


-
梵净山地处热带、亚热带生物区系向温带生物区系的生态交错区,由于独特的地理位置、复杂的生物和生态演化过程、多样化的植物类群特征、众多地方特有和濒危动植物种类等特点[1-2],引起国内外不同领域学者的广泛关注,相关研究涉及到土壤理化性质、物种种类及多样性、珍稀濒危物种种群结构特征[2-5]等多个方面。关于地下微生物群落的研究则集中在丛枝菌根真菌[6-7],对真菌群落结构及功能类群特征尚无系统报道。真菌特别是某些功能类群在促进有机质分解和养分循环、增强植物对矿物质营养元素的吸收、提高植物对生物和非生物胁迫的耐受性、维持生态系统的稳定性和生产力等方面发挥着重要作用[8-10]。此外,真菌群落组成不仅受土壤理化性质、气候等非生物因素影响[11-12],其对植物种类、群落组成和多样性等[13-14]生物因素也较为敏感,而植被类型可通过影响非生物和生物因素作用于真菌群落。尤为特殊的是,不同植被类型还会塑造特定的真菌功能类群:Tedersoo等[11]通过对全世界范围内不同植被类型的土壤真菌群落进行分析后发现,外生菌根真菌在温带落叶林中较为丰富,热带潮湿雨林和热带干旱森林中植物病原菌更为多样;Liu等[15]发现在中国东部森林地区,温带森林以外生菌根真菌为主,热带和亚热带森林中具有丰富的丛枝菌根真菌和植物病原菌;Sheng等[16]研究了神农架山地森林生态系统中的土壤真菌功能类群,发现外生菌根真菌在落叶阔叶林、针阔混交林和针叶林中尤为丰富,植物病原菌-木腐生菌主要出现在常绿阔叶林,植物病原菌和寄生性真菌在山顶灌丛中具有较高相对丰度。菌在山顶灌丛中具有较高相对丰度。
梵净山拥有的典型植物区系过渡性及地球上同纬度保存完好的森林生态系统,为丰富且独特真菌种类的形成创造良好生存条件。此外,由于水热条件等差异造就了梵净山多样且分异明显的植被类型,为探讨植被类型与真菌群落间的关系提供了天然理想实验室。本研究以梵净山国家级自然保护区典型且具有代表性的常绿阔叶林、常绿落叶阔叶混交林及位于山顶附近的亚高山矮林3种森林植被类型为研究对象,分析真菌群落结构和功能类群特征与植被类型间的关系,探讨土壤性质、细根性状和叶性状对真菌群落的影响。研究结果可为后续探讨亚热带地区不同植被类型树种的环境适应机制及地上和地下部分的养分交换策略提供来自于地下真菌群落方面的数据支撑。
-
研究区域位于梵净山国家级自然保护区(27°46′50″ N ~ 28°1′30″ N,108°35′55″ E ~ 108°48′30″ E)内,地处贵州铜仁市江口县、印江自治县、松桃自治县交界处,面积约为419 km2。亚热带季风湿润气候,年平均气温16 ℃,年平均降雨量1 300 mm,平均相对湿度80%以上,土壤以山地黄壤和黄棕壤为主。植被类型丰富,具有代表性和面积较大的是常绿阔叶林和常绿落叶阔叶混交林,前者主要植物种类为栲(Castanopsis fargesii Franch.)、米槠(C. carlesii (Hemsl.) Hayata.)、巴东栎(Quercus engleriana Seem.)、山茶(Camellia japonica L.)等,后者植物种类包括水青冈(Fagus longipetiolata Seem.)、贵州青冈(Cyclobalanopsis argyrotricha (A. Camus) Chun et Y. T. Chang ex Y. C. Hsu et H. W. Jen)和扇叶槭(Acer flabellatum Rehd.)等。亚高山矮林是位于山顶附近的特殊植被类型,以吊钟花(Enkianthus quinqueflorus Lour.)、耳叶杜鹃(Rhododendron auriculatum Hemsl.)和长萼杜鹃(Rhododendron longicalyx Fang f.)等杜鹃花科植物为主。
在常绿阔叶林海拔600 m处、常绿落叶阔叶混交林海拔1 500 m处和亚高山矮林海拔2 200 m处分别垂直于海拔高度设置1条100 m的样方线,每隔10 m设置一个采样点,中心象限法[17]采集样本,即在每个采样点设置4个方向,选择距离采样点最近的2个树种(起测胸径为2 cm),去除表层石块和落叶等杂物,从树干开始挖掘完整根系,抖落法[18]收集根际土壤。由于部分采样点树木生长在坡度较大区域或岩石缝隙中,采集根系和土壤极为困难,仅48个样本用于后续分析(每种植被类型16个)。其中,常绿阔叶林中,栲5个,巴东栎4个,山茶3个,黄丹木姜子(Litsea elongata (Wall. ex Nees) Benth. et Hook. f.)2个,四照花(Cornus kousa subsp. chinensis (Osborn) Q. Y. Xiang)1个,柃木(Eurya japonica Thunb.)1个;常绿落叶阔叶混交林中,贵州青冈5个,水青冈4个,多脉青冈(Cyclobalanopsis multinervis W.C.Cheng & T.Hong)3个,云山青冈(C. sessilifolia (Blume) Schottky)3个,青冈(C. glauca (Thunberg) Oersted)1个;亚高山矮林中,耳叶杜鹃8个,吊钟花6个,长萼杜鹃2个。同时,在每个植株上随机摘取20片向阳、完全展开、没有病虫害的叶片,用于叶性状指标分析。根系样本装在放有干冰的泡沫箱运回实验室,一部分保存在4 ℃冰箱用于分析根性状指标,另一部分保存在−70 ℃冰箱用于真菌测定。
-
根系用自来水清洗掉附着土壤后,依据Pregitzer等[19]根序分级法,按照序级进行分级,前两个序级归为吸收根并进行分离。EPSON Perfection V700(EPSON America Inc.)扫描细根样本,根系图像分析软件WinRhizo(Régent Instruments Inc., Québec, Canada)分析扫描的图片获得平均根直径/mm、根长/cm、根表面积/cm2和根体积/cm3。将扫描过的细根称量质量后,放入信封袋在70 ℃下烘48 h,再次称量质量得到根生物量/g。计算比根长(根长/生物量)/(cm·g−1)、根比表面积(根表面积/生物量)/(cm2·g−1)、根组织密度(生物量/根体积)/(g·cm−3)和根干物质含量(根干质量/根鲜质量)/(g·g−1)。
叶片放平整后利用Nikon Coolpix 5000数码相机(Nikon Corp.,Tokyo,Japan)拍照,Image J(Systat Software Inc.,Richmond,CA)软件计算叶面积/cm2。称重后,在70 °C下烘72 h,再次称量质量得到叶生物量/g。计算比叶面积(叶面积/叶生物量)/(cm2·g−1)和叶干物质含量(叶干质量/叶鲜质量)/(g·g−1)。
所有土壤、细根和叶样本均重复测定3次。土壤pH测定采用pH计法(土水比例1:2.5);土壤、细根和叶碳、氮含量测定利用CN元素分析仪(Vario EL III,Hanau,Germany);土壤全磷测定采用HCIO4-H2SO4消煮-钼锑抗比色法,细根和叶磷含量测定采用H2SO4-H2O2消煮-钼锑抗比色法[20]。
-
根系用蒸馏水冲洗10 min后,70%乙醇浸泡1 min,3.3%次氯酸钠浸泡3 min,75%乙醇浸泡30 s,无菌水反复冲洗后验证消毒效果。灭菌后根系用液氮冷冻研磨,DNeasy Plant Mini Kit(Qiagen Gmbh, Hilden, Germany)提取DNA,0.8%琼脂糖凝胶电泳检测DNA 提取质量,紫外分光光度计(Thermo Fisher Scientific, MA, USA)准确定量DNA浓度。真菌rDNA-ITS保守区引物ITS1FI2(5’-GTGARTCATCGAATCTTTG-3’)和ITS2 (5’-TCCTCCGCTTATTGATATGC-3’)进行 PCR 扩增。扩增体系包括12.5 μL Phusion® Hot Start Flex 2X Master Mix(New England Biolabs, Ipswich, USA)、10 μmol·L−1正向引物和反向引物各2.5 μL、Genomic DNA 50 ng,补充灭菌超纯水到25 μL。扩增条件:98 ℃预变性30 s;98 ℃变性10 s,54 ℃退火30 s、72 ℃延伸45 s,共 35个循环;72 ℃延伸10 min。扩增产物利用2%琼脂糖凝胶电泳检测,AxyPrepPCR Cleanup Kit(Axygen Biosciences, CA, USA)纯化,Quant-iTTM PicoGreenTM dsDNA Assay Kit(Thermo Fisher Scientific, MA, USA)定量浓度,Illumina MiSeq平台测序。
-
DADA2(divisive amplicon denoising algorithm)[21]进行序列拼接、去除错误碱基序列、去掉barcode和引物序列、过滤嵌合体后,Ribosomal Database Project (RDP) Naïve Bayesian Classifier得到扩增序列变异(amplicon sequence variants,ASVs)。ASVs相当于以100%相似度聚类OTUs(operational taxonomic units),但与OTUs比,具有较高数据精确度与物种分辨率[21]。在UNITE数据库[22]中比对,得到序列注释信息,FUNGuild[23]比对和解析真菌营养型和功能类群,置信度为“极可能”(highly probable)和“很可能”(probable)等级的功能分组被用于后续分析。
-
混合效应模型探讨植被类型对土壤理化性质、细根性状、叶性状、和真菌α多样性指数的影响,植被类型为固定因子,树种为随机因子,并检验模型残差的正态性、齐性和独立性,若不满足则对数据进行转换。LEfSe(
http://huttenhower.sph.harvard.edu/lefse/ )判别不同植被类型间真菌的显著差异类群。非度量多维标度(nonmetric multidimensional scaling,NMDS)排序和置换多元方差分析(permutational multivariate analysis of variance,PERMANOVA)对不同植被类型间真菌群落组成进行比较,Kruskal-Wallis test和Wilcoxon signed-rank tests判别植被类型间真菌Bray-Curtis相异性指数的高低。植被类型、树种、细根性状、叶性状和土壤性质对真菌α多样性指数影响的相对重要性基于ABT(aggregated boosted tree)分析,并计算其与真菌α多样性指数的Pearson相关性。降趋势对应分析(detrended correspondence analysis,DCA)检验真菌数据为单峰分布还是线性分布,DCA排序轴梯度长度最大值为2.657,小于3.0,选择线性排序模型中的冗余分析(redundancy analysis,RDA)探讨影响真菌群落的因子;执行RDA前,对所涉及解释变量进行前向选择。方差分解分析评估植被类型、土壤性质、细根性状和叶性状对真菌群落影响的大小。除特别说明外,统计分析均在R中完成。 -
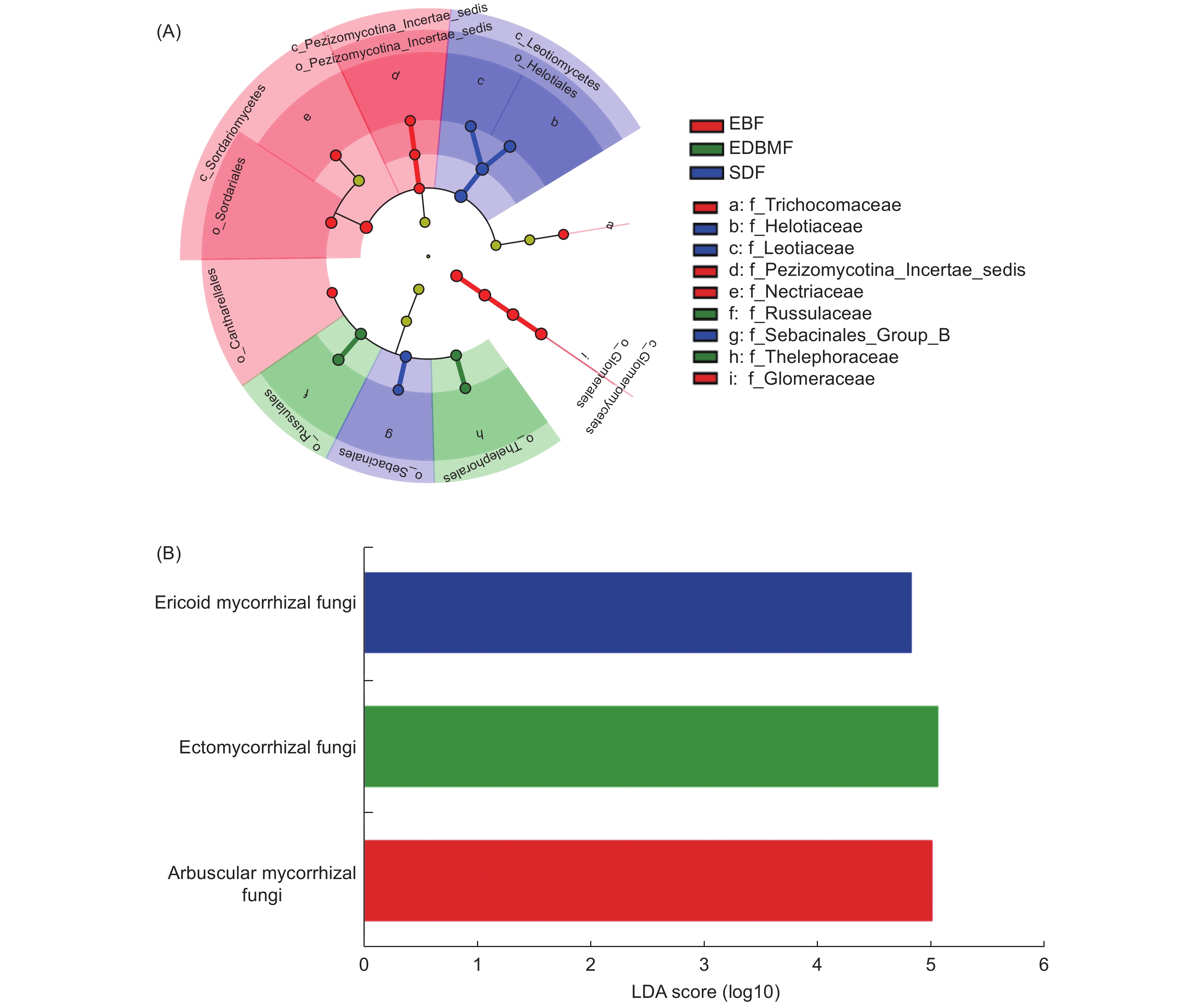
48个根系样本共得到5 103个ASVs,属于9门、193科和385个属(不包括unidentified fungi)。子囊菌门(Ascomycota)和担子菌门(Basidiomycota)为优势门,占全部ASVs数的89.93%。小蔓毛壳科(Herpotrichiellaceae)、球囊霉科(Glomeraceae)和柔膜菌科(Helotiaceae)为优势科,晶杯菌科(Hyaloscyphaceae)、Sebacinales_Group_A、被孢霉科(Mortierellaceae)、皮盘菌科(Dermateaceae)、红菇科(Russulaceae)、小菇科(Mycenaceae)、丛赤壳科(Nectriaceae)、水盘菌科(Vibrisseaceae)、锤舌菌科(Leotiaceae)、肉座菌科(Hypocreaceae)和革菌科(Thelephoraceae)相对丰度超过1%。常绿阔叶林具有最多ASVs数(2 287个),子囊菌门为优势门,球囊霉科为优势科;亚高山矮林共得到ASVs数为1 636个,子囊菌门为优势门,柔膜菌科和晶杯菌科为优势科;常绿落叶阔叶混交林ASVs数最少,为1 608个,子囊菌门和担子菌门为优势门,球囊霉科和小蔓毛壳科为优势科。LEfSe分析(图1A)显示,常绿阔叶林显著差异类群为球囊菌门(Glomeromycota)、球囊菌纲(Glomeromycetes)、粪壳菌纲(Sordariomycetes)、Pezizomycotina_Incertae_sedis纲、Pezizomycotina_Incertae_sedis目、粪壳菌目(Sordariales)、鸡油菌目(Cantharellales)、球囊霉目(Glomerales)、球囊霉科、丛赤壳科、Pezizomycotina_Incertae_sedis科和发菌科(Trichocomaceae),常绿落叶阔叶混交林中显著差异类群为革菌目(Thelephorales)、红菇目(Russulales)、革菌科和红菇科,锤舌菌纲(Leotiomycetes)、柔膜菌目(Helotiales)、蜡壳耳目(Sebacinales)、Sebacinales_Group_B科、柔膜菌科和锤舌菌科是亚高山矮林的显著差异类群。
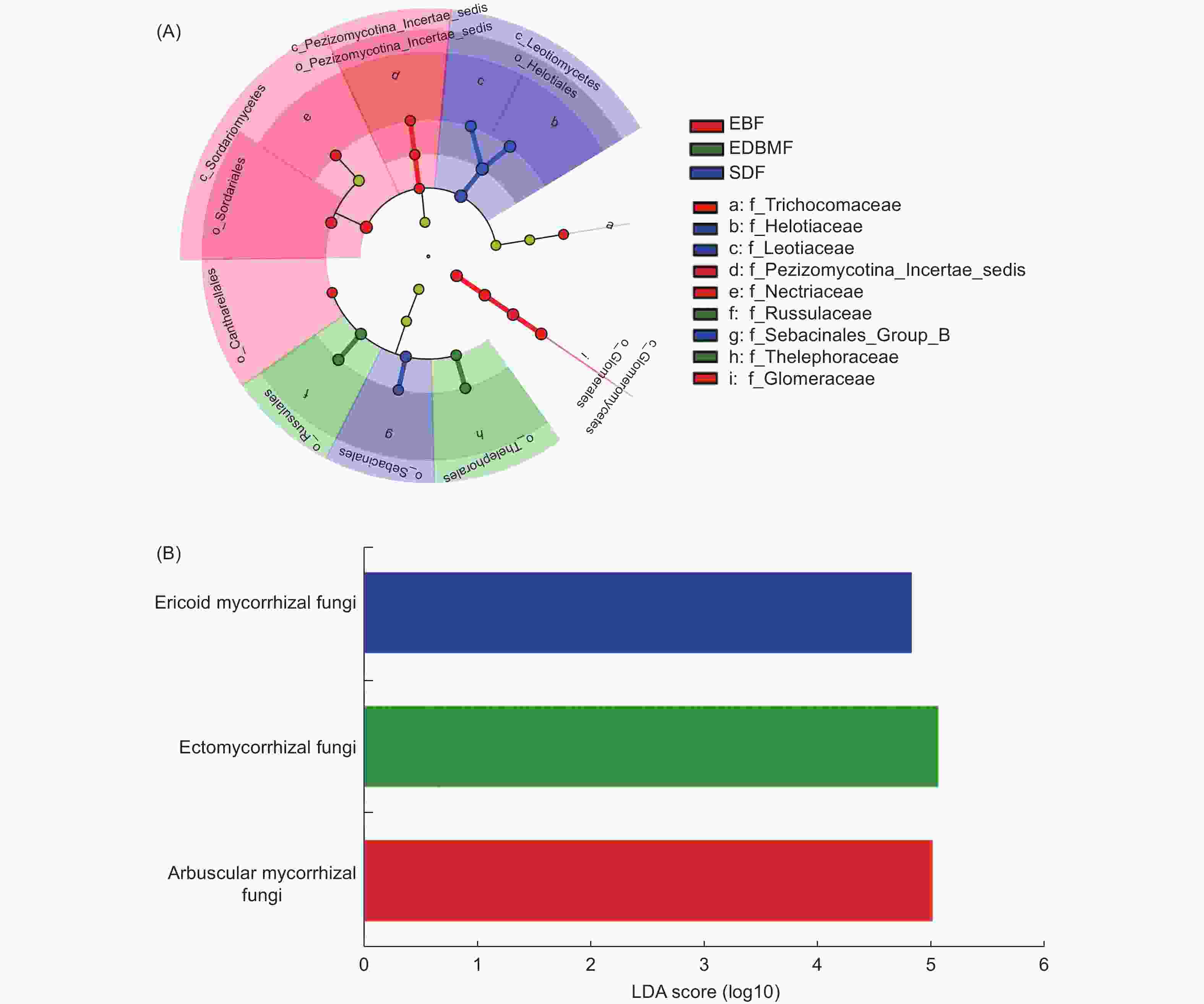
图 1 3种植被类型根系真菌的cladogram图(A)和功能类群的LDA Score图(B)(LDA阈值3.0)
Figure 1. The key root-associated fungal groups (A) and functional taxa (B) corresponding to vegetation type are identified by linear discriminant analysis (LDA) effect size (LEfSe)(LDA score= 3)
-
3种植被类型树种根系真菌在“极可能”和“很可能”水平上进行功能分组的ASVs数为2 231个,共被分为病理营养型(pathotroph)、共生营养型(symbiotroph)、腐生营养型(saprotroph)、病理-共生营养型(pathotroph-symbiotroph)和病理-腐生营养型(pathotroph-saprotroph)5个营养型。其中,41.86%的ASVs被归为腐生营养型,其次是共生营养型(28.73%)和病理营养型(15.37%),其余2个营养型所占比例较低。在功能亚类水平上,undefined saprotroph相对丰度最高,其次是丛枝菌根(arbuscular mycorrhizal)真菌、外生菌根(ectomycorrhizal)真菌、植物病原菌(plant pathogen)、内生真菌(endophyte)、寄生性真菌(fungal parasite)和杜鹃花类菌根(ericoid mycorrhizal)真菌,其余功能亚类相对丰度较低。此外,丛枝菌根真菌仅出现在常绿阔叶林和常绿落叶阔叶混交林,且在常绿阔叶林(7.77%)的相对丰度高于常绿落叶阔叶混交林(7.30%);外生菌根真菌在常绿落叶阔叶混交林(10.20%)的相对丰度明显高于常绿阔叶林(3.28%)和亚高山矮林(0.98%),内生真菌和杜鹃花类菌根真菌只出现在亚高山矮林。LEfSe分析(图1B)还发现,丛枝菌根真菌是常绿阔叶林的显著差异功能类群,外生菌根真菌是常绿落叶阔叶混交林的显著差异功能类群,亚高山矮林的显著差异功能类群为杜鹃花类菌根真菌。
-
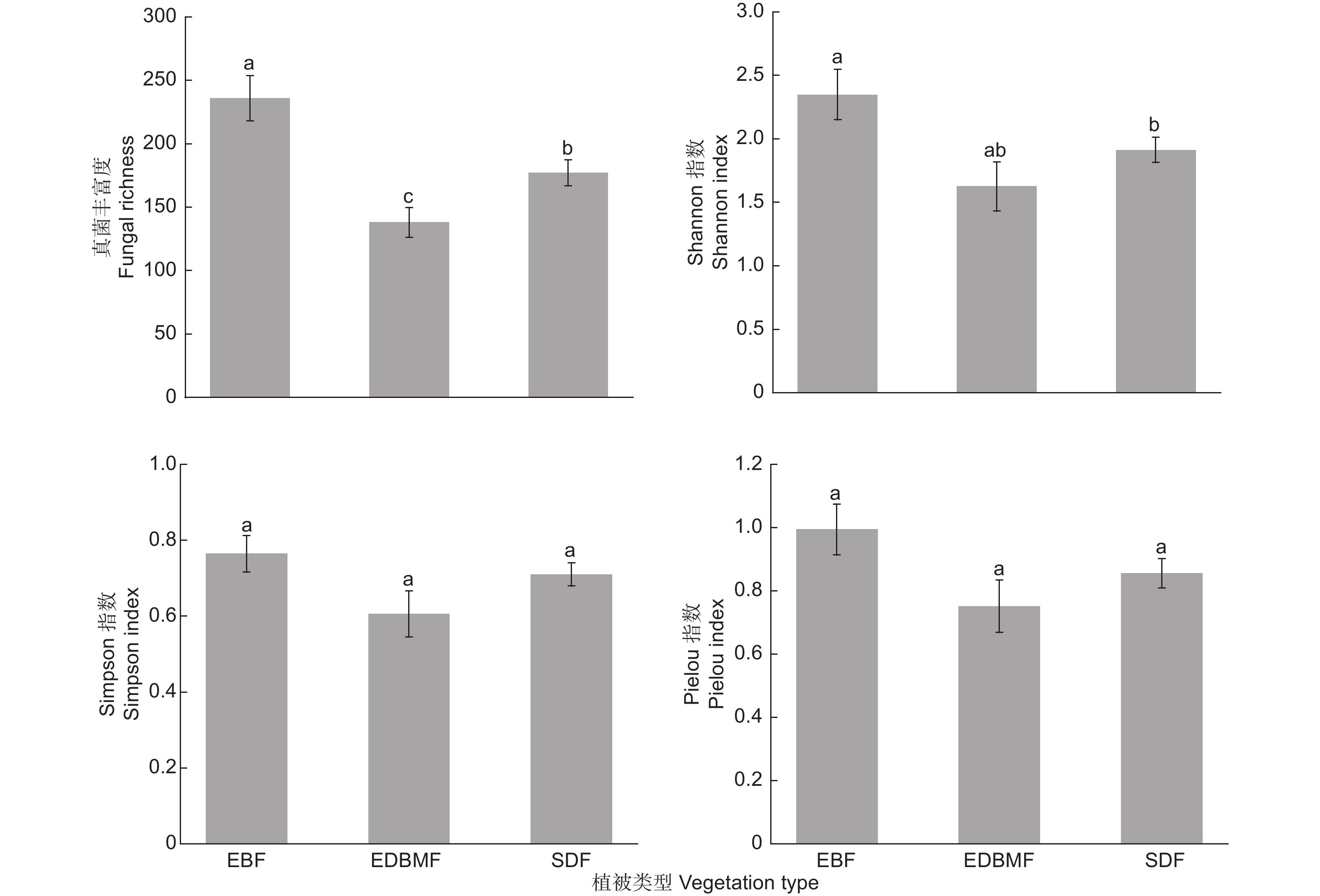
混合效应模型结果表明,植被类型能显著影响真菌丰富度和Shannon指数(P < 0.05),对Simpson指数和Pielou指数无显著影响(P > 0.05),常绿落叶林中真菌丰富度显著高于常绿落叶阔叶混交林和亚高山矮林,Shannon指数显著高于亚高山矮林(图2)。
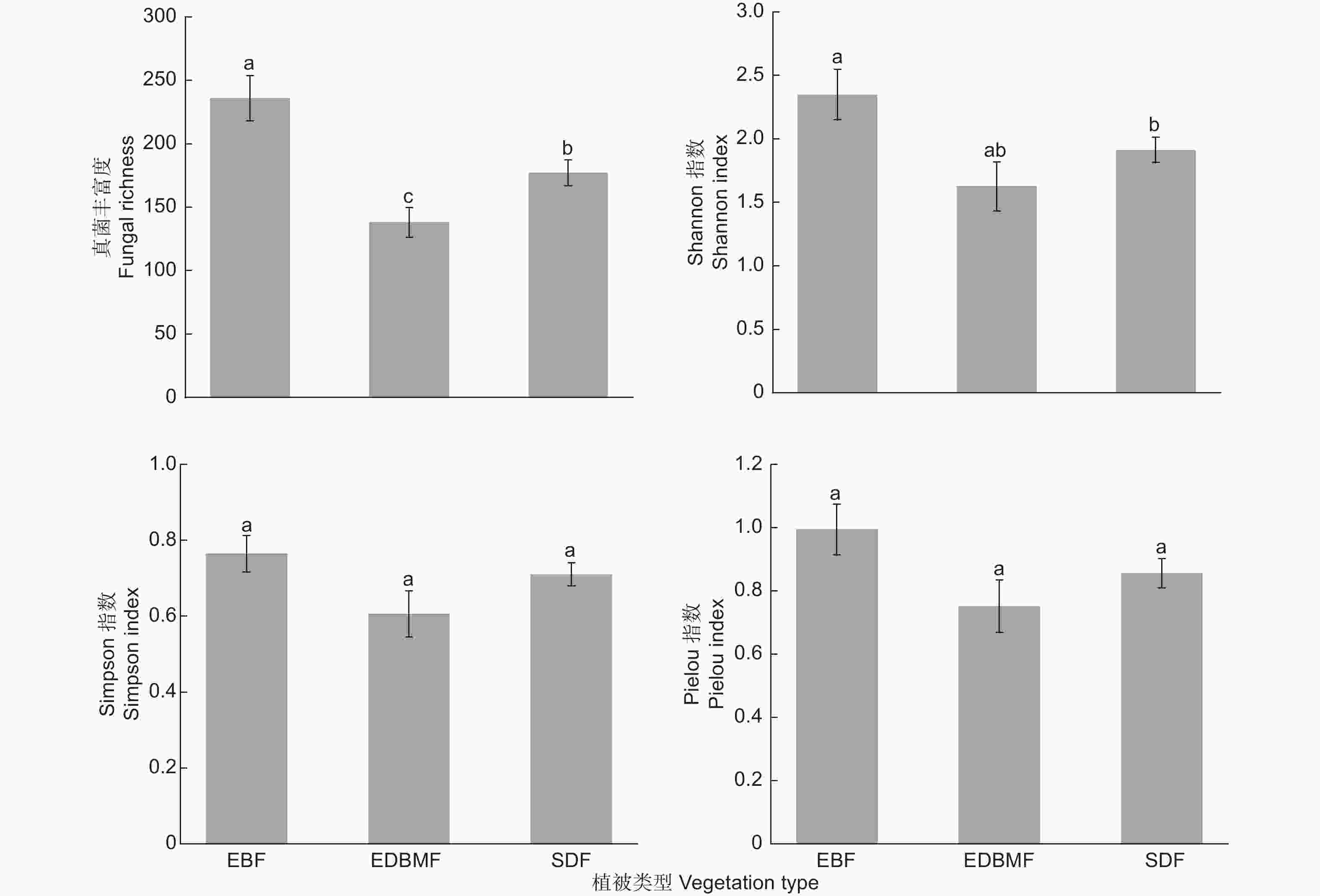
图 2 不同植被类型根系真菌多样性指数
Figure 2. The diversity indices of root-associated fungi in different vegetation types
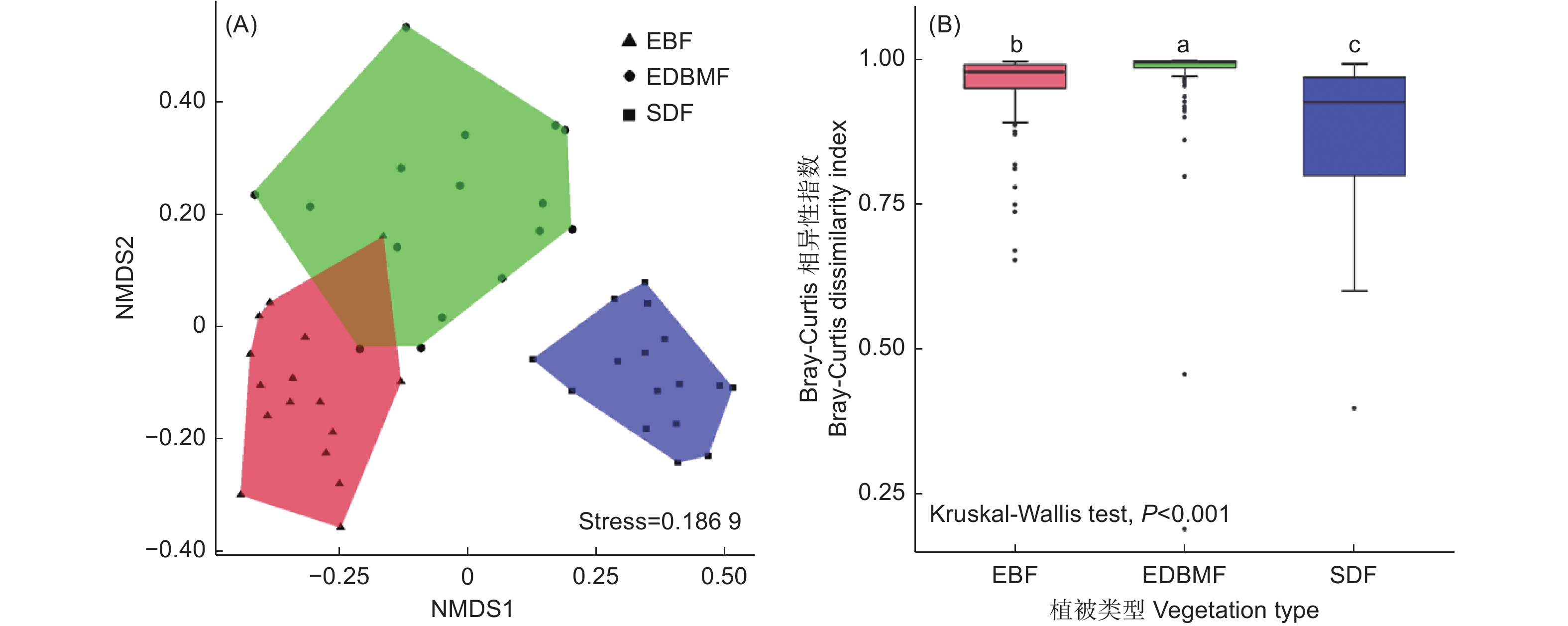
NMDS分析结果显示真菌群落组成在植被类型间存在差异(图3A),PERMANOVA揭示了相同的结果(F = 2.856,P = 0.001)。此外,Bray-Curtis相异性指数(图3B)在3种植被类型间存在极显著差异(P < 0.001),且在常绿落叶阔叶混交林中最高,常绿阔叶林次之,亚高山矮林最低,表明真菌群落组成在亚高山矮林样本间具有最高相似性,其次是常绿阔叶林,常绿落叶阔叶混交林的相似性最低,NMDS分析也印证了该结果。
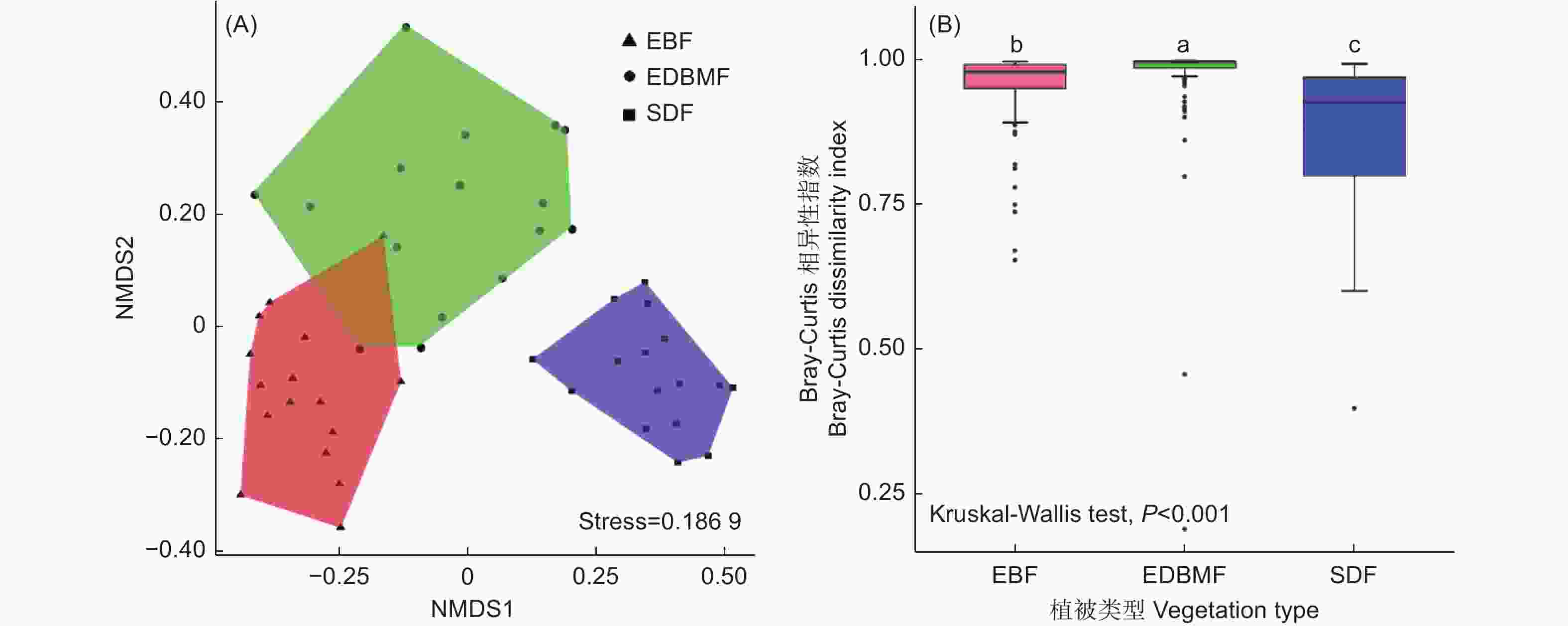
图 3 根系真菌群落的NMDS(A)和Bray-Curtis相异性指数(B)
Figure 3. Nonmetric multidimensional scaling (NMDS) plot (A) and Bray-Curtis dissimilarity index (B) of root-associated fungal community composition based on Bray-Curtis distance
-
以植被类型为固定因子,树种为随机因子构建的混合效应模型发现,植被类型可显著影响土壤pH、土壤全碳、土壤全氮、土壤全磷、比根长、根组织密度、平均根直径、根比表面积、根碳含量、根氮含量、叶面积和叶碳含量(P<0.05)。此外,常绿阔叶林和常绿落叶阔叶混交林土壤pH显著高于亚高山矮林,亚高山矮林土壤具有最高的全碳和全氮含量,常绿阔叶林具有最高的土壤全磷含量;比根长、根比表面积和根碳含量在亚高山矮林显著高于常绿阔叶林和常绿落叶阔叶混交林,常绿阔叶林具有最高的根组织密度、平均根直径和根氮含量;叶面积在常绿阔叶林和常绿落叶阔叶混交林显著高于亚高山矮林,叶碳含量在亚高山矮林显著高于常绿阔叶林和常绿落叶阔叶混交林(表1)。
表 1 不同植被类型土壤理化性质、细根性状和叶性状
Table 1. Soil properties, fine root traits and leaf traits in different vegetation types
指标
Index常绿阔叶林
EBF常绿落叶阔叶混交林
EDBMF亚高山矮林
SDF土壤 pH
Soil pH6.067 ± 0.101 a 5.858 ± 0.156 a 5.006 ± 0.106 b 土壤全碳
Soil total carbon/(g·kg−1)7.057 ± 0.312 b 12.315 ± 1.677 ab 17.598 ± 2.946 a 土壤全氮
Soil total nitrogen/(g·kg−1)0.560 ± 0.022 b 0.875 ± 0.091 ab 1.127 ± 0.166 a 土壤全磷
Soil total phosphorus/(g·kg−1)6.067 ± 0.015a 5.858 ± 0.054 b 5.006 ± 0.053 ab 根生物量
Root biomass/g0.321 ± 0.063 a 0.239 ± 0.057 a 0.246 ± 0.019 a 根干物质含量
Root dry matter content/(g·g−1)0.255 ± 0.041 a 0.274 ± 0.030 a 0.277 ± 0.009 a 比根长
Specific root length/(cm·g−1)29.242 ± 3.209 b 44.158 ± 4.993 b 71.392 ± 10.885 a 根组织密度
Root tissue density/(g·cm−3)1.115 ± 0.249 a 1.014 ± 0.120 a 0.468 ± 0.052 b 平均根直径
Mean root diameter/cm0.237 ± 0.008 a 0.196 ± 0.014 b 0.220 ± 0.009 ab 根比表面积
Specific surface area/(cm2·g−1)217.188 ± 25.446 b 253.534 ± 28.078 b 479.641 ± 29.043 a 根碳含量
Root carbon content/(g·kg−1)46.115 ± 0.423 b 47.918 ± 0.756 b 50.371 ± 0.217 a 根氮含量
Root nitrogen content/(g·kg−1)2.357 ± 0.108 a 2.134 ± 0.128 ab 1.833 ± 0.050 b 根磷含量
Root phosphorus content/(g·kg−1)0.554 ± 0.015 a 0.673 ± 0.073 a 0.730 ± 0.049 a 叶生物量
Leaf biomass/g1.741 ± 0.222 a 1.609 ± 0.198 a 1.536 ± 0.172 a 叶面积
Leaf area/cm2278.285 ± 24.778 a 299.781 ± 26.502 a 167.156 ± 16.237 b 比叶面积
Specific leaf area/(cm2·g−1)169.114 ± 14.169 a 172.458 ± 19.323 a 138.494 ± 24.582 a 叶干物质含量
Leaf dry matter content/(g·g−1)0.386 ± 0.012 a 0.439 ± 0.017 a 0.370 ± 0.028 a 叶碳含量
Leaf carbon content/(g·kg−1)47.592 ± 0.715 b 47.937 ± 0.837 b 50.751 ± 0.456 a 叶氮含量
Leaf nitrogen content/(g·kg−1)1.902 ± 0.126 a 2.052 ± 0.110 a 1.722 ± 0.131 a 叶磷含量
Leaf phosphorus content/(g·kg−1)0.547 ± 0.039 a 0.701 ± 0.063 a 0.615 ± 0.077 a 注:数据为平均值 ± 标准误,同行后不同小写字母表示在P < 0.05水平上差异显著。
Notes: Data are means ± standard error. Different lowercase letters in the same row indicate significant difference at P < 0.05 level.ABT分析用来揭示植被类型、树种、土壤理化性质、细根性状和叶性状对真菌丰富度和多样性指数影响的相对重要性(图4)。结果表明:根碳含量是影响真菌丰富度的首要因子,相对重要性可达到24.47%(图4A);影响Shannon指数、Simpson指数和Pielou指数的首要因子均为叶干物质含量,相对重要性分别为14.63%、18.08%和15.55%(图4B、图4C、图4D)。Pearson相关性分析结果指出,根碳含量与真菌丰富度(r = −0.626,P < 0.001)、Shannon指数(r = −0.407,P = 0.004)呈显著负相关,叶干物质含量与Shannon指数(r = −0.293,P = 0.043)、Simpson指数(r = −0.591,P = 0.002)、Pielou指数(r = −0.531,P = 0.0259)呈显著负相关。
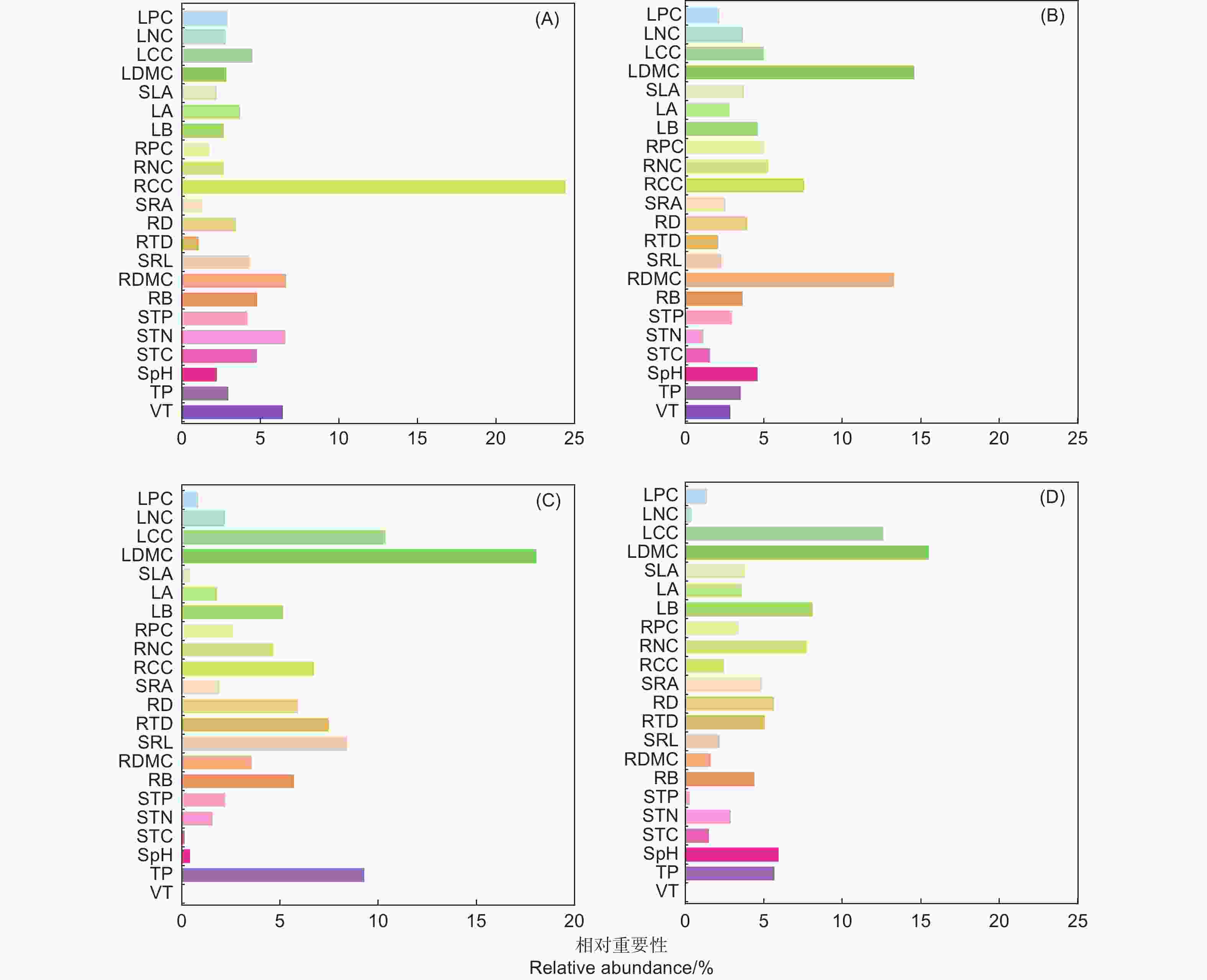
图 4 植被类型、树种、土壤性质、细根性状和叶性状对根系真菌丰富度(A)、Shannon指数(B)、Simpson指数(C)和Pielou指数(D)影响的相对重要性分析
Figure 4. Relative influences of vegetation type, tree species, soil properties, fine root traits and leaf traits on root-associated fungal richness (A), Shannon index (B), Simpson index (C) and Pielou index (D)
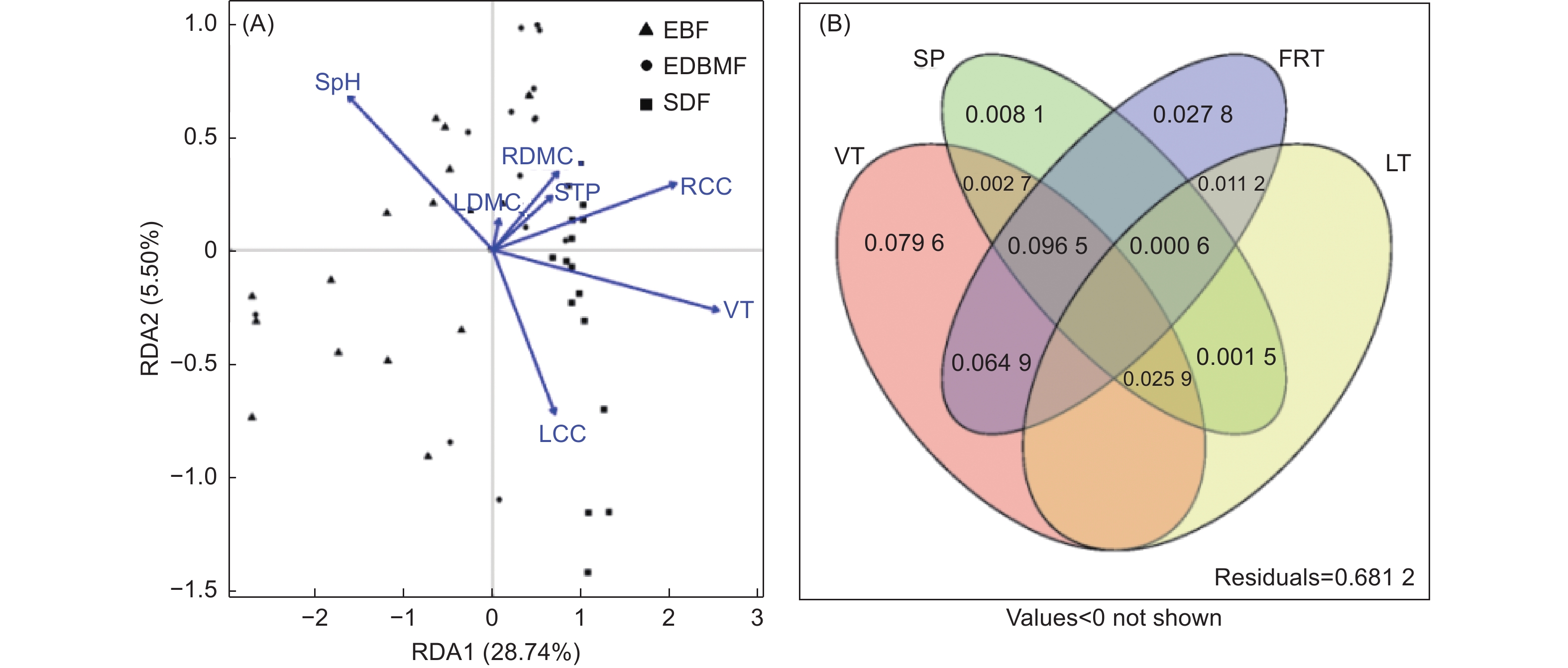
对包括植被类型和树种在内的22个解释变量进行前向选择后剩余7个,分别是植被类型、土壤pH、土壤全磷、根干物质含量、根碳含量、叶干物质含量和叶碳含量,与真菌群落构建RDA模型(图5A)。结果发现,植被类型、土壤pH、根碳含量和叶碳含量是影响真菌群落的显著因子(P < 0.05)。方差分解分析进一步揭示植被类型、土壤性质(土壤pH、土壤全磷)、细根性状(根干物质含量、根碳含量)和叶性状(叶干物质含量、叶碳含量)对真菌群落的影响(图5B),发现植被类型(26.96%)是影响真菌群落的首要因子,其次是细根性状(20.10%)和土壤性质(13.53%),叶性状(3.92%)影响最小。
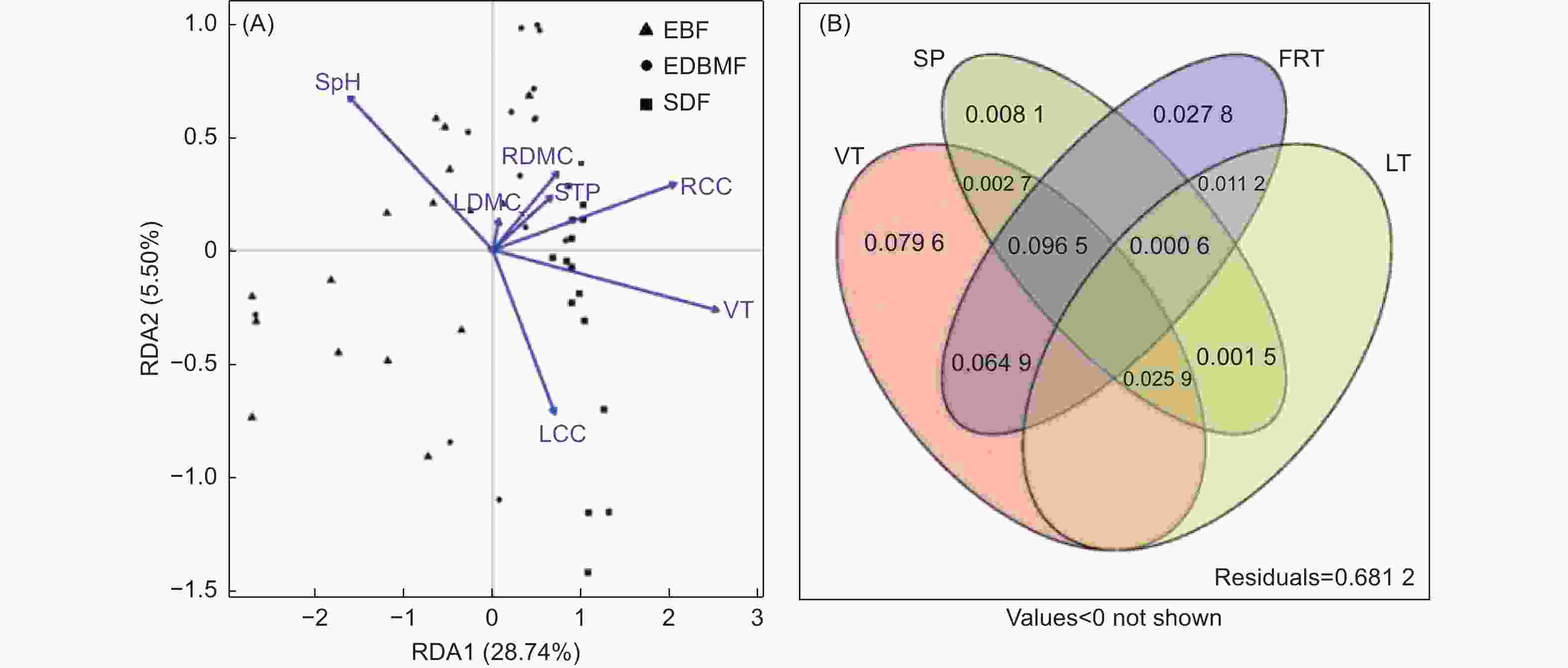
图 5 植被类型、树种、土壤性质、细根性状和叶性状与根系真菌群落关系的RDA(A)和方差分解分析(B)
Figure 5. A redundancy analysis (A) and variance partitioning analysis (B) considering the influences of vegetation type, tree species, soil properties, fine root traits and leaf traits on root-associated fungal community composition
-
本研究基于高通量测序技术探讨世界自然遗产地梵净山地区3种植被类型根系真菌群落,发现子囊菌门和担子菌门为优势真菌门,森林生态系统中枯枝落叶和枯死木是植物和微生物主要养分来源,子囊菌门和担子菌门的大多数真菌能促进纤维素和木质素等的分解和矿化从而调控生物地球化学循环[24-25]。此外,丛枝菌根真菌为常绿阔叶林的显著差异功能类群,外生菌根真菌是常绿落叶阔叶混交林的显著差异功能类群,可能因为常绿阔叶林养分周转速率一般高于落叶阔叶林,不同菌根真菌采取的养分循环策略存在差异,丛枝菌根真菌通常在养分周转速率快的地区占优势,外生菌根真菌普遍存在于养分循环慢的区域[26]。同时,本研究还发现,杜鹃花类菌根真菌主要出现在亚高山矮林,一方面梵净山地区亚高山矮林的优势植物主要以杜鹃花科为主,这为杜鹃花类菌根真菌的生存提供大量可供选择的宿主;另一方面亚高山矮林生长在山顶土层浅薄、土壤贫瘠、昼夜温差大、温度低的恶劣生境中,丰富和多样化的杜鹃花类菌根真菌可通过促进杜鹃花科植物对矿物质营养元素的吸收及产生具有抗氧化和水解作用的酶来帮助宿主植物获得复合有机物中的养分[27-28],从而增强杜鹃花科植物对山顶恶劣环境的适应能力。
-
常绿阔叶林中植物根系真菌具有最高的α多样性,可能原因如下:第一,常绿阔叶林分布区域的温度高于常绿落叶阔叶混交林和亚高山矮林,而较高的温度能够增强初级生产力及枯枝落叶物等的分解速率,为异养真菌生存提供更多资源;第二,亚热带森林中大部分树种为菌根依赖型树种,菌根真菌的存在能帮助宿主植物获取更多可利用土壤养分。虽然常绿阔叶林土壤磷含量高于其它两种植被类型,但在整个亚热带森林,磷是植物生长的重要限制元素,再加上常绿阔叶林具有最低的土壤氮含量。一方面某些菌根真菌的存在有助于溶解土壤中不能被植物吸收利用的磷酸盐[29]和固定生物氮[30],另一方面宿主植物会倾向于选择更多的菌根真菌搭档来获得生长所必须的氮和磷[29, 31];使得常绿阔叶林具有丰富且多样的真菌种类。此外,真菌群落组成在3种植被类型间存在差异,RDA结果也发现植被类型是影响真菌群落组成的显著因子。同在中国亚热带地区,Han等[32]的研究表明真菌群落组成在常绿阔叶林、落叶阔叶林、针阔混交林、针叶林和次生灌木林存在显著差异。植被类型间不仅植物种类及群落组成、植物功能性状等生物因素存在差异,凋落物厚度及成分、土壤理化性质等非生物因素也有较大差别,从而可能直接或间接影响真菌群落组成。亚高山矮林真菌具有最低的Bray-Curtis相异性指数,是由于亚高山矮林所处区域植物种类(以杜鹃花科植物为主)及土壤、气候条件的独特性而孕育出高度相似的真菌群落。
-
植物功能性状是植物长期适应外部环境变化的结果,近年来越来越多的研究关注根和叶功能性状与真菌群落的关系:Prada-Salcedo等[33]指出根氮含量和根组织密度与真菌群落组成密切相关;Sweeney等[34]发现根直径、根氮含量和比根长在驱动真菌群落组成中发挥着重要作用;而本研究发现根碳含量和叶碳含量能显著影响真菌群落组成,出现这些不同结果可能与宿主植物种类及采样部位的差异有关。当土壤养分受到限制时,宿主植物通常会将自身大量碳交易给菌根共生体,让更多菌根真菌搭档去竞争碳水化合物从而提高养分吸收效益[35],可能是根碳含量和叶碳含量显著影响真菌群落组成且根碳含量与真菌丰富度和Shannon指数呈显著负相关的原因。本研究还发现土壤pH是影响真菌群落组成的显著因子,先前同在森林生态系统中的研究也指出,土壤pH可以影响腐生真菌、丛枝菌根真菌以及外生菌根真菌[36],这是因为土壤pH会改变养分可利用性[37]。此外,也有研究进一步探讨了土壤性质、根性状和叶性状对真菌群落的影响大小:Sweeney等[34]指出与叶性状相比,根性状能决定丛枝菌根真菌和腐生真菌的丰富度和多样性;Dong等[38]发现根系真菌多样性和群落组成主要受土壤性质(土壤pH、土壤碳磷比和氮磷比)的显著影响,而与叶性状无显著相关性。在本研究中,方差分解分析结果表明,细根性状对真菌群落的影响大于土壤性质和叶性状,可能原因:一方面植物细根(吸收根)是大多数真菌的栖息场所,较高比根长和根比表面积能为其生存提供更多空间;另一方面细根是某些真菌与宿主植物进行物质交换区域,植物可能会更加依赖于共生真菌来应对亚热带地区缺磷的土壤环境,使某些真菌功能类群(如丛枝菌根真菌、外生菌根真菌和杜鹃花类菌根真菌等)对细根性状变化较为敏感。对上述原因分析只是基于当前生态学知识的推测,后续研究会从植物种类与真菌群落结构的关系、不同菌根类型与宿主植物的养分获取途径等方面深入解析梵净山地区地上植物群落和地下真菌群落的关联机制。
-
世界自然遗产地梵净山3种典型植被类型的植物根系中,子囊菌门和担子菌门为优势真菌门,小蔓毛壳科、球囊霉科和柔膜菌科为优势真菌科,腐生营养型真菌所占比例较高。丛枝菌根真菌是常绿阔叶林的显著差异功能类群,外生菌根真菌是常绿落叶阔叶混交林的显著差异功能类群,而亚高山矮林的显著差异功能类群为杜鹃花类菌根真菌。植被类型、土壤pH、根碳含量和叶碳含量显著影响根系真菌群落组成,且细根性状对真菌群落的影响大于土壤性质和叶性状。研究结果不仅为丰富世界自然遗产地的物种多样性提供了数据支撑,还能够为深入探讨亚热带地区土壤-真菌-植物养分交易策略奠定基础。
梵净山典型植被类型的根系真菌群落结构及功能类群特征研究
Community Structure of Root-associated Fungi and Functional Characteristics in Typical Vegetation Types of Fanjingshan
-
摘要:
目的 解析世界自然遗产地梵净山地区典型植被类型对真菌群落结构及功能的影响,为深入探讨可见的地上植物群落和未知的地下真菌群落结构间的关联机制提供数据支撑。 方法 以梵净山常绿阔叶林、常绿落叶阔叶混交林和亚高山矮林3种植被类型为研究对象,高通量测序技术探讨根系真菌群落结构在植被类型间的差异,FUNGuild对真菌进行功能预测,并分析真菌群落与土壤性质、细根性状和叶性状间的关系。 结果 子囊菌门和担子菌门为优势真菌门,undefined saprotroph、丛枝菌根真菌、外生菌根真菌、植物病原菌、内生真菌、寄生性真菌和杜鹃花类菌根真菌为主要功能类群。常绿阔叶林、常绿落叶阔叶混交林和亚高山矮林的显著差异功能类群分别是丛枝菌根真菌、外生菌根真菌和杜鹃花类菌根真菌。真菌群落组成在植被类型间存在显著差异,常绿阔叶林真菌多样性和Shannon指数显著高于常绿落叶阔叶混交林和亚高山矮林,而Bray-Curtis相异性指数在亚高山矮林显著低于常绿阔叶林和常绿落叶阔叶混交林。ABT分析表明,根碳含量是影响真菌丰富度的首要因子,叶干物质含量是影响真菌Shannon指数、Simpson指数和Pielou指数的首要因子。冗余分析揭示植被类型、土壤pH、根碳含量和叶碳含量显著影响真菌群落;方差分解分析表明植被类型对真菌群落的影响最大,其次是细根性状和土壤性质,叶性状影响最小。差分解分析表明植被类型对真菌群落的影响最大,其次是细根性状和土壤性质,叶性状影响最小。 结论 植被类型能显著影响真菌群落结构,真菌功能类群与植被类型间存在某种特定联系,植物性状对根系真菌群落的构建具有重要意义。 Abstract:Objective To reveal the effects of vegetation types on root-associated fungal community structure and function in the Fanjingshan World Heritage property, supportive data was provided to predict the relationships between visible vegetation types and invisible community structure in terrestrial ecosystem. Method Three representative vegetation types, including an evergreen broadleaved forest (EBF), an evergreen and deciduous broad-leaved mixed forest (EDBMF) and a subalpine dwarf shrub (SDF), were investigated in the Fanjingshan. Illumina MiSeq sequencing and the FUNGuild annotation tool were used to obtain and analyse the characteristics of the root-associated fungal community structure and function in different vegetation types, and the contributions of soil properties, fine root traits and leaf traits to variations in composition of root-associated fungal community were also analysed. Result Ascomycota and Basidiomycota were the two most abundant phyla, and undefined saprotroph, arbuscular mycorrhizal fungi, ectomycorrhizal fungi, plant pathogen, endophyte, fungal parasite and ericoid mycorrhizal fungi were the main guilds. Arbuscular mycorrhizal fungi were notably enriched in EBF, ectomycorrhizal fungi were more significantly abundant in EDBMF, and ericoid mycorrhizal fungi were significantly overrepresented in SDF. The significant difference in the composition of root-associated fungi among vegetation types was confirmed by non-metric multidimensional scaling analysis and permutational multivariate analysis of variance. A higher alpha diversity was detected in EBF, and the Bray-Curtis dissimilarity index was significantly lower in SDF than in EBF and EDBMF. Aggregated boosted tree analysis indicated that root carbon content was the primary factor influencing observed richness, while leaf dry matter content was the dominant factors associated with the changes in Shannon index, Simpson index and Pielou index of root-associated fungi. Redundancy analysis showed that changes in vegetation type, soil pH, root carbon content and leaf carbon content could significantly affect the fungal community composition. The variance partitioning analysis further revealed that vegetation type had the greatest impact on the composition of root-associated fungi, followed by fine root traits and soil properties, and leaf traits had the least impact. Conclusion Vegetation type can significantly affect root-associated fungal community structure. There is a specific relationship between vegetation type and functional taxa, and plant traits affect the construction of fungal communities. -
Key words:
- vegetation type
- / FUNGuild
- / root-associated fungi
- / functional taxa
- / fine root traits
-
图 1 3种植被类型根系真菌的cladogram图(A)和功能类群的LDA Score图(B)(LDA阈值3.0)
Figure 1. The key root-associated fungal groups (A) and functional taxa (B) corresponding to vegetation type are identified by linear discriminant analysis (LDA) effect size (LEfSe)(LDA score= 3)
图 2 不同植被类型根系真菌多样性指数
Figure 2. The diversity indices of root-associated fungi in different vegetation types
图 3 根系真菌群落的NMDS(A)和Bray-Curtis相异性指数(B)
Figure 3. Nonmetric multidimensional scaling (NMDS) plot (A) and Bray-Curtis dissimilarity index (B) of root-associated fungal community composition based on Bray-Curtis distance
图 4 植被类型、树种、土壤性质、细根性状和叶性状对根系真菌丰富度(A)、Shannon指数(B)、Simpson指数(C)和Pielou指数(D)影响的相对重要性分析
Figure 4. Relative influences of vegetation type, tree species, soil properties, fine root traits and leaf traits on root-associated fungal richness (A), Shannon index (B), Simpson index (C) and Pielou index (D)
图 5 植被类型、树种、土壤性质、细根性状和叶性状与根系真菌群落关系的RDA(A)和方差分解分析(B)
Figure 5. A redundancy analysis (A) and variance partitioning analysis (B) considering the influences of vegetation type, tree species, soil properties, fine root traits and leaf traits on root-associated fungal community composition
表 1 不同植被类型土壤理化性质、细根性状和叶性状
Table 1. Soil properties, fine root traits and leaf traits in different vegetation types
指标
Index常绿阔叶林
EBF常绿落叶阔叶混交林
EDBMF亚高山矮林
SDF土壤 pH
Soil pH6.067 ± 0.101 a 5.858 ± 0.156 a 5.006 ± 0.106 b 土壤全碳
Soil total carbon/(g·kg−1)7.057 ± 0.312 b 12.315 ± 1.677 ab 17.598 ± 2.946 a 土壤全氮
Soil total nitrogen/(g·kg−1)0.560 ± 0.022 b 0.875 ± 0.091 ab 1.127 ± 0.166 a 土壤全磷
Soil total phosphorus/(g·kg−1)6.067 ± 0.015a 5.858 ± 0.054 b 5.006 ± 0.053 ab 根生物量
Root biomass/g0.321 ± 0.063 a 0.239 ± 0.057 a 0.246 ± 0.019 a 根干物质含量
Root dry matter content/(g·g−1)0.255 ± 0.041 a 0.274 ± 0.030 a 0.277 ± 0.009 a 比根长
Specific root length/(cm·g−1)29.242 ± 3.209 b 44.158 ± 4.993 b 71.392 ± 10.885 a 根组织密度
Root tissue density/(g·cm−3)1.115 ± 0.249 a 1.014 ± 0.120 a 0.468 ± 0.052 b 平均根直径
Mean root diameter/cm0.237 ± 0.008 a 0.196 ± 0.014 b 0.220 ± 0.009 ab 根比表面积
Specific surface area/(cm2·g−1)217.188 ± 25.446 b 253.534 ± 28.078 b 479.641 ± 29.043 a 根碳含量
Root carbon content/(g·kg−1)46.115 ± 0.423 b 47.918 ± 0.756 b 50.371 ± 0.217 a 根氮含量
Root nitrogen content/(g·kg−1)2.357 ± 0.108 a 2.134 ± 0.128 ab 1.833 ± 0.050 b 根磷含量
Root phosphorus content/(g·kg−1)0.554 ± 0.015 a 0.673 ± 0.073 a 0.730 ± 0.049 a 叶生物量
Leaf biomass/g1.741 ± 0.222 a 1.609 ± 0.198 a 1.536 ± 0.172 a 叶面积
Leaf area/cm2278.285 ± 24.778 a 299.781 ± 26.502 a 167.156 ± 16.237 b 比叶面积
Specific leaf area/(cm2·g−1)169.114 ± 14.169 a 172.458 ± 19.323 a 138.494 ± 24.582 a 叶干物质含量
Leaf dry matter content/(g·g−1)0.386 ± 0.012 a 0.439 ± 0.017 a 0.370 ± 0.028 a 叶碳含量
Leaf carbon content/(g·kg−1)47.592 ± 0.715 b 47.937 ± 0.837 b 50.751 ± 0.456 a 叶氮含量
Leaf nitrogen content/(g·kg−1)1.902 ± 0.126 a 2.052 ± 0.110 a 1.722 ± 0.131 a 叶磷含量
Leaf phosphorus content/(g·kg−1)0.547 ± 0.039 a 0.701 ± 0.063 a 0.615 ± 0.077 a 注:数据为平均值 ± 标准误,同行后不同小写字母表示在P < 0.05水平上差异显著。
Notes: Data are means ± standard error. Different lowercase letters in the same row indicate significant difference at P < 0.05 level. 下载: 导出CSV
下载: 导出CSV
-
[1] 巫仁霞. 梵净山生态系统多样性全球对比分析与世界遗产价值研究[D]. 贵阳: 贵州师范大学, 2017. [2] 朱佳运. 梵净山植物多样性全球对比分析与世界遗产价值[D]. 贵阳: 贵州师范大学, 2017. [3] 李相楹, 张维勇, 刘 峰, 等. 不同海拔高度下梵净山土壤碳、氮、磷分布特征[J]. 水土保持研究, 2016, 23(3):19-24. [4] 张明明, 杨朝辉, 王 丞, 等. 贵州梵净山国家级自然保护区鸟兽红外相机监测[J]. 生物多样性, 2019, 27(7):813-818. doi: 10.17520/biods.2019131 [5] 李晓笑, 王清春, 崔国发, 等. 濒危植物梵净山冷杉野生种群结构及动态特征[J]. 西北植物学报, 2011, 31(7):1479-1486. [6] 袁 腾, 陶光耀, 江 龙. 梵净山4种林型的土壤丛枝菌根真菌多样性[J]. 东北林业大学学报, 2018, 46(3):83-86. [7] 尚 昆, 石 磊, 李海波, 等. 梵净山不同海拔丛枝菌根真菌多样性[J]. 东北林业大学学报, 2020, 48(2):76-80. [8] VAN DER HEIJDEN M G, KLIRONOMOS J N, URSIC M, et al. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity[J]. Nature, 1998, 396(6706): 69-72. doi: 10.1038/23932 [9] TRESEDER K K, LENNON J T. Fungal traits that drive ecosystem dynamics on land[J]. Microbiology and Molecular Biology Reviews, 2015, 79(2): 243-262. doi: 10.1128/MMBR.00001-15 [10] SMITH S E, READ D J. Mycorrhizal Symbiosis[M]. New York: Academic Press, 2008. [11] TEDERSOO L, BAHRAM M, PÕLME S, et al. Global diversity and geography of soil fungi[J]. Science, 2014, 346(6213): 1256688. doi: 10.1126/science.1256688 [12] 林宇岚, 李正昀, 吴 斐, 等. 不同品种油茶根际丛枝菌根真菌群落结构特征[J]. 林业科学研究, 2020, 33(5):163-169. [13] 梁明月, 苏以荣, 何寻阳, 等. 岩溶区典型灌丛植物根系丛枝菌根真菌群落结构解析[J]. 环境科学, 2018, 39(12):5657-5664. [14] GUO Y Q, HOU L J, ZHANG Z Y, et al. Soil microbial diversity during 30 years of grassland restoration on the Loess Plateau, China: Tight linkages with plant diversity[J]. Land Degradation and Development, 2019, 30(10): 1172-1182. doi: 10.1002/ldr.3300 [15] LIU S E, WANG H, TIAN P, et al. Decoupled diversity patterns in bacteria and fungi across continental forest ecosystems[J]. Soil Biology and Biochemistry, 2020, 144: 107763. doi: 10.1016/j.soilbio.2020.107763 [16] SHENG Y Y, CONG W, YANG L S, et al. Forest soil fungal community elevational distribution pattern and their ecological assembly processes[J]. Frontiers in Microbiology, 2019, 10: 02226. doi: 10.3389/fmicb.2019.02226 [17] COTTAM G, CURTIS J T. The use of distance measures in phytosociological sampling[J]. Ecology, 1956, 37(3): 451-460. doi: 10.2307/1930167 [18] BELL C, CARRILLO Y, BOOT C M, et al. Rhizosphere stoichiometry: are C: N: P ratios of plants, soils, and enzymes conserved at the plant species-level?[J]. New Phytologist, 2014, 201(2): 505-517. doi: 10.1111/nph.12531 [19] PREGITZER K S, DEFOREST J L, BURTON A J, et al. Fine root architecture of nine North American trees[J]. Ecological Monographs, 2002, 72(2): 293-309. doi: 10.1890/0012-9615(2002)072[0293:FRAONN]2.0.CO;2 [20] 鲍士旦. 土壤农化分析(第3版)[M]. 北京: 中国农业出版社, 2000. [21] CALLAHAN B J, MCMURDIE P J, ROSEN M J, et al. DADA2: high-resolution sample inference from Illumina amplicon data[J]. Nature Methods, 2016, 13(7): 581-583. doi: 10.1038/nmeth.3869 [22] KÕLJALG U, NILSSON R H, ABARENKOV K, et al. Towards a unified paradigm for sequence-based identification of fungi[J]. Molecular Ecology, 2013, 22(21): 5271-5277. doi: 10.1111/mec.12481 [23] NGUYEN N H, SONG Z, BATES S T, et al. FUNGuild: an open annotation tool for parsing fungal community datasets by ecological guild[J]. Fungal Ecology, 2016, 20: 241-248. doi: 10.1016/j.funeco.2015.06.006 [24] OSONO T. Functional diversity of ligninolytic fungi associated with leaf litter decomposition[J]. Ecological Research, 2020, 35(1): 30-43. doi: 10.1111/1440-1703.12063 [25] PURAHONG W, WUBET T, LENTENDU G, et al. Determinants of deadwood-inhabiting fungal communities in temperate forests: molecular evidence from a large scale deadwood decomposition experiment[J]. Frontiers in Microbiology, 2018, 9: 02120. doi: 10.3389/fmicb.2018.02120 [26] ZHANG H Y, LÜ X T, HARTMANN H, et al. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scales[J]. Global Ecology and Biogeography, 2018, 27(7): 875-885. doi: 10.1111/geb.12738 [27] ADAMCZYK B, AHVENAINEN A, SIETIÖ O-M, et al. The contribution of ericoid plants to soil nitrogen chemistry and organic matter decomposition in boreal forest soil[J]. Soil Biology and Biochemistry, 2016, 103: 394-404. doi: 10.1016/j.soilbio.2016.09.016 [28] PEROTTO S, DAGHINO S, MARTINO E. Ericoid mycorrhizal fungi and their genomes: another side to the mycorrhizal symbiosis?[J]. New Phytologist, 2018, 220(4): 1141-1147. doi: 10.1111/nph.15218 [29] HORTAL S, PLETT K L, PLETT J M, et al. Role of plant-fungal nutrient trading and host control in determining the competitive success of ectomycorrhizal fungi[J]. The ISME Journal, 2017, 11(12): 2666-2676. doi: 10.1038/ismej.2017.116 [30] HACK C M, PORTA M, SCHÄUFELE R, et al. Arbuscular mycorrhiza mediated effects on growth, mineral nutrition and biological nitrogen fixation of Melilotus alba Med. in a subtropical grassland soil[J]. Applied Soil Ecology, 2019, 134: 38-44. doi: 10.1016/j.apsoil.2018.10.008 [31] JOHNSON N C, WILSON G W, BOWKER M A, et al. Resource limitation is a driver of local adaptation in mycorrhizal symbioses[J]. Proceedings of the National Academy of Sciences, 2010, 107(5): 2093-2098. doi: 10.1073/pnas.0906710107 [32] HAN W J, WANG G M, LIU J L, et al. Effects of vegetation type, season, and soil properties on soil microbial community in subtropical forests[J]. Applied Soil Ecology, 2021, 158: 103813. doi: 10.1016/j.apsoil.2020.103813 [33] PRADA-SALCEDO L D, GOLDMANN K, HEINTZ-BUSCHART A, et al. Fungal guilds and soil functionality respond to tree community traits rather than to tree diversity in European forests[J]. Molecular Ecology, 2021, 30(2): 572-591. doi: 10.1111/mec.15749 [34] SWEENEY C J, DE VRIES F T, VAN DONGEN B E, et al. Root traits explain rhizosphere fungal community composition among temperate grassland plant species[J]. New Phytologist, 2021, 229(3): 1492-1507. doi: 10.1111/nph.16976 [35] BEVER J D. Preferential allocation, physio-evolutionary feedbacks, and the stability and environmental patterns of mutualism between plants and their root symbionts[J]. New Phytologist, 2015, 205(4): 1503-1514. doi: 10.1111/nph.13239 [36] VASCO-PALACIOS A M, BAHRAM M, BOEKHOUT T, et al. Carbon content and pH as important drivers of fungal community structure in three Amazon forests[J]. Plant and Soil, 2020, 450: 111-131. doi: 10.1007/s11104-019-04218-3 [37] VAN SUNDERT K, RADUJKOVIĆ D, COOLS N, et al. Towards comparable assessment of the soil nutrient status across scale-Review and development of nutrient metrics[J]. Global Change Biology, 2020, 26(2): 392-409. doi: 10.1111/gcb.14802 [38] DONG H Y, GE J F, SUN K, et al. Change in root-associated fungal communities affects soil enzymatic activities during Pinus massoniana forest development in subtropical China[J]. Forest Ecology and Management, 2021, 482: 118817. doi: 10.1016/j.foreco.2020.118817 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3824
- HTML全文浏览量: 1273
- PDF下载量: 105
- 被引次数: 0
