

-
杨树是杨柳科(Salicaceae Mirb.)杨属(Populus L.)多年生落叶乔木,因其速生成林、防风固沙、抗逆性强、木材质佳等优势,成为我国用材林、防护林主要造林树种之一[1]。我国杨树栽培规模不断扩大,但受限于栽培技术和杨树病虫害的日益频发。杨树叶锈病是一种严重的叶部病害,导致杨树的生长量和材积严重降低,造成杨树人工林巨大经济损失[2]。杨树叶锈病的病原菌多为栅锈菌属(Melampsora Castagne)锈菌。北美栅锈菌(Melampsora medusae Thümen.)原寄主为北美洲东部的美洲黑杨(Populus deltoides W. Bartram ex Marshall),后逐渐传播至世界各地,是印度、欧洲、日本等国家和地区的检疫性有害生物[3],也是我国最近发现的外来入侵种,危害极大,并有持续传播的趋势[4]。该种与我国本土流行种松杨栅锈菌(Melampsoralarici-populina Kleb.)在形态、寄主谱范围等方面高度相似,给杨树叶锈病的检验检疫带来了实际困难,因此亟待建立一种快速准确的杨树叶锈病检测技术。
目前林木真菌病原检测方法主要包括传统检测法和分子生物学法。传统检测法多为形态观察,较为直观但对一线检疫人员的要求较高。如形态结构相似的同属不同种真菌,往往需要扫描电子显微镜(SEM)才能实现有效辨别;同种病原菌在不同时期或不同生长条件下也会表现出不同的形态特征等。因此,对新病原菌或疑难种病原菌进行物种水平的辨别常常有困难。分子生物学法中广泛使用的常规PCR及以常规PCR为基础建立的多重PCR、实时荧光PCR等技术特异性强,检出率高,但依赖昂贵的仪器,耗时较长,且需要专业的技术人员进行操作[5-7]。因此,以上两种方法很难满足目前生产实践中快速准确检测的需求。
环介导等温扩增技术(loop-mediated isothermal amplification,LAMP)是Notomi等人在2000年建立的一种恒温核酸扩增技术,在恒定温度下,利用4~6条引物及Bst DNA聚合酶对模板进行扩增,经30~60 min可将微量模板扩增109倍[8-9]。LAMP结果检测方法多样化,除了琼脂糖凝胶电泳外,目前常用的方法还有浊度法、显色法、实时荧光法等,这些方法可以实现闭管检测,从而降低污染概率[10-11]。LAMP具特异性好、灵敏度高、反应速度快和操作简易便捷等特点,应用前景广阔[12-14],在林业病害检测方面已有许多报道[15-16]。另外,LAMP还广泛应用于医学[17-18]、农业[19-20]、食品[21-22]等行业。
本研究以北美栅锈菌和松杨栅锈菌为研究对象,根据LAMP的反应原理,针对两种不同锈菌的靶标序列分别设计了多组引物。经实验初步筛选引物后,再以北美栅锈菌(M. medusae)、松杨栅锈菌(M. larici-populina)、图拉斯叉钩丝壳菌(Sawadaia tulasnei (Fuck.) Homma)、芍药白粉菌(Erysiphe paeoniae Zheng&Cheng)、梨胶锈菌(Gymnosporangium asiaticum Miyabe ex Yamada)、羊肚菌(Morchella esculenta (L.) Pers.)、金针菇(Flammulina velutipes (Curt.:Fri.) Sing)的基因组DNA为模板进行特异性检测,后续进行体系优化、反应条件优化、灵敏度检测等研究,并结合羟基萘酚蓝(HNB)建立LAMP-HNB可视化检测体系,以期为松杨栅锈菌和北美栅锈菌的快速、准确的鉴别及鉴定提供技术支撑。
-
供试北美栅锈菌(M. medusae)、松杨栅锈菌(M. larici-populina)、羊肚菌(M. esculenta)、金针菇(F. velutipes),菌株由西北农林科技大学森林病理学实验室保存并提供,其余参考菌株包括图拉斯叉钩丝壳菌(S. tulasnei)、芍药白粉菌(E. paeoniae)、梨胶锈菌(G. asiaticum)均采自西北农林科技大学南校区内感病植株。等温扩增试剂盒、引物均购于生工生物(上海)股份有限公司;Bst DNA聚合酶购于NEB(北京)公司;羟基萘酚蓝购于上海麦克林生化科技有限公司;dNTPs购于宝日医(大连)公司。
-
将CTAB缓冲液置于65 ℃水浴锅中充分裂解。向盛有样品的离心管中加入500 μL CTAB缓冲液,添加适量石英砂充分研磨,再加入2 μL巯基乙醇,置于恒温孵育器中65 ℃保温1 h,每10 min颠倒均匀一次。保温结束后加入500 μL DNA提取液(氯仿∶异戊醇=24∶1),室温离心11 000 rpm,10 min,2次。取上清液,加入2倍体积无水乙醇,上下颠倒均匀,−20 ℃保存2~3 h后取出,室温离心11 000 rpm,10 min。弃上清液,加入75%乙醇750 μL,再室温离心12 000 rpm,10 min,2次,弃上清液,于超净工作台吹干后加入50 μL TE缓冲液,于−20 ℃保存,备用。
-
在NCBI上获取M. medusae和M. larici-populina的28SrDNA基因序列。根据两种菌的28srDNA序列,利用在线LAMP引物设计工具(PrimerExplorer V5)分别设计多组引物,以dimer值、引物两端自由能为依据初步选出若干组引物(每组引物包括两条内引物:FIP、BIP,两条外引物:F3、B3),送至生工生物(上海)股份有限公司合成。
根据研究目的,需确保引物在M. medusae与M. larici-populina之间不发生交叉反应,因此需要对引物进一步筛选。每组引物分别以M. medusae、M. larici-populina基因组DNA为模板进行LAMP扩增反应。参考厂家等温扩增试剂盒说明书建立反应体系。25 μL体系包括12.5 μL LAMP MasterMix、2 μL FIP、2 μL BIP、0.5 μL F3、0.5 μL B3、2 μL模板DNA、0.5 μL Bst DNA聚合酶,最后ddH2O补至25 μL。在冰上将各组分置于PCR管中并涡旋混匀,反应条件为65 ℃水浴60 min后80 ℃水浴10 min。反应结束后取7 μL产物进行1.5%琼脂糖凝胶电泳检测。通过观察LAMP梯状条带,选出仅扩增M. medusae或M. larici-populina基因组DNA的引物进行后续特异性检测。
-
在北美栅锈菌引物特异性检测中,同时以提取的松杨栅锈菌、芍药白粉菌、图拉斯叉钩丝壳菌、梨胶锈菌、羊肚菌、金针菇基因组DNA为模板进行LAMP扩增反应;同样地,松杨栅锈菌引物特异性检测中,同时以提取的北美栅锈菌及上述菌种基因组DNA为模板进行LAMP扩增反应;两组试验均以ddH2O为空白对照。反应体系和条件同1.2.2。反应结束后取7 μL产物在1.5%琼脂糖进行凝胶电泳。通过观察LAMP梯状条带判断LAMP方法的特异性。
-
参考NEB公司的Bst DNA聚合酶使用说明书建立初始反应体系(表1)。
表 1 初始反应体系
Table 1. Initial reaction system
组分及浓度
Components and Concentrations体积
Volume/μL10 × Buffer 2.5 Mg2+(100 mmol·L−1) 1.5 dNTPs(10 mmol·L−1 each) 3.5 FIP(10 μmol·L−1) 4.0 BIP(10 μmol·L−1) 4.0 F3(10 μmol·L−1) 0.5 B3(10 μmol·L−1) 0.5 模板 template 1.5 Bst DNA聚合酶
Bst DNA polymerase(8 000 U·mL−1)0.5 ddH2O 补至25 μL 在冰上将各组分置于PCR管中并涡旋混匀,反应条件为65 ℃水浴60 min后80 ℃水浴10 min。
在初始反应体系和条件的基础上,按表2设置的梯度依次对Mg2+、内外引物比例、dNTPs进行单因素优化,同时以ddH2O为空白对照,反应结束后取7 μL产物进行1.5%琼脂糖凝胶电泳检测。根据LAMP梯状条带的有无以及清晰度确定最佳浓度及比例。
表 2 组分梯度
Table 2. component gradient
因素
Factor梯度
GradientMg2+浓度
Mg2+concentration/(mmol·L−1)2、4、6、8、10 内外引物比例
ratio of internal to external primer1:1、2:1、4:1、6:1、8:1、10:1 dNTPs浓度
dNTPs concentration/(mmol·L−1)1、1.2、1.4 在优化后的体系中加入2 μL羟基萘酚蓝( 2 mmol·L−1)指示LAMP反应的结果。同时以ddH2O为空白对照,通过目视溶液颜色,判断是否发生反应(阳性为蓝色,阴性为紫色),反应结束后取7 μL产物进行1.5%琼脂糖凝胶电泳检测,观察羟基萘酚蓝的指示效果。
-
在组分优化后的体系中加入羟基萘酚蓝为显色剂,并依次进行温度和时间的优化。初始反应温度为65 ℃,初始反应时间为60 min。优化反应温度时,反应时间为初始反应时间,温度梯度设置如表3所示;优化反应时间时,以上一步得出的优化温度作为反应温度,时间梯度设置如表3所示。以ddH2O为空白对照,筛选出颜色分化时对应的最低反应温度和最短反应时间,从而进一步优化反应条件。
表 3 条件梯度
Table 3. Conditional gradient
因素
Factor梯度
Gradient温度 Temperature/℃ 61、63、65、67 时间 Time/min 30、40、50、60 -
M. medusae和M. larici-populina基因组DNA经浓度测定后进行10倍浓度的梯度稀释,在优化后的体系和反应条件中进行反应。同时以ddH2O为空白对照,观察反应后的溶液颜色以判断最低检测浓度。
-
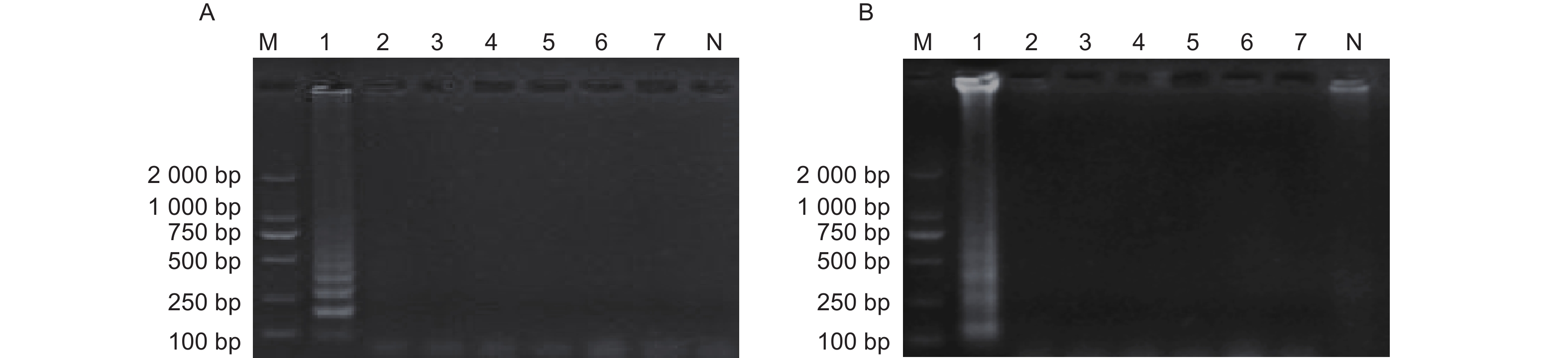
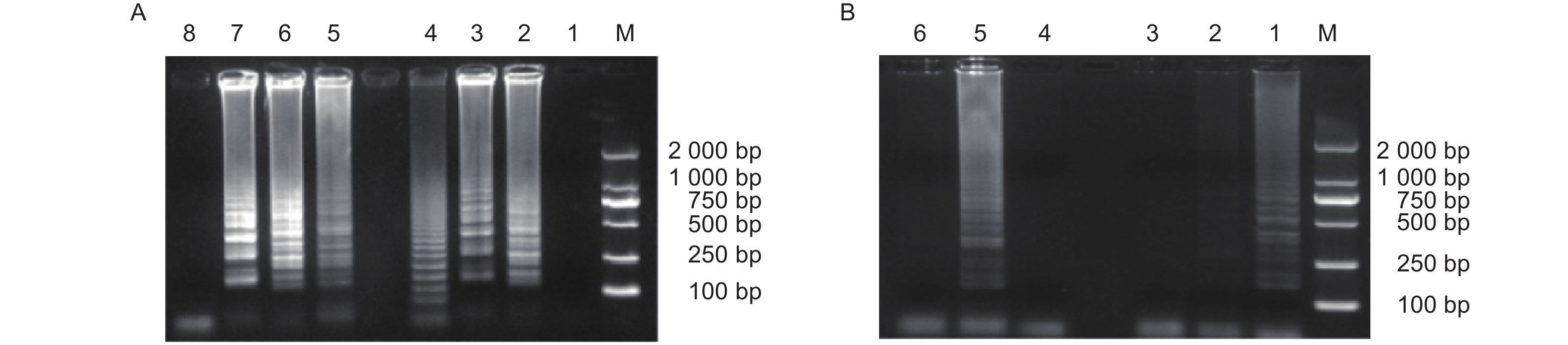
