




-
昆虫的化学感受系统在其生存和繁殖过程中起着极其重要的作用[1-3]。在过去10多年的时间里,研究者对昆虫触角嗅觉信号传导的分子机制研究有了突出的进步。在昆虫嗅觉识别时,气味分子从触角感器孔渗入,然后被气味结合蛋白(odorant binding proteins,OBPs)或化学感受蛋白(chemosensory proteins,CSPs)识别和转移,最后激活位于嗅觉感觉神经元(olfactory sensory neurons,OSNs)树突膜上的嗅觉受体(odorant receptors,ORs)或离子型受体(ionotropic receptors,IRs),产生电位,指导昆虫做出相应的行为反应[4-8]。此外,还有一些蛋白如感觉神经元膜蛋白(sensory neuron membrane proteins,SNMPs)在昆虫气味识别过程中也扮演着至关重要的作用[5]。
昆虫SNMPs也是一种膜蛋白,与脊椎动物CD36家族为同源基因,具有2个跨膜区域,其功能主要是识别和转运亲脂性气味分子如脂肪酸和脂类化合物等[9-14]。昆虫第一个SNMP基因在多音天蚕(Antheraea polyphemus Cramer)中被鉴定,并命名为ApolSNMP1[9]。随后在烟草天蛾(Manduca sexta L.)中发现SNMP的第二个亚类型,即命名为MsexSNMP2[10, 15]。紧接着SNMP的同源基因在鳞翅目Lepidoptera[10, 15-16]、双翅目Diptera[17]、鞘翅目Coleoptera[18]、直翅目Orthoptera[13]和膜翅目Hymenoptera[19]等昆虫中都有发现。一直以来被认为SNMP基因家族就2个成员,即SNMP1和SNMP2。最近昆虫SNMP家族的第三个成员SNMP3在鳞翅目中被鉴定[20-21],但认为该基因的主要功能与昆虫的免疫反应有关,这还需要进一步的去证明。
红脊长蝽(Tropidothorax elegans Distant)属半翅目(Hemiptera)长蝽科(Lygaeidae),主要为害刺槐(Robinia pseudoacacia L.)、辣椒(Capsicum annuum L.)、葫芦(Lagenaria siceraria Molina)、油菜(Brassica napus L.)、大白菜(Brassica pekinensis Lour.)和小麦(Triticum aestivum L.)等多种植物,食性较杂。关于红脊长蝽的嗅觉基因研究较少,本研究通过前期红脊长蝽触角转录组测序结果[22],鉴定了红脊长蝽的2个SNMP基因,即TeleSNMP1和TeleSNMP2,并通过荧光定量PCR技术对红脊长蝽TeleSNMP1和TeleSNMP2在不同组织中的表达情况进行分析,为进一步探索红脊长蝽SNMPs的化学通讯功能奠定基础。
HTML
-
红脊长蝽来自河南科技大学林学院昆虫实验室,该群体为自2014年7月在洛阳周边(112˚26′ E,34˚43′ N)蔬菜地采集的成虫,然后在温室内继代饲养至今。温室条件为:温度25 ± 2℃,相对湿度60% ± 5%,光周期14L∶10D。
-
选取红脊长蝽羽化后第3 d的雌雄成虫,收集触角各100头、头部各30头、胸部各20头、腹部各5头、足各50头、翅各50头,每个样品收集材料重复3次。将红脊长蝽各部分组织解剖后立即放入浸在液氮中的1.5 mL离心管内,然后保存于−80℃中。
总RNA的提取采用RNAiso Plus Kit(TaKaRa,北京)试剂盒进行,并使用RNase-free DNase I(TaKaRa,北京)对提取的RNA进行除DNA处理。采用1%的琼脂糖凝胶电泳和NanoDrop 2000c分光光度计(Thermo Scientific)进行质量检测。采用PrimeScript™ 1st Strand cDNA Synthesis Kit(TaKaRa,北京)试剂盒对RNA进行反转录。
-
从本实验室前期对红脊长蝽触角转录组测序注释结果中[21]搜素到2个SNMP基因。根据该序列设计特异性引物,TeleSNMP1-F:5′-ATGGCTGCACCACTGAGG-3′,TeleSNMP1-R:5′- CTAGTACTTTGCCGGGGGTG-3′;TeleSNMP2-F:5′-ATGACGAAGGTGCTGTTCCC-3′,TeleSNMP2-R:5′-TTAGCTTGTGAGAGTCCTTTTGA-3′,进行PCR扩增。PCR反应体系为20 μL:雌蛾触角cDNA模板1 μL,上下游引物各1.5 μL(10 μmol·L−1),dNTPs混合液1.6 μL(2.5 μmol·L−1),Ex Taq DNA聚合酶0.2 μL(TaKaRa,大连),10 × Ex Taq buffer 2 μL,ddH2O为12.2 μL。PCR反应条件为:94℃ 5 min;94℃ 30 s,58℃ 30 s,72℃ 30 s,35个循环;最后72℃ 10 min。胶回收目的片段,将目的片段克隆到pMDTM19-T载体上,转化DH5α感受态细胞,挑取阳性克隆培养过夜,送去测序。
-
核酸序列采用在线工具(
http://www.bio-soft.net/sms/ )翻译;采用(http://www.cbs.dtu.dk/services/TMHMM )在线工具进行跨膜区域预测;利用在线工具(https://web.expasy.org/protparam/ )对蛋白序列特性进行分析;蛋白亲疏水性采用在线工具(https://web.expasy.org/protscale/ )分析;序列比对采用线下DNAMAN进行多重序列比较;进化树采用线下MEGA 6.0构建。 -
通过荧光定量PCR检测红脊长蝽SNMPs基因在不同组织中的表达情况。根据红脊长蝽SNMPs基因的开放阅读框和荧光定量引物设计原则设计特异性引物,TeleSNMP1-F:TCACCATCCCTCATCCA,TeleSNMP1-R:TCTTCGCCGTCTTTCAT,TeleSNMP2-F:TGTGGGAACGGAACTCT,TeleSNMP2-R:GCACCTTGGCACTTTG。内参基因为红脊长蝽Actin基因(基因登陆号为MG322127),引物为:TeleActin-F:CAAGGACGAAACAATCA;TeleActin-R:GAGAATACACTCCCAGAAC。
荧光定量PCR被执行在ABI 7500 PCR仪(ABI,Carlsbad,CA,USA)上进行,每个反应体积20 μL,包括10 μL的2 × SYBR Green PCR Master Mix(TaKaRa,大连)、0.8 μL的正反向引物(10 μmol·L−1)、2 μL的cDNA模板(200 ng)和6.4 μL的DEPC水。PCR循环遵循95℃ 30 s,然后95℃ 5 s,53℃ 31 s,共循环40个周期。试验重复3次。
-
红脊长蝽SNMPs基因在不同组织中的相对表达量采用公式2−∆∆CT[23]计算。利用SPSS 17.0软件中的ANOVA方法对TeleSNMPs在红脊长蝽不同组织中的表达量进行显著性差异比较(新复极差法检验,P ≤ 0.05)。
1.1. 试虫的准备
1.2. 总RNA的提取与第一链cDNA的合成
1.3. 红脊长蝽SNMPs基因的克隆
1.4. 红脊长蝽SNMPs基因的序列分析
1.5. 实时定量PCR
1.6. 数据分析
-
将测序结果在NCBI上进行同源性搜索,结果表明所测序列与多种昆虫的SNMP基因序列高度同源,证明这2个基因就是红脊长蝽的SNMP基因,并分别命名为TeleSNMP1和TeleSNMP2,在NCBI基因登陆号分别为MW442946和MW442947。
这2个SNMPs基因都具有全长的开放阅读框,TeleSNMP1长度为1 497 bp,编码498个氨基酸(图1);而TeleSNMP2长度为1 686 bp,编码561个氨基酸。这2个基因都是酸性,TeleSNMP1的等电点是8.26,蛋白分子量是55.59 kD;TeleSNMP2的等电点是7.05,蛋白分子量是63.37 kD。
TeleSNMP1和TeleSNMP2在氨基酸序列的N-端和C-端各有一个跨膜区域,其中TeleSNMP2在氨基酸第158和180之间还有一个跨膜区域。在膜外区域6个保守的半胱氨酸残基位点被预测(图1),这6个保守的半胱氨酸位点与CD36基因家族相似。
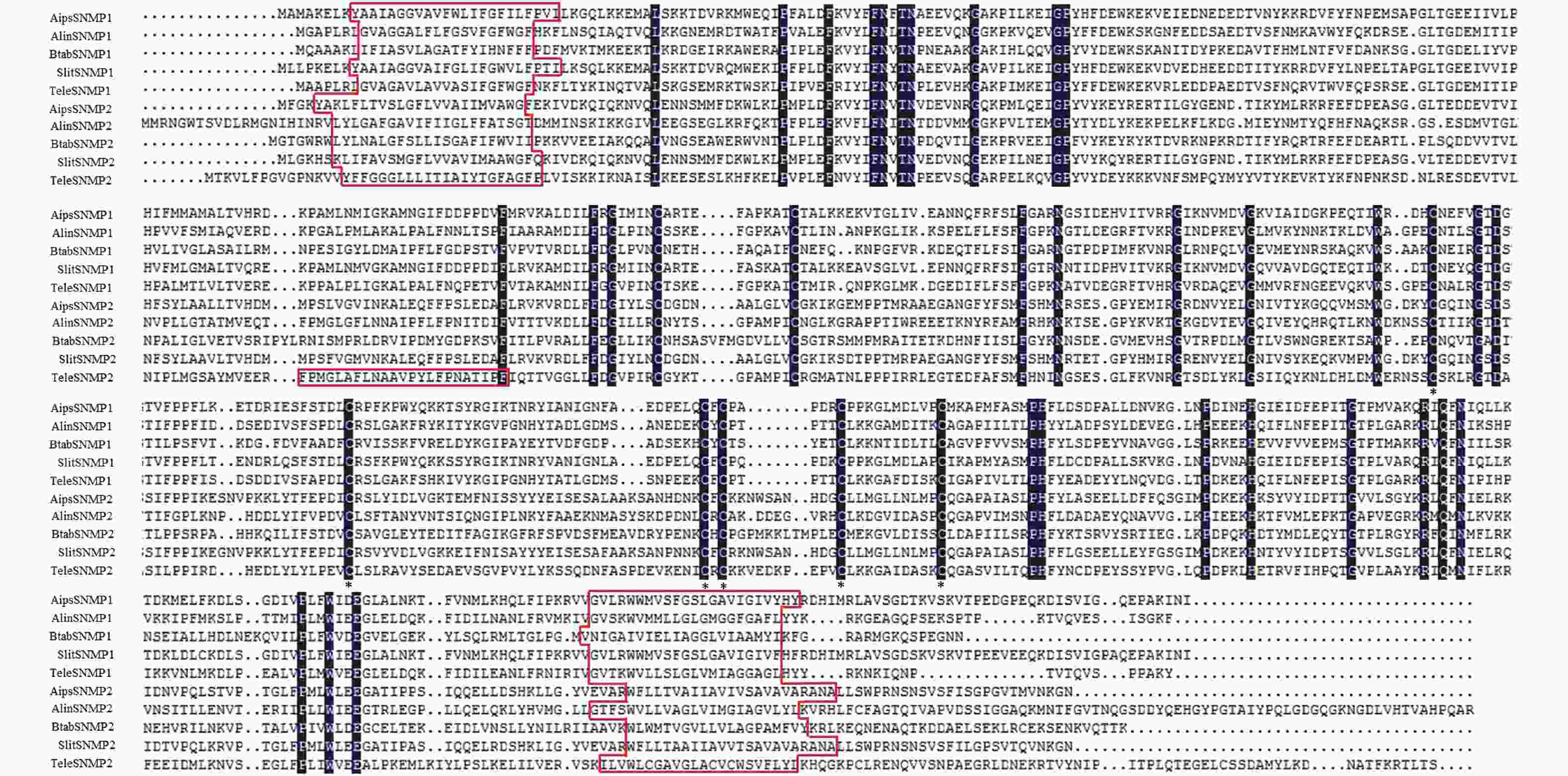
Figure 1. Alignment of Tropidothorax elegans SNMP1 and SNMP2 with SNMPs from other insects
-
将红脊长蝽TeleSNMP基因与已报道的其他昆虫SNMP基因的氨基酸序列在NCBI中的Blastp在线搜索工具进行同源性分析,发现TeleSNMP基因与不同种类昆虫SNMP基因一致性差别较大。与同目昆虫SNMP基因一致性较高,如TeleSNMP1与茶翅蝽 (Halyomorpha halys Stal) HhalSNMP1序列一致性在81.33%;与苜蓿盲蝽 (Adelphocoris lineolatus Goeze) AlinSNMP1序列一致性在68.54%;与不同目昆虫SNMP基因一致性较低,如TeleSNMP1与斑痣悬茧蜂 (Meteorus pulchricornis Wesmael) MpulSNMP1序列一致性在43.44%。但TeleSNMP2与所有昆虫SNMP基因一致性都不高,如与茶翅蝽HhalSNMP2序列一致性在48.68%;与苜蓿盲蝽AlinSNMP2序列一致性在41.18%;与德国小蠊 (Blattella germanica Linnaeus) BgerSNMP1序列一致性在46.22%。TeleSNMP2与玉带凤蝶 (Papilio polytes L.) PpolSNMP2序列一致性在30.94%;与亚洲小车蝗 (Oedaleus asiaticus Bei-Bienko) OasiSNMP2序列一致性在34.76%。红脊长蝽TeleSNMP1和TeleSNMP2之间序列一致性仅有29.66%。
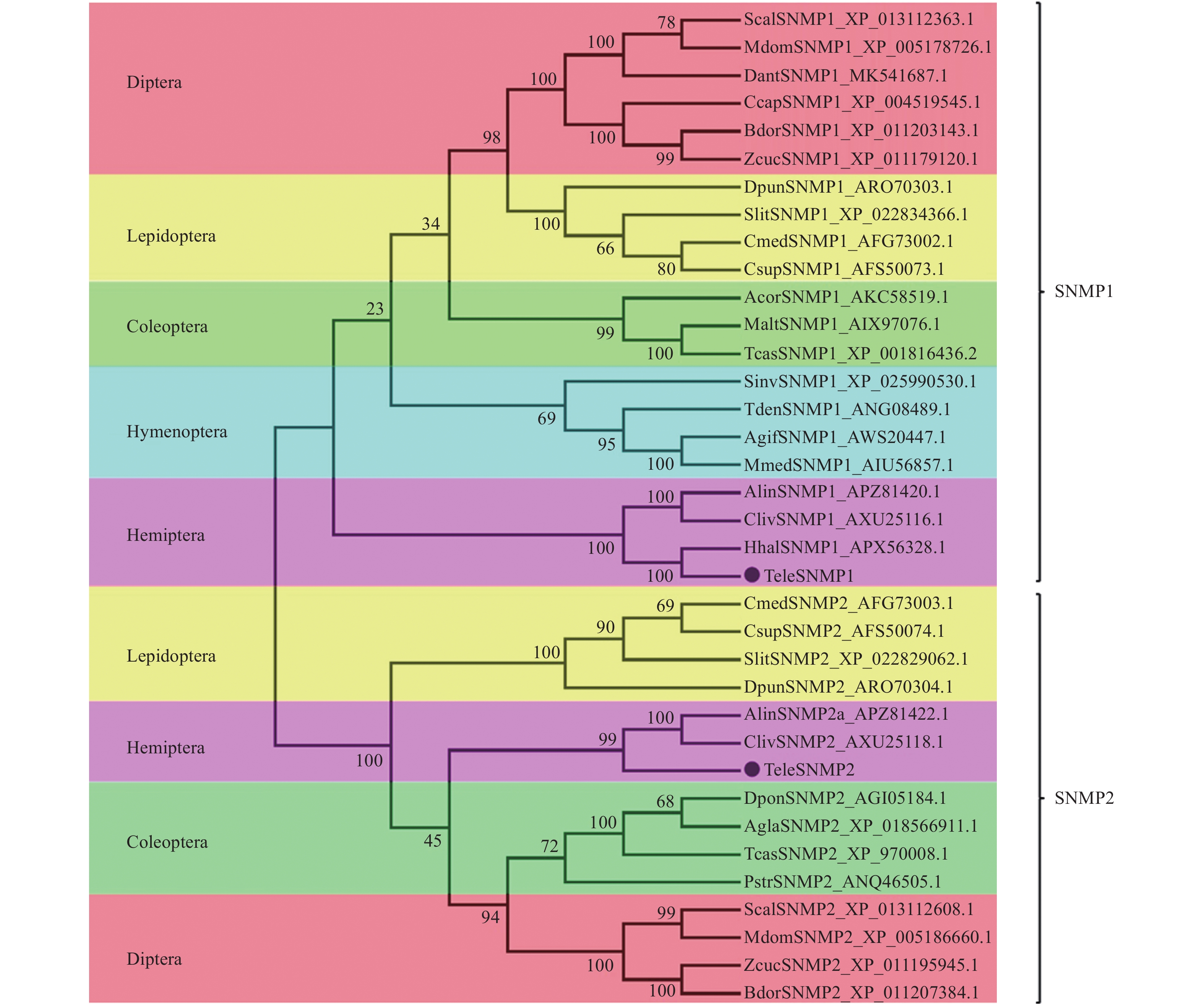
使用MEGA6.0对9个目的36种昆虫SNMP同源基因进行进化树比较。SNMPs基因被分成2个亚组,即SNMP1和SNMP2(图2),每亚组中同目昆虫SNMP基因同源性最高。红脊长蝽的TeleSNMP1和TeleSNMP2基因分别被集聚到这2个亚组,并且与同为半翅目昆虫的苜蓿盲蝽、茶翅蝽和黑肩绿盲蝽SNMP基因进化关系最近,与其他目昆虫SNMP关系较远。
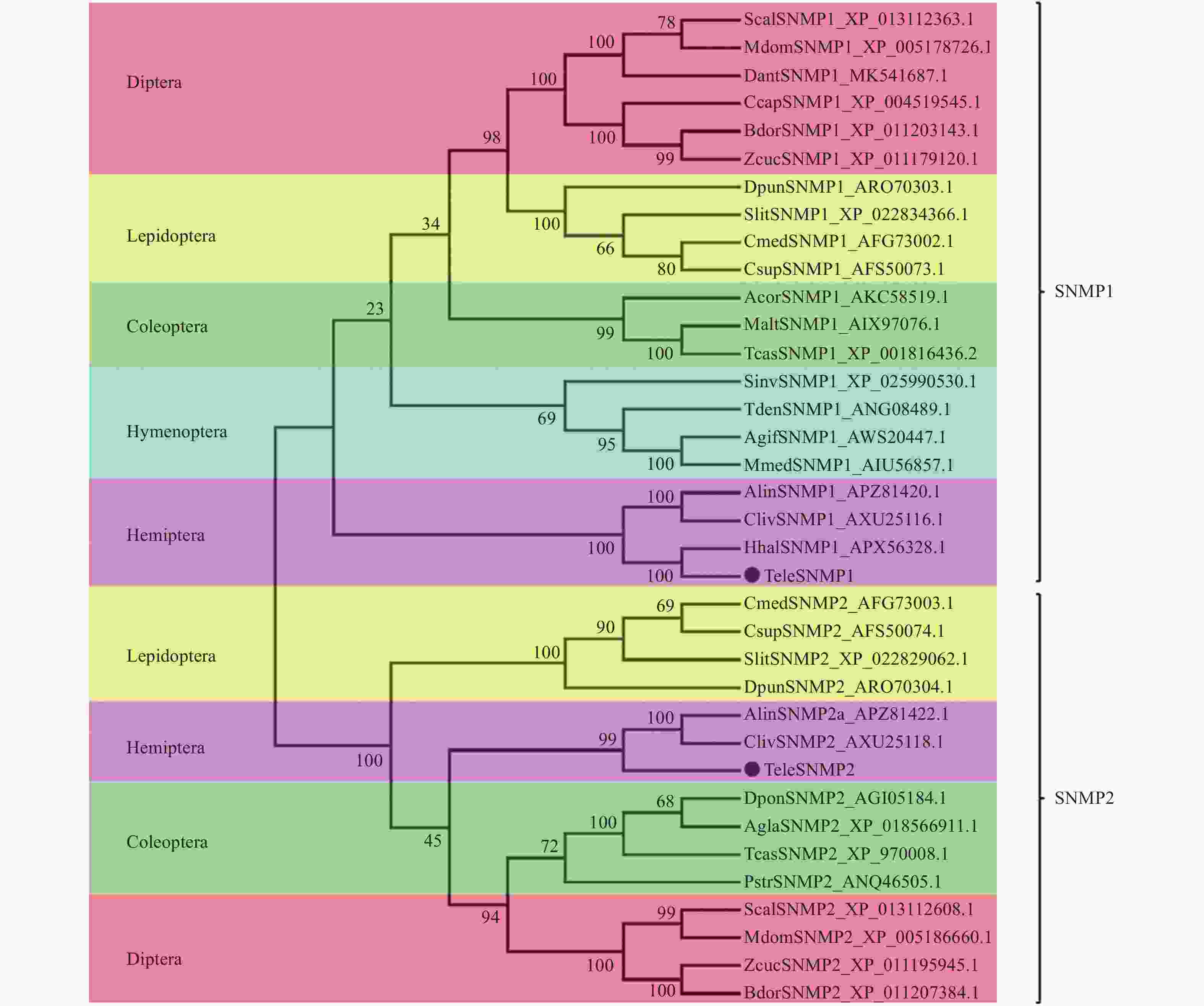
Figure 2. Phylogenetic tree of the SNMPs of Tropidothorax elegans and other insects based on amino acid sequences by using neighbor-joining method
-
使用荧光定量PCR对红脊长蝽TeleSNMP1和TeleSNMP2在不同组织中的表达情况进行分析,结果发现:TeleSNMP1和TeleSNMP2在雌雄蛾触角中高度表达,TeleSNMP1在雄蛾触角中的表达量明显高于雌蛾,TeleSNMP2的表达情况刚好相反,即TeleSNMP2在雌蛾触角中的表达量明显高于雄蛾。除触角外,TeleSNMP1几乎不在其他组织中表达或表达量甚微。TeleSNMP2除在触角中表达量丰富外,在足和翅中也有少量表达(图3)。
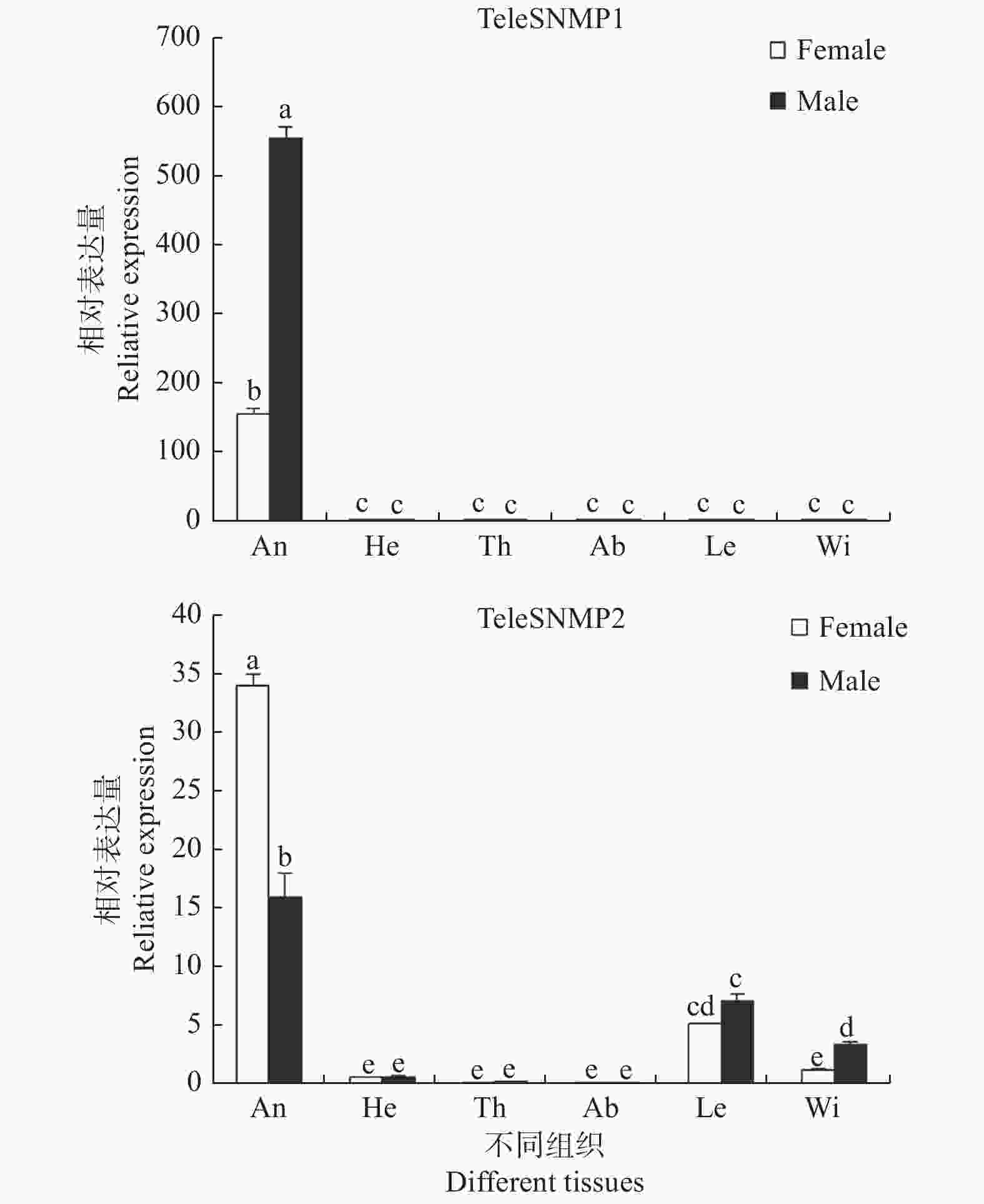
Figure 3. Expression level of SNMPs in different tissues of Tropidothorax elegans
2.1. 红脊长蝽SNMPs基因克隆及序列分析
2.2. 红脊长蝽SNMPs与其他昆虫SNMP基因的同源性及进化树分析
2.3. 红脊长蝽SNMPs基因表达谱分析

-
本研究根据前期红脊长蝽触角转录组的数据,通过BLASTX在线搜索和同源性比较鉴定出2个SNMPs基因,即TeleSNMP1和TeleSNMP2。这2个基因与其他昆虫SNMPs具有类似的特征,如在氨基酸序列N端和C端附近有2个保守的跨膜区域,并且由6个保守的半胱氨酸残基形成二硫键组成一个大的胞外环,此结构也与CD36基因家族极其相似[24-26]。根据对这2个跨膜蛋白结构投影预测,这部分的功能是转运和结合脂类分子[27-28],我们可推测红脊长蝽SNMPs的2个跨膜区具有同样的功能。但红脊长蝽TeleSNMP2在氨基酸序列第158和180之间多了1个跨膜区域,目前关于SNMPs基因具有3个跨膜区域的报道还没有,可能是鉴定的SNMPs数量还不够庞大,也可能长期进化形成的。
红脊长蝽TeleSNMP基因同源性搜索发现TeleSNMP基因与不同种类昆虫SNMP基因一致性差别较大。与同目昆虫SNMP基因一致性较高,与不同目昆虫SNMP基因一致性较低。红脊长蝽TeleSNMP1和TeleSNMP2之间分歧也比较大。进化树结果也显示,红脊长蝽TeleSNMP1和TeleSNMP2分别被集聚到SNMP1和SNMP2两个亚组,在同一组内同目昆虫的SNMP基因进化关系最近,与其他目昆虫SNMP关系较远。此结果与大多数昆虫SNMP基因特性相同[26,29-30]。在鳞翅目中曾经鉴定出SNMP3亚家族基因[20],而在红脊长蝽触角转录组数据中没有发现该基因,可能是这个基因在幼虫肠中高度表达的原因。
组织特异性表达可以为功能预测提供可靠性的参考。我们研究发现红脊长蝽SNMP1主要表达在雌雄蛾的触角中,此结果与多音蚕蛾[9]、脐橙螟(Amyelois transitella Walker)[31]、甜菜夜蛾(Spodoptera exigua Htibner)[32]和中红侧沟茧蜂(Microplitis mediator Haliday)[33]等多种昆虫SNMP1的表达模式相同。Benton等[34]在果腹黑蝇(Drosophila melanogaster wDm)中发现,DmelSNMP1能够识别集合信息素cVA,激活受体HR13。Pregitzer等[28]在烟芽夜蛾(Heliothis virescens Fabricius)中也发现,HvirSNMP1能明显增强HR13对信息素Z11-16:Ald的结合力。以上研究都暗示昆虫SNMP1的功能是调节信息素的识别和通过SNMP-OR互作转运信息素。相对于TeleSNMP1,TeleSNMP2的表达相对广泛,除嗅觉感器触角外,在非嗅觉感器足中也有少量表达,此结果与小菜蛾(Plutella xylostella L.)[35]、二化螟(Chilo suppressalis Walker)[36]、稻纵卷叶螟(Cnaphalocrocis medinalis G.)[37]、甜菜夜蛾[31]和中红侧沟茧蜂[32]一致。昆虫身体上分布有大量的味觉感器[38],而CD36蛋白的主要功能是转运脂类化合物,因此可推测TeleSNMP2和GRs在这些组织中共同表达,识别脂类分子完成味觉过程。
-
本研究首次克隆和鉴定了红脊长蝽的2个SNMPs基因,即TeleSNMP1和TeleSNMP2。同目昆虫同类SNMP同源序列一致性较高,反之,不同目昆虫不同类SNMP同源序列一致性较低。同样,TeleSNMP1和TeleSNMP2之间的序列一致性也极低。TeleSNMP1主要在雌雄触角中特异性表达,而TeleSNMP2除触角外,在非触角组织中也有表达。
 DownLoad:
DownLoad: