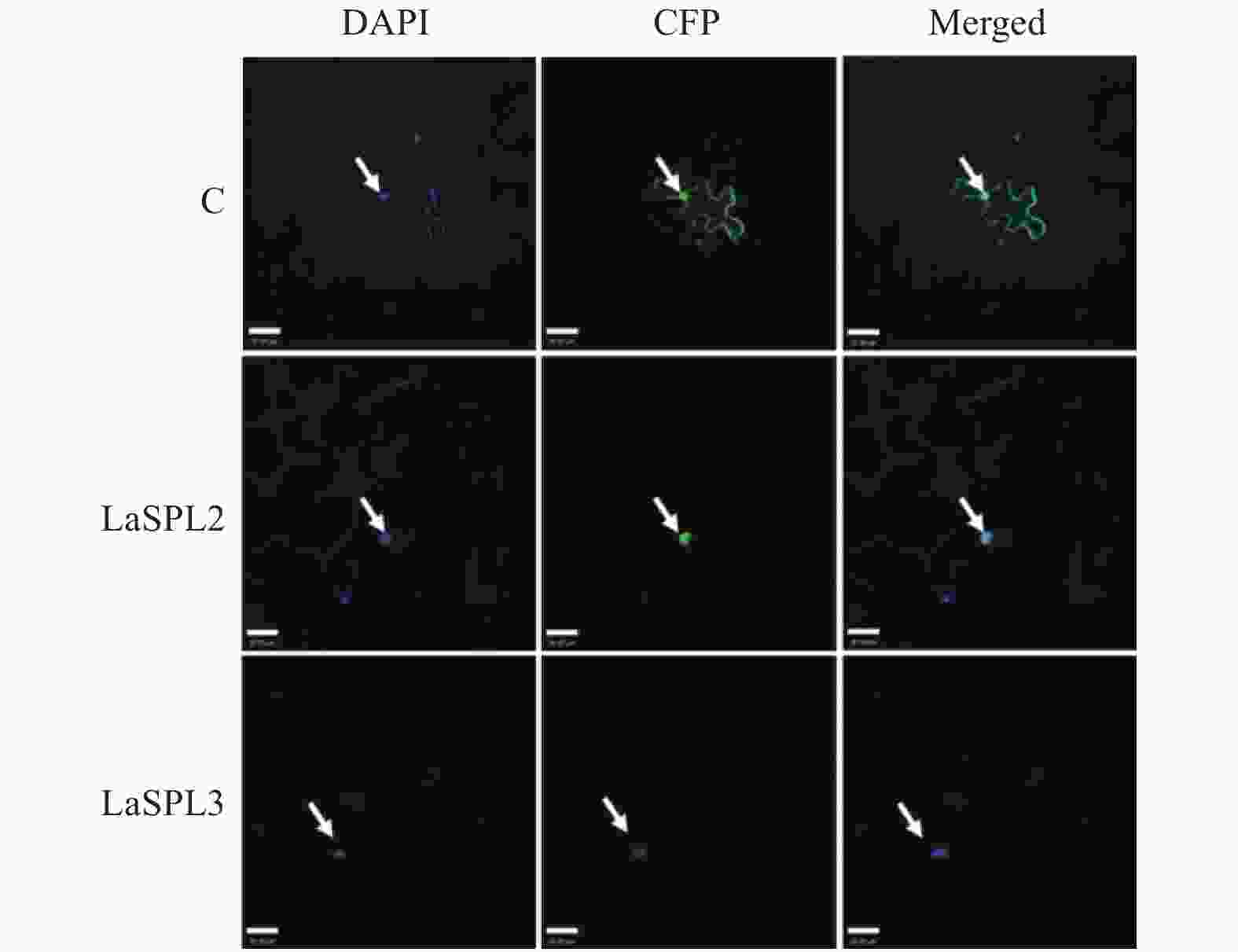
Figure 4. Subcellular localization ofLaSPL2 and LaSPL3
-
为了研究LaSPL2和LaSPL3在胚胎发育中的作用,检测了其在落叶松体细胞胚发育过程的表达模式。当ESMs转接到成熟培养基后,在植物生长调节剂(PGRs)的撤除和ABA共同作用下,促进了早期胚胎的形成。结果表明,随着PGRs的撤除及ABA处理,LaSPL2和LaSPL3均有不同程度的响应。当ESMs转到成熟培养基(PGRs撤除)的前5 d,即在体细胞胚发育早期,LaSPL2和LaSPL3的表达均有所下调(图5A、C),其中,LaSPL3下调程度较多(图5C);而在ABA缺失的情况下,其表达水平略有上调(图5B、D)。
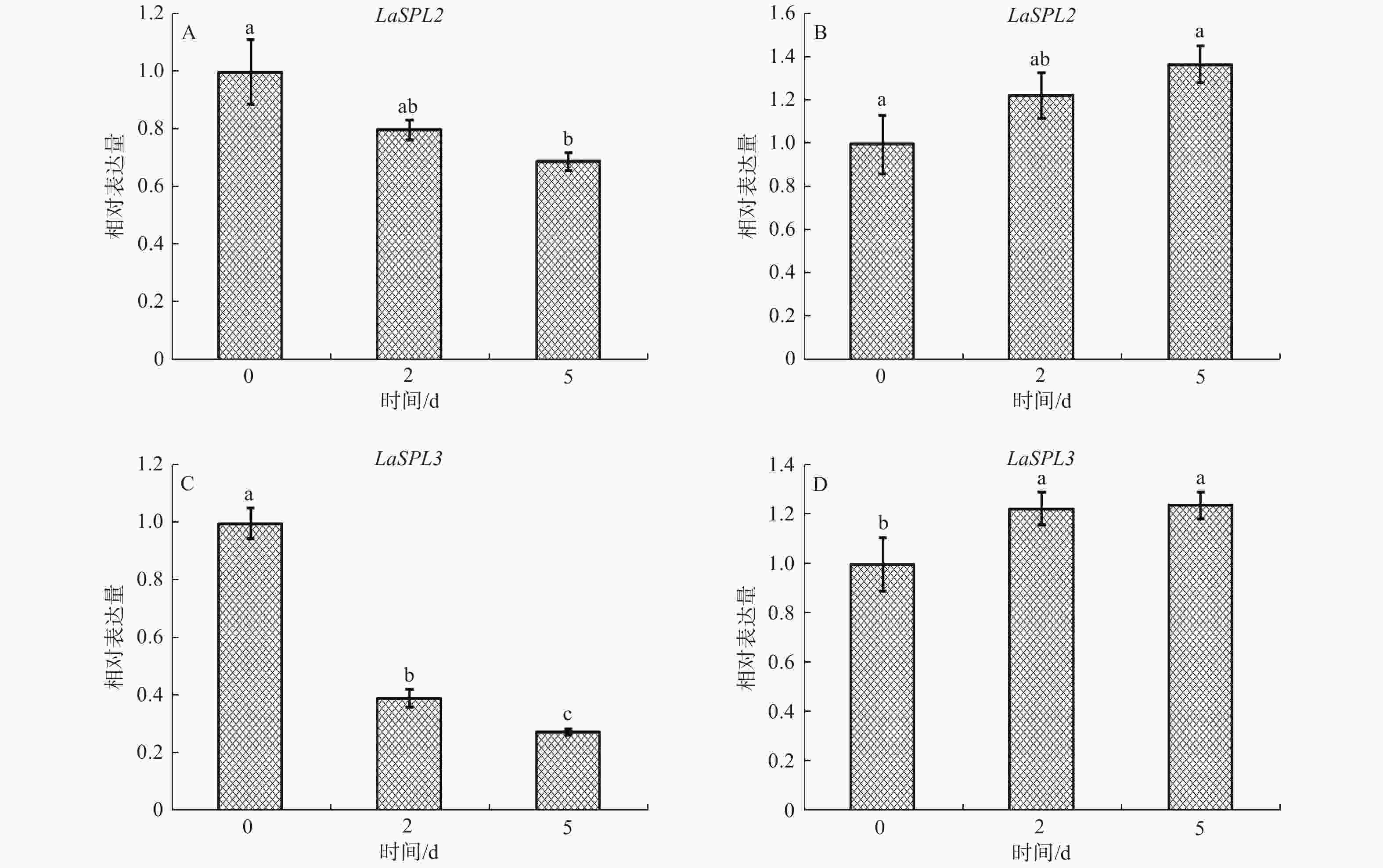
Figure 5. Accumulation of LaSPL2 (A, B)and LaSPL3 (C, D)transcripts in ABA-treated (A, C) and no ABA-treated ESMs (B, D) ESMs during the early somatic embryogenesis stage.
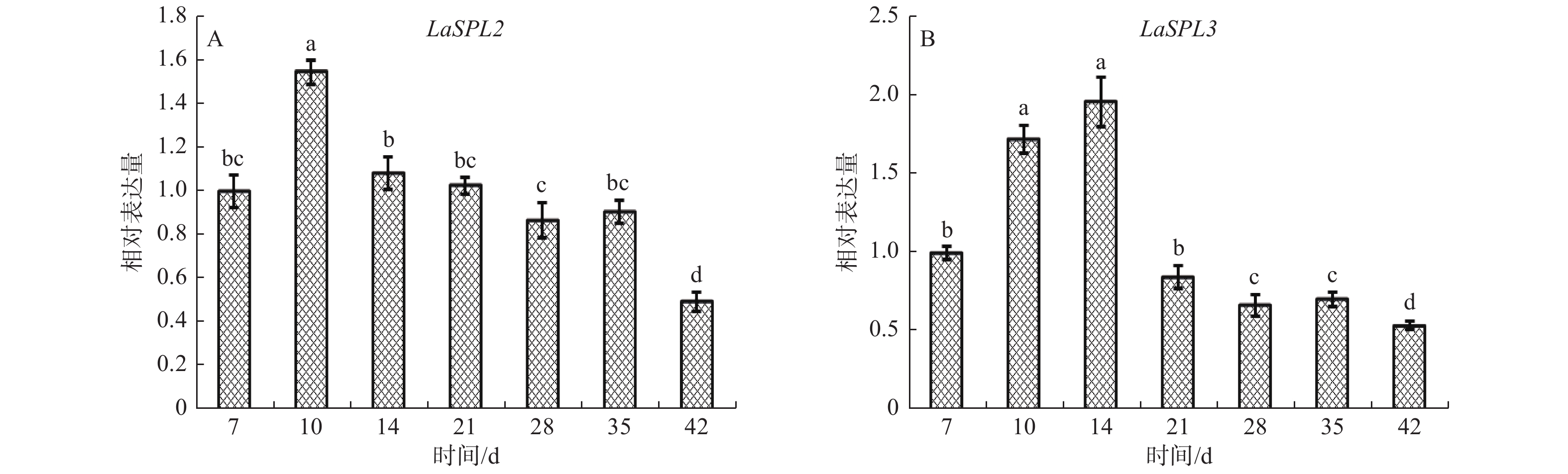
当ESMs转接到成熟培养基后,第7~10天时出现黄色胚头[31]。qRT-PCR结果(图6)表明:随着体细胞胚的进一步发育,LaSPL2的表达水平在10 d达到最高峰,14~35 d表达下降,期间变化不明显,42 d降到最低值;LaSPL3的表达水平在7~14 d逐渐升高,随后下调,42 d达到最低值;表明这2个基因可能与早期胚胎发生相关。
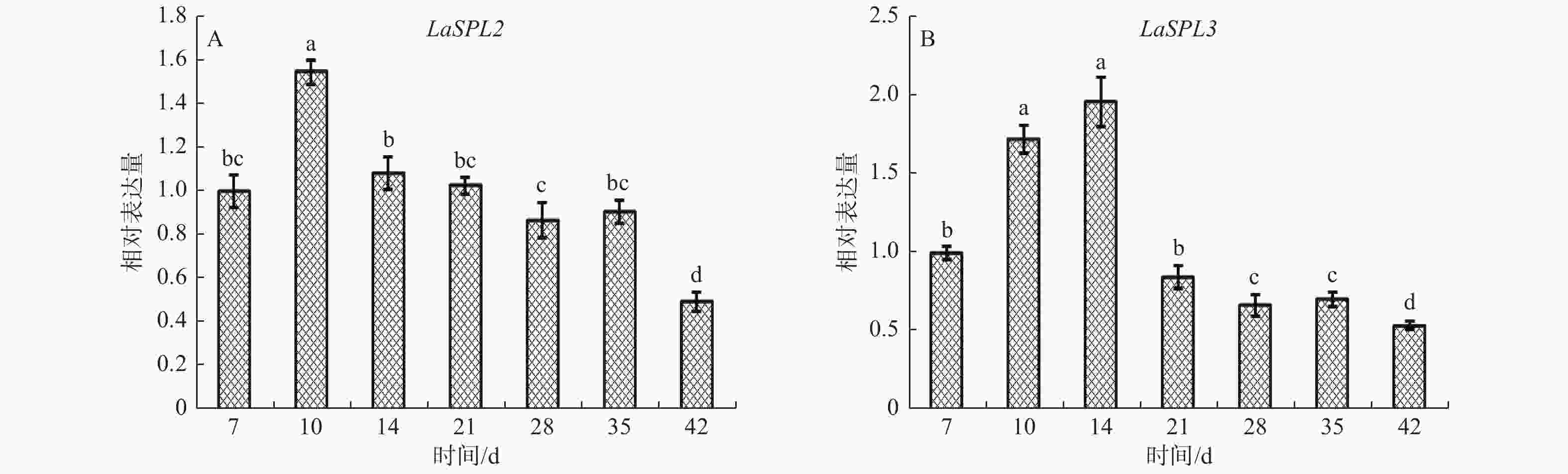
Figure 6. Expression patterns of LaSPL2 (A) and LaSPL3 (B) during somatic embryogenesis.
2.1. LaSPL2和LaSPL3全长cDNA克隆及生物信息学分析


2.2. LaSPL2和LaSPL3亚细胞定位研究
2.3. LaSPL2和LaSPL3在落叶松体细胞胚发育中的表达模式

-
miR156通过识别SPL基因家族编码区或3′非翻译区(UTR)中保守的识别位点,进行转录后或翻译水平调控[4]。拟南芥AtSPL3、AtSPL4和AtSPL5的miR156识别位点位于3′UTR区域,其余AtSPLs的miR156识别位点位于编码区[3, 32]。本研究从落叶松中鉴定了2个SPL同源基因,系统发育树分析表明,它们分别与拟南芥AtSPL2/10/11、AtSPL3/4/5最相似(图3),将其命名为LaSPL2和LaSPL3。多序列比对发现,该基因编码的蛋白质具有SPL家族蛋白的典型特征,具有保守的SBP结构域(图2),表明LaSPL2和LaSPL3与SPL家族同源。LaSPL2和LaSPL3 mRNA具有miR156识别位点(图1),其中,LaSPL2的识别位点位于编码区,而LaSPL3的识别位点位于3′UTR,这与拟南芥中的研究结果相一致,其表达在一定程度上可能受miR156调控。本研究构建了pSuper1300-LaSPL2-GFP和pSuper1300-LaSPL3-GFP融合表达载体,通过烟草瞬时表达体系进行亚细胞定位,结果显示LaSPL2和LaSPL3定位于细胞核中(图4),因具有保守的SBP结构域,表明LaSPL2和LaSPL3是SPL家族蛋白,故而推测它们的编码产物为SPL类转录因子。
ABA在落叶松体细胞胚发育中起重要调控作用[33],可以诱导H2O2和ROS的产生,并调节CAT、SOD和APX的表达[34]。H2O2可能通过细胞信号转导系统影响基因表达,进而诱导体细胞胚发育[34]。研究表明,拟南芥过表达miR156可以降低H2O2水平和SA信号通路相关基因的转录水平[35],同时降低植物对真菌的免疫抗性。上述结果表明,miR156/SPLs系统影响ROS的积累,并控制SA信号通路的激活或失活[35]。通过分析ABA缺失对LaSPL2和LaSPL3表达的影响(图5),表明ABA可能是落叶松体细胞胚发育早期LaSPL2和LaSPL3下调表达的主要因子。
miRNAs通过影响植物激素的生物合成、转运和信号转导途径而成为植物生长发育的关键调控因子[36]。生长素在植物体胚细胞重编程和体细胞胚胎发育中发挥重要作用[37-39]。前期研究表明,LaGH3的表达在一定程度上反映了落叶松体细胞胚发育过程中内源生长素的动态变化,体细胞胚发育前6 d,ABA是迅速下调LaGH3的主要因子,反应此时在ABA作用下生长素水平可能会下调,以降低细胞分裂速度,实现ESMs从快速增殖状态到胚胎发育的转变,进而促进体细胞胚的进一步发育[40]。上述研究表明,ABA可能通过调控生长素的水平而影响LaSPL2和LaSPL3的表达。在ABA缺失的情况下,LaSPL2和LaSPL3表达水平略有上调(图5B、D)。前期研究发现,在ABA缺失的培养基上生长的ESMs中H2O2含量升高,细胞由白色逐渐转变为褐色,体细胞胚发育受阻[31],这可能与ABA缺失导致的早期胁迫有关。
随着体细胞胚的进一步发育,LaGH3转录水平逐渐增加,在28 d达到高峰,表明生长素在体细胞胚的进一步发育中具有重要作用[40]。此时,LaSPL2和LaSPL3的表达水平,分别在10 d和14 d达到峰值,表明其可能与早期胚胎形成相关(图6),同时H2O2含量在10 d左右时处于低谷期[34]。因此,认为此时LaSPL2和LaSPL3的高表达可能与生长素、H2O2调控相关,进而参与体细胞胚的早期形成。
拟南芥miR156有助于促进胚胎形态建成[21, 41-42]。DICER-LIKE1(DCL1)突变体中,除了AtSPL10、AtSPL11外,AtSPL2表达水平上调了5.33倍[21]。柑橘中敲除CsSPL3可以显著提高柑橘愈伤组织的体细胞胚发生能力[22]。上述结果表明,miR156/SPL调控系统可能参与植物的胚胎发育。前期研究表明,在落叶松早期子叶胚发育的前6个阶段,miR156b前体和成熟体表达水平逐渐升高[26, 43],表明其表达可能与ABA的调控相关。拟南芥的原位杂交分析中,发现miR156信号随着合子胚胎的发育而逐渐增强[21],与落叶松体细胞胚发育中的表达模式相一致[26]。本研究发现,随着体细胞胚发育成熟,LaSPL2和LaSPL3表达量逐渐下降,并在42 d达到最低值(图6)。这与笔者前期发现miR156水平在成熟体细胞胚中达到峰值相一致[26]。因此,落叶松中ABA可能通过miR156下调LaSPL2和LaSPL3,提高体细胞胚发生能力,进而促进胚胎发育及成熟休眠。
-
本研究从落叶松中克隆了2个SPL同源基因LaSPL2和LaSPL3,均具有保守的SBP结构域,其cDNA序列中存在miR56的识别位点;亚细胞定位结果表明LaSPL2和LaSPL3定位于细胞核;qRT-PCR结果表明,ABA可能是落叶松体细胞胚发育早期LaSPL2和LaSPL3下调表达的主要因子;LaSPL2和LaSPL3在体细胞胚发育的早期阶段达到最高峰,提示其可能参与体细胞胚的早期形成。随着体细胞胚发育成熟,LaSPL2和LaSPL3表达可能受miR156的调控,在体细胞胎的成熟休眠中具有重要意义。
 DownLoad:
DownLoad: