




-
森林天然更新是森林生态系统自我繁衍恢复的手段[1],对未来森林群落的组成、结构及其生物学多样性具有十分重要的作用[2],是森林经营和森林生态学研究的重要内容之一。近年来,有关森林天然更新的研究主要集中在幼苗组成和空间分布格局[3]、更新动态以及更新影响因素[4]等方面,其中在对天然更新影响因素的研究中,主要关注环境因子[5-7](光照、土壤)、地形因子[8-9]、林分因子[10-11](郁闭度、枯落物、灌草盖度)、种内种间竞争关系[12]以及干扰[13]等方面的影响,对于林分结构尤其是林分空间结构对天然更新的影响研究较少,有研究表明:林分水平结构和垂直结构直接决定着林下的光照、降水以及土壤养分含量,进而对更新幼苗的建立、存活和生长产生不同程度的影响[14-17],是森林天然更新成功的关键因子之一[12],同时林分空间结构也是森林经营过程中最有可能调控的因子[18],因此,开展林分空间结构对天然更新影响研究,揭示林分空间结构对更新幼苗的影响机制,对人工促进天然更新,实现森林的可持续发展具有重要意义。
蒙古栎(Quercus mongolica Fisch.)林是东北地区常见的典型森林类型之一,其面积占东北有林地面积的15%~20%,在涵养水源、保持水土及维持生态稳定性方面发挥了重要作用,具有较高的经济和生态价值。近年来,由于经营不当,部分蒙古栎林出现天然更新差、幼苗存活率低,幼苗难以成树的现象。因此,本研究以辽东山区蒙古栎林为研究对象,探讨林分结构对蒙古栎林更新幼苗的影响,皆在解决蒙古栎林天然更新障碍的问题,以期为人工促进蒙古栎林天然更新提供理论依据。
HTML
-
辽东山区地处长白山脉龙岗山脉北麓,地理坐标为40°00′~43°09′ N、122°53′~125°47′ E,属温带大陆性湿润季风气候,年均气温4~11℃,年均降水量700~850 mm,年蒸发量925~1 284 mm,≥10℃的年有效活动积温为2 497.5~3 943.0℃,无霜期120~139 d,年日照时数为2 403.1 h。土壤以棕壤暗棕壤为主。研究区主要乔木树种有红松(Pinus koraiensis Sieb.)、蒙古栎、水曲柳(Fraxinus mandshurica Rupr.)、胡桃楸(Juglans mandshurica Maxim.)、沙松(Abies holophylla Maxim.)、紫杉(Taxus cuspidada Sieb. et Zucc.)、色木槭(Acer mono Maxim.)、硕桦(Betula costata Trautv.)、拧筋槭(Acer triflorum Komarov)、毛榛子(Corylus mandshurica Maxim.)、东北山梅花(Philadelphus schrenkii Rupr.)、暴马丁香(Syringa reticulata subsp. amurensis)、东北赤杨(Alnus mandshurica (Call.) Hand.-Mazz.)等;林下灌木主要有胡枝子(Lespedeza bicolor Turcz.)、龙牙楤木(Aralia elata (Miq.) Seem.)、东北山梅花(Philadelphus schrenkii Rupr.)、卫矛(Evonymus alatus (Thunb.) Sieb.)、迎红杜鹃(Rhododendron mucronulatum Turcz.)等;草本主要有关苍术(Atractylodes lancea (Thunb.) DC.)、尾叶香茶菜(Rabdosia excisa(Maxim.)Hara)、水金凤(Impatiens noli-tangere L.)、荨麻叶龙头草(Meehania urticifolia (Miq.) Makino)等。藤本植物有软枣猕猴桃(Actinidia arguta (Sieb. et Zucc.) Planch. ex Miq.)、五味子(Schisandra chinensis (Turcz.) Baill.)、南蛇藤(Celastrus orbiculatus Thunb.)等。
-
2019年和2020年的6—9月,分别在新宾县赵家林场、桓仁县八里甸子林场、本溪县碱厂林场、岫岩县刘家河镇等蒙古栎次生林内,采用典型抽样法,分别设置25 m × 25 m的矩形样地28块、0.1 hm2的圆形样地9块,共计37块样地。对样地内胸径 ≥ 5 cm的林木进行每木检尺,记录树种名称、位置、胸径、树高、冠幅、第一枝下高及健康情况。同时,在样地内随机设置3个5 m × 5 m(矩形样地)、半径 2 m 圆形样地的更新苗小样方,记录其中胸径 < 5 cm的所有乔木更新幼苗的种类、基径、苗高和盖度等。利用手持GPS记录样地的地理位置、海拔、坡位、坡度、坡向等,样地基本情况见表1。
样地号
Sample
plot
No.海拔
Altitude/
m坡度
Slope/
(°)坡向
Aspext坡位
Slope
position密度
Density/
(tree·hm−2)平均
胸径
Mean
diameter/
cm平均
树高
Mean
height/
m1 603 27 北 上 688 22.3 15.5 2 606 20 北 上 608 22.2 14.4 3 606 17 北 上 528 20.7 13.1 4 598 20 北 上 528 22.4 15.2 5 713 25 南 上 832 19.6 12.5 6 696 25 南 上 800 21.1 13.9 7 755 25 南 中 720 22.7 14.5 8 755 24 南 中 640 18.9 11.1 9 656 30 南 中 880 21.0 16.8 10 645 21 南 中 944 21.4 17.6 11 641 25 西北 中 816 23.0 20.3 12 646 20 南 中 656 23.6 18.1 13 685 27 南 上 1360 18.5 14.0 14 680 17.7 南 上 1072 19.9 15.4 15 679 20 南 上 1168 20.1 15.2 16 687 31 南 上 1152 18.2 14.0 17 588 18.9 西 中 528 19.4 13.3 18 617 28 西 中 617 18.8 12.1 19 599 11 西 中 720 19.6 12.5 20 615 22.5 西南 中 624 22.8 13.9 21 650 17 西南 下 656 25.3 14.7 22 644 16 西南 下 464 32.0 16.9 23 634 16 西南 下 480 28.8 16.2 24 638 14 西南 下 384 28.4 14.5 25 655 24 东 上 496 23.0 15.3 26 651 19 东 上 560 24.9 16.0 27 650 17 东 上 448 24.6 14.9 28 650 18 东 上 704 23.6 13.9 29 317 24 东 中 230 31.0 17.4 30 220 19 南 中 150 31.7 14.2 31 495 10 东 中 240 34.8 17.0 32 303 27 南 上 470 17.0 13.0 33 381 19 西 中 450 16.3 12.7 34 330 19 北 上 1250 10.8 10.9 35 391 25 西南 中 480 20.5 10.1 36 395 35 西 中 1040 12.7 9.1 37 214 29 西北 中 620 19.0 9.9 Table 1. Basic characteristic of the sampling plots
-
采用基面积、混交度(Mi)、角尺度(Wi)、林层指数(Si)、密集度(Ci)、Hegyi竞争指数(CIi)作为表达林分结构的参数。为避免边缘效应对林分结构的影响,设置3 m为缓冲区。本研究结构单元n取4。
混交度(Mi)表达树种空间隔离程度的指数[19]。
式中:当参照树i与相邻木j非同种时,Vij = 1,反之Vij = 0;n为最近邻木株数(下同)。
角尺度(Wi)用来反映林分林木的水平空间分布格局[20]。
式中:当第j个α角小于标准角α0时,Zij = 1,反之Zij = 0;α0 = 360/(n + 1)。
林层指数(Si)表达林层的垂直结构,其反映了空间结构单元中林层的垂直结构分布格局及林层多样性[21]。
式中:Zi为参照树的空间结构单元内林层的个数;当参照树与最近邻近木第j株相邻木属同一林层时,Sij = 0,反之Sij = 1。
密集度(Ci)用来反映林木的疏密程度[22]。
式中:当参照树i与相邻木j的树冠投影相重叠时,
$ {y}_{ij}=1 $ ,反之$ {y}_{ij}=0 $ ;Hegyi竞争指数(CIi)用来反映林木个体所承受的来自相邻木的竞争压力大小[23]。
式中:dj为最邻近木j的胸径,di为参照树i的胸径,Lij为参照树i与第j株相邻木之间的水平距离。
-
采用α多样性测度方法中的Patrick丰富度指数(S)、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)来分析更新幼苗的物种多样性水平。其计算公式如下:
式中,N为样地内乔木更新物种的总数,Pi为第i种的个体数占所有种个体总数的比例。
-
采用SPSS20.0和Origin2019b软件对数据进行统计分析。利用散点图结合拟合模型分析林分结构变量、更新幼苗密度之间的相互关系,采用决定系数(R2)来反映拟合优度检验。采用Pearson相关分析探讨林分空间结构与更新幼苗多样性之间的关系。
2.1. 样地调查
2.2. 林分结构指数计算


2.3. 物种多样性计算
2.4. 数据分析方法
-
经统计分析(表2)可知,蒙古栎林林下更新幼苗密度为10 759 株·hm−2,总体更新状况良好,但变异系数较大,为62.6%,说明不同样地间更新差异较大,更新幼苗分布不均匀,更新密度最大的样地为23 600 株·hm−2,最小的样地仅为800 株·hm−2,更新幼苗分布趋向斑块状。更新幼苗的基径为0.53~2.17 cm,苗高为37.6~207.8 cm。
项目
Items平均值
Mean最小值
Min最大值
Max标准误
SE变异系数
CV/%密度
Density/(stems·hm−2)10 759 800 23 600 1 122 62.6 基径
Basal diameter/cm1.23 0.53 2.17 0.08 37.4 苗高
Seedings height/cm104.4 37.6 207.8 7.3 42.2 Table 2. The quantitative characteristics of regeneration seedings in Q. mongolica forest
-
由图1可知,林分断面积对幼苗密度影响显著,随着林分断面积的增加,幼苗密度逐渐增加,当断面积达到31 m2·hm−2,幼苗密度增加趋势逐渐放缓;同时,林分断面积对幼苗的基径和苗高生长影响显著(图2),随着林分断面积的增加,幼苗基径和苗高呈降低趋势(R2 = 0.36,P < 0.05;R2 = 0.34,P < 0.05)。
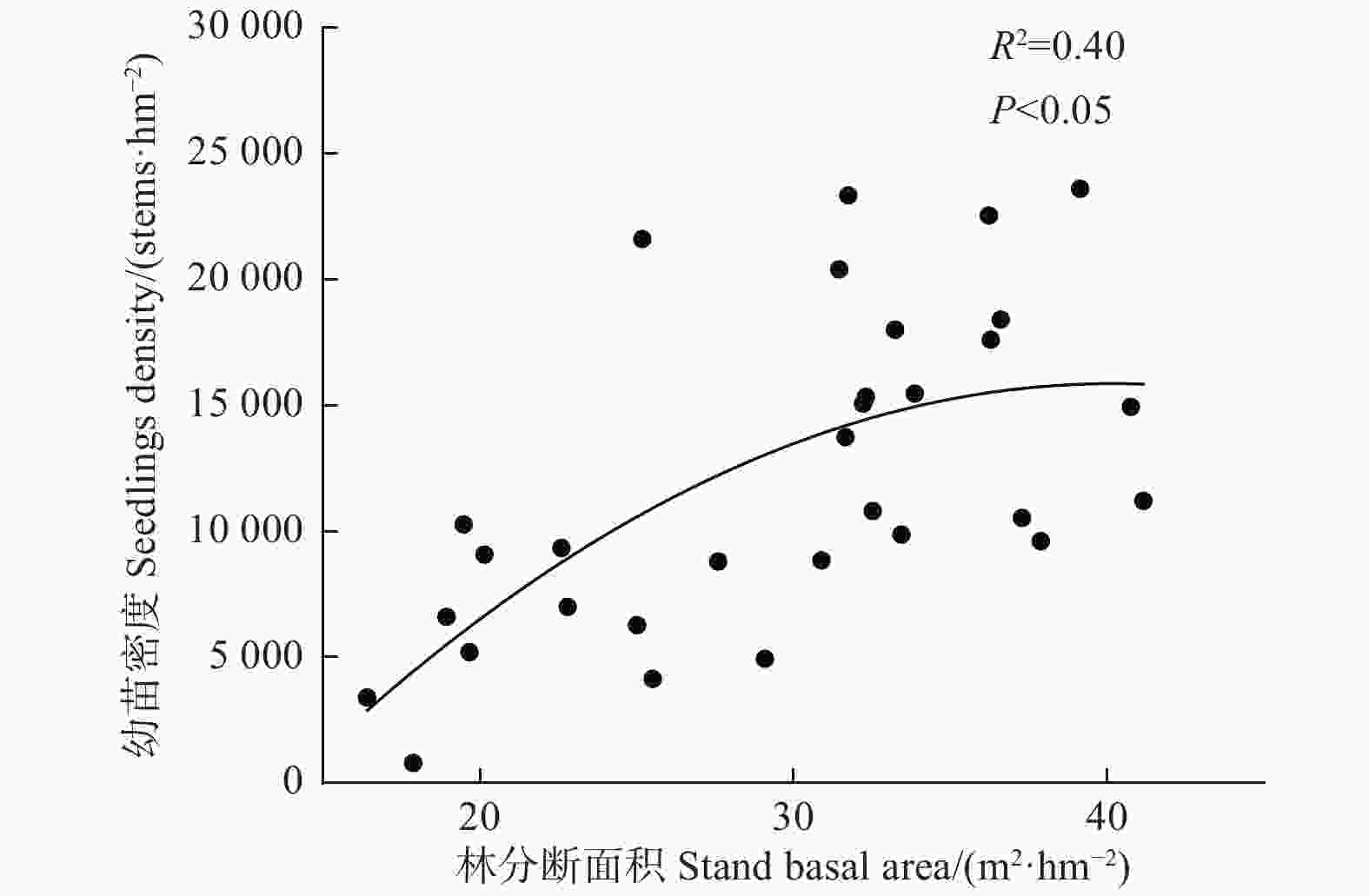
Figure 1. Relationships between stand basal area and seedling density
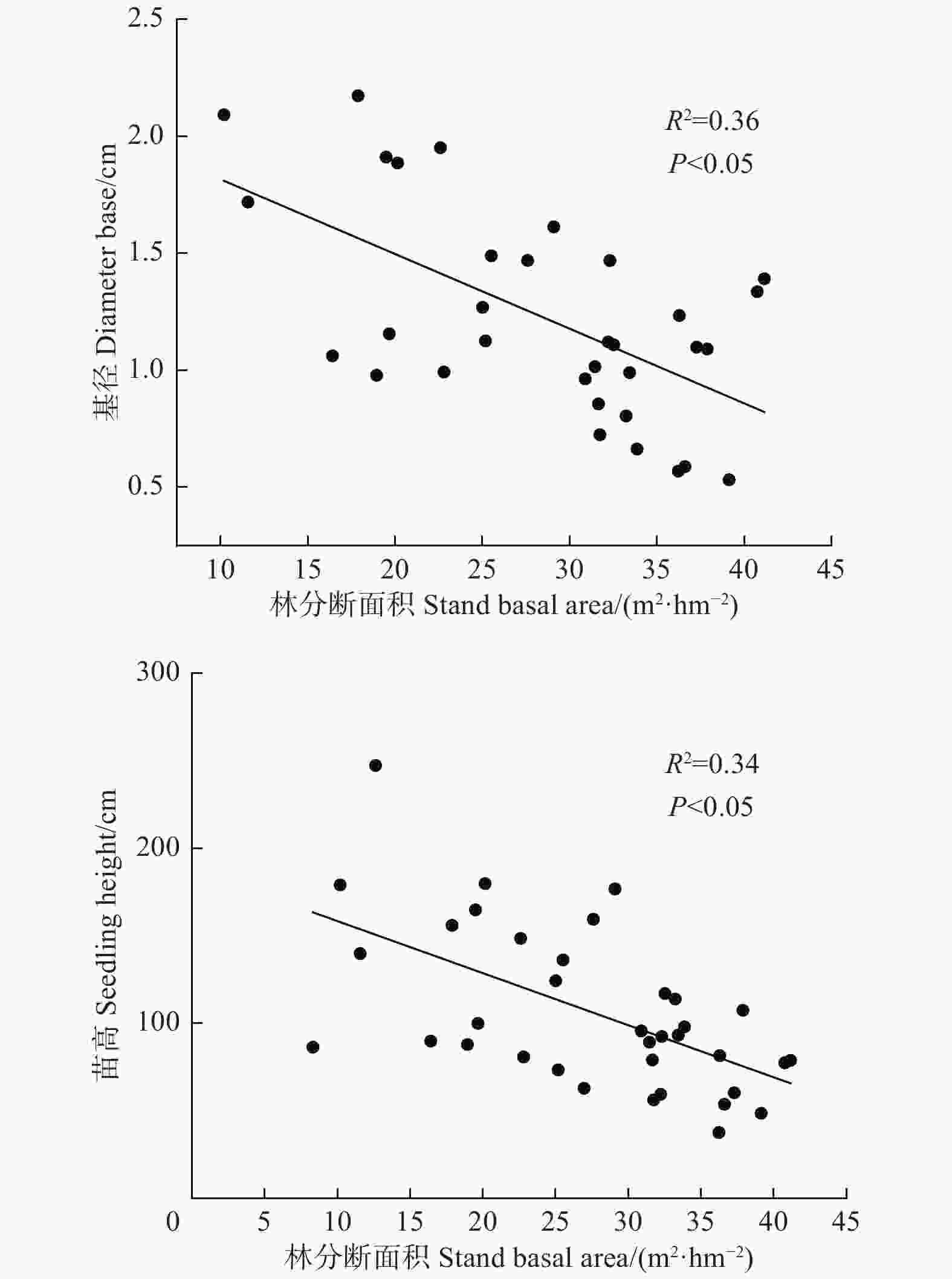
Figure 2. Relationships between stand basal area and seedling growth
-
空间结构指数(混交度、角尺度、竞争指数、林层指数、密集度)与更新幼苗密度做拟合曲线见图3。结果显示:竞争指数和密集度对更新幼苗密度影响显著,随竞争指数和密集度的增加,幼苗密度先增加后降低,表现为典型的抛物线关系,但竞争指数与更新幼苗密度的拟合程度更高(R2 = 0.4),表明与密集度相比,林分竞争指数对更新幼苗密度的影响效应更显著。其他空间结构指数对更新幼苗密度的影响均未达到显著水平(P > 0.05)。进一步分析空间结构指数对幼苗生长影响,由图4可知,竞争指数与幼苗的基径和苗高呈显著负相关(P < 0.05),随着竞争指数增加,幼苗基径和苗高逐渐降低,其他空间结构指标对更新幼苗生长影响不大(P > 0.05)。
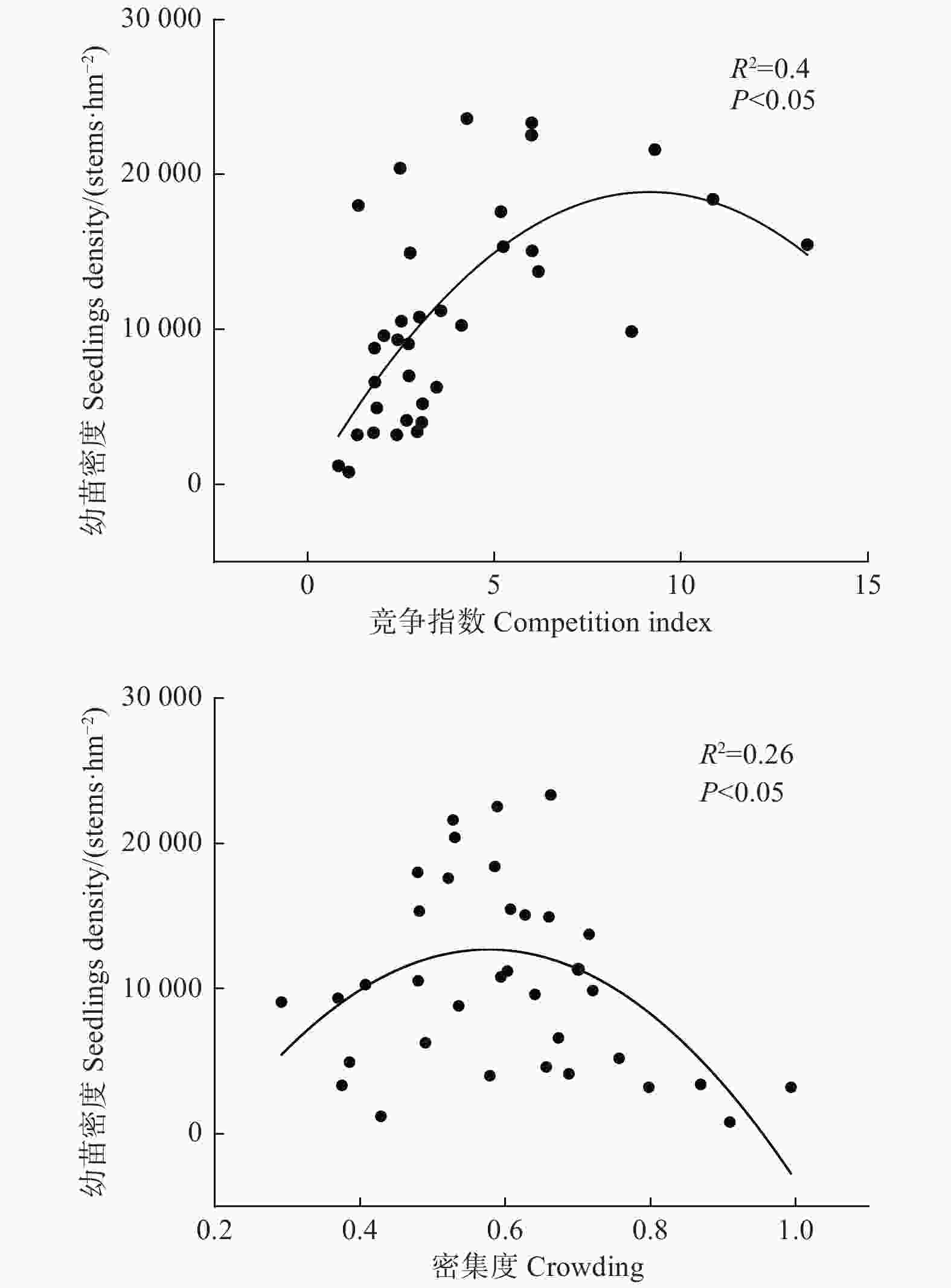
Figure 3. Relationships between indicators of stand spatial structure and seedling density
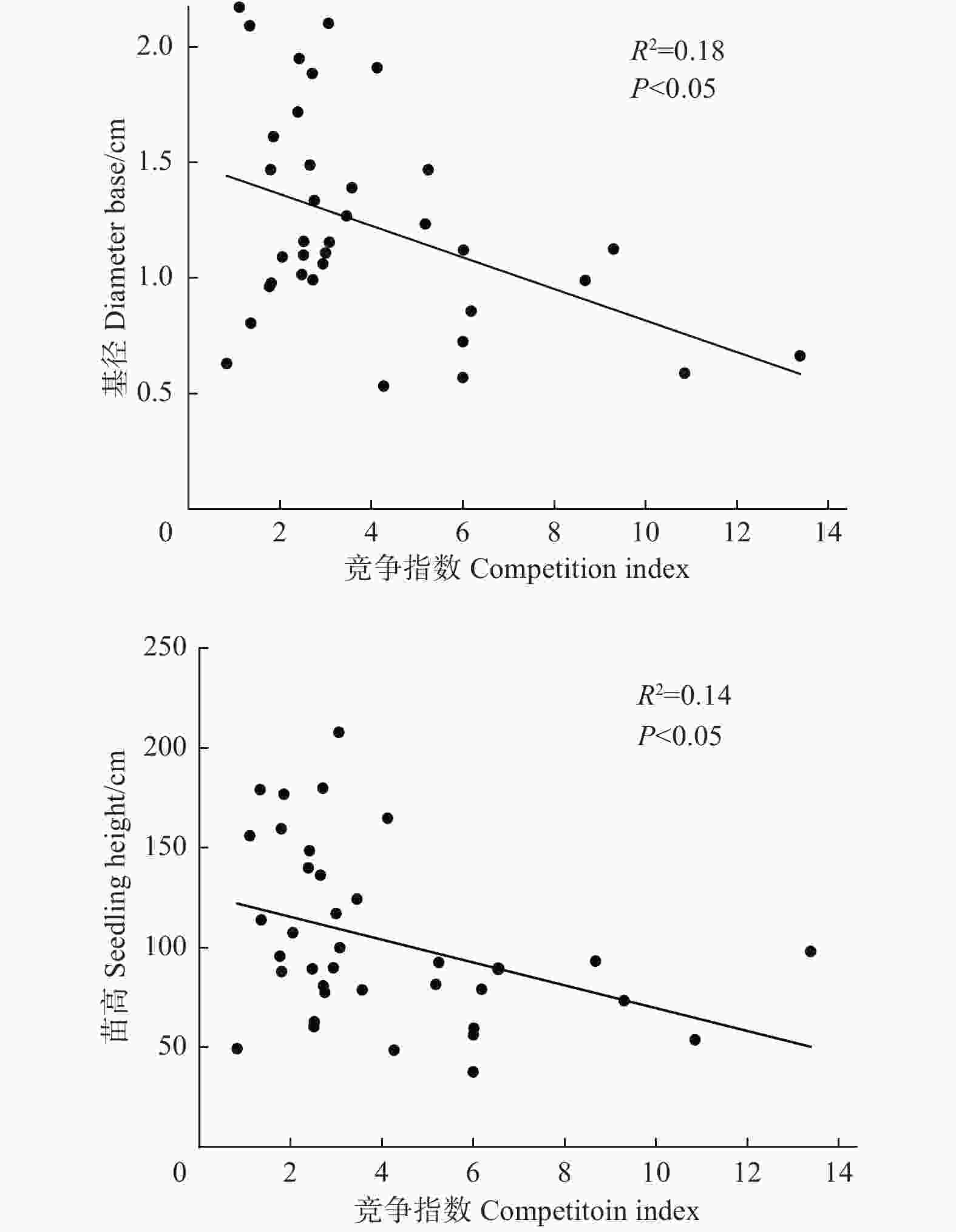
Figure 4. Relationships between competition index and seedling growth
-
对林分空间结构指数(混交度、角尺度、竞争指数、林层指数、密集度)和林下更新幼苗物种多样性指数进行相关分析,结果(表3)表明:林分空间结构对更新幼苗物种多样性影响显著,其中更新幼苗丰富度与林分混交度、竞争指数、林层指数呈极显著或显著正相关(P < 0.01或P < 0.05),与密集度呈极显著负相关(P < 0.01),与角尺度相关性不大;Shannon-Wiener多样性指数分别与林分混交度和林层指数呈极显著和显著正相关(P < 0.01、P < 0.05),与其他空间结构指数相关性不大。Pielou均匀度指数与竞争指数呈极显著负相关(P < 0.01),与其他空间结构指数相关性不大。说明林分种间隔离程度越大,林分垂直结构越复杂,林下更新幼苗的种类和多样性越高。
空间结构指标
Forest spatial index丰富度
SP Shannon-Wiener指数
Shannon-Wiener diversity indexP Pielou均匀度指数
PielouP 混交度 Mingling index 0.485 ** 0.002 0.552 ** 0.001 −0.103 0.542 角尺度 Uniform angle index 0.057 0.739 0.197 0.258 0.004 0.982 竞争指数 Competition index 0.411 * 0.012 0.059 0.737 −0.428 ** 0.008 林层指数 Canopy layer index 0.465 ** 0.004 0.350 * 0.040 −0.282 0.090 密集度 Crowding −0.462 ** 0.004 −0.251 0.145 0.135 0.426 *P < 0.05,**P < 0.01. Table 3. Pearson correlation analysis of stand spatial structure and tree species diversity of undergrowth regeneration
-
以更新幼苗密度为因变量(y),密度因素和空间结构指标为自变量(x)进行多元逐步回归分析(表4),结果显示,林分断面积、密集度和竞争指数是影响更新幼苗密度的主要因子,3个变量能够解释更新幼苗密度变异的41%。
项目
Item回归系数
Regression
coefficient标准化系数
Standardization
coefficientt值
t valueP 常数 Constant 7 105.365 1.206 0.236 林分断面积 Stand basal area 293.020 0.345 2.309 0.027 密集度 Crowding −11 810.751 −0.273 −1.921 0.063 竞争指数 Competition index 885.678 0.308 2.189 0.036 R2 = 0.41,P = 0.000 4 Table 4. The optimal multiple regression models with stepwise of seedling density and forest structure
3.1. 蒙古栎林更新幼苗数量特征
3.2. 密度因素对更新的影响

3.3. 林分空间结构对更新的影响
3.3.1. 林分空间结构对幼苗密度和生长的影响


3.3.2. 林分空间结构对幼苗物种多样性的影响
3.4. 密度因素和林分空间结构对幼苗更新的影响
-
林分密度变化形成了群落中不同的光、热、水等小环境[24],林下乔木幼苗的更新对林分密度反映剧烈。本研究表明,当林分基面积从16.41 m2·hm−2增加到31 m2·hm−2,幼苗密度逐渐增加,当林分基面积从31 m2·hm−2增加到41.75 m2·hm−2时,幼苗密度增加趋势逐渐放缓。黄朗等[25]对枹栎甜槠混交林幼苗更新和张志东等[14]对黑松种群更新的研究中也得出相似的结论。林下乔木幼苗具有一定的耐阴性,林分基面积较低时,过强光照抑制幼苗的发育。另一方面,幼苗的发育,需要一定量的水分支撑,密度较小的林分,蒸腾量较大,林地水分比较缺乏,不利于幼苗的构建,幼苗数量较少[26]。随着林分基面积的增加,林地微环境逐渐改善,母树结实能力增强,更新幼苗密度逐渐增加,但当林分基面积增加到一定程度,受林地资源配置的影响,幼苗密度随基面积的增加速度逐渐放缓。虽然幼苗密度随基面积的增加呈增加趋势,但幼苗基径和树高随基面积的增加均表现为降低趋势,这与Kabrick等[27]对短叶松更新影响因素的研究中得出的结论相同,可能是由于在林分密度较小时,林下光照和养分含量充足,有利于幼苗的生长,随着林分密度的增加,幼苗之间竞争加剧,分配到幼苗个体的可利用资源逐渐减少,从而影响幼苗的生长。
-
林木在林地上的水平或垂直分布及其属性在空间上的排列方式,可促进异质性微生境的形成、调节林内小气候,影响林木生长和稳定,在很大程度上决定着更新幼苗的分布和存活[12]。本研究结果表明,随着林分竞争指数和密集度的增加,更新幼苗密度呈先增加后降低的趋势,在林分竞争指数为4~6,密集度为中等条件下,更新幼苗密度最大,随后更新幼苗密度逐渐减少。这可能是因为当竞争指数和密集度增加到一定程度时,林木之间的竞争加剧,分配到林下的光照和养分不足,从而不利于更新幼苗的建成和存活,同时也说明过高或过低的竞争指数和密集度均不利于幼苗密度的增加。林分空间结构不仅影响更新幼苗密度,而且对更新幼苗多样性也具有一定的影响。研究结果显示,林分混交度与更新幼苗的丰富度、多样性指数呈显著正相关,随着林分混交度的增加,更新幼苗的丰富度、多样性指数也相应的增大。一方面是由于混交度高的林分,乔木树种丰富,能够为更新幼苗提供丰富的母树,另一方面混交度高低影响着乔木层各个树种的竞争程度,不同树种的生态位在一定程度上会互相抑制[28],为林下幼苗的更新释放了营养空间。林分的垂直结构对更新幼苗的多样性也具有一定的影响,研究表明,林层指数与更新幼苗的丰富度和多样性指数均呈显著的正相关关系,一方面是由于复杂的垂直结构促进异质性微生境的形成[29],从而增加幼苗定居的生态位,为更多幼苗的萌发和生长提供机会和空间,使森林容纳更多物种[30]。另一方面,树冠层深厚,容易形成较多的枯枝落叶,能够改善土壤的理化性质,提高土壤的养分含量,有利于林下更新幼苗的生长[31]。
4.1. 密度因素对更新幼苗的影响
4.2. 林分空间结构对更新幼苗的影响
-
辽东山区蒙古栎林乔木更新由密度因素和林分空间结构共同影响,随着林分断面积的增加,幼苗密度逐渐增加,当断面积达到31 m2·hm−2,幼苗密度增加趋势逐渐放缓;林分空间结构对更新幼苗的影响主要表现为随竞争指数和密集度的增加,幼苗密度呈先增加后降低趋势,林分种间隔离程度越大,林分垂直结构越复杂,林下更新幼苗的种类和多样性越高。因此,欲人工促进蒙古栎林天然更新,改善林下物种多样性,可通过调控林分断面积、竞争指数和密集度等经营方式来实现。
 DownLoad:
DownLoad: