



-
肿腿蜂属膜翅目(Hymenoptera)青蜂总科(Chrysidoidea)肿腿蜂科(Bethylidae),是我国林木蛀干类天牛和吉丁虫等害虫生物防治中应用广泛的一类寄生性天敌昆虫[1]。其中管氏肿腿蜂(Sclerodermus guani)已有40余年的研究和应用历史,是防治松褐天牛(Monochamus alternatus)和双条杉天牛(Semanotus bifasciatus)的重要天敌[2]。我国是世界上少数将肿腿蜂类寄生蜂应用到林木蛀干害虫防治的国家[3],除管氏肿腿蜂外,川硬皮肿腿蜂(S. sichuanensis)[4]和白蜡吉丁肿腿蜂(S. pupariae)[5]也在近年有较多应用,前者多被用于防治松褐天牛,后者多被用于防治白蜡窄吉丁(Agrilus planipennis)、栗山天牛(Massicus raddei)和光肩星天牛(Anoplophora glabripennis)等害虫。松褐天牛肿腿蜂(S. alternatusi)是近年来发现的肿腿蜂新种,原始种群发现于自然被寄生的松褐天牛低龄幼虫体上[6],其个体较管氏肿腿蜂和川硬皮肿腿蜂更大,雌蜂多具翅。目前,松褐天牛肿腿蜂已被用于松褐天牛[7-8]和光肩星天牛[9]的生物防治。
肿腿蜂均属多寄主型寄生蜂[10],已报道自然寄主以天牛类、吉丁虫类和象甲类蛀干害虫为主。在肿腿蜂的繁育中,原始记录的自然寄主均可用于繁蜂。一些替代寄主,如黄粉虫(Tenebrio molitor)[11]、意大利蜜蜂(Apis mellifera)[12]、大理窃蠹(Ptilineurus marmoratus)[13]、玉米螟(Pyrausta nubilalis)[14]、红铃虫(Pectinophora gossypiella)[15]、大袋蛾(Clania vartegata)[16]等也有被报道用于繁蜂。事实上,以上各种被报道的替代寄主除黄粉虫在肿腿蜂的繁育中会被使用外,大多数并不常用。在肿腿蜂的繁育中,目前常用的寄主有黄粉虫、青杨天牛(Saperda populnea)、双条杉天牛和麻竖毛天牛(Thyestilla gebleri)。黄粉虫主要被用于繁育管氏肿腿蜂和川硬皮肿腿蜂[11, 17],青杨天牛主要被用于繁育管氏肿腿蜂[18],双条杉天牛主要被用于繁育管氏肿腿蜂[3],白蜡吉丁肿腿蜂和松褐天牛肿腿蜂多用麻竖毛天牛繁育[8]。上述除黄粉虫的幼虫不能被肿腿蜂寄生,需以蛹用于繁蜂,其他替代寄主多以幼虫用于繁蜂。在上述几种常用的繁蜂寄主中,麻竖毛天牛个体较大,可提供的营养更多,单头寄主可育得肿腿蜂数较其他几种寄主更多,是当前繁育肿腿蜂最主要替代寄主[5]。然而,当前并没有人工饲料可用于饲养麻竖毛天牛。寄主的获取方式主要为每年幼虫越冬前(9—10月)大量低温储存带虫的麻根,需要繁蜂时再从麻根中剥取天牛幼虫。在长期储存麻根的过程中会面临幼虫化蛹的情况。对于已经化蛹的麻竖毛天牛,是否还能用于肿腿蜂繁育?若用于繁育肿腿蜂,繁蜂效率如何?母蜂的寄生行为、子代发育情况是否较用麻竖毛天牛幼虫作寄主时有差异?均未见评估和报道。探清这些问题是进一步筛选替代寄主和实现寄主资源利用最大化的基础。本研究以松褐天牛肿腿蜂和麻竖毛天牛为研究对象,分析和评价分别以麻竖毛天牛幼虫和蛹繁蜂时,松褐天牛肿腿蜂的寄生行为、繁蜂效率是否存在差异,以期为松褐天牛肿腿蜂的繁育和替代寄主资源的利用策略提供参考。
-
松褐天牛肿腿蜂建群种采自昆明市西南林业大学后山(25°03′ N,102°42′ E),供试蜂为在实验室用麻竖毛天牛幼虫传代繁殖的种群。传代繁殖的方法为将建群蜂接种至松褐天牛低龄幼虫繁育得到肿腿蜂的初始实验室种群,随后用麻竖毛天牛幼虫作寄主续代繁殖肿腿蜂。续代繁殖中,接蜂方式为1头母蜂接种至1头寄主,饲养条件为27 ℃,RH=60%~70%,L∶D=8∶16 h。当每一代子代蜂羽化后挑选3日龄的雌蜂继续接种,获得实验室连续传代种群。
供试寄主为麻竖毛天牛幼虫和蛹。选取健康的麻竖毛天牛老熟幼虫,置于人工气候箱中培养至化蛹。选择蛹龄为1~2 d、体质量为220 ± 20.0 mg的麻竖毛天牛蛹和体质量较蛹相对一致的天牛老熟幼虫各30头作为繁蜂寄主。将麻竖毛天牛蛹和幼虫单头装入玻璃指形管内(直径=1.0 cm,高=5.0 cm),随后接入1头3日龄且已交配的松褐天牛肿腿蜂雌蜂(肿腿蜂雌雄蜂一般在羽化后1~2 d内即完成交尾),并用脱脂棉将指形管口塞紧,饲养条件同上。
-
自接蜂时起,每隔12 h记录1次松褐天牛肿腿蜂对寄主的麻痹情况、初始产卵时间,初孵幼虫出现时间,茧蛹出现时间和子代羽化时间,以此统计松褐天牛肿腿蜂的寄生率、产卵前期和子代发育历期。寄生率为麻痹寄主且产卵母蜂的数量占处理数的比率。产卵前期为接蜂至观察到第1粒卵的间隔时期。子代发育历期共包括卵期、幼虫期、蛹期和幼期历期。其中,卵期为产下第1粒卵至初孵幼虫出现的间隔时期;幼虫期为第1头初孵幼虫至茧出现的间隔时期;蛹期为茧出现至第1头子代蜂羽化的间隔时期;幼期历期为产卵至羽化的间隔时期。子代蜂完全羽化后,统计寄生不同虫态麻竖毛天牛的松褐天牛肿腿蜂的子代羽化率和子代雌雄个体数量。子代羽化率为被寄生的寄主中最终有肿腿蜂子代羽化的比率。依据子代雌雄个体数量计算雄性比,即雄蜂占子代总数的比率。
-
Fisher's精确检验用于比较松褐天牛肿腿蜂寄生不同虫态麻竖毛天牛的寄生率和子代羽化率。松褐天牛肿腿蜂寄生不同虫态麻竖毛天牛的产卵前期、子代发育历期、子代数量和雄性比用单因素方差分析检验处理间差异。经正态性和方差同质性检验,产卵前期、子代发育历期、子代数量满足正态分布且方差齐性,其均直接用原始数据进行差异性检验。子代性比经反正弦化后满足正态分布和方差齐性,转换后的数据用于差异性检验。数据统计用SPSS 20完成。
-
麻竖毛天牛老熟幼虫和蛹均可被松褐天牛肿腿蜂寄生。所有的供试天牛幼虫均被成功寄生,4头天牛蛹未被寄生,松褐天牛肿腿蜂对不同虫态的麻竖毛天牛的寄生率无显著差异(χ2=4.286,df=1,P=0.112)(表1)。在全部被寄生的30头麻竖毛天牛幼虫中,有93.33%的寄主成功育出子代蜂;被寄生的26头天牛蛹中可育出子代蜂的比例为57.69%。用麻竖毛天牛幼虫繁育松褐天牛肿腿蜂时,子代蜂羽化率显著高于用麻竖毛天牛蛹繁蜂(χ2=9.926,df=1,P=0.003)(表1)。
寄主虫态
Host stage供试寄主数
Number of host/n寄生数
Parasitized host/n寄生率
Parasitism rate/%育出子代蜂寄主数
Parasitoid completed development/n子代羽化率
Emergence rate/%幼虫 Larva 30 30 100.00 28 93.33 蛹 Pupa 30 26 86.67 15 57.69 χ2 / / 4.286 / 9.926 P / / 0.112 / 0.003 Table 1. Successful parasitization and progeny emergence of Sclerodermus alternatusi on different stage of Thyestilla gebleri
-
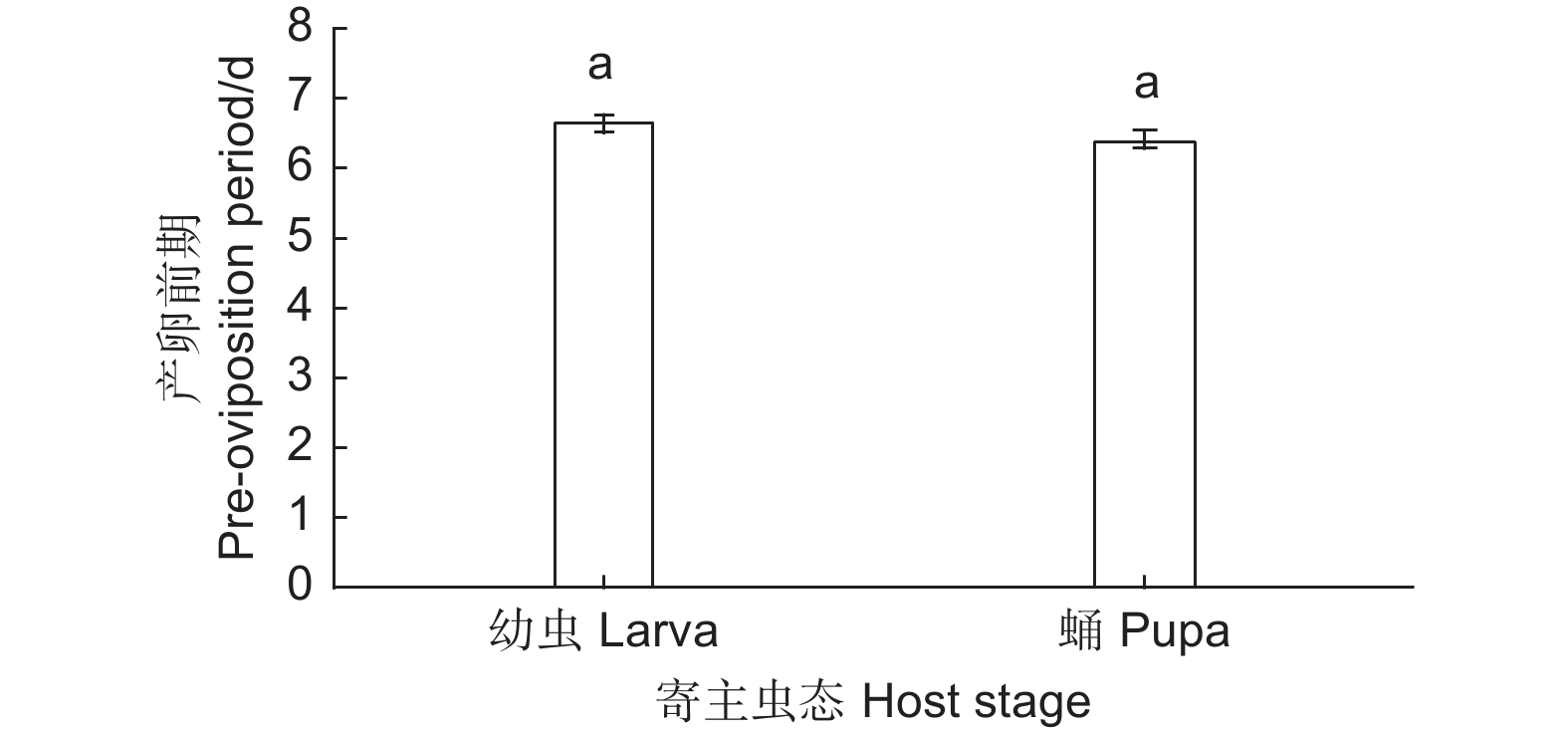
用麻竖毛天牛幼虫和蛹繁育松褐天牛肿腿蜂时,母蜂产卵前期无显著差异(F=1.560,df=1, 54,P=0.217)。在所有供试肿腿蜂中,雌蜂的产卵前期为6~8 d,其中绝大多数雌蜂产卵前期为6 d,少部分为7 d或8 d。当寄主为麻竖毛天牛幼虫时,松褐天牛肿腿蜂的产卵前期平均为6.63 d;寄主为麻竖毛天牛蛹时,产卵前期平均为6.43 d(图1)。

Figure 1. The pre-oviposition period of Sclerodermus alternatusi female on different stage of Thyestilla gebleri
-
从表2可知,在不同虫态麻竖毛天牛上发育的松褐天牛肿腿蜂子代的卵期、幼虫期、蛹期及幼期历期均无显著差异(卵期:F=1.542,df=1, 54,P=0.220;幼虫期:F=0.989,df=1, 41,P=0.325;蛹期:F=0.472,df=1, 41,P=0.496;幼期历期:F=2.562,df=1, 41,P=0.117)。当用麻竖毛天牛幼虫和蛹分别繁育松褐天牛肿腿蜂时,子代蜂的卵期分别平均为4.34 d和4.57 d,幼虫期平均为8.34 d和8.52 d,蛹期平均为19.89 d和20.07 d,幼期历期平均为32.61 d和33.40 d。
寄主虫态
Host stage卵期
Duration of egg stage/d幼虫期
Duration of larval stage/d蛹期
Duration of pupal stage/d幼期历期
Duration of immature stage/d幼虫 Larva 4.34 ± 0.12 8.34 ± 0.12 19.89 ± 0.16 32.61 ± 0.30 蛹 Pupa 4.57 ± 0.12 8.52 ± 0.12 20.07 ± 0.18 33.40 ± 0.38 F 1.542 0.989 0.472 2.562 P 0.220 0.325 0.496 0.117 Table 2. Developmental durations of Sclerodermus alternatusi progeny on different stage of Thyestilla gebleri
-
从表3可以看出,当用麻竖毛天牛幼虫和蛹分别繁育松褐天牛肿腿蜂时,单头寄主可育出的子代雌蜂数量差异显著(F=7.511,df=1, 41,P=0.009),其中单头麻竖毛天牛幼虫育出的子代雌蜂数平均为39.50头,显著高于单头麻竖毛天牛蛹育出雌性子代的30.80头。然而,麻竖毛天牛幼虫和蛹育出的雄性子代数量无显著差异(F=2.254,df=1, 41,P=0.141),单头麻竖毛天牛幼虫育出的平均雄性子代数为1.96头,单头麻竖毛天牛蛹育出的为1.40头。在麻竖毛天牛幼虫和蛹上育出的子代总数分别为41.18头和32.20头,前者显著高于后者(F=7.850,df=1, 41,P=0.008)。由图2可知,用麻竖毛天牛幼虫和蛹繁蜂时,子代蜂雄性比亦无显著差异(F=0.346,df=1, 41,P=0.560)。寄主为麻竖毛天牛幼虫时,平均子代雄性比为5.00%;寄主为天牛蛹时,平均子代雄性比为4.45%。
寄主虫态
Host stage子代雌蜂数
Female number/n子代雄蜂数
Male number/n子代总数
Progeny number/n幼虫 Larva 39.50 ± 1.99 1.96 ± 0.25 41.18 ± 1.98 蛹 Pupa 30.80 ± 2.24 1.40 ± 0.19 32.20 ± 2.33 F 7.511 2.254 7.850 P 0.009 0.141 0.008 Table 3. Progeny number of Sclerodermus alternatusi on different stage of Thyestilla gebleri
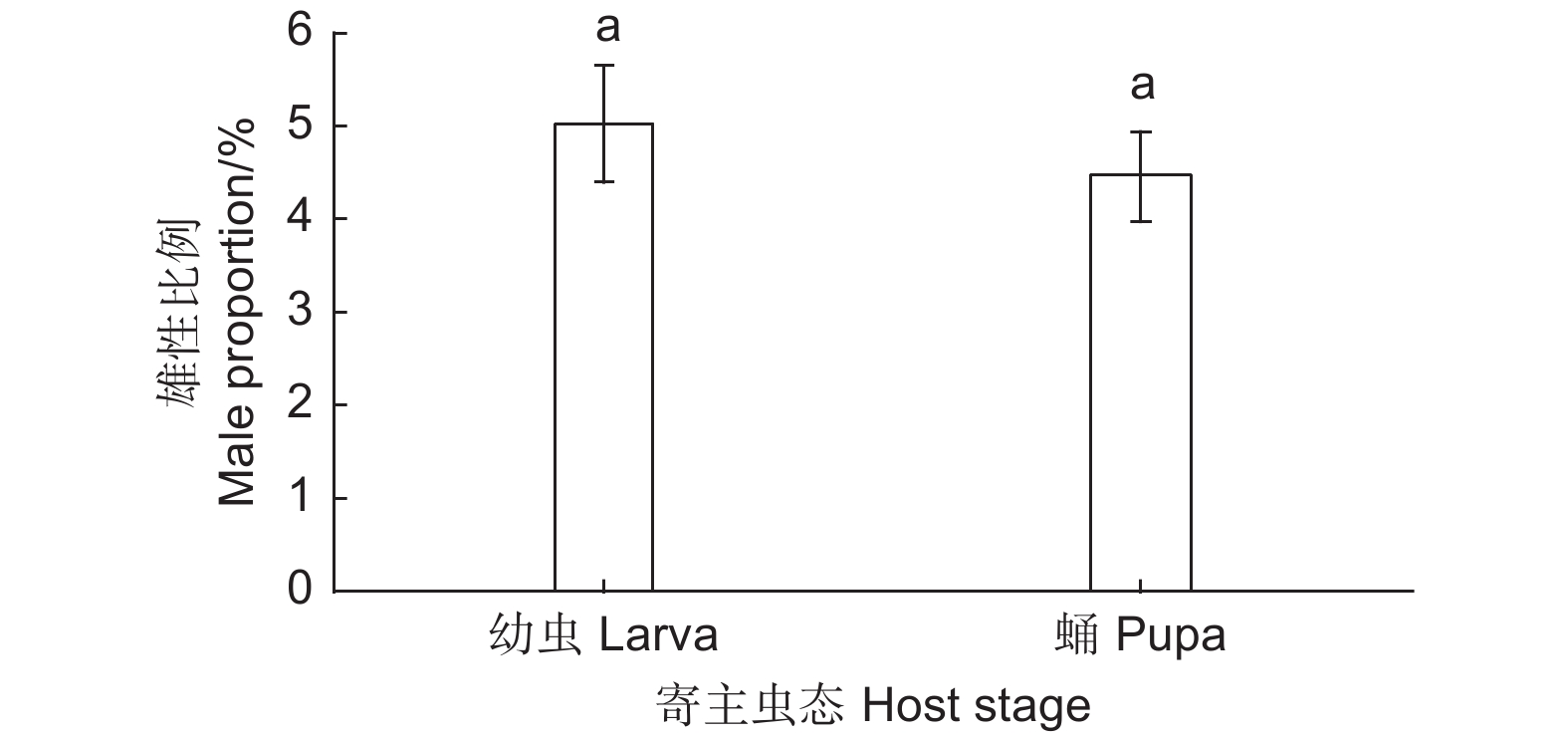
Figure 2. Male proportion of Sclerodermus alternatusi progeny on different stage of Thyestilla gebleri
-
作为多寄主型寄生蜂的代表,肿腿蜂的自然寄主包括天牛类、吉丁类、小蠹类、象甲类等[8]。在非选择条件下,其还可以寄生鞘翅目黄粉甲的蛹[11, 17]、膜翅目蜜蜂的蛹[12, 15]、鳞翅目玉米螟幼虫[14]、红铃虫幼虫[15]和大袋蛾幼虫[16]等。肿腿蜂在自然林分的环境极为复杂,需首先定位受害的树木,再搜索树木中的寄主,进而将寄主刺蜇麻痹后,才能完成产卵寄生过程[19]。上述过程对于无翅的肿腿蜂雌蜂而言(大多数肿腿蜂雌蜂无翅,仅依靠爬行寻找寄主)需耗费大量的能量,许多雌蜂在未找到寄主前就已耗尽能量[20]。当搜索到害虫后,并没有过多的机会去再次进行寄主选择。因此,多寄主型的生活史策略成为其获得最大适合度的重要途径。
肿腿蜂的多寄主性不仅体现在寄主种类的多样性,也体现在可寄生的寄主虫态的多样性。当以天牛类和吉丁类作为寄主时,肿腿蜂既能寄生幼虫态寄主,也可以寄生蛹态寄主[8]。前期对光肩星天牛生物防治的研究中观察到白蜡吉丁肿腿蜂可寄生光肩星天牛蛹,并完成生活史[5]。本研究结果发现,麻竖毛天牛幼虫和蛹均可被松褐天牛肿腿蜂寄生,且子代蜂可在两种不同虫态的寄主上完成生活史。松褐天牛肿腿蜂分别接种至麻竖毛天牛幼虫和蛹时,肿腿蜂对两种虫态寄主的寄生率无显著差异。以上结果首先表明肿腿蜂具备麻痹制服这两种虫态天牛的能力。其次,从母蜂产卵决策角度表明其认为麻竖毛天牛蛹和幼虫一样,均具备提供子代蜂完整发育的营养需求,即两种虫态的寄主对母蜂的产卵决策无影响。然而,母蜂在麻竖毛天牛幼虫或蛹体表产卵后,可成功育出子代蜂的天牛幼虫显著多于天牛蛹的数量。在试验过程中我们发现,当以麻竖毛天牛蛹为寄主时,肿腿蜂幼虫容易霉变和死亡,即较多子代蜂不能完成幼虫期,造成最终能羽化出蜂的寄主数较被寄生数大大降低。我们分析,肿腿蜂幼虫主要依靠寄主血淋巴和脂肪体所提供的营养而发育,而麻竖毛天牛化蛹初期其体内呈液态的组织增多,子代蜂幼虫因这一原因难以持续发育至成熟。同时,在麻竖毛天牛幼虫上发育最终所获得的雌性子代蜂数量也显著高于以麻竖毛天牛蛹作寄主的处理,同样也表明了天牛幼虫提供了更大的寄主适合度。虽然在麻竖毛天牛幼虫和蛹上育得的肿腿蜂子代雌蜂差异显著,但在两种虫态寄主上育得的子代雄蜂数无显著差异。通常,母蜂产卵时会对寄主质量进行评价,当寄主质量较差时,母蜂会选择产出更多的雄性后代,因为雄性后代在发育过程中受寄主质量变差的负面影响较雌性后代更弱[21-22]。本研究中几乎一致的子代雄蜂数进一步佐证了母蜂产卵时其所评价两种虫态寄主的质量无差异,麻竖毛天牛蛹所带来的肿腿蜂适合度降低主要因其不适于幼蜂发育。对抑性寄生蜂而言,子代发育历期与寄主大小紧密相关,通常在体型大的寄主上会因可获得的净营养更多使发育历期延长,以此获得体型更大的子代成蜂[22-23]。松褐天牛肿腿蜂是典型的抑性寄生蜂,本研究中2组寄主的体质量一致,故子代蜂各时期发育历期未表现出显著差异。基于以上结果,作者认为,对松褐天牛肿腿蜂繁育而言,麻竖毛天牛幼虫更适合作为繁蜂寄主。然而,若麻竖毛天牛幼虫在长期的低温储存中不可避免地存在一定比例的化蛹,仍建议将其用于繁蜂,这样可获得近60%的繁蜂成功率,可最大限度地避免寄主浪费。
此外,在应用肿腿蜂防治天牛类蛀干害虫时,以往主要在天牛幼虫期使用,较少有在天牛蛹期释放应用的案例。本研究结果表明,利用肿腿蜂防治天牛类害虫,也可以尝试在蛹期释放。肿腿蜂雌蜂搜寻到寄主后,雌蜂需取食寄主血淋巴补充营养后产卵[24],这一过程会直接致死寄主。即便是寄生天牛蛹后子代蜂的发育效率降低,但母蜂完成寄生实际也达成了害虫防治的目的。不过,许多天牛幼虫化蛹前,会将蛹室和蛀道内的虫粪排出,这可能会降低肿腿蜂对寄主的搜索效率,因为雌蜂通常需要依靠寄主虫粪所携带的化学信息来定位寄主[25]。同时,天牛在化蛹前会用细的木丝堵塞蛀道与蛹室,在林间究竟有多少比例的肿腿蜂能进入蛹室致死天牛蛹,需更多观察和研究。
-
(1)松褐天牛肿腿蜂可寄生麻竖毛天牛幼虫和蛹并完成生活史,对两种虫态的天牛的寄生率无显著差异,但以天牛幼虫作寄主时可育出子代蜂的寄主比例显著高于以天牛蛹作寄主的处理。
(2)以麻竖毛天牛老熟幼虫和蛹繁蜂时,母蜂产卵前期、子代蜂卵期、幼虫期和蛹期均无显著差异。以天牛幼虫作寄主时,可获得的子代蜂总数和雌蜂数更多。寄生不同虫态的麻竖毛天牛时,子代蜂性比均为显著的偏雌特征,子代雄蜂数和雄性比例无显著差异。
(3)利用麻竖毛天牛蛹繁育松褐天牛肿腿蜂,可获得近60%的繁蜂成功率,但其繁蜂效率低于麻竖毛天牛老熟幼虫。
Parasitism Rate and Progeny Development of Sclerodermus alternatusi (Hymenoptera: Bethylidae) on Different Stage of Thyestilla gebleri (Coleoptera: Cerambycidae)
- Received Date: 2022-03-06
- Accepted Date: 2022-04-22
- Available Online: 2022-12-20
Abstract:
 DownLoad:
DownLoad: