


-
中国内陆盐碱地以盐化和碱化混合土壤为主, 成分复杂且程度各异, 统称为盐碱地土壤[1],盐碱化已成为威胁生态环境和农业可持续发展的重要因素之一。我国盐碱地所占面积为9.9 × 107 hm2[2]。据全国第二次调查报告显示,我国盐碱地主要分布在东北(黑吉辽)、华北(京津冀)、中北(晋秦陇)、西北(青宁新)等地区[3],具有分布广、面积大、类型多的特点。河南分布着大面积的盐碱地,这严重制约了河南农林经济和植物景观建设的发展。
开发和利用盐碱地资源是进一步挖掘农业发展潜力、解决人口和环境压力的重要举措,而选用和推广耐盐植物是改良和利用大面积盐碱土壤最经济、最快捷的措施之一[4];但盐碱地区适生范围的确定,是耐盐碱植物大面积推广亟待解决的关键技术。因此,国内外众多科研工作者对研究盐碱胁迫下植物的生理响应机制愈加重视,主要集中在蔬菜、农作物或特定耐盐模式植物的研究[5-7]。
许多研究表明,盐碱胁迫对植物的生理系统影响十分复杂,植株不仅遭受着盐胁迫引起的渗透伤害和离子毒害,还受到高pH的损害[8]。慈敦伟等[9]通过试验发现,低浓度盐可以促进花生植株的生长,而高浓度的盐则会抑制其生长。张晓燕等[10]研究表明,在一定盐胁迫范围内,文冠果(Xanthoceras sorbifolia Bunge)通过调节体内渗透压变化,而表现出一定的耐盐性;此外,郭瑞等[11]研究证明,碱胁迫的高pH值影响植物对矿质元素的吸收,阻碍离子稳态重建,显著影响植物根系生长发育。因此,盐碱化植物既要通过渗透调节减缓渗透伤害和离子毒害,又要维持体内的pH平衡[12-13]。目前,文冠果多以研究雄性不育、遗传变异、快速繁殖及干旱胁迫等为主,在针对中性盐和碱性盐方面的研究鲜有报道。
文冠果是我国特有的珍稀木本油料树种,被列为制造生物柴油的八大树种之一,素有“北方油茶”的美称[14-15];其根系发达,萌蘖力强,对环境具有较强的适应性,在我国华北和东北、西北等地区均有分布[16]。为避免盲目引种造成的经济损失,研究盐碱胁迫下文冠果的生理机制显得尤为迫切。本试验选用NaCl和Na2CO3模拟河南省盐碱土壤,研究盐碱胁迫下其质膜透性、渗透调节物质及抗氧化酶活性等的生理指标,对探究文冠果耐盐碱能力及耐盐碱生理响应机制具有重要意义,也为可持续利用盐碱地资源提供理论参考。
-
本试验在河南牧业经济学院苗圃(113°65′ E,34°78′ N)进行,2018年1月选用河南济源种源的文冠果种子进行沙藏处理,2018年3月,将露白的种子直播在规格为280 mm × 180 mm的花盆内,基质V草炭土∶V珍珠岩 = 5∶1,花盆配置的托盘有效收集其渗出液并及时回浇至盆中,以避免土壤盐分的流失。将种植的幼苗置于3 m × 3 m的避雨棚下,并在花盆底部作垫高处理,所有植株统一管理。
-
2019年7月选取长势良好且基本一致的81株幼苗进行盐碱处理。将试验材料随机分成9份,每份9株,根据河南地区常见中性盐(NaCl)和碱性盐(Na2CO3)以及Na+浓度[17],设置NaCl浓度分别为:70、140、210、280 mmol·L−1;Na2CO3浓度分别为:35、70、105、140 mmol·L−1,共8个盐处理,以浇等体积的清水为对照(CK),每3株为1组,3次重复。各处理每次浇相应浓度的盐、碱溶液300 mL,为避免盐激发生,盐、碱溶液从低浓度(为最终浓度的1/4)开始加入。每3 d递增1个梯度,4次后累计浇灌盐、碱溶液的总量达到设计各处理的浓度,开始计算处理时间。处理后的7、12、17、22 d时采样进行各生理指标的测定。
-
采用刘正祥等[18]的方法进行生物量和根冠比的测定;质膜透性的测定采用电导仪[19]测定;丙二醛(MDA)含量采用硫代巴比妥酸法[20]测定;脯氨酸含量采用酸性茚三酮比色法[20]测定;可溶性蛋白含量采用考马斯亮蓝法[21]测定;超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)光化学还原法[21]测定。
-
采用SPSS24.0进行数据分析,并用WPS 2019整理以及绘制图表,进行双因素方差分析(Two-way ANOVA)检验不同浓度、时间和二者交互作用的影响以及单因素方差分析(One-way ANOVA)检验不同处理间的差异,LSD法进行多重比较,并对各指标进行相关性分析和主成分分析。
-
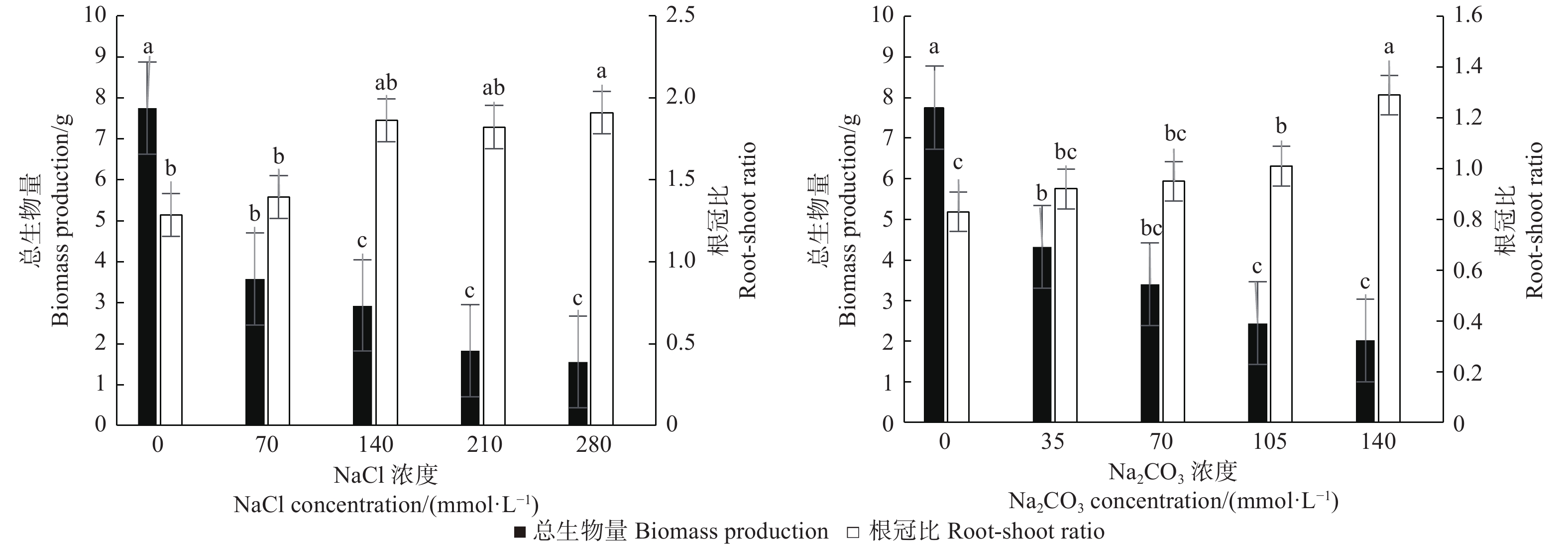
图1表明:不同盐浓度下植株的总生物量、根冠比存在差异,随着盐碱胁迫加剧,文冠果总生物量呈降低趋势,而根冠比呈上升趋势;不同浓度的中性盐和碱性盐对植株生长影响不同,280 mmol·L−1的中性盐和140 mmol·L−1碱性盐处理下,植株的总生物量最低,根冠比最高。此外,280 mmol·L−1中性盐和140 mmol·L−1碱性盐处理比CK的总生物量分别显著降低79.9%、73.9%(P < 0.05),而根冠比比CK分别显著增加48.5%、55.4%(P < 0.05)。
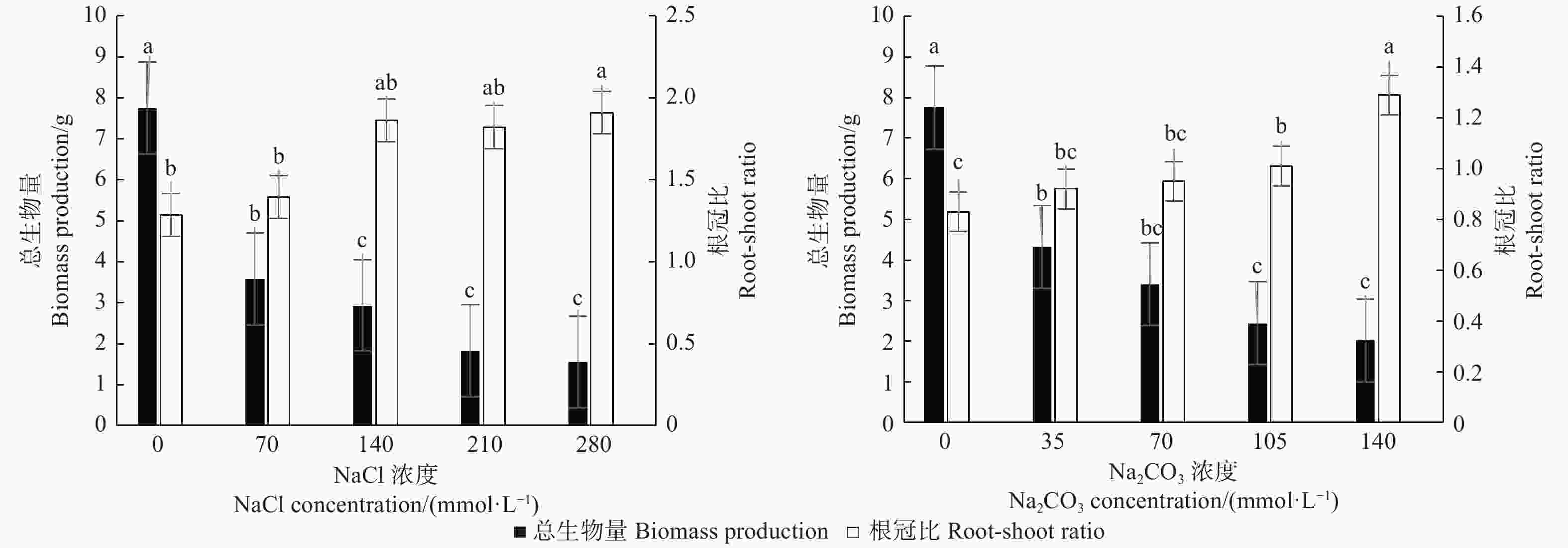
图 1 盐碱处理对文冠果幼苗总生物量和根冠比的影响
Figure 1. The effect of saline-alkali treatment on biomass production and root-shoot ratio of Xanthoceras sorbifolia seedlings
-
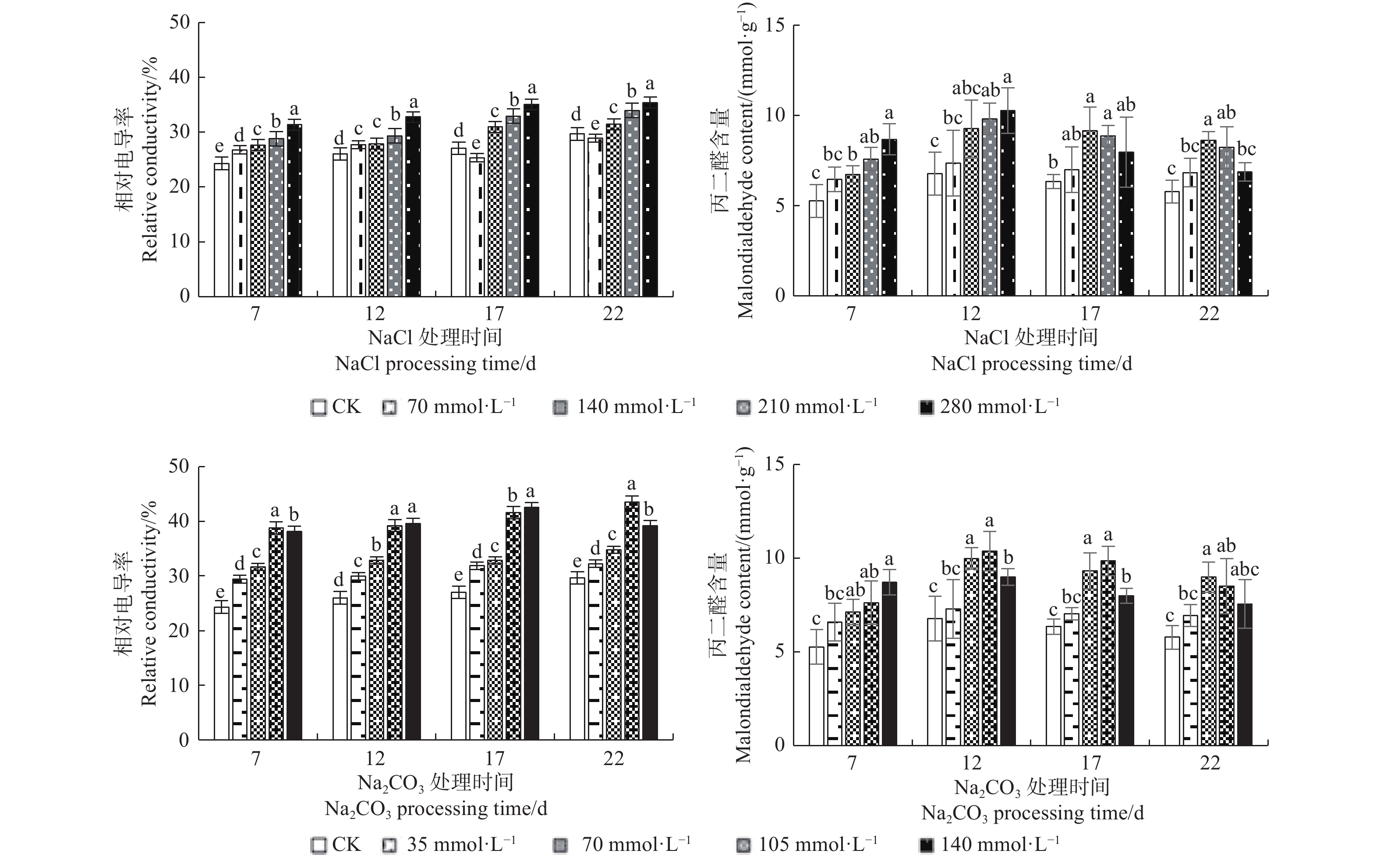
由图2可知:盐碱处理浓度较低且处理时间较短时,文冠果叶片相对电导率、丙二醛含量均呈上升趋势,相对电导率显著高于对照组,在碱胁迫22 d、140 mmol·L−1时,相对电导率开始降低;在NaCl浓度为210 mmol·L−1、Na2CO3浓度为70 mmol·L−1 胁迫12 d时丙二醛含量比CK显著增多,且在NaCl浓度为280 mmol·L−1、Na2CO3浓度为105 mmol·L−1时达最高值;此外,高浓度(NaCl 210、280 mmol·L−1和Na2CO3 105、140 mmol·L−1)长时间(17、22 d)的处理下,丙二醛含量呈下降趋势。说明文冠果具有一定的抗盐能力,且在高浓度盐碱胁迫下植物细胞抵御外界逆境的能力增强,但胁迫后期幼苗叶片依旧受害严重,甚至使植株死亡。

图 2 盐碱处理对文冠果幼苗叶片相对电导率和丙二醛的影响
Figure 2. The effect of saline-alkali treatment on relative electrical conductivity and Malondialdehyde in the leaves of Xanthoceras sorbifolia seedlings
-
图3表明:随着NaCl处理浓度的增加及处理时间的延长,文冠果幼苗叶片脯氨酸及可溶性蛋白含量均呈上升趋势,在浓度为280 mmol·L−1、处理22 d时达到峰值,分别为30.35 μg·g−1、3.724 mg·g−1,与对照(CK)相比分别增加了211.70%、647.80%。在高浓度(105、140 mmol·L−1)Na2CO3胁迫下,脯氨酸含量随着时间的延长呈先升后降趋势,两浓度处理间差异不显著(P > 0.05),在140 mmol·L−1,处理17 d时达到最大值(26.97 μg·g−1);可溶性蛋白含量随处理时间的延长呈上升趋势,在高浓度(140 mmol·L−1)长时间(22 d)处理下达到峰值(3.765 mg·g−1)。由此说明,盐碱的类型、浓度及处理时间均对文冠果的生理状况有一定影响,随着处理浓度的增加及处理时间的延长,幼苗被破坏程度逐步加深,最终导致植株死亡。
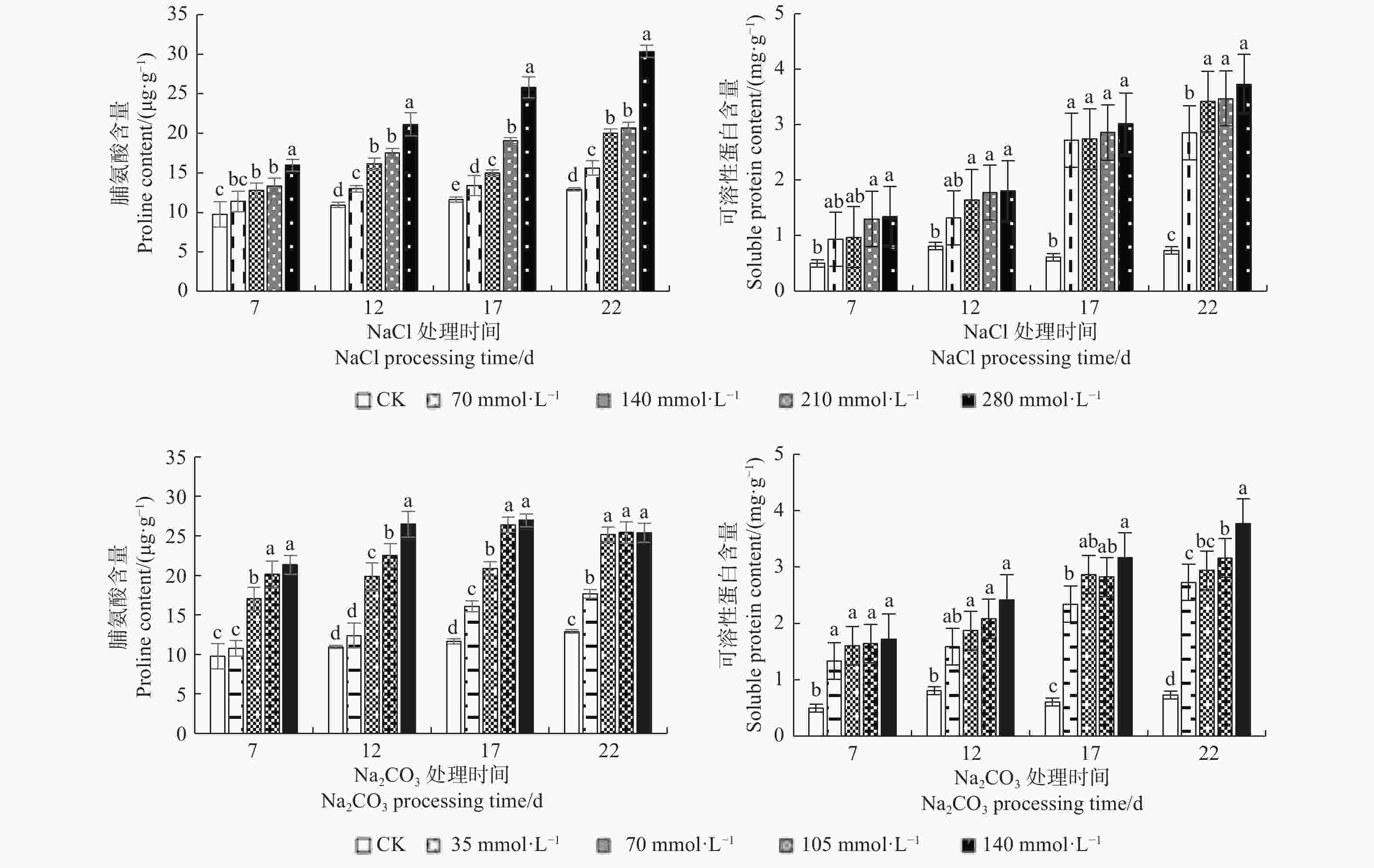
图 3 盐碱处理对文冠果幼苗叶片脯氨酸和可溶性蛋白含量的影响
Figure 3. The effect of saline-alkali treatment on the content of proline and soluble protein in the leaves of Xanthoceras sorbifolia seelings
-
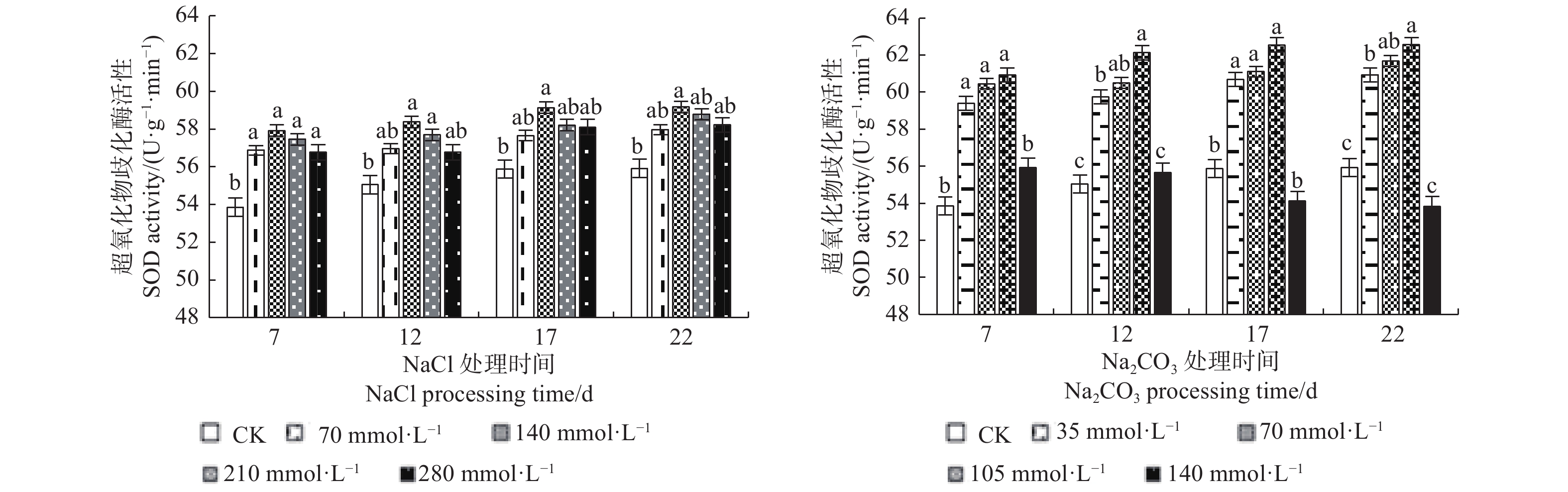
由图4可见:NaCl和Na2CO3处理下的文冠果超氧化物歧化酶(SOD)活性变化规律基本一致,随浓度的增大均呈先升后降的趋势,且表现为碱胁迫>盐胁迫;在NaCl浓度为140 mmol·L−1、Na2CO3浓度为105 mmol·L−1处理17 d时SOD活性均达到峰值,比CK分别增加了5.81%、11.95%。
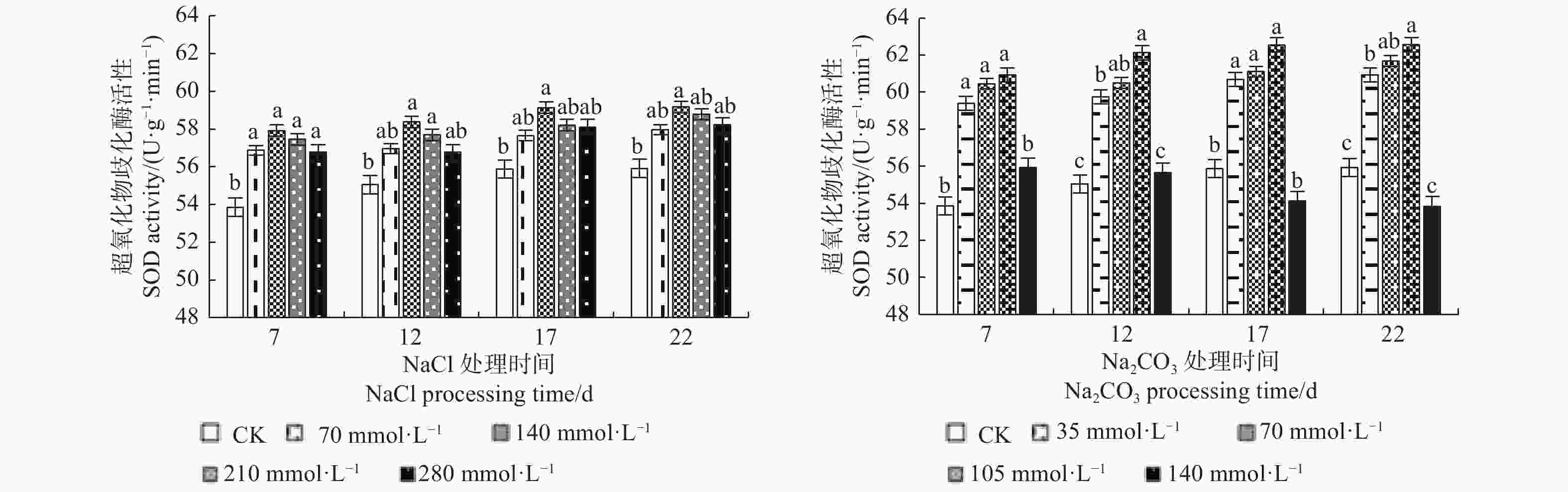
图 4 盐碱处理对文冠果幼苗叶片SOD活性的影响
Figure 4. The effect of saline-alkali treatment on SOD activity in the leaves of Xanthoceras sorbifolia seedlings
-
由表1可知:时间和浓度对盐碱胁迫下的文冠果相对电导率、脯氨酸均有极显著交互效应(P < 0.01),而可溶性蛋白仅在盐胁迫下受到时间和浓度交互作用的影响(P < 0.01)。盐碱胁迫的时间和浓度双因素对丙二醛和SOD均无显著的交互作用(P > 0.05)。
表 1 盐碱处理下各生理指标的双因素方差分析
Table 1. Two-way ANOVA of various physiological indexes under saline-alkali treatment
处理
Treatment影响因子
Impact factors相对电导率
Relative conductivity丙二醛
Malondiadehyde脯氨酸
Proline可溶性蛋白
Soluble protein超氧化物歧化酶
SODNaCl 时间 Time 1636.66** 14.03** 317.79** 62.39** 3.92* 浓度 Concentration 802.63** 7.82** 169.83** 101.64** 10.72** 时间 × 浓度 Time × concentration 68.42** NS 15.65** 6.15** NS Na2CO3 时间 Time 472.36** 8.81** 64.30** 48.78** NS 浓度 Concentration 5602.78** 24.49** 309.37** 74.78** 53.25** 时间 × 浓度 Time × concentration 48.51** NS 4.52** NS NS 注:*表示差异显著(P < 0.05),**表示差异极显著(P < 0.01),NS表示差异不显著;表中数据代表F值,误差项省略。
Notes:*, **mean significance at 0.05 level and 0.001 level,respectively,NS mean no significance. The data in the table represent the F value, and the error item is omitted. -
由表2可知:NaCl处理下,丙二醛、脯氨酸、可溶性蛋白与相对电导率及丙二醛与脯氨酸、可溶性蛋白、SOD均呈极显著正相关(P < 0.01);可溶性蛋白与SOD、脯氨酸均呈极显著正相关(P < 0.01);SOD与相对电导率、脯氨酸无显著相关(P > 0.05)。Na2CO3处理下,丙二醛、脯氨酸、可溶性蛋白与相对电导率及丙二醛与脯氨酸、可溶性蛋白均呈极显著正相关(P < 0.01);SOD仅与丙二醛呈显著正相关(P < 0.05),SOD与相对电导率、脯氨酸、可溶性蛋白无显著相关(P > 0.05)。
表 2 盐碱处理下各生理指标的相关性分析
Table 2. Correlation analysis of various physiological indexes under saline-alkali treatment
处理
Treatment指标
Index相对电导率
Relative conductivity丙二醛
Malondiadehyde脯氨酸
Proline可溶性蛋白
Soluble protein超氧化物歧化酶
SODNaCl 相对电导率 Relative conductivity 1 丙二醛 Malondialdehyde 0.806** 1 脯氨酸 Proline 0.979** 0.760** 1 可溶性蛋白 Soluble protein 0.720** 0.844** 0.754** 1 超氧化物歧化酶 SOD 0.424 0.739** 0.440 0.826** 1 Na2CO3 相对电导率 Relative conductivity 1 丙二醛 Malondialdehyde 0.804** 1 脯氨酸 Proline 0.951** 0.867** 1 可溶性蛋白 Soluble protein 0.828** 0.809** 0.876** 1 超氧化物歧化酶 SOD 0.291 0.553* 0.260 0.446 1 注: **表示极显著相关(P < 0.01),*表示显著相关(P < 0.05)。
Note: **Correlation is significant at the 0.01 level, * correlation is significant at the 0.05 level. -
由表3可看出:NaCl处理下特征值为4.121,累计贡献率为82.423%,综合了丙二醛、可溶性蛋白、相对电导率、脯氨酸和超氧化物歧化酶的信息;Na2CO3处理下特征值为3.886,累计贡献率为77.725%,主要综合了丙二醛、可溶性蛋白、相对电导率、脯氨酸的信息,具有较好的代表性。因此,可用此5个指标评价文冠果耐盐性能力[22]。
表 3 主成分分析旋转后的成分载荷矩阵
Table 3. Principal component analysis rotated component loading matrix
指标
IndexNaCl胁迫
NaCl stress
PC1Na2CO3胁迫
Na2CO3 stress
PC1丙二醛 Malondialdehyde 0.968 0.966 可溶性蛋白 Soluble protein 0.953 0.949 相对电导率 Relative conductivity 0.902 0.930 脯氨酸 Proline 0.896 0.956 超氧化物歧化酶 SOD 0.813 0.000 特征值
Eigen value4.121 3.886 方差贡献率/%
Variance contribution rate82.423 77.725 累计贡献率/%
Cumulative contribution rate82.423 77.725 由表4综合得分[23]可看出:不同浓度NaCl处理对文冠果生理指标的影响由大到小依次为:280、210、140、70、0 mmol·L−1;不同浓度Na2CO3处理对文冠果生理指标的影响由大到小依次为:105、140、70、35、0 mmol·L−1。
表 4 NaCl、Na2CO3胁迫下文冠果的综合得分及排名
Table 4. NaCl、Na2CO3 processing the comprehensive score and ranking of the following
处理
Treatment浓度
Concentration/
(mmol·L−1)PC1
(F1)综合得分
Overall ratings综合得分排名
Overall score
rankingNaCl 0(CK) −1.5154 −124.9038 5 70 −0.4671 −38.4998 4 140 0.3908 32.2109 3 210 0.6143 50.6324 2 280 0.9774 80.5602 1 Na2CO3 0(CK) −1.5215 −118.2586 5 35 −0.4659 −36.2121 4 70 0.4026 31.2921 3 105 0.9508 73.9009 1 140 0.6340 49.2777 2 -
盐胁迫产生离子毒害的同时引起次生作用[24]。在盐碱胁迫下,植物组织离子浓度过高,导致离子失衡,从而引发植物水分缺失[25]。大量Na+的积累会产生离子毒害[26],引发诸如植株生长、光合反应及渗透物质等一系列的生理生化反应,最终表现在植物的生长和生物量累积[5,27]。本研究表明,在盐碱胁迫下,文冠果总生物量呈降低趋势,根冠比呈上升趋势,胁迫后期叶片失水萎蔫,变黄干枯,甚至死亡。这可能是植株在调节生物量的分配模式[28]来应对胁迫环境。相对电导率和丙二醛含量可反映植物细胞膜在逆境条件的受损伤程度[29-30]。本研究中,中性盐70 mmol·L−1处理17、22 d文冠果幼苗的相对电导率略有下降,这可能是低浓度盐对植物的生长具有促进作用[31];碱性盐处理下,相对电导率呈显著升高趋势(图2),说明高浓度碱性盐会破坏文冠果的细胞膜系统,影响文冠果的生长,该结果与齐琪[32]的研究结论基本一致。在盐碱胁迫下,时间和浓度2种影响因子对丙二醛无显著的交互作用(P > 0.05);在盐碱胁迫后期,高浓度时丙二醛含量呈下降趋势,这可能与适度胁迫会产生一系列“防卫性”生理反应有关,该反应阻碍了伤害的加深,因此,丙二醛含量较低[33],但随着胁迫的持续加重,势必引发不可逆的伤害。
脯氨酸、可溶性蛋白的积累是植物在盐碱胁迫下诱发的一种防御行为,也是植物遭受逆境胁迫的信号[34]。随着盐浓度的增大,脯氨酸和可溶性蛋白含量持续上升,这表明植物通过脯氨酸、可溶性蛋白调节细胞渗透势,缓解渗透胁迫对植物生长的抑制作用[35]。在碱胁迫下,脯氨酸含量随碱浓度的增加呈先上升后下降的趋势,说明高浓度的Na2CO3处理会破坏文冠果渗透调节物质的积累。
活性氧(ROS)作为渗透胁迫和离子毒害的副产物,能导致细胞膜功能丧失和细胞的死亡[36]。植物在逆境胁迫下,活性氧代谢加剧体内ROS过度积累[37]。胁迫初期,植物体内的抗氧化酶系统被激活,表现为随处理浓度的增大,文冠果的SOD活性呈先升后降趋势。本研究中,文冠果幼苗在低浓度盐碱胁迫下能够通过提高 SOD 活性增强对活性氧的清除能力;当盐碱含量过高时,SOD活性下降,致使活性氧清除能力受到抑制,这一结果与林雪锋等[38]对海滨植物 SOD 活性的研究结论基本一致。除碱性盐140 mmol·L−1外,随胁迫时间的延长,植物抗氧化酶活性均持续升高。在中性盐处理下,SOD活性升至58 U·g−1·min−1左右保持稳定,而碱性盐SOD活性升至61 U·g−1·min−1左右趋于稳定。碱性盐140 mmol·L−1处理下SOD活性显著下降表明其已达到阈值,植物体的代谢出现紊乱,活性氧清除机制遭到破坏,酶活性降低,SOD不足以清除植物体内产生的活性氧、自由基等[39]。由于华北地区平均土壤含盐量在0.3%左右,最高达到1.39%[40],因此,在园林绿化中,华北平原轻、中度盐渍化土壤可选择文冠果应用推广。
-
综上所述,盐浓度在70、140 mmol·L−1,碱浓度在35、70 mmol·L−1时文冠果通过调节生物量分配模式,积累渗透调节物质和提高抗氧化酶系统来保护细胞膜系统共同抵御盐碱胁迫。在高浓度盐胁迫(210、280 mmol·L−1)、碱胁迫(105、140 mmol·L−1)下,文冠果主要通过积累渗透调节物质来抵御逆境。此外,与中性盐NaCl相比,高浓度碱性盐Na2CO3对文冠果的伤害更严重。双因素方差分析显示:时间和浓度的交互作用对相对电导率、脯氨酸的影响极显著(P < 0.01),且主成分分析和相关性分析表明,丙二醛、可溶性蛋白、相对电导率、脯氨酸、SOD等指标可评价文冠果具有一定的盐碱耐受性。
盐碱胁迫下文冠果生理响应机制研究
Study on Growth and Physiological Response Mechanism of Xanthoceras sorbifolia Bunge under Salt-alkali Stress
-
摘要:
目的 研究文冠果(Xanthoceras sorbifolia Bunge)盐碱条件生理响应机制,为其在盐碱区造林及管理提供科学依据。 方法 以1年生文冠果实生苗为材料,采用盆栽浇盐法,分别设置70、140、210、280 mmol·L−1中性盐(NaCl)及35、70、105、140 mmol·L−1碱性盐(Na2CO3)处理,分析其相对电导率、脯氨酸含量和超氧化物歧化酶(SOD)活性等生理指标。 结果 随盐碱浓度的增大,相对电导率、脯氨酸含量和可溶性蛋白含量均升高,且高浓度胁迫显著高于对照;在NaCl浓度为210 mmol·L−1、Na2CO3浓度为105 mmol·L−1 胁迫12 d后丙二醛含量比CK显著增多,且在NaCl浓度为280 mmol·L−1、Na2CO3浓度为105 mmol·L−1时达到峰值;SOD活性随处理时间延长而升高,但处理浓度增大其呈先升后降规律。双因素方差分析表明: 时间和浓度对2种类型处理下的相对电导率、脯氨酸含量有极显著交互效应(P < 0.01)。相关性分析显示: 盐碱胁迫下各生理指标均存在显著正相关(P < 0.05)。对指标标准化后进行主成分分析,2种盐类型处理分别提取出特征值大于1的1个主成分,NaCl、Na2CO3处理下累计方差贡献率分别为82.423%、77.725%。 结论 文冠果在盐碱胁迫下主要通过大量积累渗透调节物质、增大抗氧化酶活性来保护细胞膜系统的完整性,抵抗盐碱胁迫。在抗氧化酶系统中,低浓度胁迫时SOD活性较高,高浓度胁迫会破坏SOD活性,具有一定阈值。 Abstract:Objective To explore the physiological response mechanism of Xanthoceras sorbifolia Bunge under saline-alkali conditions, so as to provide reference for silviculture in saline-alkali land. Method The relative conductivity, the activity of proline and superoxide dismutase (SOD) were studied to observe the change of physiological indicators. The influences of different concentrations of NaCl (70, 140, 210, 280 mmol·L−1) and Na2CO3 (35, 70, 105, 140 mmol·L−1) on the growth and physiological characteristics of one-year-old X. sorbifolia seedlings were observed by pot experiment. Result With the increase of saline-alkali concentration, the relative conductivity, the contents of proline and soluble protein changed significantly. The experimental data was larger than that of control group under high concentrations stress. Compared with the CK, the content of malondialdehyde increased significantly after 12 days of treatment with 210 mmol·L−1 salt and 105 mmol·L−1 alkali, and reached a peak with the concentrations of NaCl and Na2CO3 were 280 mmol·L−1 and 105mmol·L−1 respectively. Moreover, the SOD activity continued to increase under the stress, while the activity increased and then declined with the strengthen of the treatment concentration. Additionally, Two-way ANOVA showed that the time duration and concentration had extremely significant correlated with relative conductivity and proline content (P < 0.01). Correlation analysis demonstrated that the five physiological indicators measured in the leaves of seedlings under salt-alkali stress had significant positive correlation (P < 0.05). Principal component analysis was also applied after standardization of all index data. A principal component with eigenvalue greater than 1 was extracted under two different salt treatments. The cumulative variance contribution rate under NaCl treatment reached 82.423%, and the cumulative variance contribution rate under Na2CO3 treatment was 77.725%. Conclusion Under salt and alkali stress, X. sorbifolia Bunge mainly protects the integrity of the cell membrane system by accumulating a large amount of osmotic adjustment substances and increasing the activity of antioxidant enzymes. In the antioxidant enzyme system, the activity of SOD is higher under low-concentration stress than that of control group. However, the enzyme activity is destroyed under high concentration stress, which shows an obvious phenomenon of threshold value. -
图 1 盐碱处理对文冠果幼苗总生物量和根冠比的影响
Figure 1. The effect of saline-alkali treatment on biomass production and root-shoot ratio of Xanthoceras sorbifolia seedlings
图 2 盐碱处理对文冠果幼苗叶片相对电导率和丙二醛的影响
Figure 2. The effect of saline-alkali treatment on relative electrical conductivity and Malondialdehyde in the leaves of Xanthoceras sorbifolia seedlings
图 3 盐碱处理对文冠果幼苗叶片脯氨酸和可溶性蛋白含量的影响
Figure 3. The effect of saline-alkali treatment on the content of proline and soluble protein in the leaves of Xanthoceras sorbifolia seelings
图 4 盐碱处理对文冠果幼苗叶片SOD活性的影响
Figure 4. The effect of saline-alkali treatment on SOD activity in the leaves of Xanthoceras sorbifolia seedlings
表 1 盐碱处理下各生理指标的双因素方差分析
Table 1. Two-way ANOVA of various physiological indexes under saline-alkali treatment
处理
Treatment影响因子
Impact factors相对电导率
Relative conductivity丙二醛
Malondiadehyde脯氨酸
Proline可溶性蛋白
Soluble protein超氧化物歧化酶
SODNaCl 时间 Time 1636.66** 14.03** 317.79** 62.39** 3.92* 浓度 Concentration 802.63** 7.82** 169.83** 101.64** 10.72** 时间 × 浓度 Time × concentration 68.42** NS 15.65** 6.15** NS Na2CO3 时间 Time 472.36** 8.81** 64.30** 48.78** NS 浓度 Concentration 5602.78** 24.49** 309.37** 74.78** 53.25** 时间 × 浓度 Time × concentration 48.51** NS 4.52** NS NS 注:*表示差异显著(P < 0.05),**表示差异极显著(P < 0.01),NS表示差异不显著;表中数据代表F值,误差项省略。
Notes:*, **mean significance at 0.05 level and 0.001 level,respectively,NS mean no significance. The data in the table represent the F value, and the error item is omitted. 下载: 导出CSV
下载: 导出CSV
表 2 盐碱处理下各生理指标的相关性分析
Table 2. Correlation analysis of various physiological indexes under saline-alkali treatment
处理
Treatment指标
Index相对电导率
Relative conductivity丙二醛
Malondiadehyde脯氨酸
Proline可溶性蛋白
Soluble protein超氧化物歧化酶
SODNaCl 相对电导率 Relative conductivity 1 丙二醛 Malondialdehyde 0.806** 1 脯氨酸 Proline 0.979** 0.760** 1 可溶性蛋白 Soluble protein 0.720** 0.844** 0.754** 1 超氧化物歧化酶 SOD 0.424 0.739** 0.440 0.826** 1 Na2CO3 相对电导率 Relative conductivity 1 丙二醛 Malondialdehyde 0.804** 1 脯氨酸 Proline 0.951** 0.867** 1 可溶性蛋白 Soluble protein 0.828** 0.809** 0.876** 1 超氧化物歧化酶 SOD 0.291 0.553* 0.260 0.446 1 注: **表示极显著相关(P < 0.01),*表示显著相关(P < 0.05)。
Note: **Correlation is significant at the 0.01 level, * correlation is significant at the 0.05 level.
下载: 导出CSV
表 3 主成分分析旋转后的成分载荷矩阵
Table 3. Principal component analysis rotated component loading matrix
指标
IndexNaCl胁迫
NaCl stress
PC1Na2CO3胁迫
Na2CO3 stress
PC1丙二醛 Malondialdehyde 0.968 0.966 可溶性蛋白 Soluble protein 0.953 0.949 相对电导率 Relative conductivity 0.902 0.930 脯氨酸 Proline 0.896 0.956 超氧化物歧化酶 SOD 0.813 0.000 特征值
Eigen value4.121 3.886 方差贡献率/%
Variance contribution rate82.423 77.725 累计贡献率/%
Cumulative contribution rate82.423 77.725
下载: 导出CSV
表 4 NaCl、Na2CO3胁迫下文冠果的综合得分及排名
Table 4. NaCl、Na2CO3 processing the comprehensive score and ranking of the following
处理
Treatment浓度
Concentration/
(mmol·L−1)PC1
(F1)综合得分
Overall ratings综合得分排名
Overall score
rankingNaCl 0(CK) −1.5154 −124.9038 5 70 −0.4671 −38.4998 4 140 0.3908 32.2109 3 210 0.6143 50.6324 2 280 0.9774 80.5602 1 Na2CO3 0(CK) −1.5215 −118.2586 5 35 −0.4659 −36.2121 4 70 0.4026 31.2921 3 105 0.9508 73.9009 1 140 0.6340 49.2777 2
下载: 导出CSV
-
[1] Zhang X X, Shi Z Q, Tian Y J, et al. Salt stress increases content and size of glutenin macropolymers in wheat grain[J]. Food Chemistry, 2016, 197: 516-521. doi: 10.1016/j.foodchem.2015.11.008 [2] 徐云兵. 盐碱地的改良和应用[J]. 现代园艺, 2016, 39(12):147-147. [3] 张祎曼. 浅析中国西北地区土壤盐碱化现状及修复对策[J]. 当代化工研究, 2018, 18(2):26-27. [4] 孙海菁, 王树凤, 陈益泰. 盐胁迫对6个树种的生长及生理指标的影响[J]. 林业科学研究, 2009, 22(3):315-324. [5] 郭丽丽, 郝立华, 贾慧慧, 等. NaCl胁迫对两种番茄气孔特征、气体交换参数和生物量的影响[J]. 应用生态学报, 2018, 29(12):3949-3958. [6] 鲁 艳, 雷加强, 曾凡江, 等. NaCl胁迫对大果白刺幼苗生长和抗逆生理特性的影响[J]. 应用生态学报, 2014, 25(3):711-717. [7] 毛桂莲, 李国旗, 许 兴, 等. NaHCO3胁迫下3种灌木Na+、K+、Ca2+的吸收及转运[J]. 应用生态学报, 2014, 25(3):718-724. [8] Paz R C, Reinoso H, Espasabdin F D, et al. Akaline, saline and mixed saline-alkaline stresses induce physiological and morpho-anatomical changes in Lotus tenuis shoots[J]. Plant Biol, 2014, 16(6): 1042-1049. [9] 慈敦伟, 张智猛, 丁 红, 等. 花生苗期耐盐性评价及耐盐指标筛选[J]. 生态学报, 2015, 35(3):805-814. [10] 张晓燕. 神东矿区不同种源地文冠果生长适宜性及耐盐性研究[D]. 呼和浩特: 内蒙古农业大学, 2012. [11] 郭 瑞, 李 峰, 周 际, 等. 亚麻响应盐、碱胁迫的生理特征[J]. 植物生态学报, 2016, 40(1):69-79. [12] Munns R. Comparative physiology of salt and water stress[J]. Plant, Cell& Environment, 2002, 25(2): 239-250. [13] Wang L X, Fang C, Wang K. Physiological responses of Leymus chinensis to long-term salt, alkali and mixed salt-alkali stresses[J]. Journal of Plant Nutrition, 2015, 38(4): 526-540. doi: 10.1080/01904167.2014.937874 [14] 王 峰. 生物质能源林的首选树种--文冠果[J]. 甘肃林业, 2011(2):37-38. [15] 李 茂. 山西北中部文冠果生物质能源林建设规划研究[D]. 山西: 山西农业大学, 2014. [16] 侯元凯, 刘松杨, 黄 琳, 等. 我国生物柴油树种选择与评价[J]. 林业科学研究, 2009, 22(1):7-13. [17] 张 丽. 平欧杂种榛抗盐碱生理机制研究及其耐盐性评价[D]. 北京: 中国林业科学研究院, 2015. [18] 刘正祥, 张华新, 杨秀艳, 等. Na2SO4胁迫对沙枣幼苗生长和光合生理的影响[J]. 林业科学研究, 2014, 27(2):186-194. [19] 马 剑, 刘贤德, 张芬琴, 等. NaCl胁迫对文冠果生长及生理生化指标的影响[J]. 干旱区资源与环境, 2018, 32(2):182-187. [20] 王学奎. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2006. [21] 高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006. [22] 李红峥, 曹红霞, 郭莉杰, 等. 沟灌方式和灌水量对温室番茄综合品质与产量的影响[J]. 中国农业科学, 2016, 49(21):4179-4191. [23] 张会慧, 张秀丽, 李 鑫, 等. NaCl和Na2CO3胁迫对桑树幼苗生长和光合特性的影响[J]. 应用生态学报, 2012, 23(3):625-631. [24] 赵 婷, 张 瑞, 吴玉霞, 等. 外源H2S处理对盐碱胁迫下垂丝海棠幼苗生理特性的影响[J]. 果树学报, 2020, 37(8):66-71. [25] 郝 汉, 曹 磊, 陈伟楠, 等. 盐胁迫对槲树(Quercus dentata)幼苗离子平衡及其生理生化特性的影响[J]. 生态学报, 2020, 40(19):6897-6904. [26] Saqib M, Akhar J, Qureshi R H. Na+ exclusion and salt resistance of wheat (Triticum aestivum) in saline-waterlogged conditions are improved by the development of adventitious nodal roots and cortical root aerenchyma[J]. Plant Science, 2005, 169: 125-130. doi: 10.1016/j.plantsci.2005.03.003 [27] 甘红豪, 赵 帅, 杨泽坤, 等. 刺槐幼苗对NaCl胁迫的生理生化响应[J]. 林业科学研究, 2020, 33(4):75-82. [28] Van Zandt P A, Tobler M A, Mouton E, et al. Positive and negative consequences of salinity stress for the growth and reproduction of the clonal plant, Iris hexagona[J]. Journal of Ecology, 2003, 91(5): 837-846. doi: 10.1046/j.1365-2745.2003.00806.x [29] 朱燕芳, 王延秀, 胡 亚, 等. 多效唑对水分胁迫下苹果砧木八棱海棠光合及抗氧化酶活性等生理特性的影响[J]. 干旱地区农业研究, 2018, 36(4):178-186. [30] Ehlert B, Hincha D K. Chlorophyll fluorescence imaging accurately quantifies freezing damage and cold acclimation responses in Arabidopsis leaves[J]. Plant Methods, 2008, 4(1): 1-7. doi: 10.1186/1746-4811-4-1 [31] 李志萍, 张文辉, 崔豫川. NaCl和Na2CO3 胁迫对栓皮栎种子萌发及幼苗生长的影响[J]. 生态学报, 2015, 35(3):742-751. [32] 齐 琪. 蒙古柳雌雄株扦插苗对盐碱胁迫的响应[D]. 哈尔滨: 东北林业大学, 2019. [33] 朱志梅, 杨 持. 沙漠化过程中四个共有种的生长和抗氧化系统酶类变化[J]. 应用生态学报, 2004, 15(12):2261-2266. [34] 刘 铎, 丛日春, 党宏忠, 等. 柳树幼苗渗透调节物质对中碱性钠盐响应的差异性[J]. 生态环境学报, 2014, 23(9):1531-1535. [35] 程淑娟, 钱虹妹, 唐东芹, 等. NaCl胁迫对4种园林植物生长及渗透调节物质的影响[J]. 上海交通大学学报, 2012, 29(2):88-94. [36] Bohnert H J, Jensen R G. Strategies for engineering water-stress tolerance in plants[J]. Trends in Biotechnology, 1996, 14(3): 89-97. doi: 10.1016/0167-7799(96)80929-2 [37] 司廉邦, 李嘉敏, 黎桂英, 等. 茶多酚对盐胁迫下小麦幼苗叶片生理特性的影响[J]. 生态学报, 2020, 40(11):3747-3755. [38] 林雪锋, 颉洪涛, 虞木奎, 等. 盐胁迫下3种海滨植物形态和生理响应特征及耐盐性差异[J]. 林业科学研究, 2018, 31(3):95-103. [39] 林 龙, 罗佳佳, 刘酉琳, 等. 水盐胁迫对榕树的生理生化影响的研究[J]. 西南林业大学学报: 自然科学版, 2020, 40(1):46-51. [40] 周在明, 赵淑惠. 华北半干旱平原区表层土壤盐分累积的影响因素分析[J]. 干旱区地理, 2015, 38(5):976-984. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5087
- HTML全文浏览量: 3383
- PDF下载量: 70
- 被引次数: 0
