


-
丛枝菌根真菌(Arbuscular mycorrhizal fungi, AMF)能够增强植物根系对养分的获取能力,而植物为其提供生长所需的碳源,两者形成互惠共生的丛枝菌根[1]。研究表明,AMF能与90%的维管植物形成丛枝菌根,其中AMF侵染率和孢子密度被用作表征共生关系的紧密程度[2-3]。AMF的定殖和产孢与营养元素和地下生物量存在密切的关系[4-5]。此外,AMF侵染率和孢子密度也可以一定程度反映AMF对生存策略的选择,对了解寄主植物的生长发育状况具有重要意义。以往研究主要集中在农田生态系统中,如冬小麦(Triticum aestivum Linn.)、蚕豆(Vicia faba Linn.)等[6-8]。由于森林根系较复杂,目前对森林生态系统的研究较少。
毛竹(Phyllostachys edulis (Carrière) J. Houz.)是我国亚热带地区分布最为广泛的竹种,其面积为467.8万hm2,占我国竹林面积的73.0%[9],且有不断增加的趋势[10]。毛竹具有独特复杂的根鞭系统,根冠比(0.37~0.50)远高于其他森林生态系统[11],这可能导致毛竹林土壤AMF特征与其他森林存在差异。因此,亟需加强对毛竹林土壤AMF特征的研究。此外,毛竹林分布区是我国氮沉降最严重的地区之一(30.7 kg·hm−2·yr−1)[12],大量的氮输入会导致土壤pH、有机质含量和C:N降低[13],势必会影响AMF的特征。然而,关于氮添加对AMF侵染率和孢子密度影响的研究存在争议[6,14-15]。例如,陈良华等[14]发现施氮显著降低了杉木(Cunninghamia lanceolata (Lamb.) Hook.)AMF侵染率,而汪鹞雄等[15]发现氮添加会显著提高杉木AMF侵染率,Wang等[6]发现长期施氮显著降低了玉米(Zea mays Linn.)和小麦的AMF侵染率和孢子密度。目前,氮添加对毛竹林AMF的影响知之甚少。
生物炭是由生物质在缺氧条件下热裂解而成的芳香类化合物[16],具有多孔道、表面积巨大的特性[17]。生物炭作为一种土壤添加剂,已被广泛用于农业生态系统中。研究表明,施用生物炭提高了土壤的孔隙度[18]、pH[19]和含水率[20],同时增加土壤中营养物质的含量,尤其是氮、磷含量[21]。王策等[22]研究发现添加生物炭可以有效提高土壤中有机质含量,改善土壤团粒结构和微生物群落结构。然而,在大气氮沉降导致的土壤酸化和养分失衡背景下,添加生物炭对毛竹林土壤AMF的影响及其调控机制尚不清楚,相关研究亟待开展。本文以毛竹林为研究对象,探讨了不同水平的氮和生物炭添加对毛竹林AMF侵染率和孢子密度以及土壤理化性质的影响,以期从AMF共生关系角度为全球变化背景下毛竹林的可持续经营提供科学参考。
-
本研究样地位于浙江省杭州市临安区毛竹林长期氮沉降实验平台(29°56′ N, 118°51′ E)。该地区为亚热带季风气候,雨热充沛,四季温和湿润,地貌主要以低山丘陵为主,土壤类型为酸性黄壤。年平均气温为15.6 ℃,年平均降水量为1 420.0 mm,全年无霜期230 d左右,年均日照1 847 h。20世纪70年代,毛竹林样地由常绿阔叶林改造而成,胸径为10.16~10.66 cm,立竹密度为3 000~4 500 株·hm−2。林下植被共有11种,以早开堇菜(Viola prionantha Linn.)为主,覆盖率为5%,生物量为14.6 kg·ha−1。经营措施为除草、伐竹、挖笋、施肥(尿素450 kg·ha−1、过磷酸钙450 kg·ha−1、氯化钾150 kg·ha−1)[11,23-24]。毛竹林样地表层土壤pH为4.45,土壤密度为0.97 g·cm−3,土壤全氮为1.16 g·kg−1,土壤全磷为0.55 g·kg−1。
-
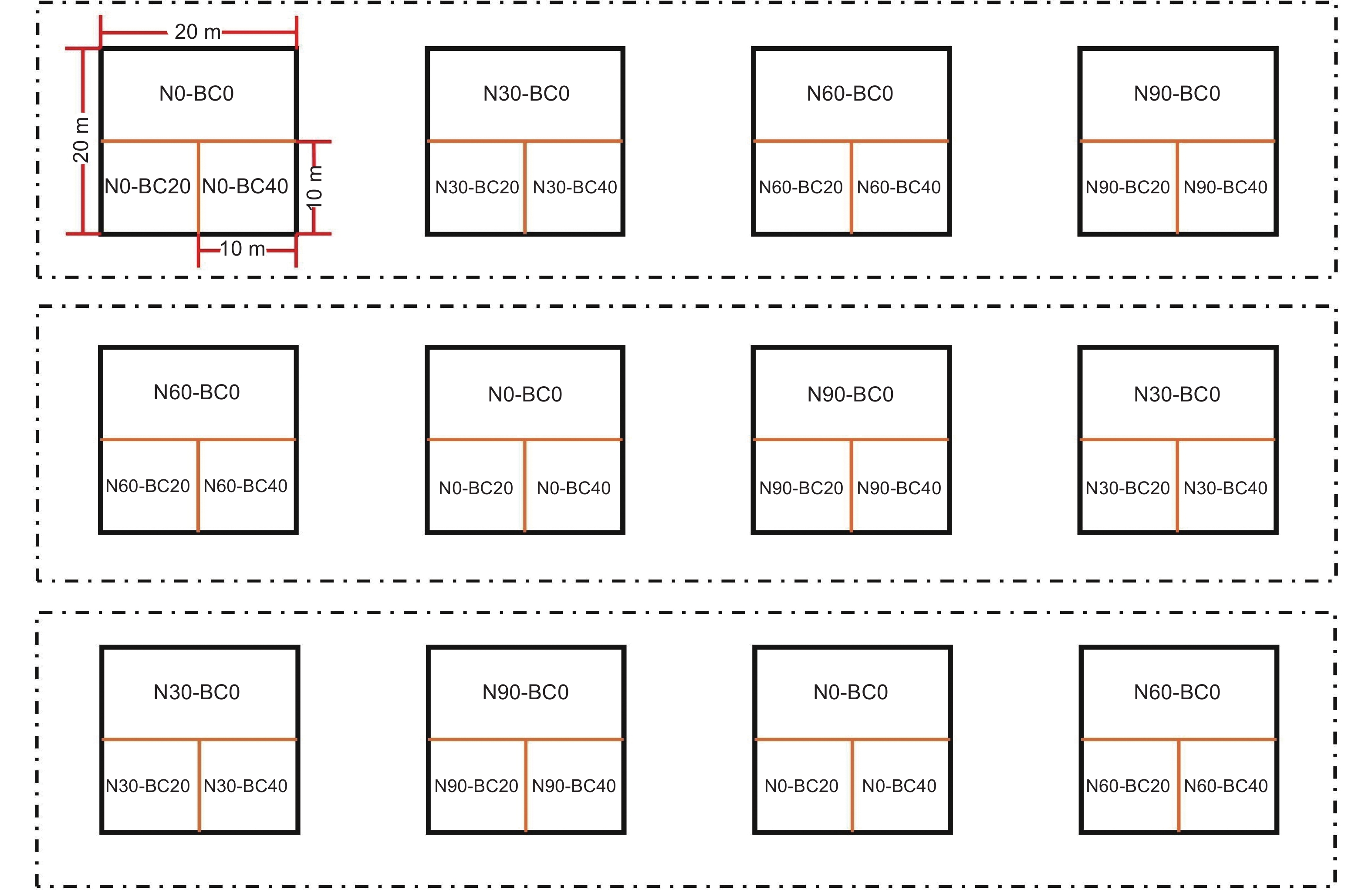
2012年11月,建立12个林分条件和环境状况相似的代表性样方(20 m × 20 m),随机区组排列,样方间设置20 m以上的缓冲区。参考莫江明研究团队氮沉降模拟的试验方法[25],根据我国亚热带地区实际大气氮沉降水平(30.7 kg·hm−2·yr−1)和未来增长趋势[24],采取倍增法设置4个梯度水平:对照(N0:0 kg·hm−2·yr−1)、低氮(N30:30 kg·hm−2·yr−1)、中氮(N60:60 kg·hm−2·yr−1)、高氮(N90:90 kg·hm−2·yr−1)。从2013年1月开始,每月配置定量硝酸铵(NH4NO3)溶液,对样地进行模拟氮沉降。同时为了避免因水分条件不同所产生的差异[26],向对照组喷洒等量自来水,每个水平重复3次。在2014年9月(模拟氮沉降21个月)后,根据前人研究[27],在每个样地内建立1个20 m × 10 m次样地和2个10 m × 10 m次样地,分别依次添加BC0(0 t·hm−2)、BC20(20 t·hm−2)、BC40(40 t·hm−2)生物炭后均匀翻耕(深度为30 cm)(图1)。生物炭由毛竹碎片在600 ℃温度下热解生成,主要理化特征如下:pH(H2O)=9.67;体积密度为0.53 g·cm−3;阳离子交换量(CEC)= 14.9 cmol·kg−1;碳含量=81.73%;氮含量=0.57%;C:N 比=143.4[17]。受疫情影响,2020年1月至5月未喷氮。
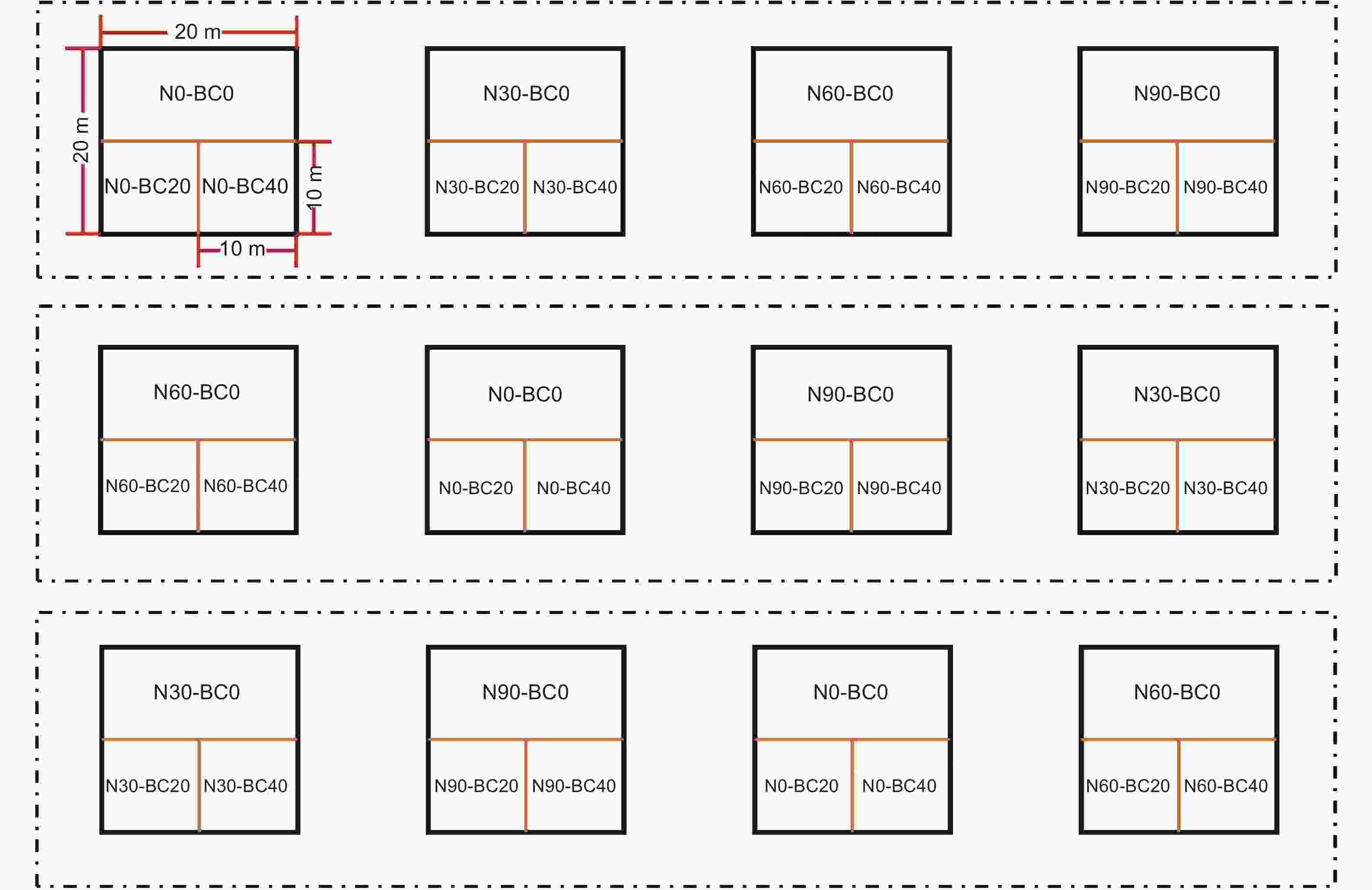
图 1 试验样地布设
Figure 1. Layout of experiment site
-
2020年5月,采用土钻法在每个样地0~20 cm土层随机取10钻土,采集毛竹根际土和根系样品。用抖根法收集根系表面的土壤,将根系和根际土样品分别置于无菌袋内用4 ℃便携式恒温箱带回实验室。剔除土壤样品中的石块、枯叶等杂物后,过2 mm土筛,一部分鲜土放置在4 ℃冰箱内低温保存,用于测定土壤微生物量碳(MBC)、可溶性有机氮(DON)和含水率(SWC),另一部分自然风干后测定AMF孢子密度和其他土壤理化性质。根系样品用于测定AMF侵染率。
采用改良的酸性品红染色法染色后用网格交叉法测定AMF侵染率[28];采用湿筛倾析-蔗糖离心法测定孢子密度[29]。采用氯仿熏蒸-K2SO4浸提法测定MBC含量[30];采用TOC分析仪测定DON含量;采用烘干法测定SWC [31];土壤pH测定采用1:2.5土水质量比,使用pH计(FE20;Mettler Toledo,Switzerland)测定[32];土壤有效氮(AN)含量经过碱性溶液水解后用扩散法测定[33];采用盐酸氟化胺浸提-钼锑抗比色法测定土壤有效磷(AP)含量[17]。
-
利用SPSS 25软件进行统计分析。利用单因素方差分析(One-way ANOVA)和最小显著差法(LSD)分析不同处理下土壤理化性质、AMF侵染率和孢子密度的差异显著性。可重复双因素方差分析氮和生物炭添加及其交互作用对土壤理化性质、AMF侵染率和孢子密度的影响。利用Pearson相关系数分析土壤理化性质与AMF侵染率和孢子密度的相关性。图由Origin 2019软件制作。图表中数据为平均值 ± 标准差。
-
与对照相比,氮添加显著提高了土壤微生物量碳(38.5%~110.3%)、有效磷(75.2%~144.5%)和有效氮(27.9%~66.7%)含量,显著降低了土壤含水率(5.6%~15.2%)和pH(7.6%~11.0%)(P < 0.05,表1)。生物炭添加(BC20和BC40)显著提高了土壤pH和有效磷含量,显著降低了可溶性有机氮、微生物量碳和有效氮含量(P < 0.05)。在氮添加处理下,生物炭添加显著提高了土壤含水率(2.6%~18.5%)和pH(2.6%~11.0%),显著降低了土壤有效氮(2.4%~22.5%)含量(P < 0.05)。可重复双因素方差分析表明氮和生物炭添加及其交互作用均显著影响土壤含水率、pH、可溶性有机氮、微生物量碳、有效磷和有效氮(表2,P < 0.05)。
表 1 不同氮和生物炭添加处理下毛竹林的土壤特性
Table 1. Soil properties under different N and biochar addition treatments in the Moso bamboo plantations
处理
Treatment含水率
SWC/%酸碱度
pH可溶性有机氮
DON/(mg·kg−1)微生物量碳
MBC/(mg·kg−1)有效磷
AP/(mg·kg−1)有效氮
AN/(mg·kg−1)N0-BC0 32.68±1.77 Aa 4.62±0.02 Ab 42.07±0.81 Aa 554.18±8.19 Da 7.05±0.16 Dc 54.87±2.34 Ca N0-BC20 28.66±0.07 b 4.75±0.04 a 30.71±0.56 c 518.10±14.59 b 12.11±0.96 b 49.84±2.03 b N0-BC40 32.03±0.16 a 4.79±0.05 a 34.84±0.69 b 514.55±6.25 b 17.1±0.12 a 49.75±1.20 b N30-BC0 27.7±0.4 Cb 4.27±0.02 Bb 30.54±1.38 Ca 1 165.48±1.82 Aa 14.98±0.61 Bb 70.19±7.41 Ba N30-BC20 32.82±1.63 a 4.36±0.02 b 33.39±1.4 a 1 165.19±5.13 a 17.79±0.89 a 68.53±1.08 ab N30-BC40 30.82±0.75 a 4.52±0.09 a 32.28±2.21 a 944.24±16.77 b 12.22±1.83 c 60.67±2.12 b N60-BC0 30.84±0.23 Bb 4.19±0.07 Cb 40.39±0.56 Aab 971.07±23.59 Bb 12.35±0.59 Cc 77.67±2.24 Ba N60-BC20 32.56±0.2 a 4.31±0.07 a 37.13±3.59 b 1 074.50±29.27 a 16.53±0.24 a 64.15±2.23 b N60-BC40 33.25±0.6 a 4.30±0.03 ab 42.84±0.74 a 955.33±8.37 b 13.95±0.24 b 63.6±2.84 b N90-BC0 30.39±0.5 Bb 4.11±0.02 Dc 37.04±1.66 Ba 767.8±26.62 Ca 17.24±0.32 Aa 91.46±2.43 Aa N90-BC20 31.96±0.49 a 4.19±0.01 b 35.86±0.93 a 747.54±27.41 a 16.77±0.53 a 83.94±4.08 b N90-BC40 31.19±1.03 ab 4.23±0.02 a 31.16±1.98 b 598.9±11.49 b 14.56±2.61 a 70.88±1.27 c 注:不同的大写字母表示在无生物炭添加处理下,不同氮添加处理之间存在显著差异(P < 0.05)。不同的小写字母表示在相同的氮添加处理下,不同生物炭添加处理之间存在显著差异(P < 0.05)
Notes: Different capital letters indicate significant differences among different N treatments without biochar addition treatments (P < 0.05). Different lowercase letters indicate significant differences among different biochar addition treatments under the same N addition treatments (P < 0.05)表 2 氮和生物炭添加对毛竹林土壤理化性质、AMF侵染率和孢子密度影响的双因素方差分析
Table 2. Two-way ANOVA of effects of N and biochar addition on soil properties, colonization rate and spore density of AMF in the Moso bamboo plantations
差异源
Source of
Variables氮添加
N addition生物炭
Biochar
addition交互作用
InteractionF P F P F P SWC 6.778 0.002 9.377 0.001 15.816 0.000 pH 259.735 0.000 41.902 0.000 2.606 0.043 DON 38.440 0.000 12.541 0.000 15.959 0.000 MBC 1 979.948 0.000 179.365 0.000 35.666 0.000 AP 24.463 0.000 22.993 0.000 28.372 0.000 AN 148.885 0.000 48.265 0.000 5.668 0.002 侵染率
Colonization rate30.763 0.000 4.319 0.025 19.014 0.000 孢子密度
Spore density20.873 0.000 11.595 0.000 12.888 0.000 -
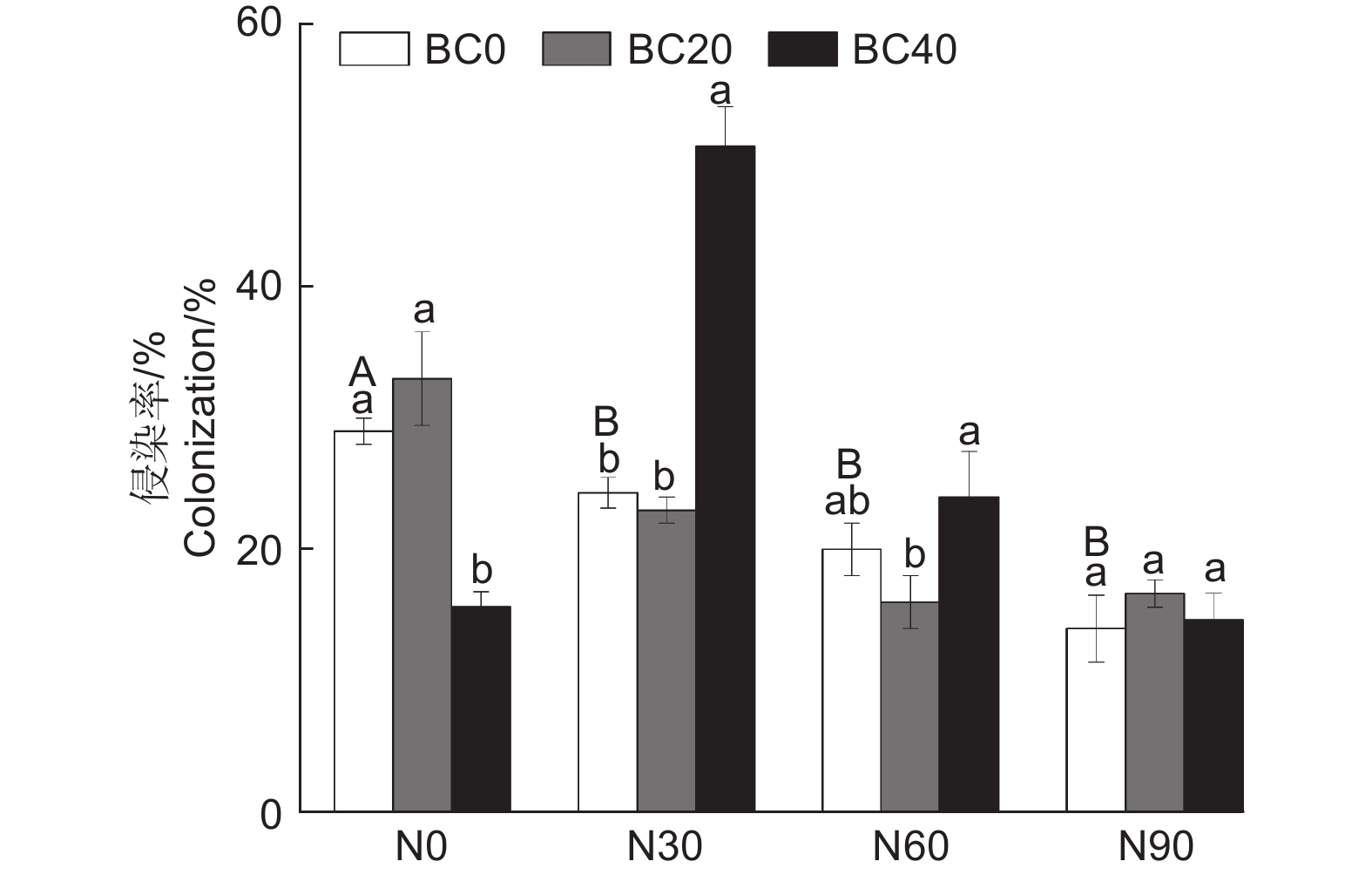
与对照相比,氮和高生物炭添加均显著降低了AMF侵染率(P < 0.05,图2)。在N30处理下,BC40处理显著提高了AMF侵染率(108.2%,P < 0.05)。在N60和N90处理下,BC20和BC40处理对AMF侵染率没有显著影响(P > 0.05)。可重复双因素方差分析表明氮和生物炭添加及其交互作用均显著影响毛竹林土壤AMF侵染率(表2,P < 0.05)。
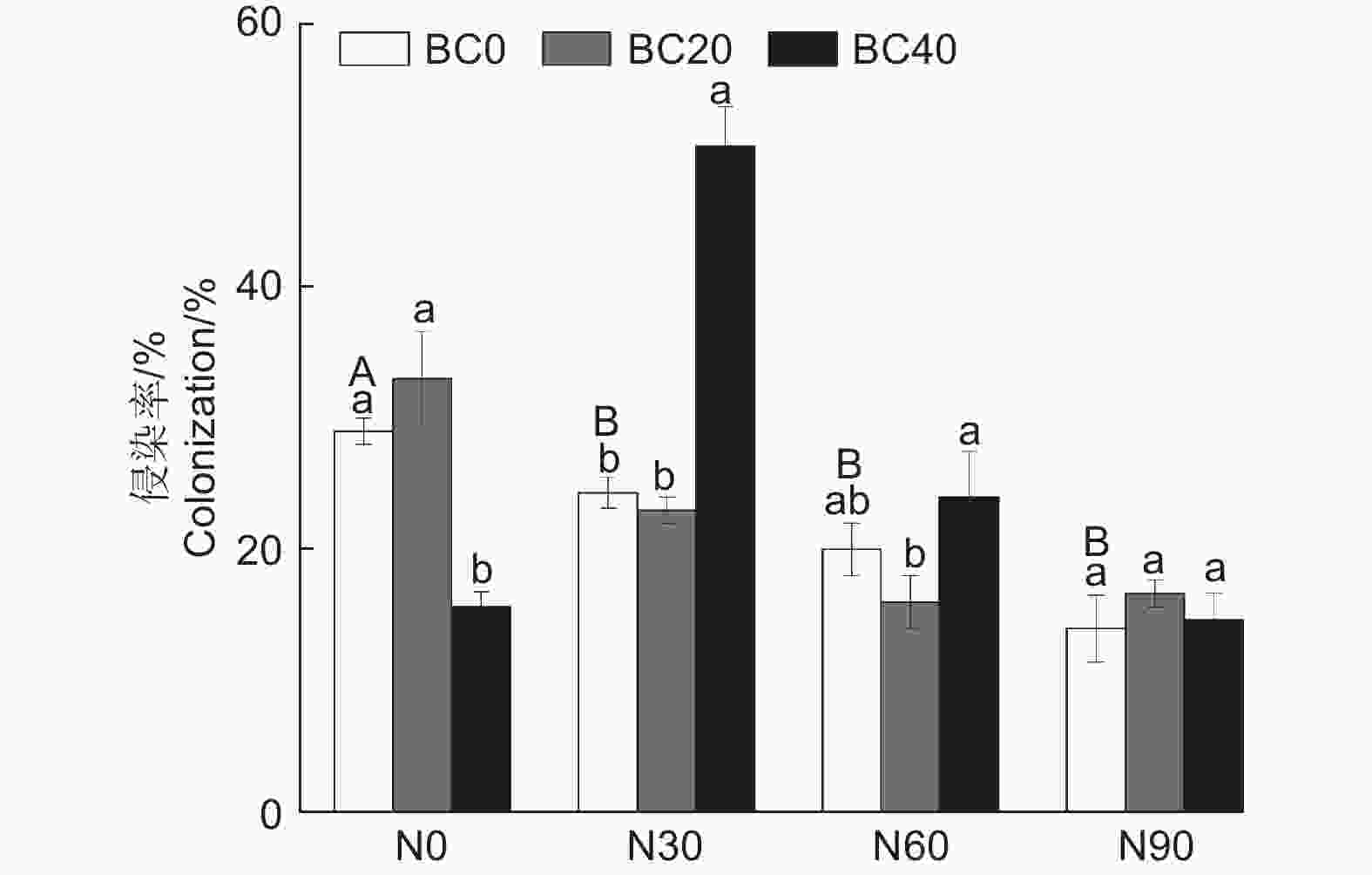
图 2 不同氮和生物炭添加对毛竹林土壤AMF侵染率的影响
Figure 2. Effects of N and biochar addition on colonization rate of AMF in the Moso bamboo plantations
-
与对照相比,氮和生物炭添加提高了AMF孢子密度,尤其是N30和BC40处理, 达到了统计意义上的显著性水平(P < 0.05,图3)。在N30处理下,BC20处理显著提高了孢子密度(23.7%,P < 0.05)。在N60和N90处理下,BC20和BC40处理对孢子密度没有显著影响(P > 0.05)。可重复双因素方差分析表明氮和生物炭添加及其交互作用均极显著影响毛竹林土壤AMF孢子密度(表2,P < 0.01)。

图 3 不同氮和生物炭添加对毛竹林土壤AMF孢子密度的影响
Figure 3. Effects of N and biochar addition on spore density of AMF in the Moso bamboo plantations
-
Pearson相关性分析表明,AMF侵染率与土壤pH呈极显著正相关关系(P < 0.01),与AP和AN呈显著负相关关系(P < 0.05);孢子密度与pH和AP呈极显著正相关关系(P < 0.01),与可溶性有机氮(DON)呈显著负相关关系(P < 0.05)(表3)。
表 3 毛竹林土壤AMF侵染率、孢子密度与土壤理化特征的相关性分析
Table 3. Correlations between colonization rate and spore density of AMF and soil properties
SWC pH DON MBC AP AN 侵染率
Colonization rate−0.208 0.429** −0.207 0.067 −0.478** −0.403* 孢子密度
Spore density0.037 0.373** −0.358* 0.229 0.461** −0.286 注/Notes:*: P < 0.05; **: P < 0.01 -
本研究发现,自然状态下毛竹林土壤AMF侵染率为29.0%,孢子密度为2.0个·g−1。在夏季,牛利敏[34]调查发现毛竹AMF侵染率为46.8%,孢子密度为3.3个·g−1。这表明毛竹AMF侵染率和孢子密度存在季节变化。研究表明,与其他季节相比,夏季的AMF的侵染率和孢子密度最高[35-36]。这可能是因为夏季是植物的生长季,植物需要从土壤中获取大量的营养物质,从而导致根系与AMF的共生关系更加紧密。此外,研究发现柠条(Caragana korshinskii kom)、杉木和水杉(Metasequoia glyptostroboides Hu & W. C. Cheng)的AMF侵染率分别为92.4%[37]、78.9%[38]和70.0%[39]。上述结果说明毛竹AMF侵染率低于其他森林类型,这可能是与毛竹林有独特发达的根鞭系统从而对AMF的依赖较小有关。
氮添加抑制了AMF侵染率,且随着氮添加水平增加这种抑制作用变强,这与陈良华[14]的研究结果相似。黄园园等[40]对高寒草甸植物的研究,表明随着土壤有效氮含量的增加,植物吸收氮素的成本降低,对AMF的依赖性减弱,从而导致AMF侵染率降低。有研究也发现施氮导致土壤有效氮含量升高,土壤中的NO3−被还原为NO2−并在根系附近积累后对宿主植物造成伤害,进而降低AMF侵染率[41]。在本研究中,土壤AN的含量与AMF侵染率呈显著负相关关系(表3),且氮添加显著增加了土壤AN的含量(表1),支持了上述观点。此外,我们发现土壤pH与AMF侵染率呈显著正相关关系。氮添加通过降低土壤pH,从而抑制了AMF侵染率,原因可能是低的pH抑制了AMF和根系的活性。研究表明,长期氮添加会改变微生物群落结构,减少土壤真菌生物量[42],这也可能是AMF侵染率下降的原因。
本研究中,N30处理显著增加了孢子密度。这与马俊卿等[3]对多种C3、C4植物的研究结论不一致。这可能是因为宿主不同,其根系的AMF种类和群落结构也不完全一致。不同的AMF种类对外界环境的响应不同,在孢子萌发和侵染根系等方面所需的环境因素条件也有所差异[37]。本试验中N30处理下土壤AP显著增加,削弱了植物对AMF的依赖性,植物因而减少对AMF的碳供应,侵染率下降。而孢子是AMF的一种繁殖和幸存体,同时也是重要的磷库[5],此时产生更多的孢子可能是AMF对生存策略的选择[37]。在本研究中,土壤AP的含量与AMF侵染率呈显著负相关关系,与孢子密度呈显著正相关关系(表3),且氮添加显著增加了土壤AP的含量(表1),支持了上述观点。然而在N60和N90梯度下,孢子密度的降低可以抑制真菌的无性繁殖,降低能量消耗,有助于AMF在逆境中生存[43]。
-
生物炭(40 t·hm−2)添加显著降低了AMF侵染率,但提高了孢子密度,这与高粱(Sorghum bicolor (L.) Moench)[44]和辣椒(Capsicum annuum Linn.)[45]对生物炭添加的响应相似。原因是生物炭添加增加了土壤AP(表1),毛竹吸收磷元素的成本降低,从而使其对AMF的依赖性减弱[40],导致AMF侵染率降低。研究发现生物炭较大的孔隙度和表面积增加了土壤孔隙度,改善了土壤酸度[22]和微生物群落结构[46],促进了AMF的繁殖。然而,Zhang等[47]对冬小麦施用生物炭的研究结果表明添加生物炭可以增加AMF的生长繁殖,提高侵染率和孢子密度,并且随着用量增加,效果更明显。AMF侵染率和孢子密度对生物炭添加的响应不一致的原因可能是生物炭施加量、植物类型、季节等不同。
在氮添加处理下,生物炭添加提高了AMF侵染率和孢子密度,主要归因于氮和生物炭的复合作用显著降低了土壤AN含量(表1),从而导致了AMF侵染率增加。此外,氮和生物炭添加的复合作用促进了毛竹的光合速率,提高了生物量[48],导致了大量的光合产物转移到根系,进而为AMF提供了更多的碳水化合物,促进AMF的繁殖,产生了更多的孢子[49-51]。
-
氮和生物炭添加提高了土壤养分含量,降低了毛竹AMF侵染率,增加了孢子密度,削弱了毛竹与AMF的共生关系。在氮沉降背景下,生物炭添加提高了AMF侵染率,增强了毛竹与AMF的共生关系紧密程度,表明添加生物炭可以缓解氮沉降对AMF定殖和产孢造成的负面影响,为应对氮沉降背景下亚热带毛竹林可持续经营提供参考和科学依据。在将来的研究中我们可以探讨AMF定殖和产孢特性以及群落结构的年际变化,以便于更好地理解氮和生物炭添加背景下毛竹与AMF的共生关系及其调控机制。
氮和生物炭添加对毛竹林AMF侵染率和孢子密度的影响
Effects of Nitrogen and Biochar Addition on Arbuscular Mycorrhizal Fungi Colonization Rate and Spore Density in Moso Bamboo Plantations
-
摘要:
目的 探讨氮沉降和生物炭添加对毛竹林丛枝菌根真菌特征的影响,为全球变化背景下毛竹林的可持续经营提供科学参考。 方法 以毛竹林为研究对象,探讨了不同梯度的氮沉降(0、30、60和90 kg·hm−2·yr−1)和生物炭添加(0、20和40 t ·hm−2)及其复合作用对毛竹林AMF侵染率和孢子密度以及土壤理化性质的影响。 结果 与对照(0 kg·hm−2·yr−1 N + 0 t·hm−2 BC)相比,氮添加显著降低了AMF侵染率(16.1%~51.7%)。高生物炭添加(40 t·hm−2)显著降低了AMF侵染率(46.0%),但提高了孢子密度(162.5%)。在氮添加处理下,生物炭添加提高了AMF侵染率,对孢子密度无显著影响。AMF侵染率与土壤pH呈显著正相关关系,与有效氮和有效磷呈显著负相关关系。AMF孢子密度与有效磷呈显著正相关关系。 结论 在氮添加处理下,生物炭添加提高了AMF侵染率,增强了毛竹与AMF的共生关系,表明在氮沉降背景下生物炭添加是实现毛竹林可持续经营的有效措施之一。 Abstract:Objective To investigate the effects of nitrogen (N) deposition and biochar addition on arbuscular mycorrhizal fungi (AMF) characteristics in Moso bamboo (Phyllostachys edulis (Carrière) J. Houz.) plantations for providing scientific reference for sustainable management of Moso bamboo plantations under global change. Methods We investigated the effects of different N addition rates (0, 30, 60, and 90 kg N·hm−2·yr−1), biochar addition rates (0, 20, and 40 t BC·hm−2), and their combination on the AMF colonization rate, spore density and soil properties of Phyllostachys edulis plantations. Results Compared with the control (0 kg N·hm−2·yr−1 + 0 t BC·hm−2), N addition significantly reduced the colonization rate of AMF (16.1%~51.7%). High biochar addition (40 t BC·hm−2) significantly reduced the colonization rate of AMF (46.0%), but increased the spore density (162.5%). Under the N addition treatments, biochar addition increased the colonization rate of AMF, and had no significant effect on spore density. The colonization rate of AMF was significantly positively correlated with soil pH, and negatively correlated with available N and available phosphorus. The spore density of AMF was significantly positively correlated with available phosphorus. Conclusion Under the N addition treatments, biochar addition increased the colonization rate of AMF and enhanced the symbiotic relationship between Moso bamboo and AMF, indicating that biochar addition is an effective measure to achieve sustainable management of Moso bamboo plantations under the background of N deposition. -
图 2 不同氮和生物炭添加对毛竹林土壤AMF侵染率的影响
Figure 2. Effects of N and biochar addition on colonization rate of AMF in the Moso bamboo plantations
图 3 不同氮和生物炭添加对毛竹林土壤AMF孢子密度的影响
Figure 3. Effects of N and biochar addition on spore density of AMF in the Moso bamboo plantations
表 1 不同氮和生物炭添加处理下毛竹林的土壤特性
Table 1. Soil properties under different N and biochar addition treatments in the Moso bamboo plantations
处理
Treatment含水率
SWC/%酸碱度
pH可溶性有机氮
DON/(mg·kg−1)微生物量碳
MBC/(mg·kg−1)有效磷
AP/(mg·kg−1)有效氮
AN/(mg·kg−1)N0-BC0 32.68±1.77 Aa 4.62±0.02 Ab 42.07±0.81 Aa 554.18±8.19 Da 7.05±0.16 Dc 54.87±2.34 Ca N0-BC20 28.66±0.07 b 4.75±0.04 a 30.71±0.56 c 518.10±14.59 b 12.11±0.96 b 49.84±2.03 b N0-BC40 32.03±0.16 a 4.79±0.05 a 34.84±0.69 b 514.55±6.25 b 17.1±0.12 a 49.75±1.20 b N30-BC0 27.7±0.4 Cb 4.27±0.02 Bb 30.54±1.38 Ca 1 165.48±1.82 Aa 14.98±0.61 Bb 70.19±7.41 Ba N30-BC20 32.82±1.63 a 4.36±0.02 b 33.39±1.4 a 1 165.19±5.13 a 17.79±0.89 a 68.53±1.08 ab N30-BC40 30.82±0.75 a 4.52±0.09 a 32.28±2.21 a 944.24±16.77 b 12.22±1.83 c 60.67±2.12 b N60-BC0 30.84±0.23 Bb 4.19±0.07 Cb 40.39±0.56 Aab 971.07±23.59 Bb 12.35±0.59 Cc 77.67±2.24 Ba N60-BC20 32.56±0.2 a 4.31±0.07 a 37.13±3.59 b 1 074.50±29.27 a 16.53±0.24 a 64.15±2.23 b N60-BC40 33.25±0.6 a 4.30±0.03 ab 42.84±0.74 a 955.33±8.37 b 13.95±0.24 b 63.6±2.84 b N90-BC0 30.39±0.5 Bb 4.11±0.02 Dc 37.04±1.66 Ba 767.8±26.62 Ca 17.24±0.32 Aa 91.46±2.43 Aa N90-BC20 31.96±0.49 a 4.19±0.01 b 35.86±0.93 a 747.54±27.41 a 16.77±0.53 a 83.94±4.08 b N90-BC40 31.19±1.03 ab 4.23±0.02 a 31.16±1.98 b 598.9±11.49 b 14.56±2.61 a 70.88±1.27 c 注:不同的大写字母表示在无生物炭添加处理下,不同氮添加处理之间存在显著差异(P < 0.05)。不同的小写字母表示在相同的氮添加处理下,不同生物炭添加处理之间存在显著差异(P < 0.05)
Notes: Different capital letters indicate significant differences among different N treatments without biochar addition treatments (P < 0.05). Different lowercase letters indicate significant differences among different biochar addition treatments under the same N addition treatments (P < 0.05) 下载: 导出CSV
下载: 导出CSV
表 2 氮和生物炭添加对毛竹林土壤理化性质、AMF侵染率和孢子密度影响的双因素方差分析
Table 2. Two-way ANOVA of effects of N and biochar addition on soil properties, colonization rate and spore density of AMF in the Moso bamboo plantations
差异源
Source of
Variables氮添加
N addition生物炭
Biochar
addition交互作用
InteractionF P F P F P SWC 6.778 0.002 9.377 0.001 15.816 0.000 pH 259.735 0.000 41.902 0.000 2.606 0.043 DON 38.440 0.000 12.541 0.000 15.959 0.000 MBC 1 979.948 0.000 179.365 0.000 35.666 0.000 AP 24.463 0.000 22.993 0.000 28.372 0.000 AN 148.885 0.000 48.265 0.000 5.668 0.002 侵染率
Colonization rate30.763 0.000 4.319 0.025 19.014 0.000 孢子密度
Spore density20.873 0.000 11.595 0.000 12.888 0.000
下载: 导出CSV
表 3 毛竹林土壤AMF侵染率、孢子密度与土壤理化特征的相关性分析
Table 3. Correlations between colonization rate and spore density of AMF and soil properties
SWC pH DON MBC AP AN 侵染率
Colonization rate−0.208 0.429** −0.207 0.067 −0.478** −0.403* 孢子密度
Spore density0.037 0.373** −0.358* 0.229 0.461** −0.286 注/Notes:*: P < 0.05; **: P < 0.01
下载: 导出CSV
-
[1] 柳 威, 吴强盛, 翟华芬, 等. 丛枝菌根真菌与土壤盐碱植物的关系[J]. 北方园艺, 2010(2):226-228. [2] GIOVANNETTI M, MOSSE B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots[J]. New Phytologist, 1980, 84(3): 489-500. doi: 10.1111/j.1469-8137.1980.tb04556.x [3] 马俊卿, 侯 宁, 孙晨瑜, 等. 宿主不同对丛枝菌根真菌扩繁效应的影响[J]. 中国农学通报, 2022, 38(1):7-14. [4] 杨 康, 孙建茹, 王 妍, 等. 入侵植物与本地植物互作对丛枝菌根真菌AMF侵染率的影响[J]. 菌物学报, 2019, 38(11):1938-1947. [5] 任 玮, 杨 韧, 唐 明. 不同培养基质和寄主植物对AMF扩繁的影响[J]. 菌物研究, 2017, 15(1):46-52. doi: 10.13341/j.jfr.2014.1708 [6] WANG M Y, HU L B, WANG W H, et al. Influence of long-term fixed fertilization on diversity of arbuscular mycorrhizal fungi[J]. Pedosphere, 2009, 19(5): 663-672. doi: 10.1016/S1002-0160(09)60161-2 [7] 赵 飞, 唐 明, 张好强. 丛枝菌根真菌对宁夏枸杞生长和氮代谢的影响[J]. 西北林学院学报, 2022, 37(1):166-171,252. [8] CHAMBERS C A, SMITH S E, SMITH F A. Effects of ammonium and nitrate ions on mycorrhizal infection, nodulation and growth of trifolium subterraneum[J]. New Phytologist, 1980, 85(1): 47-62. doi: 10.1111/j.1469-8137.1980.tb04447.x [9] 李玉敏, 冯鹏飞. 基于第九次全国森林资源清查的中国竹资源分析[J]. 世界竹藤通讯, 2019, 17(6):45-48. [10] 程明圣, 邹 娜. 毛竹扩张对森林生态的影响及其管控研究进展[J]. 江汉大学学报(自然科学版), 2021, 49(3):49-55. doi: 10.16389/j.cnki.cn42-1737/n.2021.03.007 [11] SONG X Z, PENG C H, CIAIS P, et al. Nitrogen addition increased CO2 uptake more than non-CO2 greenhouse gases emissions in a Moso bamboo forest[J]. Science Advances, 2020, 6: eaaw5790. doi: 10.1126/sciadv.aaw5790 [12] GALLOWAY J N, TOWNSEND A R, ERISMAN J W, et al. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions[J]. Science, 2008, 320(5878): 889-892. doi: 10.1126/science.1136674 [13] 张维娜, 廖周瑜. 氮沉降增加对森林植物影响的研究进展[J]. 环境科学导刊, 2009, 28(3):21-24. doi: 10.3969/j.issn.1673-9655.2009.03.005 [14] 陈良华, 刘选茹, 李西悦, 等. 杉木细根AMF侵染特征和锰、镉含量对短期氮添加的响应[J]. 四川农业大学学报, 2022, 40(2):1-14. doi: 10.16036/j.issn.1000-2650.202201046 [15] 汪鹞雄, 李 全, 沈益康, 等. 模拟氮沉降对杉木丛枝菌根真菌侵染率和球囊霉素的影响[J]. 生态学报, 2021, 41(1):194-201. [16] JOHANNES L. A handful of carbon[J]. Nature, 2007, 447(7141): 143-144. [17] 李 全. 长期氮和生物炭添加对毛竹林土壤温室气体通量的影响[D]. 杨凌: 西北农林科技大学, 2021. [18] LU X K, MAO Q G, GILLIAM F S, et al. Nitrogen deposition contributes to soil acidification in tropical ecosystems[J]. Global Change Biology, 2014, 20(12): 3790-3801. doi: 10.1111/gcb.12665 [19] CHINTALA R, SCHUMACHER T E, KUMAR S, et al. Molecular characterization of biochars and their influence on microbiological properties of soil[J]. Journal of Hazardous Materials, 2014, 279: 244-256. doi: 10.1016/j.jhazmat.2014.06.074 [20] 屈田华. 氮沉降与生物质炭输入的交互作用对毛竹林土壤N2O通量的影响[D]. 杭州: 浙江农林大学, 2021. [21] WALKER C. Systematics and taxonomy of the arbuscular endomycorrhizal Fungi (glomales)-a possible way forward[J]. Agronomie, 1992, 12(10): 887-897. doi: 10.1051/agro:19921026 [22] 王 策, 郭绍霞, 刘润进, 等. 生物质炭和AM真菌联合施用对连作甜瓜生长和土壤理化性质的影响[J]. 中国瓜菜, 2021, 34(7):29-34. doi: 10.16861/j.cnki.zggc.2021.0170 [23] 崔坤凯. 氮和生物质炭添加对毛竹林土壤生态酶活性及其化学计量特征的影响[D]. 杭州: 浙江农林大学, 2021. [24] SONG X Z, ZHOU G M, GU H H, et al. Management practices amplify the effects of N deposition on leaf litter decomposition of the Moso bamboo forest[J]. Plant and Soil, 2015, 395(1): 391-400. [25] MO J M, ZHANG W, ZHU W X, et al. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China[J]. Global Change Biology, 2008, 14(2): 403-412. doi: 10.1111/j.1365-2486.2007.01503.x [26] REAY D S, DENTENER F, SMITH P, et al. Global nitrogen deposition and carbon sinks[J]. Nature Geoscience, 2008, 1(2): 430-437. [27] Sun L Y, Chen Z Z, Wang J Y, et al. Combined effects of nitrogen deposition and biochar application on emissions of N2O, CO2 and NH3 from agricultural and forest soils[J]. Soil Science and Plant Nutrition, 2014, 60(2): 254-265. doi: 10.1080/00380768.2014.885386 [28] KORMANIK P P, BRYAN W C, SCHULTZ R C. Procedures and equipment for staining large numbers of plant root samples for endomycorrhizal assay[J]. Nrc Research Press Ottawa, Canada, 1980, 26(4): 536-538. [29] AN Z Q, HENDRIX J W, HERSHMAN D E, et al. Evaluation of the “most probable number” (MPN) and wet-sieving methods for determining soil-borne populations of endogonaceous mycorrhizal fungi[J]. Mycologia, 2018, 82(5): 576-581. [30] BROOKES P C, LANDMAN A, PRUDEN G, et al. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil[J]. Pergamon, 1985, 17(6): 837-842. [31] Knegt R D, Brink H V D. Improvement of the drying oven method for the determination of the moisture content of milk powder[J]. International Dairy Journal, 1998, 8(8): 733-738. doi: 10.1016/S0958-6946(97)00110-6 [32] PENG X Q, WANG W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of Northern China[J]. Soil Biology and Biochemistry, 2016, 98: 74-84. doi: 10.1016/j.soilbio.2016.04.008 [33] XIAO Y S, PENG Y, PENG F T, et al. Effects of concentrated application of soil conditioners on soil–air permeability and absorption of nitrogen by young peach trees[J]. Soil Science and Plant Nutrition, 2018, 64(3): 423-432. doi: 10.1080/00380768.2018.1439697 [34] 牛利敏. 毛竹入侵及经营方式对土壤丛枝菌根真菌群落的影响及其机理研究[D]. 杭州: 浙江农林大学, 2017. [35] 彭 雪, 韩 涛, 郭健桦, 等. 暖季型草坪草AMF侵染与孢子密度季节动态研究[J]. 生态科学, 2016, 35(6):1-5. doi: 10.14108/j.cnki.1008-8873.2016.06.001 [36] 陈科宇. 增温对不同季节高寒草甸丛枝菌根真菌群落的影响[D]. 成都: 西南民族大学, 2020. [37] 刘永俊, 郑 红, 何 雷, 等. 柠条根系中丛枝菌根真菌的季节性变化及影响因素[J]. 应用生态学报, 2009, 20(5):1085-1091. doi: 10.13287/j.1001-9332.2009.0184 [38] LIN C Y, WANG Y X, LIU M H, et al. Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia Lanceolata)[J]. Scientific Reports, 2020, 10(1): 12260. doi: 10.1038/s41598-020-69213-6 [39] 张 英, 郭良栋, 刘润进. 都江堰地区丛枝菌根真菌多样性与生态研究[J]. 植物生态学报, 2003(4):537-544. [40] 黄园园, OLBRECHT L, 杨晓霞, 等. 养分添加对青藏高原高寒草甸丛枝菌根真菌的影响[J]. 北京大学学报:自然科学版, 2014, 50(5):911-918. [41] HODGE A, CAMPBELL C D, FITTER A H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material[J]. Nature, 2001, 413(6853): 297-299. doi: 10.1038/35095041 [42] 张天安. 全球氮沉降对土壤微生物和凋落物分解的影响以及生态预测中驱动变量的贡献[D]. 南京: 南京林业大学, 2020. [43] 沈益康. 氮磷添加对杉木丛枝菌根真菌和叶片化学计量特征的影响[D]. 杭州: 浙江农林大学, 2021. [44] MARÍA V, PEDRO M, CLARA M, et al. The effects of biochar on indigenous arbuscular mycorrhizae fungi from agroenvironments[J]. Plants, 2021, 10(5): 950. doi: 10.3390/plants10050950 [45] GYÖNGYI B, ANDRÁS M, TÜNDE T, et al. Biochar alters soil physical characteristics, arbuscular mycorrhizal fungi colonization, and glomalin production[J]. Agronomy, 2020, 10(12): 1933-1933. doi: 10.3390/agronomy10121933 [46] 郭康颖. 不同原料和热解温度制备的生物质炭对雷竹林土壤微生物群落结构及功能的影响[D]. 杭州: 浙江农林大学, 2021. [47] ZHANG Q Z, DIJKSTRA F A, LIU X R et al. Effects of biochar on soil microbial biomass after four years of consecutive application in the North China Plain[J]. Plos One, 2017, 9(7): e102062. [48] 刘 宁, 彭春菊, 雷赵枫, 等. 氮沉降和生物质炭对毛竹叶片光合特性的影响[J]. 浙江农林大学学报, 2019, 36(4):704-712. [49] SPERATTI A B, JOHNSON M S, SOUSA H M, et al. Biochars from local agricultural waste residues contribute to soil quality and plant growth in a Cerrado region (Brazil) Arenosol[J]. GCB Bioenergy, 2018, 10(4): 272-286. doi: 10.1111/gcbb.12489 [50] AGEGNEHU G, BASS A M, NELSON P N,, et al. Biochar and biochar-compost as soil amendments: Effects on peanut yield, soil properties and greenhouse gas emissions in tropical North Queensland, Australia[J]. Agriculture, Ecosystems and Environment, 2015, 213: 72-85. doi: 10.1016/j.agee.2015.07.027 [51] TRUPIANO D, COCOZZA C, BARONTI S, et al. The effects of biochar and its combination with compost on lettuce (Lactuca Sativa L.) growth, soil properties, and soil microbial activity and abundance[J]. International Journal of Agronomy, 2017, 2017: 1-12. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3100
- HTML全文浏览量: 993
- PDF下载量: 65
- 被引次数: 0