通过在线引物设计软件共设计了12组引物,经过初步筛选,得到M. medusae引物BM① 、BM② 、BM③ 、BM④以及M. larici-populina引物SY⑤ 、SY⑥ 、SY⑦ 。进一步的引物筛选结果如图1所示。图1-A显示M. medusae的4组引物中,BM①所属的电泳道中仅以M. larici-populina基因组DNA为模板进行LAMP反应时出现梯状条带,表明BM①无法扩增目标菌;BM②和BM③所属的电泳道均出现了梯状条带,表明这两组引物既能以M. medusae基因组DNA为模板进行扩增,又能以M. larici-populina基因组DNA为模板进行扩增,因此不具有特异性;BM④所属的两个电泳道中,仅以M. medusae基因组DNA为模板进行LAMP反应时出现了梯状条带。因此,4组引物中只有BM④能够特异性扩增M. medusae。图1-B显示M. larici-populina的3组引物中,SY⑤所属的电泳道中仅以M. medusae基因组DNA为模板进行LAMP反应时出现梯状条带,表明SY⑤不能扩增目标菌;SY⑥所属的两个电泳道中,仅以M. larici-populina基因组DNA为模板进行LAMP反应时出现了梯状条带;SY⑦所属的两个电泳道中均无梯状条带,表明SY⑦发生无效扩增;因此,3组引物中只有SY⑥能够特异性扩增M. larici-populina。综上,仅BM④和SY⑥能够有效区分M. medusae和M. larici-populina,因此可用于建立LAMP检测体系。二者序列如表4所示。

图 1 供试引物筛选
Figure 1. Tested primer’s selection
表 4 特异性引物序列
Table 4. Specific primer sequences
编号
No.引物名称
Primer name引物序列
Primer sequencesBM④ F3 AGCAAGTCAACATCAGTCT B3 CTAATACTCGCAAGCATGT FIP TCGAGGTCCCAAGCTATAACAGAGTGTTGGAAAAAGGGC BIP GTTCGAAAGAGCCTCCTTACTCTTGGTCCGTGTTTCAAG SY⑥ F3 AAGCTTTGAGCGGATTCGA B3 AATGATCCAGACTGGTCGA FIP CCGTGTTTCAAGACGGGTCGGATCTCCTTACTATGGATGTTGG BIP ACCAAGGAGTCTAACATGCTTGCGGATCACATCTGAAATTCACT -
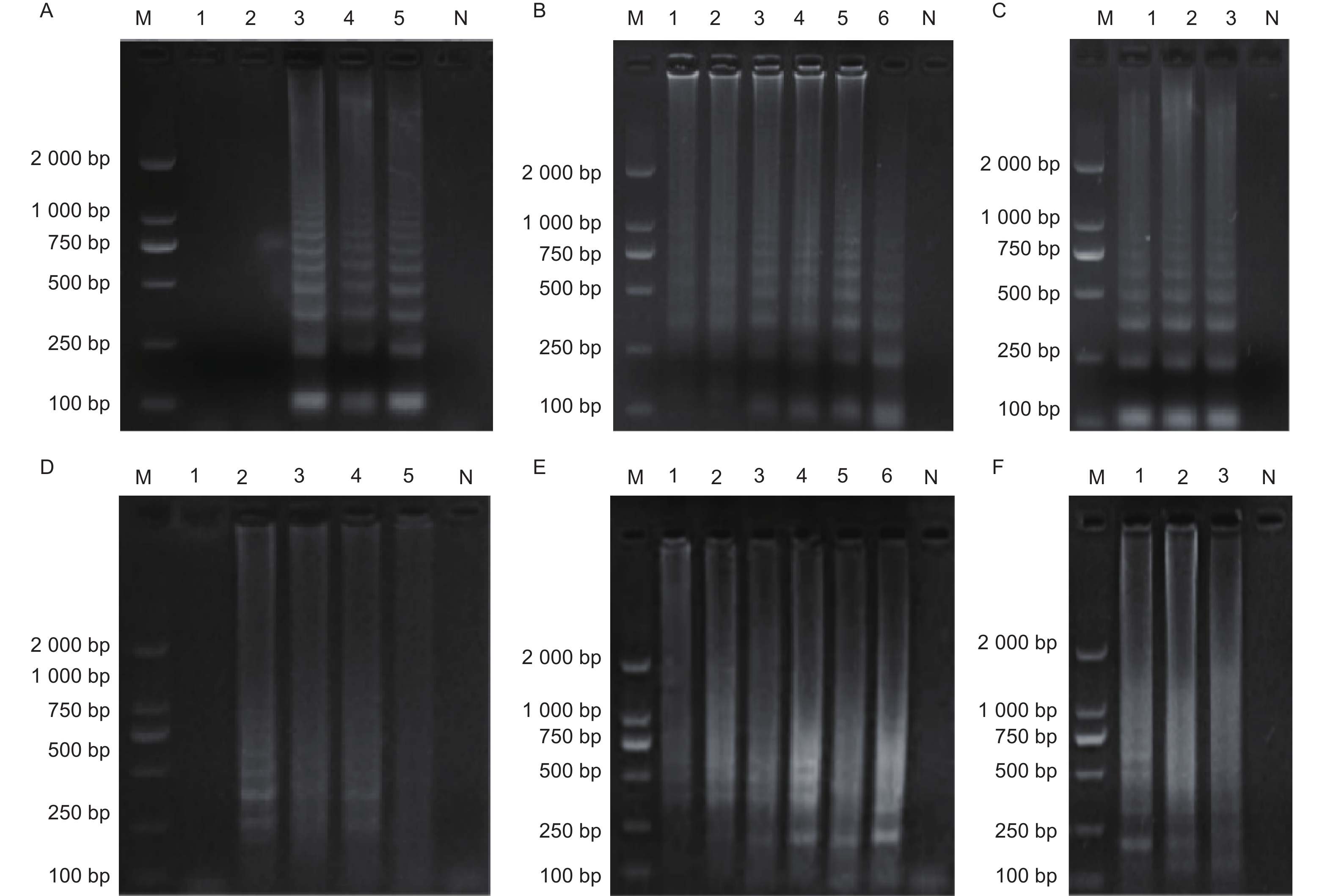
图2-A显示,BM④只有以M. medusae基因组DNA为模板进行LAMP扩增时出现了典型的梯状条带,M. larici-populina、E. paeoniae、S. tulasnei等菌株以及空白对照均未出现梯状条带;图2-B显示,SY⑥只有以M. larici-populina基因组DNA为模板进行LAMP扩增时出现了典型的梯状条带,M. medusae、E. paeoniae、S. tulasnei等菌株以及空白对照均未出现梯状条带。综上,BM④和SY⑥具备特异性扩增目标菌的能力。

图 2 引物特异性检测
Figure 2. Primer-specific detections
-
图3-A第3泳道、图3-B第5泳道、图3-C第2泳道梯状条带最为清晰明亮,因此确定25 μL M. medusae体系最佳Mg2+浓度为6 mmol·L−1,最佳内外引物比例为8:1,最佳dNTPs浓度为1.2 mmol·L−1;图3-D第2泳道、图3-E第4泳道、图3-F第1泳道中梯状条带最为清晰明亮,因此确定25 μL M. larici-populina体系最佳Mg2+浓度为4 mmol·L−1,最佳内外引物比例为6∶1,最佳dNTPs浓度为1 mmol·L−1。

图 3 反应体系优化
Figure 3. Reaction system optimization
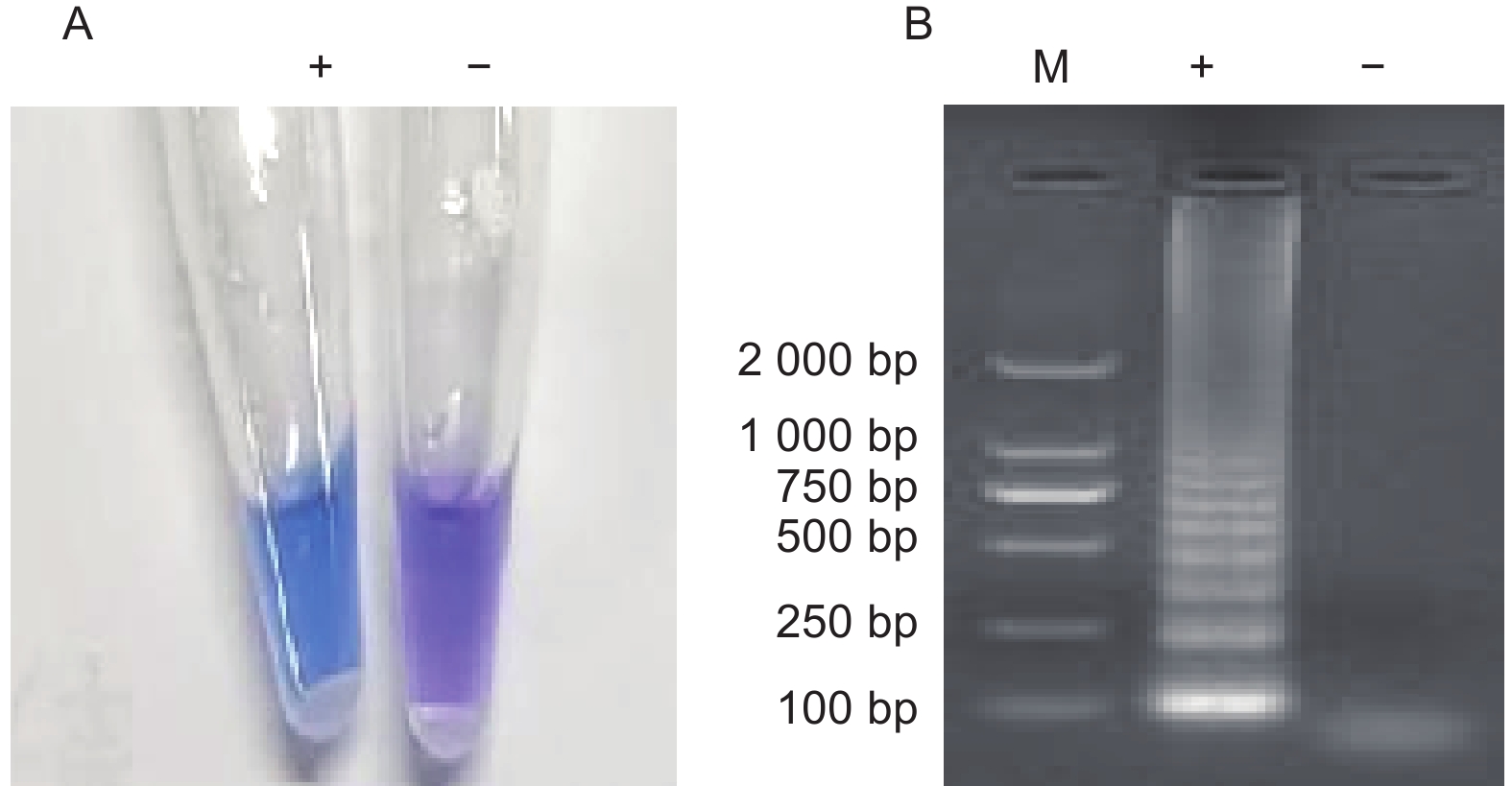
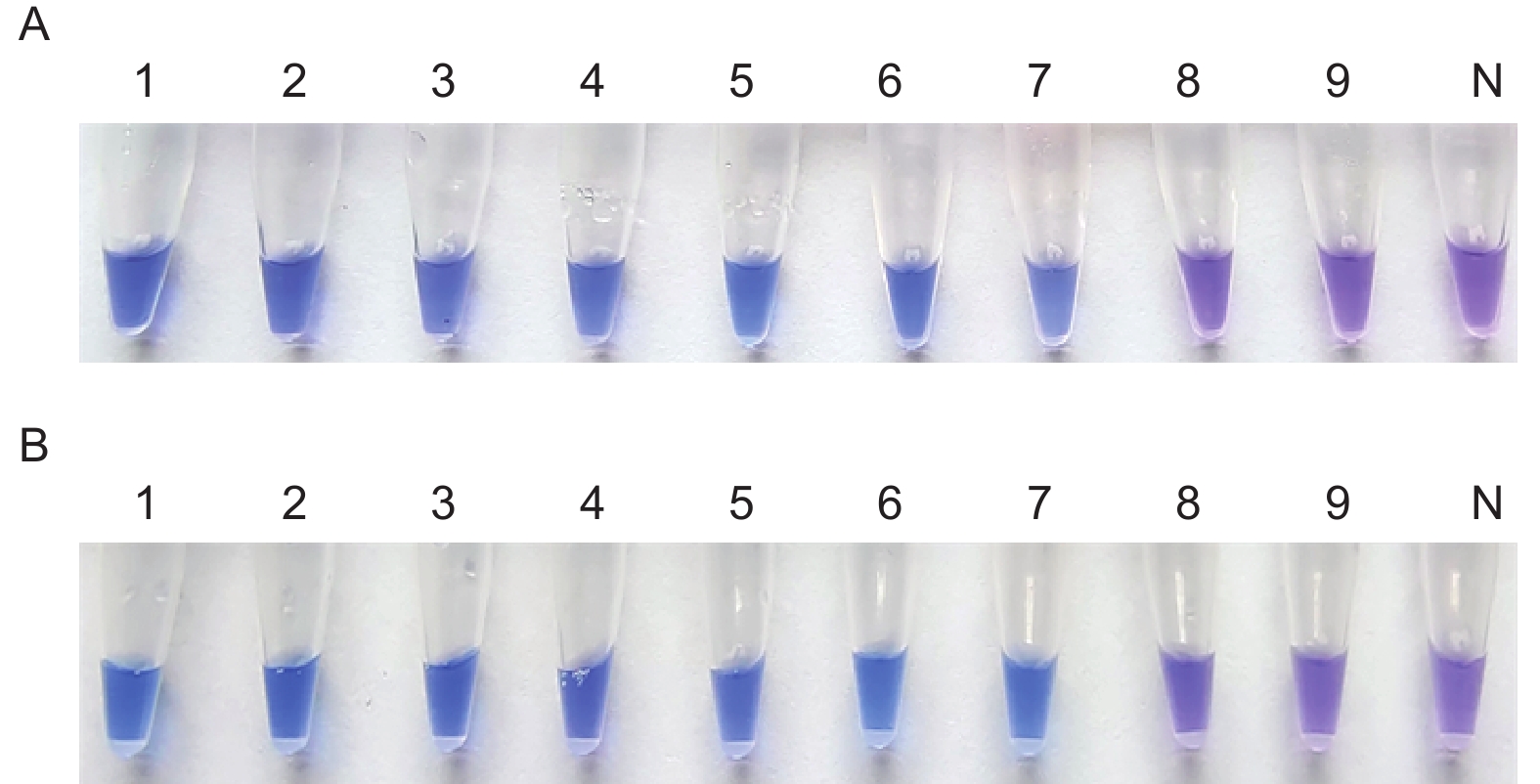
图4-A左PCR管中溶液呈蓝色为阳性,对应图4-B左泳道出现典型的梯状条带;图4-A右PCR管中溶液呈紫色为阴性,对应图4-B右泳道无任何条带,显色结果与电泳结果相互印证,不同结果颜色区分明显。

图 4 可视化体系建立
Figure 4. Establishment of visualization system
-
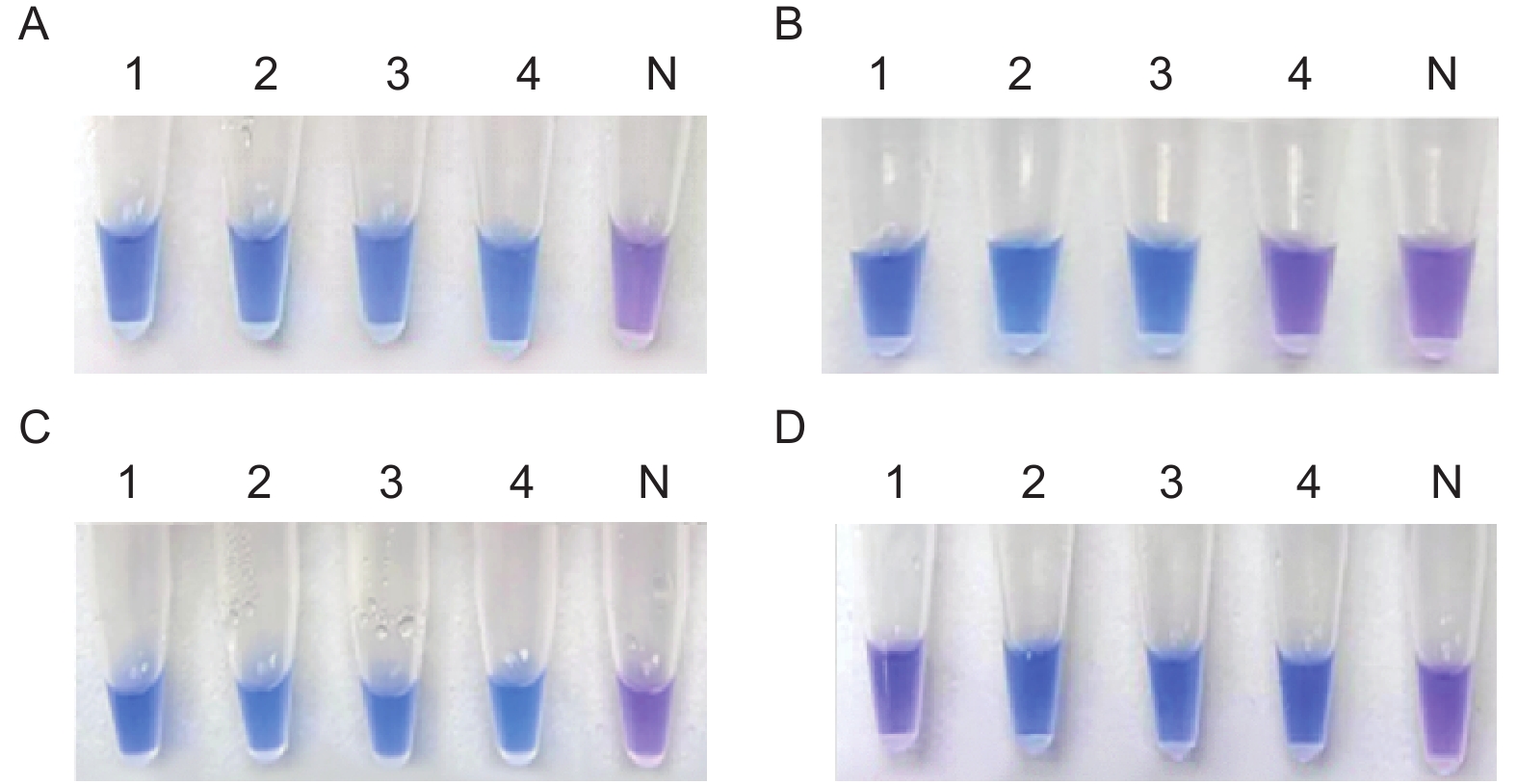
图5-A中1~4号PCR管中溶液呈明显蓝色为阳性结果,表明M. medusae体系在61 ℃、63 ℃、65 ℃、67 ℃的温度条件下均能发生扩增反应且最低温度为61 ℃;图5-C中1~4号PCR管中溶液呈明显蓝色为阳性结果,表明温度为61 ℃时,M. medusae体系在30 min、40 min、50 min、60 min的时间条件下均能充分扩增且最短时间为30 min。

图 5 反应条件优化
Figure 5. The optimization of reaction conditions
图5-B中1~3号PCR管中溶液呈明显蓝色为阳性结果,4号PCR管中溶液呈紫色为阴性结果,表明温度为67 ℃时不利于M. larici-populina体系进行扩增,因此能够保证M. larici-populina体系发生扩增的反应温度为61 ℃、63 ℃、65 ℃,最低温度为61 ℃;图5-D中1号PCR管中溶液呈紫色为阴性结果,2~4号PCR管中溶液呈明显蓝色,为阳性结果,表明在温度为61 ℃,反应时间为30 min时,M. larici-populina体系由于时间过短无法充分扩增而导致HNB显色不佳,因此能够保证M. larici-populina体系充分反应的时间为40 min、50 min、60 min,最短时间为40 min。
最终得到体系和反应条件如表5所示。
表 5 最终体系及反应条件
Table 5. Terminal optimization of reaction system and reaction conditions
组分及浓度
Components and Concentrations体积/μL
VolumeM.medusae M. larici-populina 10 × Buffer 2.5 2.5 Mg2+(100 mmol·L−1) 1.0 0.5 dNTPs (10 mmol·L−1 each) 3.0 2.5 FIP (10 μmol·L−1) 4.0 3.0 BIP (10 μmol·L−1) 4.0 3.0 F3 (10 μmol·L−1) 0.5 0.5 B3 (10 μmol·L−1) 0.5 0.5 模板 template 1.5 1.5 Bst DNA聚合酶 (8 000 U·ml−1)
Bst DNA polymerase0.5 0.5 羟基萘酚蓝 (2 mmol·L−1)
Hydroxynaphthol blue2.0 2.0 ddH2O 补至25 μL 反应条件
Reaction conditions61 ℃水浴30 min后80 ℃水浴10 min
Water bath at 61 ℃ for 30 min followed 80 ℃ for 10 min61 ℃水浴40 min后80 ℃水浴10 min
Water bath at 61 ℃ for 40 min followed 80 ℃ for 10 min -
图6-A所示1~7管呈现蓝色为阳性,其余呈紫色为阴性,即将出现颜色分化时对应的最低浓度梯度为34 fg·μL−1。图6-B所示1~7管呈现蓝色为阳性,其余呈紫色为阴性,即将出现颜色分化时对应的最低浓度梯度为60 fg·μL−1。因此M. medusae反应体系灵敏度可达34 fg·μL−1,M. larici-populina反应体系灵敏度可达60 fg·μL−1。

图 6 LAMP灵敏度检测
Figure 6. Sensitivity detection of LAMP
-
一直以来,松杨栅锈菌和北美栅锈菌引起的杨树叶锈病困扰着杨树产业发展,建立快速的杨树叶锈病分子检测方法在口岸检疫、产地检疫及锈病流行监测等方面具有重要意义。2019年,陆红艳以松杨栅锈菌HMJAU85核糖体RNA基因为靶序列设计了LAMP特异性引物,在试剂盒的基础上建立了基于钙黄绿素-Mn2+显色的松杨栅锈菌可视化检测体系,在65 ℃下反应50 min即可观察结果,为青杨锈病的检测提供了重要的技术支持[23]。但在栅锈菌属的鉴别上,特别是北美栅锈菌的鉴别上,国内外均缺乏相关研究。本研究建立了能够相互区别的北美栅锈菌和松杨栅锈菌的LAMP检测体系并且可以实现对这两种锈菌的鉴定。与陆红艳的研究相比,本研究中LAMP检测的灵敏度略低,为60 fg·μL−1,但在显色剂的选择、反应条件、检测成本3个方面均有所改进。首先,钙黄绿素作为LAMP常用显色剂之一,与MnCl2搭配使用,二者之间的配比要极为精细才有明显效果,因此需要深入研究离子配比;有研究表明,Mn2+会降低LAMP反应效率,使Bst DNA聚合酶的错配率达0.8%[10,24],本研究使用的显色剂为羟基萘酚蓝,具有颜色区分度好、配置方便,无需优化离子配比等优势,且对LAMP反应过程无影响;在配置体系时最后加入Mg2+并混匀,可得到更好的显色效果。其次,本研究将松杨栅锈菌的检测温度降低至61 ℃,检测时间缩短至40 min,在反应温度和时间上得以进一步优化,也可以降低发生非特异性扩增的概率。最后,以试剂盒为基础建立的反应体系将导致检测经济成本提高,本研究通过自配试剂并探索最佳检测体系及反应条件,在降低检测成本的同时也保证了引物和各组分间的高效配合。
具有特异性的有效引物是建立检测体系的关键。LAMP引物设计难度较高,引物序列长,一般需要通过实验检验其特异性。大多数LAMP体系建立过程中,往往先建立好最优体系,再进行特异性检测。若最终检测结果表明引物无特异性,则需要重新设计引物,这会导致人力、物力及时间的浪费。因此本研究将引物特异性检测放至建立体系之前,先用通用试剂盒建立的稳定体系对引物进行筛选及特异性检测,若引物不具备特异性,则可以直接排除,从而提高了引物筛选效率。
LAMP因其特异性强、灵敏度高、操作简易而备受关注;同时,LAMP技术也存在一定的局限性。LAMP扩增的产物量大,如有操作不慎,极易造成气溶胶污染,导致假阳性结果。有研究表明,建立UDG-dUTP反应系统可有效解决气溶胶污染问题,可尝试在本研究建立的体系基础上加入UDG酶以及dUTP,以增强反应特异性[25]。为了将LAMP检测技术应用于现场快速检测,本研究可以结合锈菌夏孢子快速提取DNA技术[26],建立更高效的杨树叶锈病LAMP检测技术,为病害的检疫和防控争取更多的主动性。
-
本研究通过环介导等温扩增技术(LAMP)建立了特异性强、灵敏度高的可视化检测体系,实现了北美栅锈菌和松杨栅锈菌快速、准确的鉴定及区分。本研究设计的两组引物可特异性识别相应的靶基因序列。在体系中加入160 μM羟基萘酚蓝后可以直观地对供试的病原菌进行鉴别,极大地克服了传统电泳检测的弊端,并且检测体系灵敏度分别达到了34 fg·μL−1和60 fg·μL−1。基于本研究建立的可视化体系,在61 ℃恒温条件下,分别反应30 min与40 min后即可观察结果,可达到快速检测的目的。
北美栅锈菌和松杨栅锈菌可视化检测体系建立
Establishment of a Visual Detection System for Melampsora medusae and Melampsora larici-populina
-
摘要:
目的 为实现北美栅锈菌和松杨栅锈菌快速有效的鉴别及鉴定。 方法 本研究根据两种锈菌的28SrDNA基因序列设计若干组LAMP引物。经筛选得到的引物以北美栅锈菌、松杨栅锈菌、图拉斯叉钩丝壳菌、芍药白粉菌、梨胶锈菌、羊肚菌、金针菇的基因组DNA为模板进行LAMP反应并完成特异性检测;首先建立初始LAMP反应体系,再进一步优化LAMP反应体系的组分和反应条件,加入羟基萘酚蓝实现可视化检测;最后确定检测体系灵敏度。 结果 表明,筛选出的引物具有特异性;25 μL北美栅锈菌LAMP检测体系最佳Mg2+浓度为6 mmol·L−1,最佳内外引物比例为8:1,最佳dNTPs浓度为1.2 mmol·L−1;同样地,25 μL松杨栅锈菌LAMP检测体系最佳Mg2+浓度为4 mmol·L−1,最佳内外引物比例为6:1,最佳dNTPs浓度为1 mmol·L−1。加入160 μM羟基萘酚蓝(HNB)可清晰地指示反应结果,两种检测体系在61 ℃条件下,分别反应30 min和40 min可实现目视判断结果,且灵敏度分别可达34 fg·μL−1和60 fg·μL−1。 结论 通过建立两种锈菌的可视化LAMP-HNB检测体系可实现对北美栅锈菌和松杨栅锈菌进行区分及鉴定,为快速鉴定和区分重要杨树锈病提供了技术支撑和实践参考。 Abstract:Objective To achieve the rapid and effective differentiation and identification of M. medusae and M. larici-populina. Methods In this study, several sets of primers for loop-mediated isothermal amplification (LAMP) were designed based on the 28S ribosomal DNA gene of the two rust fungi. The putative primers selected from experiments were employed for LAMP reactions, and for the specific detection with the controls of genomic DNA from M. medusae, M. larici-populina, Sawadaia tulasnei, Erysiphe paeoniae, Gymnosporangium asiaticum, Morchella esculenta and Flammulina velutipes. The initial LAMP reaction system was established firstly and then the reaction system components and reaction conditions were further optimized. Hydroxynaphthol blue dye (HNB) was supplied into the reaction system to realize visual detection. Finally the sensitivity of the detection system was determined by the lowest DNA substrate. Results The results showed that the selected primers had specificity-species. The optimal Mg2 + concentration in the 25 μL LAMP detection system for M. medusae was 6 mmol·L−1, the optimal internal and external primer ratio was 8:1, and the optimal dNTPs concentration was 1.2 mmol·L−1. Meanwhile, the correspondent case for M. larici-populina was 4 mmol·L−1, 6:1 and 1 mmol·L−1. And all reaction products could be clearly detected with 160 μM hydroxynaphthol blue dying(HNB). The two rusts detection system can be visually determined at 61 ℃ for 30 min and 40 min, respectively, each corresponding to the lowest DNA substrate concentration of 34 fg·μL−1 and 60 fg·μL−1. Conclusion Through establishing a visual LAMP-HNB detection system, M. medusae and M. larici-populina can be differentiated and identified by the LAMP technology. This study provides a technological support in practice for rapid identification and detection of the important poplar rust diseases. -
Key words:
- Poplar
- / M. medusae
- / M. larici-populina
- / LAMP
- / Hydroxynaphthol blue dying
-
表 1 初始反应体系
Table 1. Initial reaction system
组分及浓度
Components and Concentrations体积
Volume/μL10 × Buffer 2.5 Mg2+(100 mmol·L−1) 1.5 dNTPs(10 mmol·L−1 each) 3.5 FIP(10 μmol·L−1) 4.0 BIP(10 μmol·L−1) 4.0 F3(10 μmol·L−1) 0.5 B3(10 μmol·L−1) 0.5 模板 template 1.5 Bst DNA聚合酶
Bst DNA polymerase(8 000 U·mL−1)0.5 ddH2O 补至25 μL  下载: 导出CSV
下载: 导出CSV
表 2 组分梯度
Table 2. component gradient
因素
Factor梯度
GradientMg2+浓度
Mg2+concentration/(mmol·L−1)2、4、6、8、10 内外引物比例
ratio of internal to external primer1:1、2:1、4:1、6:1、8:1、10:1 dNTPs浓度
dNTPs concentration/(mmol·L−1)1、1.2、1.4
下载: 导出CSV
表 3 条件梯度
Table 3. Conditional gradient
因素
Factor梯度
Gradient温度 Temperature/℃ 61、63、65、67 时间 Time/min 30、40、50、60
下载: 导出CSV
表 4 特异性引物序列
Table 4. Specific primer sequences
编号
No.引物名称
Primer name引物序列
Primer sequencesBM④ F3 AGCAAGTCAACATCAGTCT B3 CTAATACTCGCAAGCATGT FIP TCGAGGTCCCAAGCTATAACAGAGTGTTGGAAAAAGGGC BIP GTTCGAAAGAGCCTCCTTACTCTTGGTCCGTGTTTCAAG SY⑥ F3 AAGCTTTGAGCGGATTCGA B3 AATGATCCAGACTGGTCGA FIP CCGTGTTTCAAGACGGGTCGGATCTCCTTACTATGGATGTTGG BIP ACCAAGGAGTCTAACATGCTTGCGGATCACATCTGAAATTCACT
下载: 导出CSV
表 5 最终体系及反应条件
Table 5. Terminal optimization of reaction system and reaction conditions
组分及浓度
Components and Concentrations体积/μL
VolumeM.medusae M. larici-populina 10 × Buffer 2.5 2.5 Mg2+(100 mmol·L−1) 1.0 0.5 dNTPs (10 mmol·L−1 each) 3.0 2.5 FIP (10 μmol·L−1) 4.0 3.0 BIP (10 μmol·L−1) 4.0 3.0 F3 (10 μmol·L−1) 0.5 0.5 B3 (10 μmol·L−1) 0.5 0.5 模板 template 1.5 1.5 Bst DNA聚合酶 (8 000 U·ml−1)
Bst DNA polymerase0.5 0.5 羟基萘酚蓝 (2 mmol·L−1)
Hydroxynaphthol blue2.0 2.0 ddH2O 补至25 μL 反应条件
Reaction conditions61 ℃水浴30 min后80 ℃水浴10 min
Water bath at 61 ℃ for 30 min followed 80 ℃ for 10 min61 ℃水浴40 min后80 ℃水浴10 min
Water bath at 61 ℃ for 40 min followed 80 ℃ for 10 min
下载: 导出CSV
-
[1] 曾伟生, 陈新云, 杨学云, 等. 我国人工杨树生物量建模和生产力分析[J]. 林业科学, 2019, 55(11):1-8. [2] 何经纬, 张伊莹, 田呈明, 等. 区域景观格局对杨树锈病为害流行的影响—以北京延庆地区银白杨为例[J]. 林业科学, 2020, 56(4):99-108. [3] BOUTIGNY A L, GUINET C, VIALLE A,et al. A sensitive real-time PCR assay for the detection of the two Melampsora medusae formae speciales on infected poplar leaves[J]. European Journal of Plant Pathology, 2013, 136(3): 433-441. doi: 10.1007/s10658-013-0180-0 [4] ZHENG W, NEWCOMBE G, HU D,et al. The first record of a north American poplar leaf rust fungus, Melampsora medusae, in China[J]. Forests, 2019, 10(2): 182. doi: 10.3390/f10020182 [5] 王云飞, 张 昊, 杨美欣, 等. 南方镰孢Fusarium meridionale特异性PCR检测方法的建立与应用[J]. 植物保护, 2022, 48(2):162-166,172. [6] 杨玉文, 乔 培, 刘德华, 等. 基于多重PCR技术的西瓜噬酸菌分组检测方法及其应用[J]. 植物保护, 2022, 48(4):211-219. [7] 陈鑫源. 木材腐朽菌的DNA提取及PCR检测研究[D]. 哈尔滨: 东北林业大学, 2019. [8] NAGAMINE K, HASE T, NOTOMI T. Accelerated reaction by loop-mediated isothermal amplification using loop primers[J]. Molecular and Cellular Probes, 2002, 16(3): 223-229. doi: 10.1006/mcpr.2002.0415 [9] NOTOMI T, OKAYAMA H, MASUBUCHI H,et al. Loop-mediated isothermal amplification of DNA[J]. Nucleic Acids Research, 2000, 28(12): e63. doi: 10.1093/nar/28.12.e63 [10] GOTO M, HONDA E, OGURA A,et al. Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxynaphthol blue[J]. Biotechniques, 2009, 46(3): 167-172. doi: 10.2144/000113072 [11] ZHOU S, HUANG Q, YU M,et al. Rapid visual detection of Aeromonas salmonicida by loop-mediated isothermal amplification with hydroxynaphthol blue dye[J]. Journal of Fish Diseases, 2021, 44(12): 1993-2001. doi: 10.1111/jfd.13513 [12] LAI M Y, OOI C H, LAU Y L. Validation of SYBR green I based closed-tube loop-mediated isothermal amplification (LAMP) assay for diagnosis of knowlesi malaria[J]. Malaria Journal, 2021, 20(1): 1-6. doi: 10.1186/s12936-020-03550-9 [13] LIU H, YU H, PENG Z Q,et al. Visual and rapid detection of Plesiomonas shigelloides using loop-mediated isothermal amplification method[J]. Letters in Applied Microbiology, 2019, 69(6): 411-416. doi: 10.1111/lam.13225 [14] WONG Y, OTHMAN S, LAU Y,et al. Loop-mediated isothermal amplification (LAMP): A versatile technique for detection of micro-organisms[J]. Journal of Applied Microbiology, 2018, 124(3): 626-643. doi: 10.1111/jam.13647 [15] 连文旭, 王 萌, 张 宇, 等. 橡胶树胶孢炭疽菌复合群lamp检测方法的建立及应用[J]. 植物病理学报, 2022, 52(5):841-848. [16] 纪 睿, 曾丹丹, 廖太林, 等. 基于环介导等温扩增技术检测雪松疫霉根腐病菌[J]. 植物检疫, 2017, 31(1):42-47. [17] SAITO Y, MATSUI A, MICHIYUKI S,et al. Loop-Mediated isothermal amplification as Point-of-Care testing for EGFR-Mutated lung adenocarcinoma[J]. Micromachines, 2022, 13(6): 897. doi: 10.3390/mi13060897 [18] ZHU H H, LI Y, WU L X,et al. Internal heating method of loop-mediated isothermal amplification for detection of HPV-6 DNA[J]. Mikrochimica Acta, 2022, 189(5): 212. doi: 10.1007/s00604-022-05283-9 [19] YASUHARA-BELL J, MARRERO G, De SILVA A,et al. Specific detection of Pectobacterium carotovorum by loop-mediated isothermal amplification[J]. Molecular Plant Pathology, 2016, 17(9): 1499-1505. doi: 10.1111/mpp.12378 [20] REN J, ZHU Y, RAN C,et al. Development of a reverse transcription loop-mediated isothermal amplification assay for rapid detection of strawberry crinkle virus[J]. Journal of Phytopathology, 2021, 169(11-12): 733-739. doi: 10.1111/jph.13045 [21] CHEN Y, LI H, YANG L,et al. Rapid detection of Clostridium botulinum in food using Loop-Mediated isothermal amplification (LAMP)[J]. International Journal of Environmental Research and Public Health, 2021, 18(9): 4401. doi: 10.3390/ijerph18094401 [22] SHANG Y, YE Q, CAI S,et al. Loop-mediated isothermal amplification (LAMP) for rapid detection of Salmonella in foods based on new molecular targets[J]. LWT-food science and tecnology, 2021, 142: 110999. doi: 10.1016/j.lwt.2021.110999 [23] 陆红艳. 五种杨树病害病原的lamp检测技术研究[D]. 泰安: 山东农业大学, 2019. [24] 廖 红, 陆 剑, 刘选明, 等. Mn2 +驱动的不同长度DNA片段PCR随机突变研究[J]. 湖南大学学报(自然科学版), 2013, 40(6):92-95. [25] 徐敬潇. 印第安纳沙门菌快速检测方法的建立[D]. 北京: 中国农业科学院, 2020. [26] 余仲东, 高爱琴, 曹支敏. 锈菌夏孢子DNA的微量快速提取方法[J]. 生物技术通讯, 2005, 16(1):48-50. -

 点击查看大图
点击查看大图
图(6) / 表(5)
计量
- 文章访问数: 2123
- HTML全文浏览量: 1117
- PDF下载量: 69
- 被引次数: 0
