



-
林木性状变异受一定程度的遗传控制,有效的遗传变异决定了遗传改良的潜力,这种遗传变异广泛存在于家系及个体间[1]。优良家系选择是林木遗传改良的重要环节[2-3],揭示家系性状的遗传变异规律是科学制定育种策略的依据,也是评价亲本材料遗传价值和估算遗传参数的有效途径,通过家系间的遗传变异分析,可以根据子代生长表现回选优良亲本,为林木遗传改良创造一个基本群体[4-5],也可为下一代育种提供优良家系,提高人工林生产力[6-7]。不同家系林木在不同林龄阶段生长性状具有差异性,开展优良家系早期筛选,是缩短育种周期和加速育种选择进程的有效方法之一[8-9]。国内外已开展桉树(Eucalyptus robust Smith)[10]、木荷(Schima superba Garden. et Champ.)[6, 11]和日本落叶松(Larix kaempferi (Lamb.) Carr.)[12]等多种速生树种早期优良家系选择的研究,证实不同家系间存在显著遗传变异。研究林木家系间随林龄的变化趋势,进行优良家系早期选择,对于林木遗传改良具有重要意义。
麻栎(Quercus acutissima Carruth.)属壳斗科(Fagaceae)落叶乔木,广泛分布于东亚地区,是我国温带和亚热带森林生态系统中的重要组成树种[13]。麻栎耐干旱瘠薄,具有较强的适应性,在山区、丘陵均可栽植,是荒山瘠地造林的先锋树种和营造水土保持林的优良树种[14]。麻栎材质坚硬、不易变形、耐腐蚀,用途极其广泛,也是我国重要的硬阔叶能源树种和优良用材树种[15-16]。近年来,安徽、河南等地栎炭产业发展较快,麻栎作为优质木炭的原材料日益受到关注,而当前炭用林培育缺乏优良种质材料,是制约其发展的重要因素之一。前期已对麻栎不同种源幼林生长和生物量随林龄的变化进行了研究[17],而目前针对不同家系生长随林龄的变化尚未有深入研究。因此,本试验依据江西永丰、浙江开化和安徽滁州3个试验点麻栎优树自由授粉家系试验林生长表现,展开不同家系生长性状及其稳定性随林龄变化特征研究,以期筛选出该区域麻栎优良家系,为麻栎短轮伐期炭用林高效培育提供理论依据。
-
试验地位于江西省永丰县官山林场(27°19′ N, 115°25′ E)、浙江省开化县林场(29°09′ N, 118°25′ E)和安徽省滁州市红琊山国有林场(32°10′ N, 118°04′ E)。江西永丰年均气温18 ℃,年均降水量1718 mm,试验地平均海拔169 m,土壤为红壤。浙江开化年均气温16.4 ℃,年均降水量1831 mm,试验地平均海拔146 m,土壤类型以黄红壤为主。安徽滁州年均气温15.4 ℃,年降水量1 075 mm,试验地平均海拔150 m,土壤为黄棕壤。
-
试验林18个优树自由授粉家系来源于广西融水(25°04′ N,109°15′ E),该区属亚热带季风气候,年平均气温19.3℃,年降水量1 824 mm,海拔1 000~1 500 m,土壤类型以红壤为主。2006年在该区天然林中选择林龄30 a以上的优树作为采种母树,所选优树树干通直圆满,间隔50 m以上,无病虫害,高、径生长明显大于周围的3~5株对比木。2007年春集中播种育苗。2008年3月选取1年生健壮裸根实生苗(苗高45.17~79.76 cm,地径0.59~1.09 cm)用于各试验点造林,栽植株行距为2 m × 3 m。采用完全随机区组设计,6株单列小区,6次重复。
-
分别于2008、2010、2013、2015和2018年测定麻栎树高和胸径(地径),在试验区邻近的麻栎人工林内选择1年生、3年生、5年生、8年生、10年生和13年生林分,共建立15 m × 15 m临时样地12块,在每个样地内选择5株平均木,每个林龄共10株,伐倒标准木测定地上单株生物量,并通过生物量与树高、胸(地)径的实测值构建生物量模型:
1年生、3年生:W = 0.00710.9382 × D(R2 = 0.9908)
6年生、8年生和11年生: W = 0.116 105(D2H)0.832 016(R2 = 0.978 5)
式中:W为地上单株生物量(kg),D为胸径或地径(1~3年生)(cm),H为树高(m)[17]。
采用DPS 9.01数据处理系统,开展麻栎年份、地点和家系间的3因素方差分析,方差分析模型为:
$ \begin{split} &\qquad X_{i j k l}=\mu + \tau_{i} + v_{j} + (\tau v)_{i j} + w_{k} + (\tau v)_{i k} + (v w)_{j k} + \\ &(\tau v w)_{i j k} + \rho_{j k l} + \varepsilon_{i j k l} \end{split} $
式中:Xijkl为第i个家系在第j地点、第k年份、第l区组的观测值,μ为群体的平均值,τi为家系i的效应值,vj为地点j的效应,(τv)ij为家系 × 地点互作效应,(τv)ik为家系 × 年份的互作效应,(vw)jk为地点 × 年份的互作效应,(τvw)ijk为家系 × 地点 × 年份的互作效应,ρjkl为家系 × 地点 × 年份的互作效应,εijkl为随机误差。
运用AMMI模型对11年生麻栎的地上单株生物量进行稳定性分析,模型如下:
$ Y_{i j k}=u + \alpha_{i} + \beta_{j} + \sum\limits_{r=1}^{n} \quad \lambda_{r} \psi_{i r} \sigma_{j r} + \rho_{i j} + \varepsilon_{i j k} $
式中:Yijk是第i个家系在第j个地点的第k次重复的观测值;u为总体平均值;αi为第i个家系均值与总体均值的离差(即家系主效应);βj为第j个环境与总平均的离差(即环境主效应);n为保留在模型中的主分量轴的个数,即表示能包含大部分基因交互作用信息所需要的乘积项数目;λr为第r个交互效应主成分轴(IPCA)的特征值或奇异值,代表交互作用平方和中可由该轴解释的部分;ψir为第r轴的家系特征矢量值;σjr为第r轴环境特征矢量值;ρij为提取n个主分量轴后留下的残差(交互剩余项);εijk为试验误差。
家系与地点间的相对稳定性参数:
$ D_{g(e)}=\sqrt{\sum\limits_{i=1}^{n}(\text { IPCA })^{2} g(e)} $
式中:n为显著的主分量个数;Dg(e)为第g个基因型或第e个环境在n个IPCA上的得分,Dg(e)大小度量了第g个基因型或第e个环境的相对稳定性[18]。
在SAS 8.0软件中,调用PROC VARCOMP过程中的REML方法计算各方差分量,进而估算下列遗传参数。
$家系遗传力:h_{F}^{2}=\dfrac{\sigma_{F}^{2}}{\dfrac{\sigma_{E}^{2}}{n b}+\dfrac{\sigma_{F B}^{2}}{b}+\sigma_{F}^{2}} $
$单株遗传力:h_{S}^{2}=\frac{4 \sigma_{F}^{2}}{\sigma_{E}^{2}+\sigma_{F B}{ }^{2}+\sigma_{F}{ }^{2}} $
式中:n为小区单株数的调和值,b为区组数量,
$ {\sigma _{{F}}^2} $ 为家系方差分量,$ {\sigma _{{E}}^2} $ 为机误,$ {\sigma _{{{FB}}}^2} $ 为家系与区组互作的方差分量。$ 遗传变异系数:C V_{G}=\frac{\sqrt{\sigma_{g}^{2}}}{\overline{X}} \times 100 {\text{%}}$
$ 表型变异系数:C V_{P}=\frac{\sigma_{P}}{\overline{X}} \times 100 {\text{%}} $
式中:
$ {\sigma _g^2} $ 为遗传方差分量,$ {\sigma _{{P}}} $ 为表型标准差,$ \overline {{X}} $ 为性状的均值。$表型相关:r_{P}=\frac{{Cov}_{P(j, m)}}{\sqrt{\sigma_{P j}^{2} \sigma_{p m}^{2}}}$
$ 遗传相关:r_{g}=\frac{{Cov}_{g(j, m)}}{\sqrt{\sigma_{g j}^{2} \sigma_{g m}^{2}}}$
式中:
$ Co{v_{P(j,m)}} $ 和$ Co{v_{g(j,m)}} $ 分别为早期性状与晚期性状间的表型协方差和遗传协方差,$ {\sigma _{pj}^2} $ 和$ {\sigma _{gj}^2 }$ 分别为早期性状的表型方差分量和遗传方差分量,$ {\sigma _{pm}^2 }$ 和$ {\sigma _{gm}^2 }$ 分别为晚期性状的表型方差分量和遗传方差分量[6]。$ \begin{split} & 入选家系的遗传增益:\qquad \qquad \qquad \qquad \\ & \Delta G=\left(X_{i}-\overline{X}_{j}\right) / \overline{X} \;\times \;h_{F}^{2} \;\times \;100 {\text{%}} \end{split} $
式中:Xi为入选家系性状的平均值,
$ \overline {{X}} $ j为各试验地性状的群体平均值,$ \overline {{X}} $ 为性状的均值,$h_F^2 $ 为家系遗传力。 -
试验表明,江西永丰、浙江开化和安徽滁州3个试验点不同林龄的麻栎胸(地)径、树高和地上单株生物量存在极显著家系遗传差异,这为优良家系选择提供了丰富的变异基础(表1)。江西永丰、浙江开化和安徽滁州3个试验点3年生麻栎地上单株生物量比1年生分别增加895.09%、382.85%和671.96%,6年生地上单株生物量比3年生分别增加1013.56%、7000.66%和3171.11%,11年生地上单株生物量比6年生分别增加100.19%、74.92%和91.63%;地上单株生物量在3~6 a间的生长量最大,6~11 a间生长量相对较小,说明3~6 a间麻栎生长速度较快。1年生、3年生、6年生、8年生和11年生时,江西永丰试验点地上单株生物量均值最大家系比最小家系分别高出55.00%、248.54%、212.79%、760.05%和127.00%,浙江开化试验点其分别高出7.13%、14.87%、643.25%、1147.38%和174.66%,安徽滁州试验点其分别高出8.42%、38.06%、743.23%、1338.60%和2810.72%。
表 1 3个试验地点不同林龄家系胸(地)径、树高和地上单株生物量的统计和方差分析
Table 1. Statistics and variance analysis of DBH(D), H and AGB for family with stand ages in 3 sites
试验点
Site性状
Trait林龄
Stand age/a均值
Mean变幅
Variable rang变异系数
CV/%方差分量百分比
Variance component percentage /%家系
Family区组
Block区组 × 家系
Block × Family机误
Error江西永丰
Yongfeng,Jiangxi胸(地)径
DBH(D)/cm1 1.58 1.33~1.79 6.99 21.48** 2.03** 0.86** 0.74 3 4.09 3.61~4.94 8.93 38.98** 3.95** 0.71** 0.19 6 3.69 2.72~4.97 14.56 2.47** 0.48** 0.19** 0.04 8 5.48 4.10~6.51 10.50 24.82** 1.46* 1.26** 0.33 11 7.50 6.02~8.71 8.75 8.26 13.65** 5.24** 2.97** 树高
H/m1 1.16 1.00~1.30 6.07 11.48** 1.61** 0.28** 0.07 3 2.52 1.80~2.72 8.50 50.71** 4.17** 1.27** 0.47 6 4.26 3.78~4.94 7.39 2.43** 0.71** 0.29** 0.04 8 5.56 4.73~6.24 7.28 4.68** 2.63** 0.69** 0.19 11 6.86 5.93~7.61 5.97 6.39** 0.98** 0.25** 0.01 地上单株生物量
AGB/kg1 0.03 0.02~0.04 13.62 0.37** 0.12** 0.08** 0.01 3 0.35 0.21~0.73 38.14 1.36** 0.96** 0.13** 0.02 6 3.48 1.91~5.96 29.90 1.36** 0.96** 0.23* 0.12 8 8.32 4.42~12.02 22.56 3.32** 0.29** 0.12** 0.09 11 16.66 10.13~23.03 19.03 6.39** 0.98** 0.25** 0.01 浙江开化
Kaihua,
Zhejiang胸(地)径
DBH(D)/cm1 1.07 0.83~1.24 7.93 13.25** 2.05** 1.61** 0.03 3 2.68 2.01~3.54 14.25 15.57** 3.72** 0.47** 0.36 6 5.32 4.19~6.56 12.96 35.55** 2.35** 1.07* 0.16 8 7.08 5.66~8.24 8.24 14.61** 1.87** 2.33** 0.50 11 8.96 7.28~11.38 12.07 65.11** 4.08** 3.69** 0.43 树高
H/m1 0.95 0.74~1.14 9.64 26.22** 1.09** 0.31** 0.01 3 2.03 1.83~2.35 8.10 30.20** 0.72** 0.33** 0.13 6 4.50 4.07~5.21 6.42 12.67** 0.85** 0.92** 0.03 8 6.51 5.56~7.32 6.93 58.30** 0.89** 1.15** 0.05 11 7.86 6.72~9.37 9.54 47.53** 2.28** 1.79** 0.25 地上单株生物量
AGB/kg1 0.01 0.01~0.02 7.55 6.39** 2.56** 1.45** 0.12 3 0.09 0.05~0.19 36.44 1.87** 1.22** 0.12** 0.04 6 6.66 4.06~10.49 25.82 0.63** 0.28** 0.09** 0.04 8 14.47 8.84~20.31 18.43 38.53** 3.98** 0.10** 0.08 11 25.31 15.43~42.37 27.71 6.22* 2.59** 1.20** 0.63 安徽滁州
Anhui, Chuzhou胸(地)径
DBH(D)/cm1 1.72 1.43~1.72 5.31 46.89** 2.08** 0.76** 0.04 3 4.43 2.47~4.43 12.59 20.33** 0.58** 0.53** 0.13 6 6.45 4.07~6.45 12.07 59.96** 1.83** 1.69** 0.19 8 9.52 6.98~9.52 8.16 30.04** 2.46** 2.36** 0.28 11 12.73 9.32~12.73 9.31 42.71** 4.99** 3.89** 0.34 树高
H/m1 1.18 1.05~1.29 5.36 30.15** 1.31** 0.25** 0.14 3 4.42 3.72~4.94 6.84 16.96 + 3.13** 0.13** 0.20 6 5.28 3.73~6.20 14.61 29.93** 0.62** 0.56** 0.25 8 6.81 4.94~7.63 11.94 11.30** 1.61** 0.93** 0.06 11 8.75 7.6~10.13 7.83 21.78** 2.73** 1.65** 0.09 地上单株生物量
AGB/kg1 0.03 0.02~0.03 7.80 1.87** 0.22** 0.11** 0.05 3 0.24 0.07~0.45 38.90 2.25** 0.32* 0.11** 0.01 6 7.95 4.36~11.79 27.41 1.68** 0.22** 0.12* 0.06 8 19.14 11.89~25.27 18.99 0.93** 0.39** 0.08** 0.01 11 36.67 26.84~54.95 22.10 0.69** 0.15* 0.09** 0.06 注:**表示0.01水平差异显著,*表示0.05水平差异显著。下同。
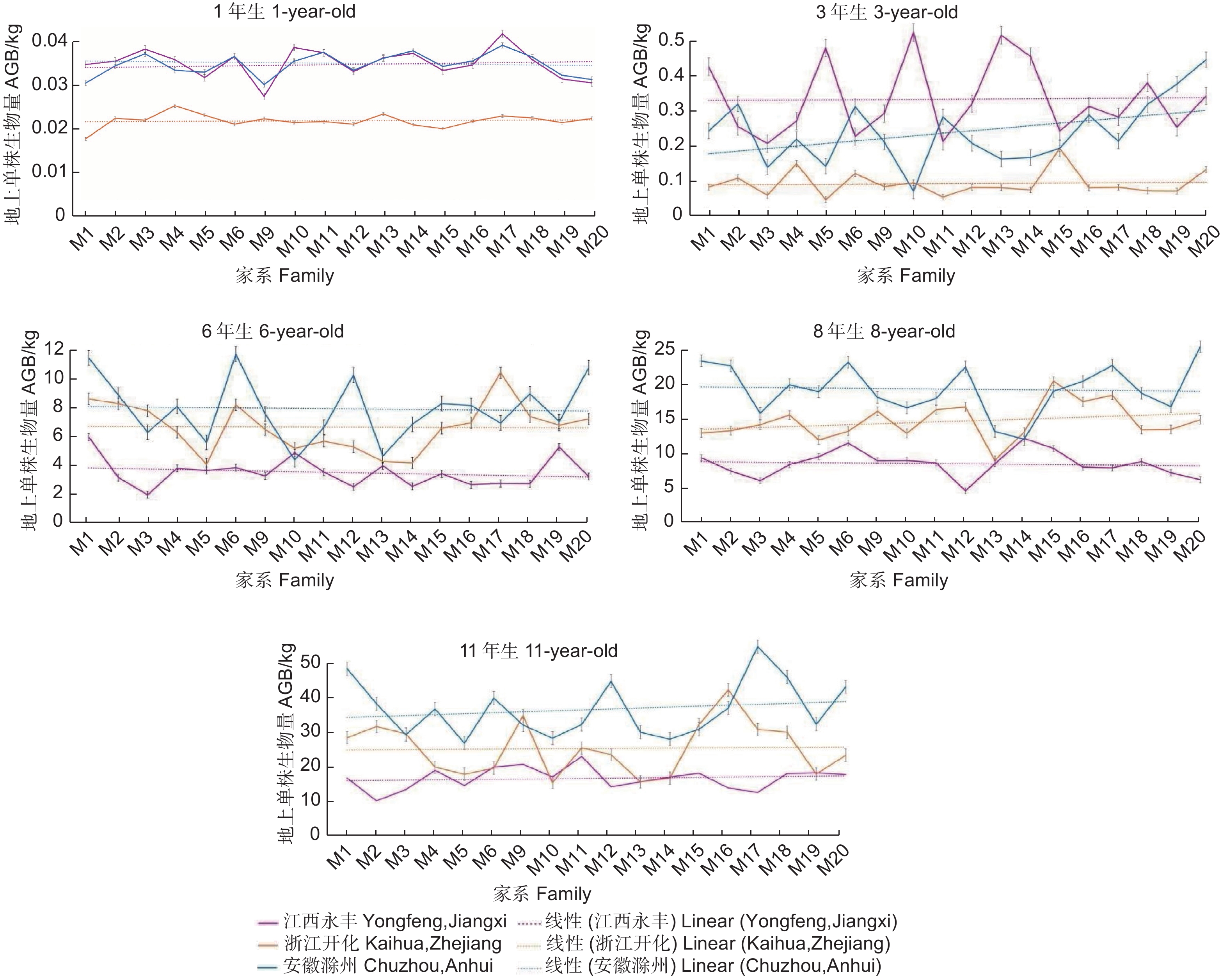
Note: **indicates significant difference at P<0.01, *indicates significant difference at P<0.05. The same below.对3个试验点麻栎各家系间生物量的生长动态分析发现,1~3年生时,江西永丰试验点生长较好,年均相对生长量比浙江开化和安徽滁州试验点分别高493.89%和85.08%。随着林龄增加,江西永丰试验点的生长优势逐渐减弱,6~11年生时在各试验点中表现最差;浙江开化试验点各家系在前期生长表现较差,而6年生后,在3个试验点表现中等;安徽滁州试验点各家系前3年生长表现较差,6~11年生时生长表现最好,江西永丰、浙江开化和安徽滁州试验点的年相对生长量分别为1.79、3.29和6.16 kg(图1)。
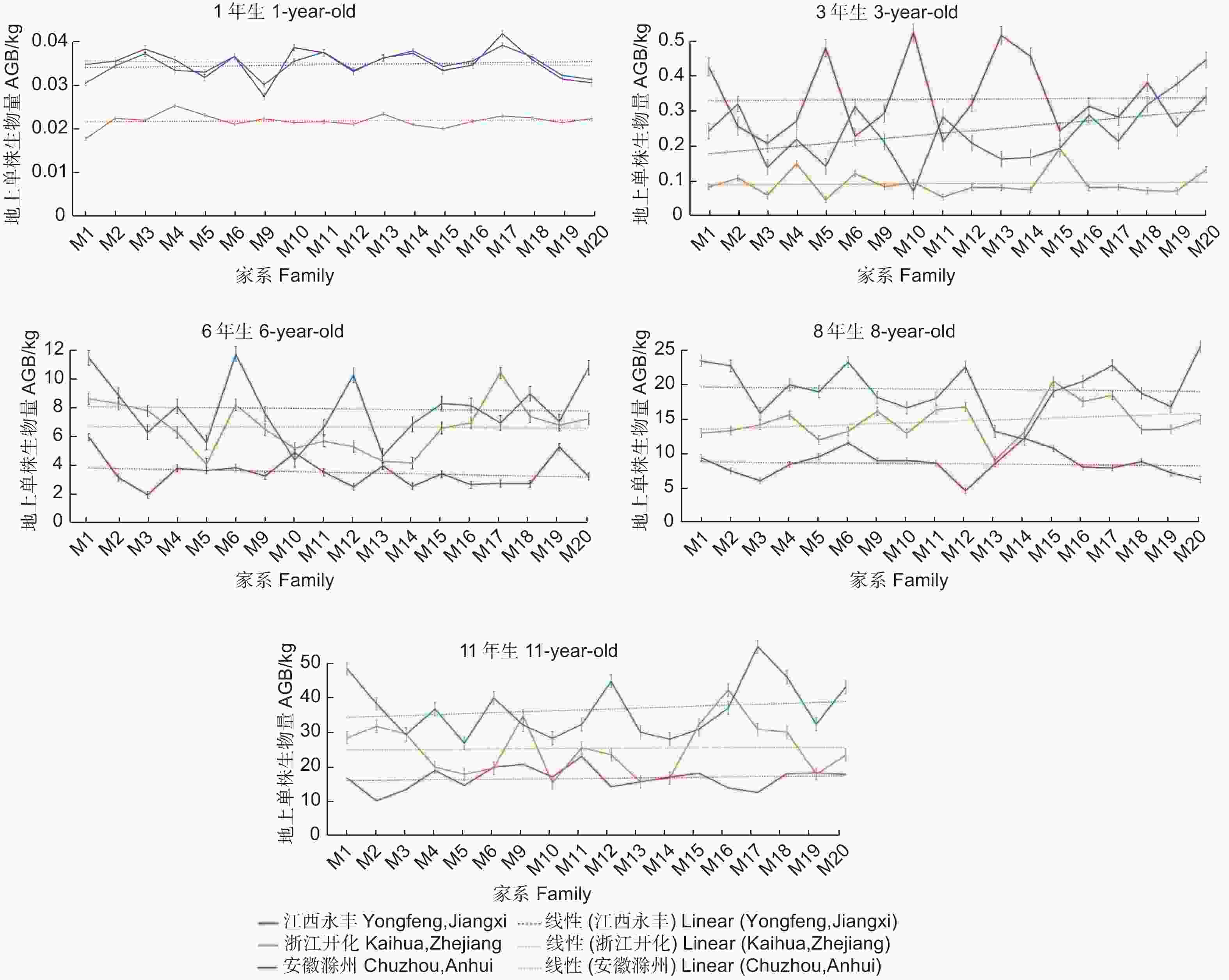
图 1 3个地点不同家系地上单株生物量随林龄的变化特征
Figure 1. Variation characteristics of AGB per plant in 3 sites among different families with the increase of stand age
为了更清晰地反映麻栎家系间生长动态变化,以地上单株生物量为例,分别从江西永丰、浙江开化和安徽滁州试验点早期(1~3年生)地上单株生物量年增长的变化范围内,选择高、中和低各两个家系,进行家系间生长趋势分析(图2)。早期地上单株生物量长势较好的部分家系,在后续生长过程中,优势生长地位相对降低,如江西永丰试验点M1和M10家系,浙江开化试验点M4家系及安徽滁州试验点M6和M18家系;早期生长缓慢的家系,在后续生长过程中仍一直保持较差水平,如江西永丰试验点M2和M17家系,浙江开化试验点M5家系及安徽滁州试验点M9和M10家系;而早期生长处于中等水平的家系,在后续生长中,有些生长为优势家系,而有些仍保持中等水平,如江西永丰试验点M9家系地上单株生物量1年生、3年生、6年生和8年生时分别比优势家系M1低42.66%、146.32%、50.31%和0.44%,而到11年生时高21.58%;浙江开化试验点M17家系1年生、3年生时分别比优势家系M15低13.48%和135.03%,而到6年生、8年生和11年生时分别比其高59.38%、11.36%和6.92%;安徽滁州试验点M17家系1年生、3年生、6年生和8年生时分别比优势家系M10低42.66%、146.32%、50.31%和0.44%,而到11年生时比其高21.58%。
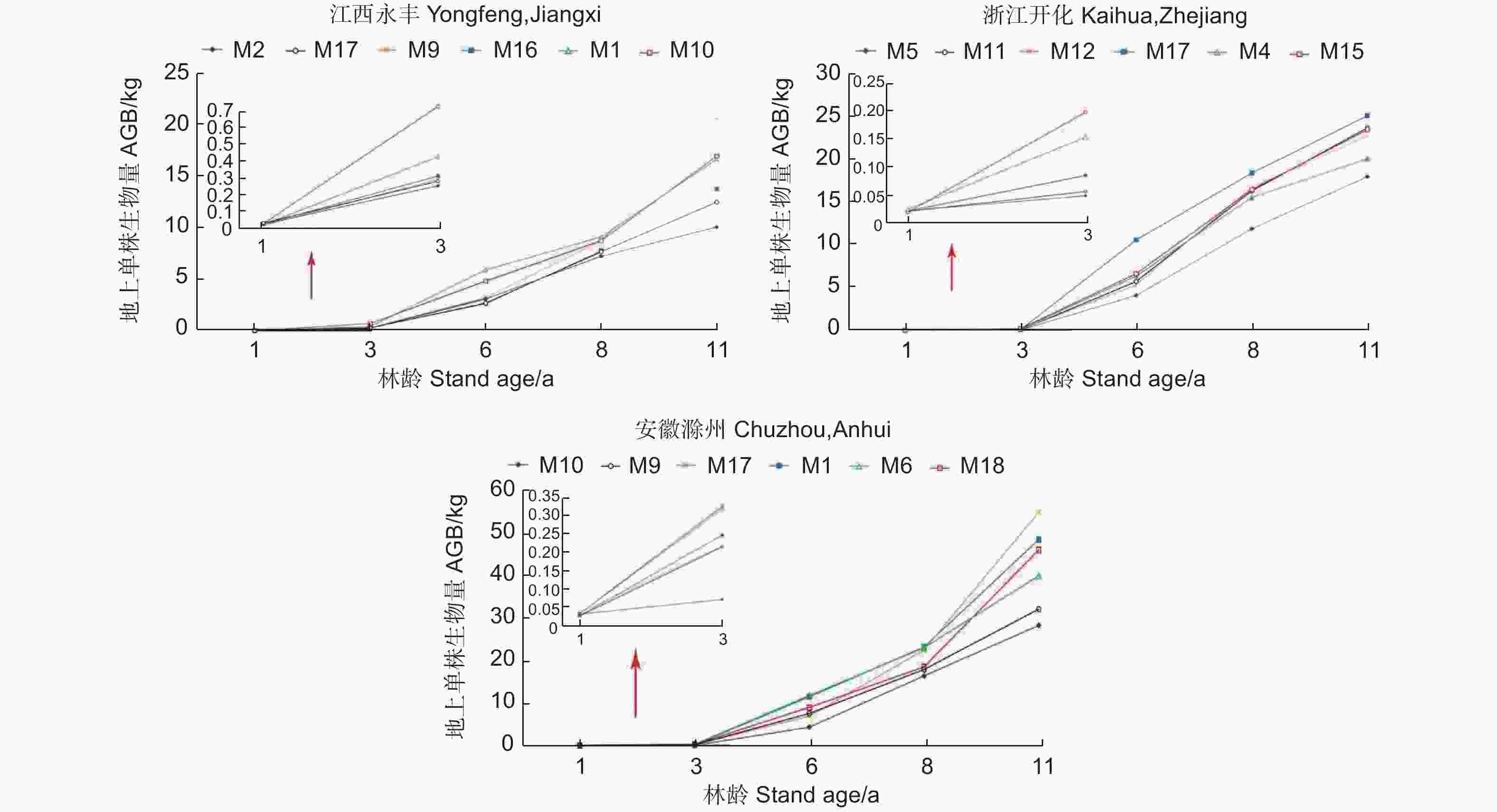
图 2 3个试验点典型家系地上单株生物量随林龄变化
Figure 2. Variation of AGB per plant of typical families in 3 test sites with the increase of stand age
-
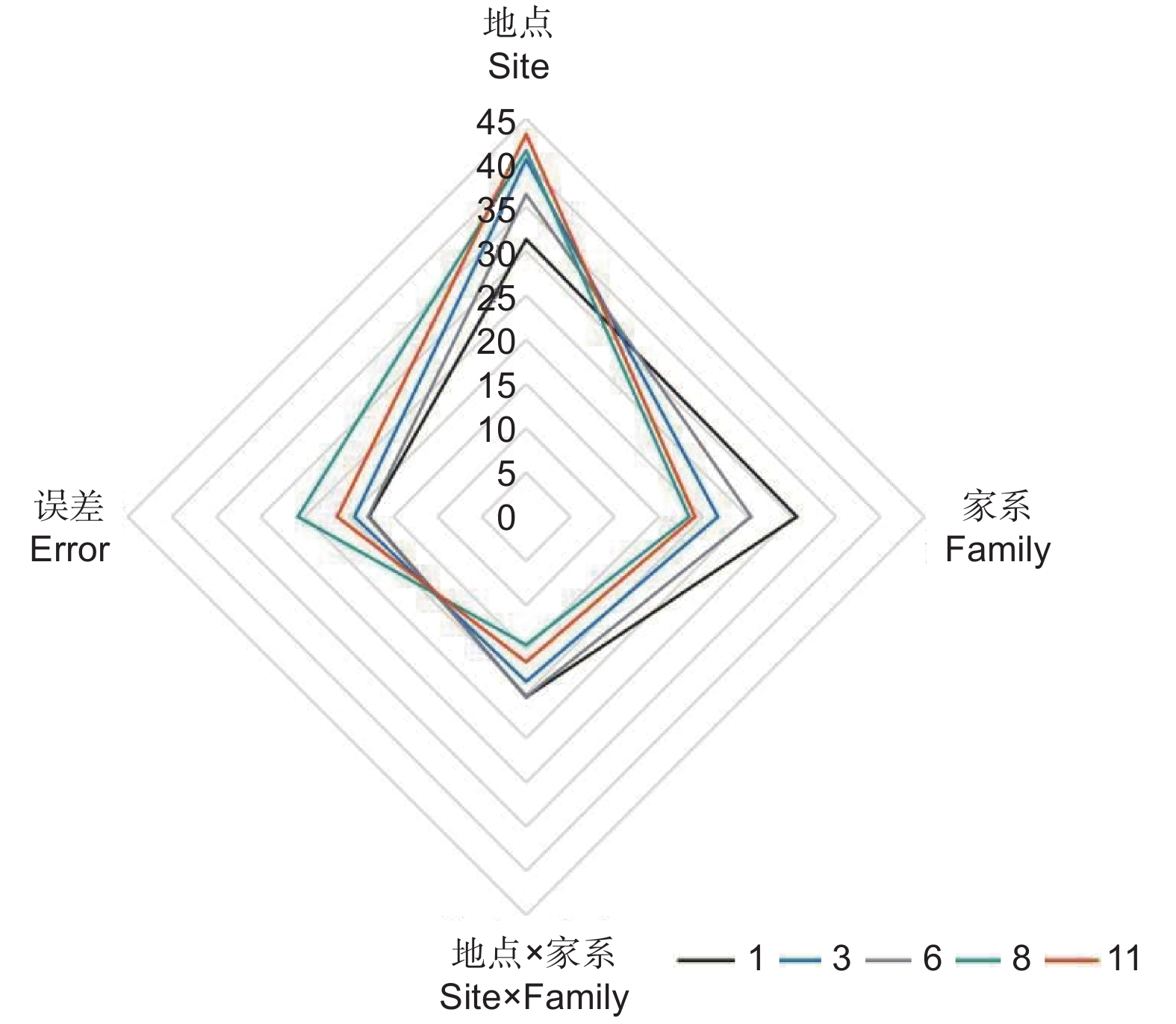
以麻栎地上单株生物量为例,分析年份、地点、家系及其交互作用的影响,结果表明:林龄、地点、家系、地点 × 林龄、家系 × 林龄、地点 × 家系以及地点 × 家系 × 林龄的交互作用均具有极显著差异(P<0.01)(表2)。方差分析发现:地点对不同林龄麻栎地上单株生物量影响最大(1年生、3年生、6年生、8年生和11年生时的方差分量分别为31.32%、 40.36%、36.42%、 41.35%和43.20%),且随着林龄增大影响程度呈增加趋势;其次为家系(1年生、3年生、6年生、8年生和11年生时的方差分量分别为30.57%、21.65%、25.4%、18.4%和19.06%);地点 × 家系交互作用对生长的影响相对较小(1年生、3年生、6年生、8年生和11年生时的方差分量分别为20.36%、18.61%、20.33%、14.47%和16.40%),随着林龄增加呈减少趋势(图3)。
表 2 参试家系地上单株生物量的3因素方差分析
Table 2. Three-way variance analysis of the AGB per plant in the test families
生长性状
Growth trait变异来源
Source of variation自由度
df均方
Mean SquareF 总的 Total 321 64.27 地上单株生物量
AGB林龄 Stand age 4 44323.95 769.02** 地点 Site 2 2722.79 94.12** 家系 Family 17 109.33 102.49** 地点 × 林龄 Site × Stand age 8 1737.27 30.14** 家系 × 林龄 Family × Stand age 68 43.74 212.75** 地点 × 家系 Site × Family 34 77.27 155.12** 地点 × 家系 × 林龄 Site × Family × Stand age 136 49.81 86.44** 误差 Error 72 31.06 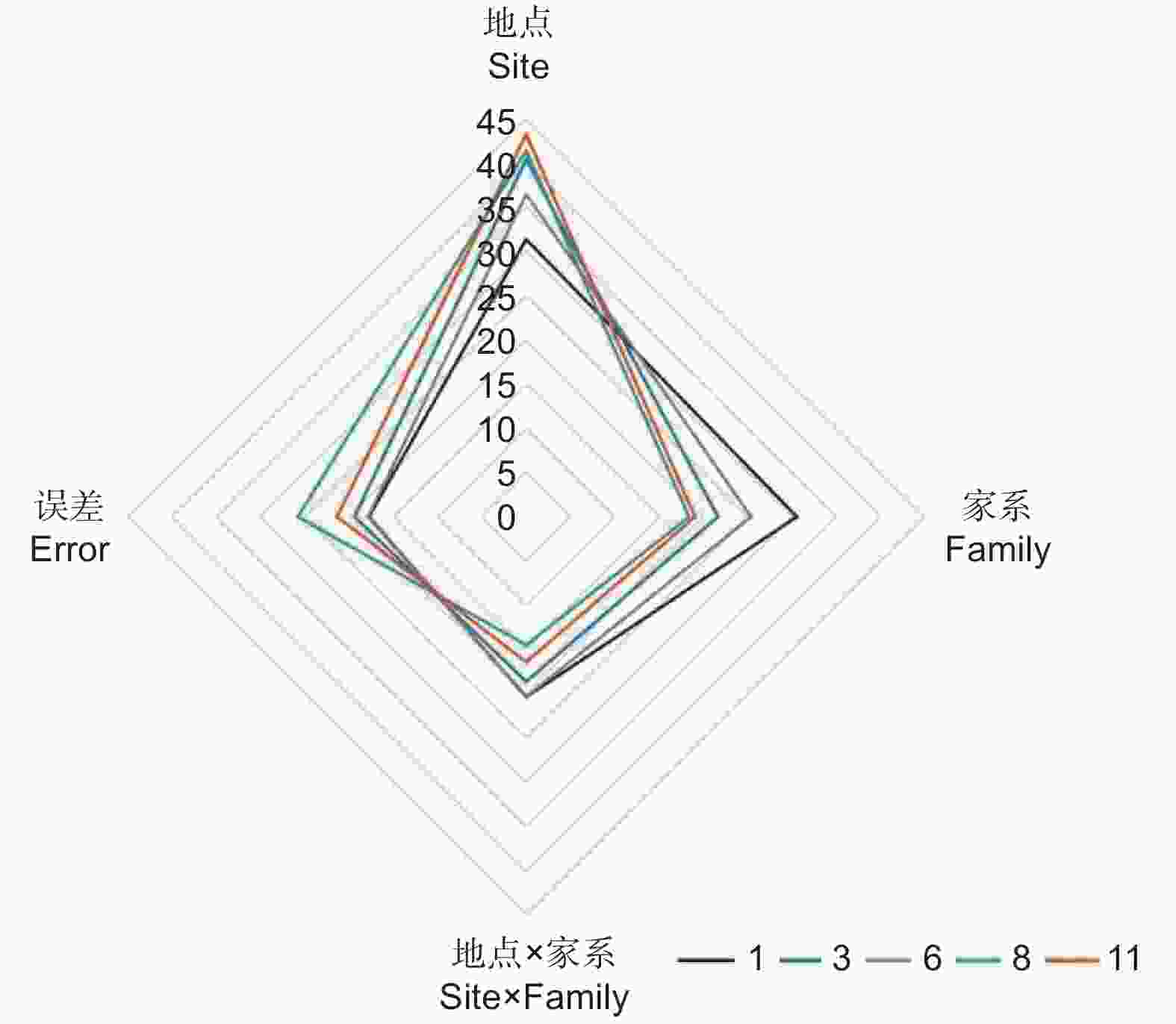
图 3 地点、家系及互作效应对地上单株生物量的相对影响随林龄的变化特征
Figure 3. Effect of site, family and their interaction on AGB per plant with the increase of stand age
-
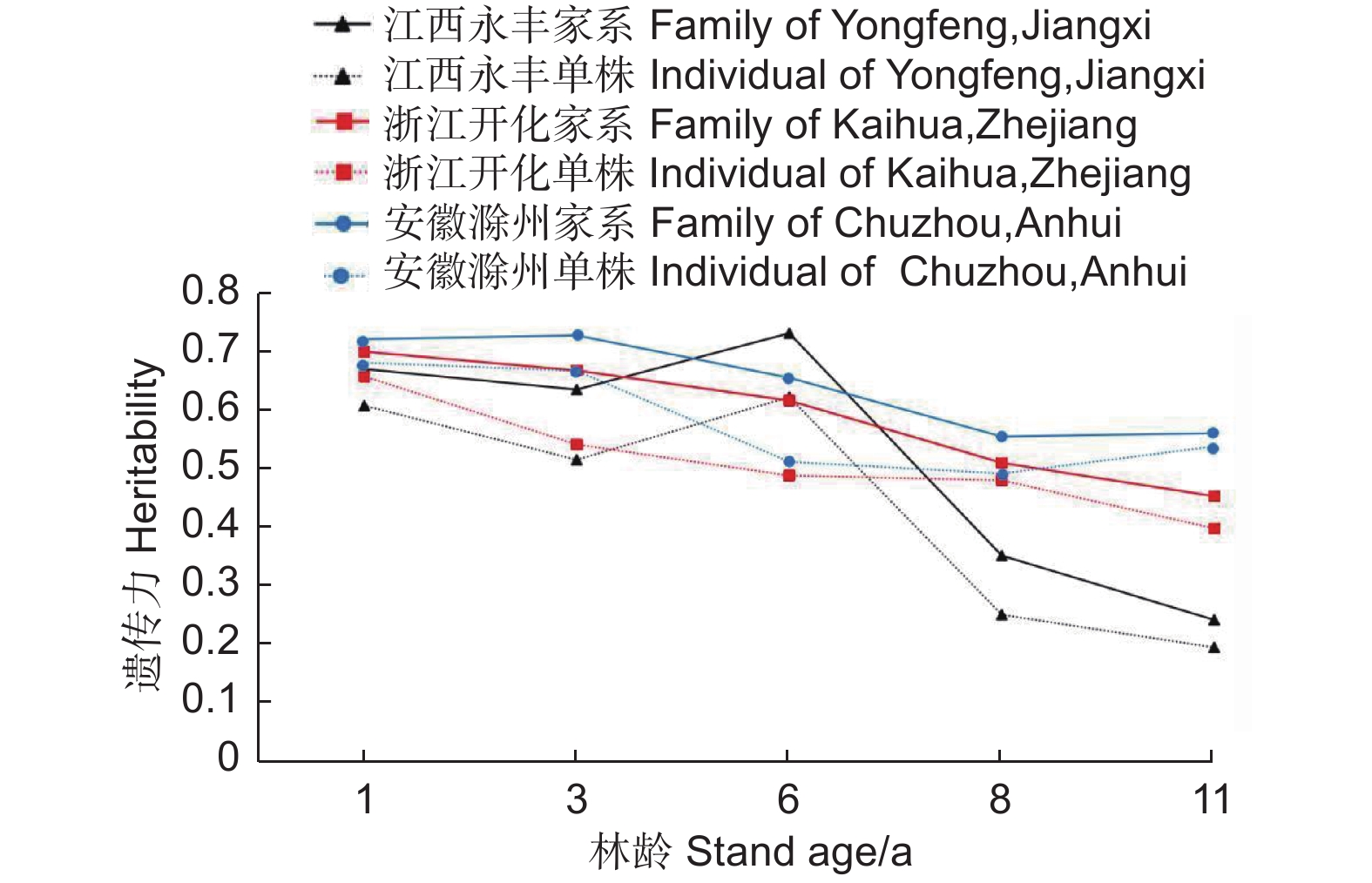
麻栎不同家系地上单株生物量家系遗传力随林龄增长呈减小趋势(图4)。江西永丰、浙江开化和安徽滁州试验点地上单株生物量家系遗传力均值分别为0.53、0.59和0.64,说明不同地点家系生长受较强的遗传控制。家系遗传力在1~6年生时(试验点遗传力变化区间:江西永丰0.63~0.73,浙江开化0.61~0.70,安徽滁州0.65~0.73)受遗传控制较强,相对较稳定,但在8~11年生时(试验点遗传力变化区间:江西永丰0.24~0.35,浙江开化0.45~0.51,安徽滁州0.55~0.56)所受遗传控制相对减小,说明麻栎生长前期受遗传影响较大。单株遗传力和家系遗传力具有相似的变化趋势,家系选择可配合单株进行,为优良家系筛选提供科学依据。
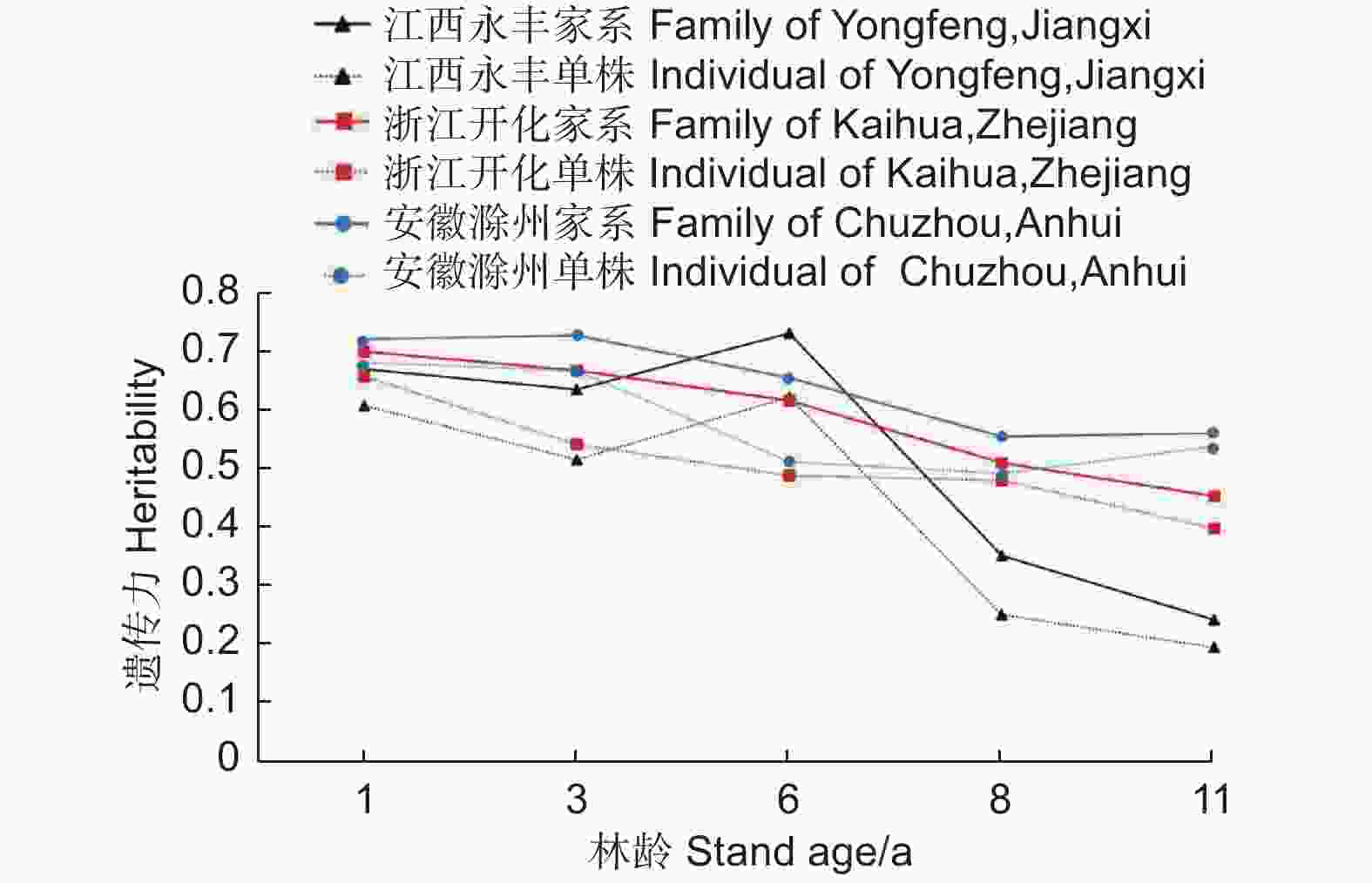
图 4 地上单株生物量的遗传力随林龄的变化趋势
Figure 4. Variation heritability of AGB per plant with the increase of stand age
不同林龄麻栎家系地上单株生物量的表型变异系数(均值23.06%)均大于遗传变异系数(均值9.08%)(图5)。总体上,地上单株生物量表型变异系数和遗传变异系数呈先增加后减小趋势,3年生时表型变异系数和遗传变异系数达最大,6~11年生时3个试验点间的遗传变异系数均趋于稳定,这表明随着林龄增长,家系间生长差异相对减小,受遗传控制因素影响也保持相对稳定。
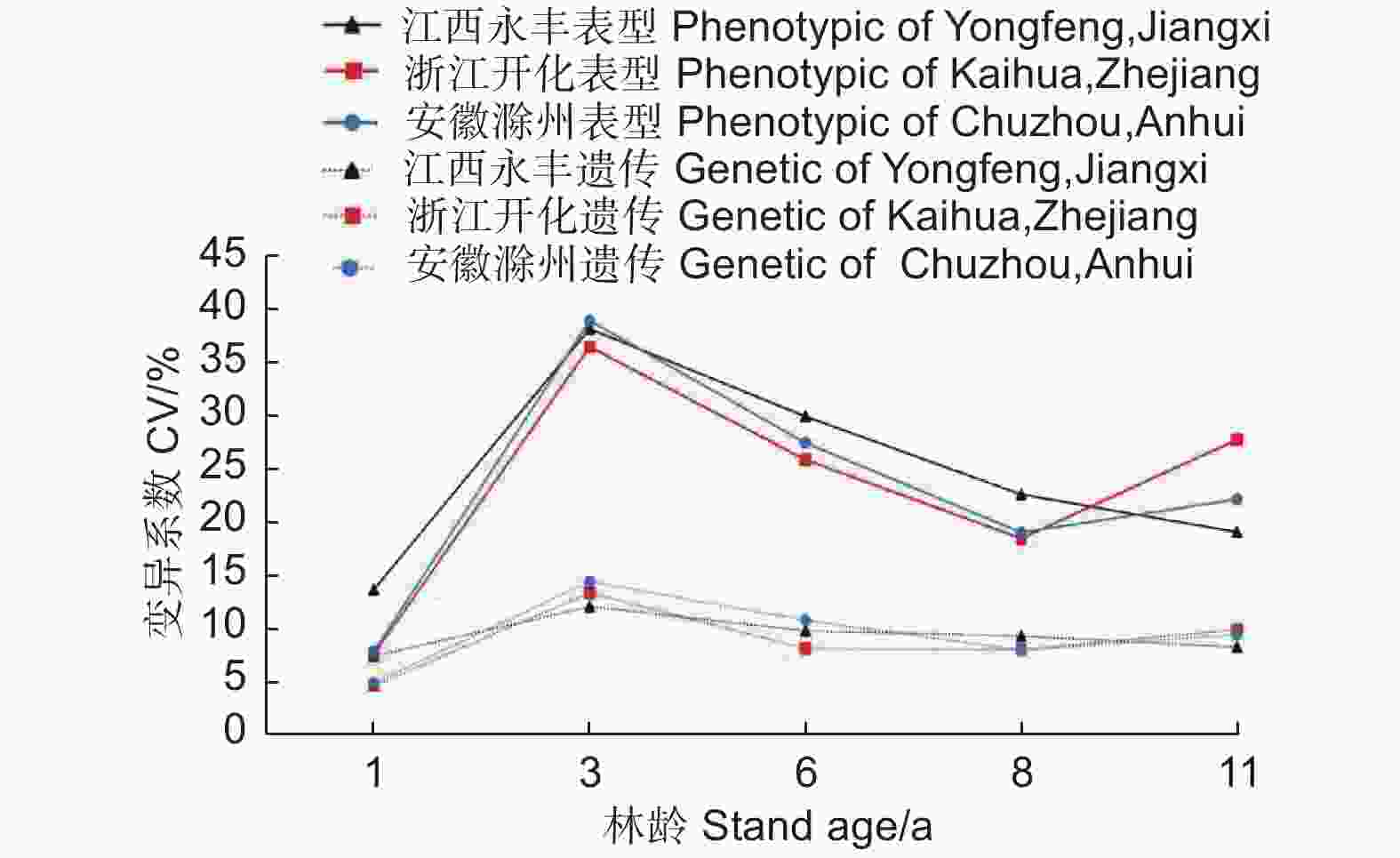
图 5 地上单株生物量变异系数随林龄的变化趋势
Figure 5. Variation coefficient of AGB per plant with the increase of stand age
-
以11年生麻栎地上单株生物量为参数进行家系稳定性分析。AMMI模型分析发现:仅用模型第1主成分分量就能解释59.46%的交互作用,且达到了显著水平(P<0.01),表明应用AMMI模型解释不同家系麻栎生长性状基因型与环境间的互作效应是可行的,利用该模型作双标图对参试家系进行稳定性分析是较可靠的(表3)。
表 3 11年生麻栎不同家系AMMI分析
Table 3. AMMI analysis of 11-year-old Q. accutissim from different families
分析方法
Analysis method变异来源
Source of variation自由度
df平方和
Sum of square方差分量
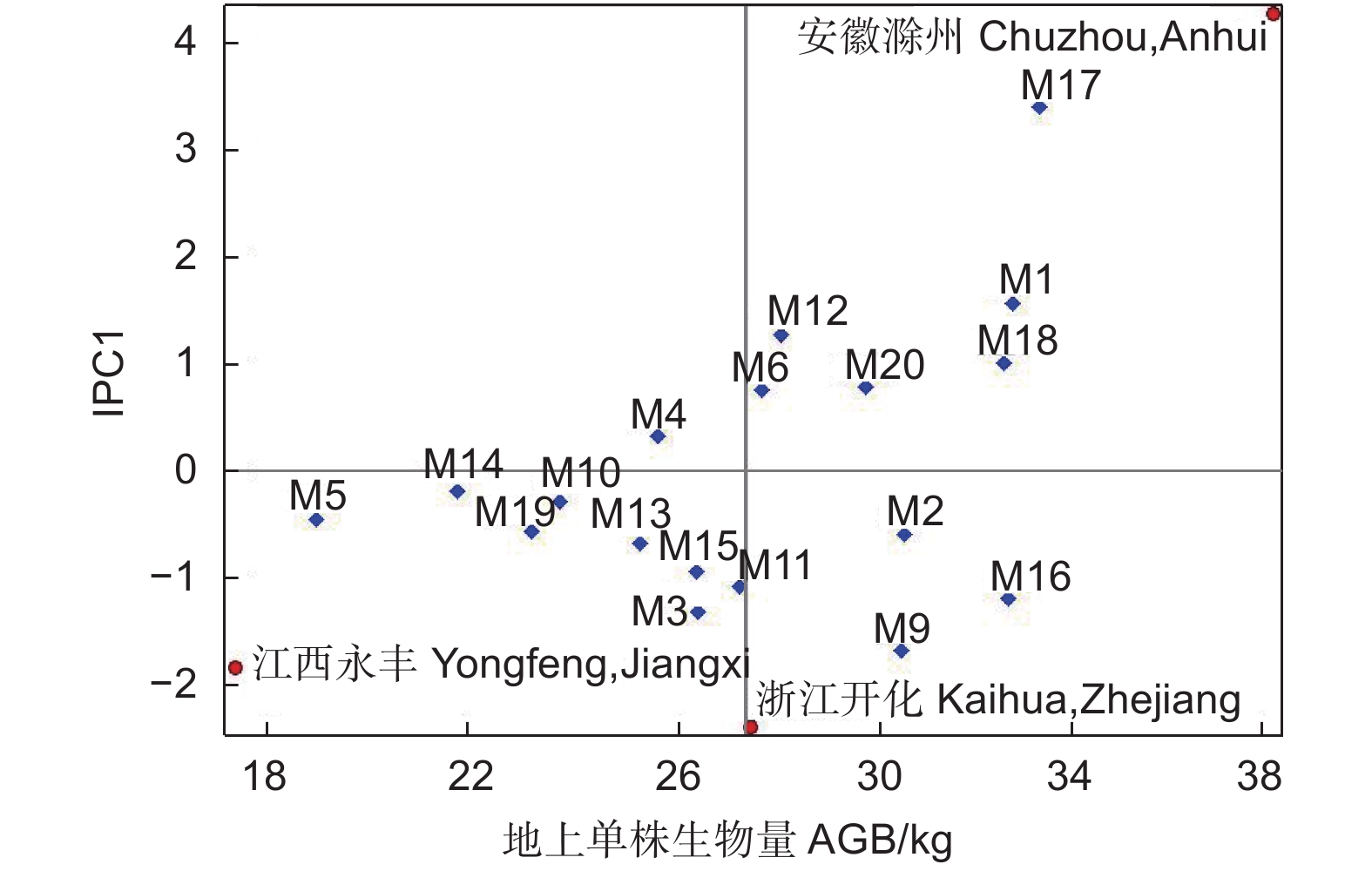
Variance component/%F AMMI模型 AMMI model PCA1 17 3732.42 59.46 9.71** 残差 21 1420.21 误差 101 1124.46 以各家系地上单株生物量值为横坐标,以家系与地点的IPC1值为纵坐标作双标图(图6)。图中X轴绝对值越大,家系生物量越大,Y轴绝对值越大,家系的稳定性越差。AMMI双标图显示:在横轴方向上,M17、M1、M18和M16等家系生长性状绝对值较大,生长相对较好,M5、M14、M19和M10等家系均值较小,生长相对较差;在纵轴方向上,M17、M9和M1等家系的IPC1值较大,说明这些家系与环境间的互作效应最强,对环境条件较为敏感,稳定性弱,而M14、M10和M4等家系的IPC1值最小,说明其受环境条件的影响小,稳定性较高。在横轴方向上,地点间分散程度均大于家系间分散程度,这说明同一家系在不同地点的生长表现差异较大。在纵轴方向上,以IPC1=0为分界线,安徽滁州试验点地上单株生物量对M17、M1和M18等家系具有正向的交互作用。江西永丰和浙江开化试验点对M9、M16和M2等家系也具有正向交互作用。

图 6 3个试验点麻栎地上单株生物量的双标图
Figure 6. Biplot of AGB per plant of Q. acutissima at 3 sites
为了更准确比较家系地点间的稳定性,分析了各家系的稳定性参数Dg(e),Dg(e)值越小,家系的稳定性越好。分析结果(表4)表明:M14、M10、M4家系的稳定性较好,Dg(e)均值为0.27;M20、M15和M18家系的稳定性中等,Dg(e)均值为0.90;M9和M17家系稳定性较差,Dg(e)均值为2.22。3个试验点地上单株生物量稳定性排名依次为:江西永丰>浙江开化>安徽滁州。
表 4 各试验点参试家系地上单株生物量交互作用主成分轴分量值及稳定性参数
Table 4. Principal component axial component value and stability parameter of the AGB per plant interaction of families in each test site
来源
Source of variation地上单株生物量均值
Mean AGB/kg离差
DeviationPCA1 稳定性参数
Dg(e)家系
FamilyM1 32.8 9 5.014 2 1.553 5 1.553 5 M2 30.7 9 2.910 0 −0.595 7 0.595 7 M3 26.7 8 −1.098 6 −1.314 5 1.314 5 M4 26.0 1 −1.867 5 0.316 7 0.316 7 M5 19.3 8 −8.492 6 −0.464 2 0.464 2 M6 28.0 2 0.144 3 0.749 0 0.749 0 M9 30.7 2 2.847 5 −1.678 0 1.678 0 M10 24.1 2 −3.755 4 −0.298 5 0.298 5 M11 27.6 0 −0.279 1 −1.089 8 1.089 8 M12 28.3 9 0.514 2 1.255 6 1.255 6 M13 25.6 8 −2.198 7 −0.675 8 0.675 8 M14 22.1 3 −5.751 7 −0.199 2 0.199 2 M15 26.7 6 −1.113 6 −0.940 6 0.940 6 M16 32.8 0 4.919 1 −1.199 7 1.199 7 M17 33.4 0 5.519 5 3.380 2 3.380 2 M18 32.7 2 4.843 3 0.993 4 0.993 4 M19 23.5 6 −4.319 8 −0.565 8 0.565 8 M20 30.0 4 2.164 8 1.553 5 0.773 5 地点 Site 江西永丰 Yongfeng, Jiangxi 17.8 4 −10.037 9 −1.842 1 1.842 1 浙江开化 Kaihua, Zhejiang 27.8 4 −0.041 4 −2.403 4 2.403 4 安徽滁州 Anhui, Chuzhou 37.9 6 10.079 4 4.245 5 4.245 5 -
以11年生麻栎地上单株生物量高出各试验点生物量均值的前30%为标准进行家系选择,江西永丰、浙江开化和安徽滁州试验点优良家系分别有3、2和3个。
江西永丰试验点筛选的优良家系为M11、M9和M15,分别高出该试验点生物量均值5.50 kg(遗传增益为7.45%)、3.81 kg(遗传增益为5.16%)和3.01 kg(遗传增益为4.08%)。浙江开化试验点筛选的优良家系为M16和M2,分别高出该试验点生物量均值22.97 kg(遗传增益为37.02%)和17.38 kg(遗传增益为28.01%)。安徽滁州试验点筛选的优良家系为M17、M1和M18,分别高出该试验点生物量均值20.37 kg(遗传增益为30.29%),12.00 kg(遗传增益为17.82%)和10.32 kg(遗传增益为15.05%)。
对3个试验点麻栎家系地上单株生物量生长和稳定性进行综合分析,在参试18个家系中,生长稳定且生长优势相对均较好的家系为M18和M20,地上单株生物量均值为31.38 kg,Dg(e)均值为0.88。
-
林木遗传变异是遗传改良的基础,并决定了林木在改良过程中的潜力[19]。研究表明,具有相同遗传结构的树种在个体、家系和种源间均存在较大的遗传变异,不同的环境和栽培措施下生长性状具有差异性[20-21]。本试验发现,麻栎优树自由授粉家系的树高、胸径和地上单株生物量在家系间均存在极显著的差异,表明生长性状存在丰富的家系变异基础,这为优良家系的选择提供了空间。林木间的生长差异是在遗传和环境的共同作用下形成的,欧阳芳群等[22]研究发现,13年生青海云杉(Picea crassifolia Kom.)自由授粉家系的树高等生长性状间存在家系与试验点的互作效应,明确了青海云杉家系的生长不仅受遗传控制,还受遗传与试验地环境互作的影响。此外,在较大的地理尺度上,气候条件和试验地土壤状况是影响植物分布和生长的关键制约因子[23]。本试验发现麻栎优树自由授粉家系地上单株生物量受到遗传和试验地环境的共同影响,且试验地环境对生长的影响较大,随着林龄增长呈增加趋势,这表明不同地点的环境条件是影响麻栎地上单株生物量的主要因素。
遗传力是最重要遗传参数之一,它反映了在群体中根据表型优劣选择基因型的可靠程度,通过估算家系和单株遗传力可以确定选择方式和选择强度,对于育种策略制定具有重要的指导意义[24]。Diao等[11]研究表明,日本落叶松生长性状的家系遗传力和单株遗传力随林龄增长呈现动态变化。任华东等[25]研究发现,黑荆树(Acacia mearnsii De Wilde)家系生长变化较平稳,生长差异主要受遗传控制。本试验发现,随着林龄增加,麻栎地上单株生物量的家系遗传力和单株遗传力均有下降趋势,受遗传控制逐渐减弱,家系遗传力和单株遗传力间的差距呈减少趋势,试验地环境因子对于麻栎家系生长的影响逐渐增大。麻栎自由授粉家系地上单株生物量的表型变异系数和遗传变异系数先升高后降低,说明在不同林龄阶段地上单株生物量受遗传效应影响是不同的。
研究发现,林木早期与后期生长关系密切,早期的生长变异可以为后期生长选择提供参考[26],对林木进行早期相关评价可以间接估测后期生长表现,以此加快育种进程[27-28]。本试验对麻栎优树动态生长表现分析发现,1~3年生与6~11年生时的生长表现并不一致,如在前期(1~3年生)生长较好的种源,在后期(6~11年生)的表现却一般。麻栎优树自由授粉家系早期生长并不能完全反映后期的生长表现,因此,针对麻栎短轮伐期炭用林培育(≤10 a),在3年生后开展麻栎优良家系的筛选较为适宜。林木遗传稳定性能够调节其表型的特征,使之能适应环境变化,维持平稳的生理和生殖特性,从而保持稳定状态[29]。本试验利用稳定性并结合地上单株生物量分析,对麻栎优树自由授粉家系进行了生长量及稳定性的评价,并分别为江西、浙江和安徽地区筛选出优良家系,可作为下世代育种亲本。
-
以不同林龄麻栎家系试验林为研究对象,通过对18个优树自由授粉家系在3个试验点的树高、胸径、地上单株生物量的生长研究发现,麻栎家系间生长差异显著,1~3年生与6~11年生优良家系表现具有巨大差异,随着林龄增长,麻栎优树自由授粉家系生长性状存在显著的地点和家系互作效应,其中,试验地环境因素对家系生长影响较大。本试验筛选出江西永丰试验点优良家系为M11、M9和M15,浙江开化试验点优良家系为M16和M2,安徽滁州试验点优良家系为M17、M1和M18,在3个试验地生长稳定且生长优势相对均较好的家系为M18和M20。
麻栎优树自由授粉家系生长性状3地点间动态变异及优良家系选择
Dynamic Variation of Growth Traits in Open-pollinated Families of Quercus acutissima Superior Tree and Selection of Superior Families Among Three Sites
-
摘要:
目的 研究麻栎优树自由授粉家系随林龄变化的生长性状遗传变异规律,为麻栎人工林高效培育筛选优良种质材料。 方法 在江西永丰、浙江开化和安徽滁州3个试验点,以来自广西融水的18个优树自由授粉家系试验林为研究对象,分析家系间林木生长变异规律、主要经济性状(地上单株生物量)随林龄的动态变化,基于AMMI模型进行生长性状稳定性评价,并进行优良家系选择。 结果 3个试验点麻栎家系间的树高、胸(地)径和地上单株生物量均具有显著差异。麻栎地上单株生物量受地点、家系、家系 × 地点交互作用显著影响,其中地点对生长变异影响最大,其次为家系和家系 × 地点。不同试验地点麻栎1~3年生和6~11年生家系的生长表现具有较大差异。根据11年生时地上单株生物量分别筛选试验地表现较好的家系,江西永丰试验点优良家系为M11、M9和M15,遗传增益为4.08%~7.45%;浙江开化试验点优良家系为M16和M2,遗传增益为28.01%~37.02%;安徽滁州试验点优良家系为M17、M1和M18,遗传增益为15.05%~30.29%。综合生长表现和稳定性分析发现,M18和M20家系适宜在3个试验点栽培,地上单株生物量均值为31.38 kg,Dg(e)均值为0.88。 结论 麻栎优树自由授粉家系间生长受地点和家系交互作用影响,筛选出一批适于不同区域人工林培育的优良家系。 Abstract:Objective To study the genetic variation of growth traits in superior open-pollinated families of Quercus acutissima with the increase of stand age, and to select the superior families for Q. acutissima plantation cultivation. Method Eighteen superior tree open-pollination families coming from Rongshui of Guangxi Zhuang Autonomous Region were planted in 3 test sites (Yongfeng of Jiangxi Province, Kaihua of Zhejiang Province and Chuzhou of Anhui Province). The variation in tree growth among the families and the dynamic changes of above-ground biomass of individual tree (AGB) with stand age were investigated. The stability of growth traits was evaluated using the AMMI model, and then the superior families were selected. Result The tree height, diameter at breast (or ground) height, and the AGB significantly differed among the 3 test sites. The AGB of Q. acutissima trees was significantly affected by the site, family, and family × site. The site had the greatest impact on growth variation, followed by family and family × site. The growth performance of 1-3 years old and 6-11 years old families had great difference among test sites. Based on the AGB at the 11st year, the superior families in Yongfeng were M11, M9 and M15, their genetic gain was 4.08%-7.45%; the superior families in the Kaihua were M16 and M2, their genetic gain was 28.01%-37.02%; the superior families in the Chuzhou were M17, M1 and M18, their genetic gain was 15.05%-30.29%. According to growth performance and stability analysis, it was showed that the families M18 and M20 were suitable for cultivation at all the 3 test sites. The average biomass of the AGB was 31.38 kg, and the average Dg(e) was 0.88. Conclusion The growth of superior open-pollinated families of Q. acutissima will be affected by the interaction of site and family, and some superior families were selected for Q. acutissima plantation cultivation. -
Key words:
- Quercus acutissima
- / open-pollinated family
- / growth trait
- / genetic variation
-
图 1 3个地点不同家系地上单株生物量随林龄的变化特征
Figure 1. Variation characteristics of AGB per plant in 3 sites among different families with the increase of stand age
图 2 3个试验点典型家系地上单株生物量随林龄变化
Figure 2. Variation of AGB per plant of typical families in 3 test sites with the increase of stand age
图 3 地点、家系及互作效应对地上单株生物量的相对影响随林龄的变化特征
Figure 3. Effect of site, family and their interaction on AGB per plant with the increase of stand age
图 4 地上单株生物量的遗传力随林龄的变化趋势
Figure 4. Variation heritability of AGB per plant with the increase of stand age
图 5 地上单株生物量变异系数随林龄的变化趋势
Figure 5. Variation coefficient of AGB per plant with the increase of stand age
表 1 3个试验地点不同林龄家系胸(地)径、树高和地上单株生物量的统计和方差分析
Table 1. Statistics and variance analysis of DBH(D), H and AGB for family with stand ages in 3 sites
试验点
Site性状
Trait林龄
Stand age/a均值
Mean变幅
Variable rang变异系数
CV/%方差分量百分比
Variance component percentage /%家系
Family区组
Block区组 × 家系
Block × Family机误
Error江西永丰
Yongfeng,Jiangxi胸(地)径
DBH(D)/cm1 1.58 1.33~1.79 6.99 21.48** 2.03** 0.86** 0.74 3 4.09 3.61~4.94 8.93 38.98** 3.95** 0.71** 0.19 6 3.69 2.72~4.97 14.56 2.47** 0.48** 0.19** 0.04 8 5.48 4.10~6.51 10.50 24.82** 1.46* 1.26** 0.33 11 7.50 6.02~8.71 8.75 8.26 13.65** 5.24** 2.97** 树高
H/m1 1.16 1.00~1.30 6.07 11.48** 1.61** 0.28** 0.07 3 2.52 1.80~2.72 8.50 50.71** 4.17** 1.27** 0.47 6 4.26 3.78~4.94 7.39 2.43** 0.71** 0.29** 0.04 8 5.56 4.73~6.24 7.28 4.68** 2.63** 0.69** 0.19 11 6.86 5.93~7.61 5.97 6.39** 0.98** 0.25** 0.01 地上单株生物量
AGB/kg1 0.03 0.02~0.04 13.62 0.37** 0.12** 0.08** 0.01 3 0.35 0.21~0.73 38.14 1.36** 0.96** 0.13** 0.02 6 3.48 1.91~5.96 29.90 1.36** 0.96** 0.23* 0.12 8 8.32 4.42~12.02 22.56 3.32** 0.29** 0.12** 0.09 11 16.66 10.13~23.03 19.03 6.39** 0.98** 0.25** 0.01 浙江开化
Kaihua,
Zhejiang胸(地)径
DBH(D)/cm1 1.07 0.83~1.24 7.93 13.25** 2.05** 1.61** 0.03 3 2.68 2.01~3.54 14.25 15.57** 3.72** 0.47** 0.36 6 5.32 4.19~6.56 12.96 35.55** 2.35** 1.07* 0.16 8 7.08 5.66~8.24 8.24 14.61** 1.87** 2.33** 0.50 11 8.96 7.28~11.38 12.07 65.11** 4.08** 3.69** 0.43 树高
H/m1 0.95 0.74~1.14 9.64 26.22** 1.09** 0.31** 0.01 3 2.03 1.83~2.35 8.10 30.20** 0.72** 0.33** 0.13 6 4.50 4.07~5.21 6.42 12.67** 0.85** 0.92** 0.03 8 6.51 5.56~7.32 6.93 58.30** 0.89** 1.15** 0.05 11 7.86 6.72~9.37 9.54 47.53** 2.28** 1.79** 0.25 地上单株生物量
AGB/kg1 0.01 0.01~0.02 7.55 6.39** 2.56** 1.45** 0.12 3 0.09 0.05~0.19 36.44 1.87** 1.22** 0.12** 0.04 6 6.66 4.06~10.49 25.82 0.63** 0.28** 0.09** 0.04 8 14.47 8.84~20.31 18.43 38.53** 3.98** 0.10** 0.08 11 25.31 15.43~42.37 27.71 6.22* 2.59** 1.20** 0.63 安徽滁州
Anhui, Chuzhou胸(地)径
DBH(D)/cm1 1.72 1.43~1.72 5.31 46.89** 2.08** 0.76** 0.04 3 4.43 2.47~4.43 12.59 20.33** 0.58** 0.53** 0.13 6 6.45 4.07~6.45 12.07 59.96** 1.83** 1.69** 0.19 8 9.52 6.98~9.52 8.16 30.04** 2.46** 2.36** 0.28 11 12.73 9.32~12.73 9.31 42.71** 4.99** 3.89** 0.34 树高
H/m1 1.18 1.05~1.29 5.36 30.15** 1.31** 0.25** 0.14 3 4.42 3.72~4.94 6.84 16.96 + 3.13** 0.13** 0.20 6 5.28 3.73~6.20 14.61 29.93** 0.62** 0.56** 0.25 8 6.81 4.94~7.63 11.94 11.30** 1.61** 0.93** 0.06 11 8.75 7.6~10.13 7.83 21.78** 2.73** 1.65** 0.09 地上单株生物量
AGB/kg1 0.03 0.02~0.03 7.80 1.87** 0.22** 0.11** 0.05 3 0.24 0.07~0.45 38.90 2.25** 0.32* 0.11** 0.01 6 7.95 4.36~11.79 27.41 1.68** 0.22** 0.12* 0.06 8 19.14 11.89~25.27 18.99 0.93** 0.39** 0.08** 0.01 11 36.67 26.84~54.95 22.10 0.69** 0.15* 0.09** 0.06 注:**表示0.01水平差异显著,*表示0.05水平差异显著。下同。
Note: **indicates significant difference at P<0.01, *indicates significant difference at P<0.05. The same below. 下载: 导出CSV
下载: 导出CSV
表 2 参试家系地上单株生物量的3因素方差分析
Table 2. Three-way variance analysis of the AGB per plant in the test families
生长性状
Growth trait变异来源
Source of variation自由度
df均方
Mean SquareF 总的 Total 321 64.27 地上单株生物量
AGB林龄 Stand age 4 44323.95 769.02** 地点 Site 2 2722.79 94.12** 家系 Family 17 109.33 102.49** 地点 × 林龄 Site × Stand age 8 1737.27 30.14** 家系 × 林龄 Family × Stand age 68 43.74 212.75** 地点 × 家系 Site × Family 34 77.27 155.12** 地点 × 家系 × 林龄 Site × Family × Stand age 136 49.81 86.44** 误差 Error 72 31.06
下载: 导出CSV
表 3 11年生麻栎不同家系AMMI分析
Table 3. AMMI analysis of 11-year-old Q. accutissim from different families
分析方法
Analysis method变异来源
Source of variation自由度
df平方和
Sum of square方差分量
Variance component/%F AMMI模型 AMMI model PCA1 17 3732.42 59.46 9.71** 残差 21 1420.21 误差 101 1124.46
下载: 导出CSV
表 4 各试验点参试家系地上单株生物量交互作用主成分轴分量值及稳定性参数
Table 4. Principal component axial component value and stability parameter of the AGB per plant interaction of families in each test site
来源
Source of variation地上单株生物量均值
Mean AGB/kg离差
DeviationPCA1 稳定性参数
Dg(e)家系
FamilyM1 32.8 9 5.014 2 1.553 5 1.553 5 M2 30.7 9 2.910 0 −0.595 7 0.595 7 M3 26.7 8 −1.098 6 −1.314 5 1.314 5 M4 26.0 1 −1.867 5 0.316 7 0.316 7 M5 19.3 8 −8.492 6 −0.464 2 0.464 2 M6 28.0 2 0.144 3 0.749 0 0.749 0 M9 30.7 2 2.847 5 −1.678 0 1.678 0 M10 24.1 2 −3.755 4 −0.298 5 0.298 5 M11 27.6 0 −0.279 1 −1.089 8 1.089 8 M12 28.3 9 0.514 2 1.255 6 1.255 6 M13 25.6 8 −2.198 7 −0.675 8 0.675 8 M14 22.1 3 −5.751 7 −0.199 2 0.199 2 M15 26.7 6 −1.113 6 −0.940 6 0.940 6 M16 32.8 0 4.919 1 −1.199 7 1.199 7 M17 33.4 0 5.519 5 3.380 2 3.380 2 M18 32.7 2 4.843 3 0.993 4 0.993 4 M19 23.5 6 −4.319 8 −0.565 8 0.565 8 M20 30.0 4 2.164 8 1.553 5 0.773 5 地点 Site 江西永丰 Yongfeng, Jiangxi 17.8 4 −10.037 9 −1.842 1 1.842 1 浙江开化 Kaihua, Zhejiang 27.8 4 −0.041 4 −2.403 4 2.403 4 安徽滁州 Anhui, Chuzhou 37.9 6 10.079 4 4.245 5 4.245 5
下载: 导出CSV
-
[1] 白天道, 徐立安, 王章荣, 等. 马尾松实生种子园自由授粉子代测定及亲本家系选择增益估算[J]. 林业科学研究, 2012, 25(4):449-449. doi: 10.3969/j.issn.1001-1498.2012.04.007 [2] White T L, Adams W T, Neale D B. 森林遗传学[M]. 崔建国, 李火根, 译. 北京: 科学出版社, 2013. [3] 解懿妮, 刘青华, 蔡燕灵, 等. 5年生马尾松生长性状3地点家系变异及评价[J]. 林业科学研究, 2020, 33(5):2-10. [4] 袁虎威. 油松高轮次遗传改良关键技术与策略[D]. 北京: 北京林业大学, 2016. [5] 季孔庶, 樊民亮, 徐立安. 马尾松无性系种子园半同胞子代变异分析和家系选择[J]. 林业科学, 2005, 41(6):43-49. doi: 10.3321/j.issn:1001-7488.2005.06.007 [6] 王云鹏, 张 蕊, 周志春, 等. 木荷优树自由授粉家系早期生长性状遗传变异动态规律[J]. 林业科学, 2020, 56(9):78-86. [7] 韩东花, 杨桂娟, 肖 遥, 等. 楸树无性系早期生长变异和优选[J]. 林业科学研究, 2019, 32(4):97-104. [8] 刘 宇, 徐焕文, 李志新, 等. 白桦杂交子代家系生长变异及稳定性分析[J]. 植物研究, 2015, 35(6):937-944. doi: 10.7525/j.issn.1673-5102.2015.06.023 [9] 赖 猛, 孙晓梅, 张守攻. 日本落叶松及其杂种无性系间的物候变异与早期选择[J]. 林业科学, 2014, 50(7):53-57. [10] 汤珧华, 李志辉. 耐寒性桉树早期选择——Ⅰ. 耐寒性桉树早期选择研究综述[J]. 中南林学院学报, 2000, 20(3):70-74. [11] Diao S, Hou Y M, Xie Y H, et al. Age trends of genetic parameters, early selection and family by site interactions for growth traits in Larix kaempferi open-pollinated families[J]. Bmc Genetics, 2016, 17(1): 104-116. doi: 10.1186/s12863-016-0400-7 [12] 辛娜娜, 张 蕊, 范辉华, 等. 5年生木荷生长和形质性状的家系变异和选择[J]. 林业科学研究, 2014, 27(3):316-316. [13] 张兴旺, 李 垚, 方炎明. 麻栎在中国的地理分布及潜在分布区预测[J]. 西北植物学报, 2014, 34(8):1685-1692. [14] 成向荣, 虞木奎, 葛 乐, 等. 不同间伐强度下麻栎人工林碳密度及其空间分布[J]. 应用生态学报, 2012, 23(5):1175-1180. [15] 王子寅, 唐罗忠, 赵 丹, 等. 两种立地条件下麻栎人工林地上部分养分的积累和分配[J]. 植物生态学报, 2010, 34(6):661-670. doi: 10.3773/j.issn.1005-264x.2010.06.005 [16] 王 霞, 胡海波, 张世豪, 等. 不同林龄麻栎林地下部分生物量与碳储量研究[J]. 生态学报, 2019, 39(22):8556-8564. [17] 苑海静, 邢文黎, 徐海东, 等. 麻栎不同种源幼林生长和生物量的变异[J]. 林业科学研究, 2020, 33(5):97-105. [18] 唐启义. DPS数据处理系统[M]. 北京: 科学出版社, 2013. [19] 林 磊, 周志春, 范辉华, 等. 木荷优树子代苗期生长遗传和变异研究[J]. 林业科学研究, 2009, 22(2):155-160. doi: 10.3321/j.issn:1001-1498.2009.02.001 [20] Zhang Z, Jin G, Feng Z, et al. Joint influence of genetic origin and climate on the growth of Masson pine (Pinus massoniana Lamb. ) in China[J]. Scientific Reports, 2020, 10(1): 4653. doi: 10.1038/s41598-020-61597-9 [21] 杨桂娟, 胡海帆, 孙洪刚, 等. 林分年龄, 造林密度和林分自然稀疏对杉木人工林个体大小分化和生产力关系的影响[J]. 林业科学, 2019, 55(11):126-136. doi: 10.11707/j.1001-7488.20191114 [22] 欧阳芳群, 祁生秀, 蔡启山, 等. 青海云杉自由授粉家系遗传评价与选择[J]. 林业科学研究, 2018, 31(6):29-35. [23] Zizumbo-Villarreal D, Piñero D. Pattern of morphological variation and diversity of Cocos nucifera (Arecaceae) in Mexico[J]. American Journal of Botany, 1998, 85(6): 855-865. doi: 10.2307/2446421 [24] Baltunis B S, Gapare W J, Wu H X et al. Genetic parameters and genotype by environment interaction in radiata pine for growth and wood quality traits in Australia[J]. Silvae Genetica, 2010, 59(6): 113-124. [25] 任华东, 姚小华, 康文玲, 等. 黑荆树种源和家系的遗传变异与早期选择[J]. 林业科学, 2010, 46(3):153-160. doi: 10.11707/j.1001-7488.20100324 [26] 肖 遥, 姚淑均, 杨桂娟, 等. 滇楸无性系早期木材性状变异与评价[J]. 林业科学研究, 2019, 32(4):79-87. [27] 赵 阳, 毕泉鑫, 句 娇, 等. 文冠果种子及苗期生长性状地理种源变异[J]. 林业科学研究, 2019, 32(1):163-171. [28] 廖怀建, 邓 疆, 杜 婷, 等. 引进种源印度黄檀优株选择及优良无性系选择与评价[J]. 林业科学研究, 2017, 30(6):916-920. [29] 李红盛, 汪阳东, 徐刚标, 等. 山苍子家系幼林生长性状遗传变异及稳定性分析[J]. 林业科学研究, 2018, 31(5):168-175. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4754
- HTML全文浏览量: 2673
- PDF下载量: 85
- 被引次数: 0

























