


-
杉木(Cunninghamia lanceolate (Lamb.) Hook.)是我国南方特有的优良速生用材树种,据国家林业和草原局数据显示,杉木人工林种植面积达990.20万hm2[1]。为满足市场需求,多代连栽、大面积单一种植杉木人工林成为南方红壤区的普遍现象,并且杉木种植区气候属于亚热带季风气候,降雨丰沛,雨季土壤含水率高,土壤氧化还原反应剧烈。在各种因素影响下杉木种植区产生地力衰退、生产力下降等问题[2-3]。其中土壤有效磷素亏缺是杉木林地力衰退的重要因素[4]。
杉木种植区土壤以酸性红壤为主,酸性红壤中最稳定和最易被生物利用的正磷酸盐和次级磷酸盐离子会通过化学作用吸附在红壤矿物表面,形成铁磷酸盐沉淀,或者被Fe2O3胶膜包被形成闭蓄态磷,无法直接供植物利用[5]。土壤磷素有多种形态,在不同的土壤环境下能够相互转化[6]。张虹等[7]研究发现,闭蓄态磷在杉木人工林红壤中占比大于47.1%。土壤铁磷转化受环境影响较大,在好氧环境下Fe(III)通过吸附或沉淀对磷酸盐阴离子有高亲和力,但在嫌气条件下铁磷酸盐与闭蓄态磷能够随Fe(III)矿物的还原而溶解[8-9]。铁磷转化同样受微生物影响,有研究发现,土壤中存在一些电子穿梭体(腐殖酸、生物炭等)能不断地把电子从微生物传递到Fe(III)氧化物[10],加速微生物对Fe(III)氧化物的还原,从而提高土壤闭蓄态铁磷的释放[11-12]。但在降雨充沛条件下土壤微生物影响土壤铁磷转化的研究不够深入,因此探究厌氧条件下微生物对有效磷素的转化是有必要的。
生物炭具有储存碳和改良土壤养分有效性的作用,其多孔特性能吸附土壤磷素,降低土壤磷素有效性[13],然而生物炭又能使吸附在铁铝氧化物上的磷素溶解,提高土壤磷素有效性[14]。Zhou等[15]发现,生物炭能明显促进红壤闭蓄态铁磷向有效态磷转化。令狐荣云等[16]发现,向土壤中添加铁还原菌能够促进土壤Fe(III)的还原,释放闭蓄态磷和铁铝磷中的有效磷素。但目前对生物炭如何促进Fe-P等难溶态磷向有效态磷转化机制还不清楚,因此探究南方红壤难溶性铁磷有效化利用是有必要的。
鉴于此,本文以亚热带杉木人工林红壤为对象,模拟研究在降雨丰沛、红壤水分长期饱和条件下生物炭对红壤铁还原、磷形态转化的影响,分析红壤铁还原菌、解磷菌群落结构变化规律,旨在明确生物炭对红壤铁还原的影响与磷形态转化的关系。
-
供试土壤采集于福建省南平市延平区(26°39′ N,117°54′ E),属于亚热带季风气候,年平均气温19.3 ℃,年平均降水量1 669 mm,相对湿度83%。选择由花岗岩发育而来的杉木二代人工林林下表层红壤(0~20 cm)为供试土壤,土层厚度超过1.0 m[17]。除去采集土样中的石块、根系等杂物,过2 mm尼龙筛后放入−80 ℃冰箱备用。
-
叶片生物炭在调节土壤养分和微生物群落结构方面优于木屑炭[15],所以本研究选择杉木叶凋落物作为生物炭的原料。杉木叶洗净滤干后于60 ℃烘箱中烘干至质量恒定,然后粉碎过100目筛,得到制备生物炭的原材料。取过筛杉木叶装入带盖陶瓷坩埚,在马弗炉中以15 ℃·min−1的速率分别升温至目标温度(300 ℃和500 ℃),保持在高温缺氧条件下炭化4 h,将生物炭按照热解温度不同得到300 ℃生物炭(B300)和500 ℃生物炭(B500)。所得生物炭B300与B500的灰分含量分别为12.03%、21.74%,pH值分别为6.69、8.47,全碳含量分别为546.4、572.6 g·kg−1,全氮含量分别为19.75、15.77 g·kg−1,全磷含量分别为0.84、1.49 g·kg−1。
-
以红壤为基础均匀添加B300和B500生物炭,设置0、1%、3%3种梯度。将混匀土样放入聚丙烯塑料盆中,保持70%的田间持水量,设置淹水处理进行对照试验,进行为期40 d的室内培养。共10个处理组,每组3个重复,具体如下:(1)红壤保持70%的田间持水量(N-CK)、(2)红壤加1% 300 ℃生物炭,保持70%的田间持水量(N-1%B300)、(3)红壤加3% 300 ℃生物炭,保持70%的田间持水量(N-3%B300)、(4)红壤加1% 500 ℃生物炭,保持70%的田间持水量(N-1%B500)、(5)红壤加3% 500 ℃裂解的生物炭,保持70%的田间持水量(N-3%B500)、(6)红壤保持淹水状态(D-CK)、(7)红壤加1% 300 ℃裂解的生物炭,保持淹水状态(D-1%B300)、(8)红壤加3% 300 ℃裂解的生物炭,保持淹水状态(D-3%B300)、(9)红壤加1% 500 ℃裂解的生物炭,保持淹水状态(D-1%B500)、(10)红壤加3% 500 ℃裂解的生物炭,保持淹水状态(D-3%B500)。
-
土壤pH值[18]使用pH计(STARTER 2100,美国)测定、土壤全碳全氮采用全自动元素分析仪(Vario Macro Cube,Vario Macro Cubeelementar,德国)测定、土壤全钾采用电感耦合等离子体发射光谱仪(ICP-OES,ICAP6000 Radial,Thermo,英国)测定、土壤全磷采用钼锑抗比色法测定、土壤亚铁离子采用邻啡啰啉比色法测定、分级磷采用修正后的Hedley[19]方法,其中H2O表示水溶态、NaHCO3表示碳酸氢钠态、NaOH表示氢氧化钠态、HCl表示盐酸态、Residual-P表示残渣态磷、TP表示全磷、Po表示有机磷、Pi表示无机磷。土壤活性磷由水溶态磷和碳酸氢钠态磷组成,土壤中等活性磷由与土壤铁铝化合物化学吸附及粘粒结合的无机磷(NaOH-Pi)、与土壤腐殖酸结合的有机磷(NaOH-Po)组成,土壤稳定态磷由盐酸态磷和残渣态磷组成。
-
本研究选取处理组N-CK、N-3%B300、N-3%B500、D-CK、D-3%B300和D-3%B500检测土壤铁还原菌与解磷菌。用绝对定量方法检测样品中铁还原菌基因拷贝数。利用高通量测序技术测定土壤解磷菌群落数量和组成。
-
利用SPSS 25.0对红壤化学性质、解磷菌与铁还原菌数据进行单因素(ANOVA)方差分析,选择LSD法显著性检验;通过Alpha多样性分析反映解磷菌群落的丰富度和多样性;基于Bray-Curtis 距离矩阵的非度量多维尺度分析(NMDS)确定解磷菌群落β多样性差异;使用R语言“random Forest”包进行随机森林模型分析,使用“plspm”包进行偏最小二乘法路径模型(PLS-PM)分析。
-
淹水条件下施用生物炭显著影响了土壤磷形态(P<0.05,图1)。淹水处理后土壤活性磷即水溶态磷与碳酸氢钠提取态磷含量增加,其中随生物炭添加量的增加H2O-Pi与NaHCO3-Pi含量减少,H2O-Po与NaHCO3-Po含量增加;NaHCO3-Pi含量随生物炭制备温度的升高而减少(图1AB)。淹水处理后土壤NaOH-Pi含量减少,NaOH-Po含量整体上增加,非淹水条件下施加不同温度制备的生物炭均增加了NaOH-Pi含量,但NaOH-Po含量却随B500添加量的增加呈下降趋势(图1C)。淹水后盐酸态磷含量上升,非淹水处理下添加生物炭提高了HCl-Pi含量,但不同处理组之间无显著差异,淹水处理的HCl-Po含量显著高于对照组(图1D)。淹水处理后残渣态磷含量随生物炭的添加而下降(图1E),但不显著。添加生物炭后,土壤全磷含量随生物炭制备温度与使用量的升高而增加,但经过淹水处理后对照组与B300处理组的全磷含量低于非淹水处理组。(图1F)。

图 1 不同处理土壤有机磷、无机磷组分特征(平均值 ± 标准误)
Figure 1. Soil organic and inorganic phosphorus fractions under different treatments(mean ± SE)
-
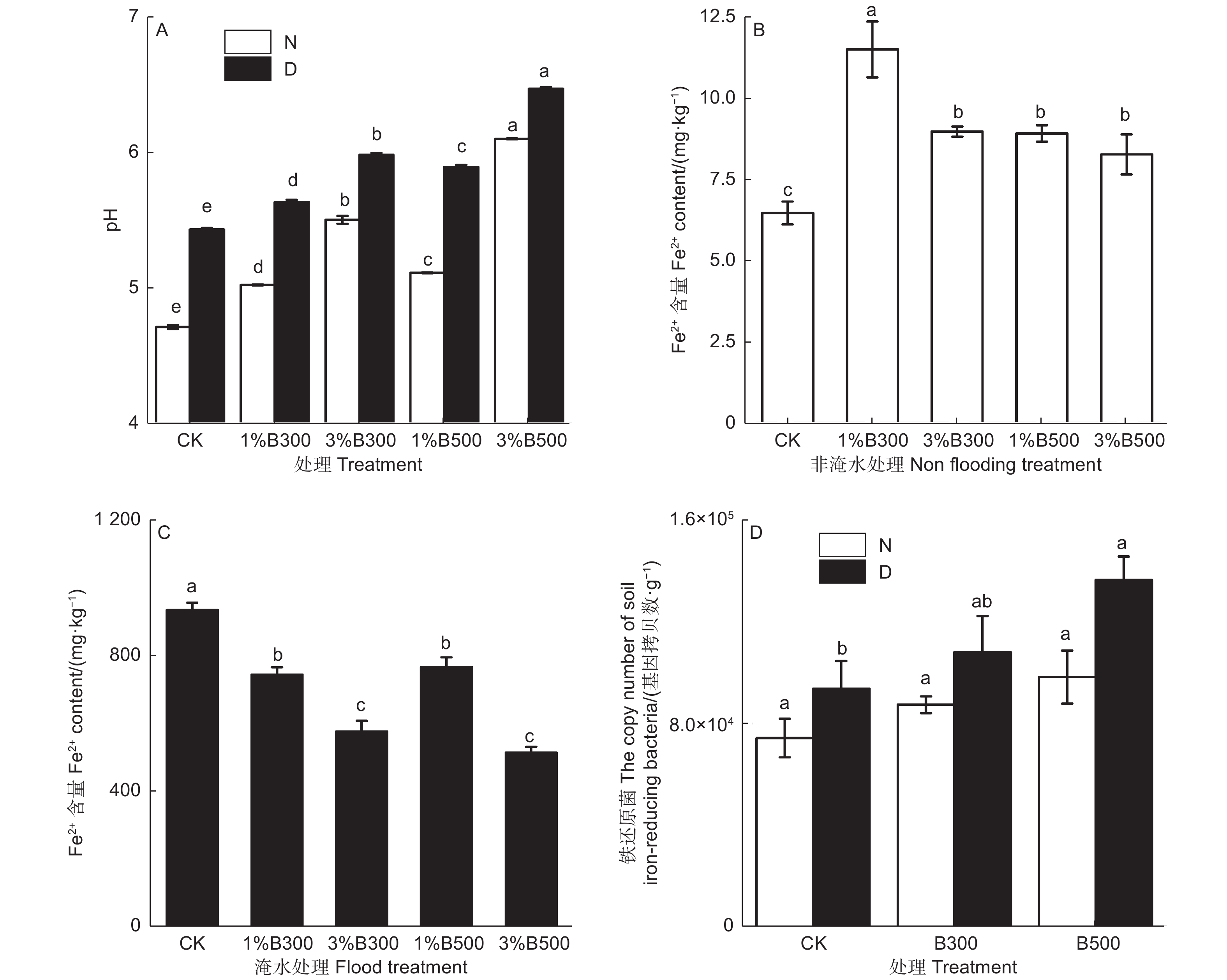
不同处理显著影响土壤化学性质(P<0.05,图2A)。淹水厌氧处理组pH值高于非淹水处理组,添加生物炭均显著提高土壤pH值。不同水分状态下施加生物炭显著影响土壤亚铁离子含量与铁还原菌群落结构(P<0.05,图2BCD)。淹水处理组土壤亚铁离子含量高于非淹水处理组,土壤亚铁离子含量在非淹水条件下,添加生物炭处理组高于对照组;在淹水条件下,随生物炭添加的增加而降低,并在B300与B500处理下效果相似。淹水处理组的铁还原菌数量高于非淹水处理,不同水分处理下随生物炭的添加与制备温度的升高土壤中铁还原菌数量均增加,其中淹水处理有明显的促进作用,而非淹水处理无显著差异。

图 2 不同处理土壤pH、亚铁离子含量和土壤铁还原菌基因拷贝数(平均值 ± 标准误)
Figure 2. Soil pH,ferrous ion content of soil under and the copy number of soil iron-reducing bacteria under different treatments(mean ± SE)
-
添加生物炭能显著影响土壤解磷菌群落结构。对97%相似水平下的OTUs进行生物信息统计分析,共产生7 569个OTUs,经过抽平处理剩余7 240个。由表1可以看出:OTUs的数量淹水处理组高于非淹水处理组。土壤中解磷菌丰富度指数Chao1,在非淹水处理下对照组高于生物炭处理组,淹水处理下生物炭处理组高于对照组。淹水处理下的群落多样性指数Shannon显著高于非淹水处理,D-B500处理最高。基于OUTs丰富度的非度量多维度分析得出解磷菌的Beta多样性指数NMDS1与NMDS2,结果表明,淹水厌氧处理组和非淹水处理组分别位于NMDS1的正负两侧,添加生物炭处理组与未添加生物炭处理组分别位于NMDS2的正负两侧,说明淹水与非淹水下添加生物炭对解磷菌群落有显著影响。
表 1 不同处理下土壤解磷菌群落OTUs、α多样性和β多样性指数(平均值 ± 标准误)
Table 1. OTUs, α diversity and β diversity indices of soil phosphorus solubilizing bacteria communities under different treatments(mean ± SE)
处理组
Treatment group分类操作单元
OTUsα多样性指数
Alpha exponentβ多样性指数
Beta exponentChao1指数
Chao1 indexShannon指数
Shannon indexNMDS1指数
NMDS1 indexNMDS2指数
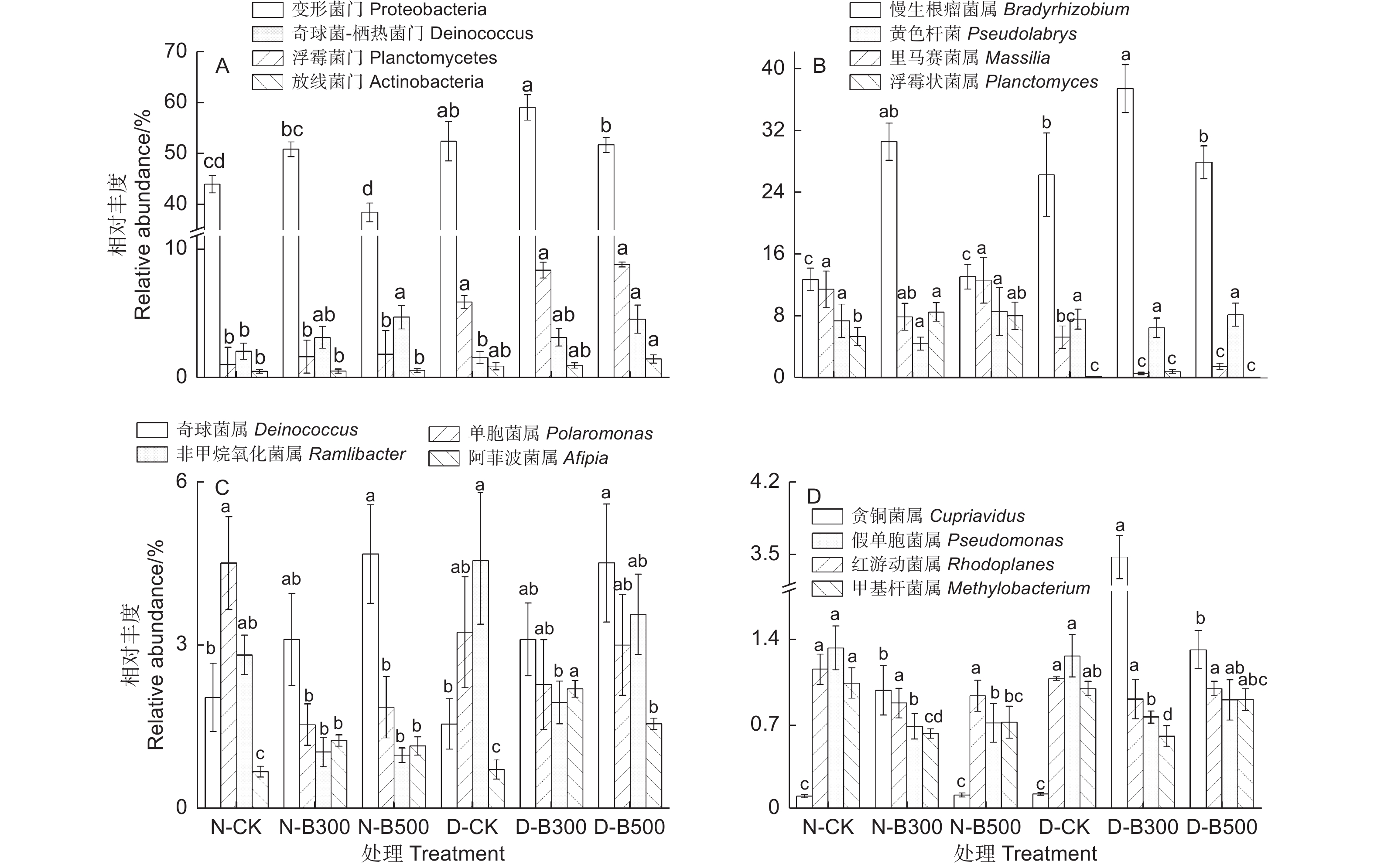
NMDS2 indexN-CK 1 918 ± 31.4 c 2 713.5 ± 48.9 a 7.2 ± 0.0 d −0.6 ± 0.2 −0.7 ± 0.1 N-B300 1903.3 ± 36.6 c 2 492.2 ± 39.8 ab 7.6 ± 0.0 c −0.6 ± 0.1 0.8 ± 0.1 N-B500 1 982.7 ± 36.4 abc 2 638.9 ± 47.3 ab 7.2 ± 0.2 d −1.1 ± 0.2 0.2 ± 0.1 D-CK 1 939 ± 47.2 bc 2 450.1 ± 148.4 b 7.7 ± 0.1 bc 0.5 ± 0.2 −0.8 ± 0.2 D-B300 2 053.3 ± 35.5 ab 2 604.3 ± 51.1 ab 8.0 ± 0.0 ab 1.1 ± 0.16 0.5 ± 0.2 D-B500 2 061.3 ± 36.8 a 2 601.5 ± 35.0 ab 8.0 ± 0.1 a 0.6 ± 0.1 0.1 ± 0.1 模拟厌氧条件下添加生物炭显著影响了土壤解磷菌的群落组成(P>0.05)。在门分类水平下,解磷菌以变形菌门(Proteobacteria)(38.48%~59.07%)、浮霉菌门(Planctomycetes)(1.02%~8.76%)、奇球菌-栖热菌门(Deinococcus−Thermus)(1.56%~4.69%)和放线菌门(Actinobacteria)(0.49%~1.45%)为主。淹水处理后土壤中变形菌门、浮霉菌门与放线菌门的相对丰度增加,变形菌门在添加B300后增高,添加B500后降低;浮霉菌门与放线菌门添加生物炭后各处理组的相对丰度无显著差异。奇球菌-栖热菌门相对丰度随制备温度的升高有上升趋势(图3A)。

图 3 不同处理下土壤解磷菌在门、属水平上的相对丰度变化(平均值 ± 标准误、相对丰度>1%)
Figure 3. Changes in relative abundance of soil phosphorus solubilizing bacteria at the phylum and genus level under different treatments (mean ± SE,relative abundance>1%)
不同处理下土壤解磷菌优势菌属见图3BCD。土壤添加生物炭增加了慢生根瘤菌属(Bradyrhizobium)、奇球菌属(Deinococcus)、阿菲波菌属(Afipia)和贪铜菌属(Cupriavidus)的相对丰度。淹水处理增加了慢生根瘤菌属、阿菲波菌属和贪铜菌属的相对丰度,降低里马赛菌属(Massilia)和浮霉状菌属(Planctomyces)的相对丰度,淹水处理下添加生物炭后降低了非甲烷氧化菌属(Ramlibacter)、假单胞菌属(Pseudomonas)和甲基杆菌属(Methylobacterium)的相对丰度。
-
随机森林模型展示出土壤化学因子、解磷菌Alpha、解磷菌Beta和铁还原菌对活性磷(A)、中等活性磷(B)和稳定态磷(C)活性的重要性程度(Increase in MSE%)。由图4可知:Beta MNDS1、Fe(II)和Shannon index对活性磷和稳定态磷的重要性影响程度最大,而Beta MNDS2对中等活性磷的重要性影响程度最大,其次是pH、Shannon index、TC、Fe(II)、Chao1 index、Beta MNDS1,这说明解磷菌群落结构特征对各磷形态起到重要作用,其中Fe(II)、FRB对稳定态磷与活性磷也起到重要作用,而对于中等活性磷来说环境因子pH、TC所起作用大于Fe(II)、FRB等。

图 4 随机森林模型
Figure 4. Random forest model
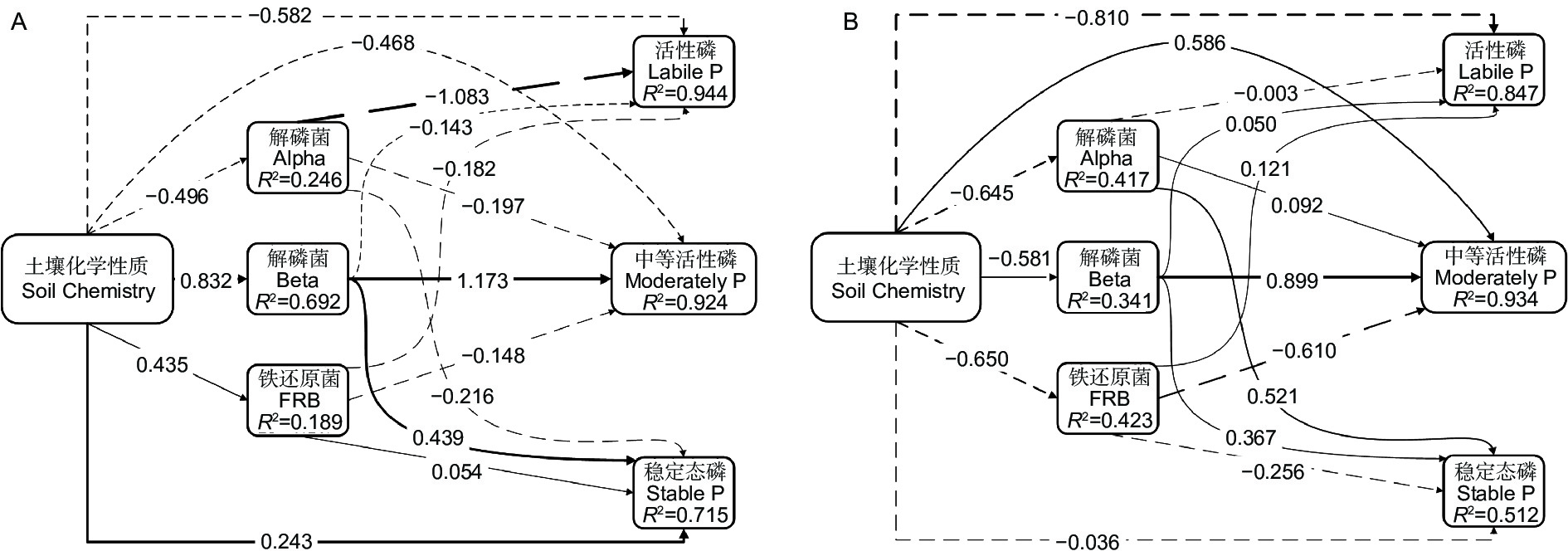
偏最小二乘路径模型(PLS-PM)(图5)展示了非淹水(A)与淹水(B)处理下土壤化学性质、土壤解磷菌、铁还原菌与土壤磷形态之间的关系,进而解释各因子对活性磷、中等活性磷和稳定态磷的影响程度。图中线条粗细代表路径系数的直接效应绝对值大小,实线代表正效应,虚线代表负效应。非淹水与淹水模型的拟合度分别为0.654 4、0.707 5。非淹水状态下,化学指标对稳定态磷、解磷菌Beta的直接影响均高于淹水状态。铁还原菌对各形态磷的直接效应低于0.2。淹水状态下,土壤化学指标对活性磷、中等活性磷的直接效应大于非淹水状态,淹水处理后对稳定态磷的直接效应由正效应转变为负效应。化学指标对铁还原菌的直接影响为−0.650,解磷菌Alpha与Beta对各形态磷的直接效应相比于非淹水状态有减少,但解磷菌Alpha对稳定态磷的直接效应增加、铁还原菌对中等活性磷与稳定态磷的直接效应增加,为0.521、−0.256。这说明淹水处理能改变土壤中解磷菌与铁还原菌对磷形态的转化。

图 5 偏最小二乘法路径模型(PLS-PM)
Figure 5. Partial least squares path model
-
本研究通过添加生物炭增加了有效磷含量,可能是因为生物炭本身附带的磷素溶解到土壤中[20],也可能是生物炭的添加改变了土壤环境,使土壤磷素形态发生了变化[14]。不同温度制备的生物炭对土壤磷素的影响不同,淹水条件下B500处理组H2O-Pi含量最低,H2O-Po含量最高,说明500 ℃下制备的生物炭H2O-Po向H2O-Pi转换的量与300 ℃下制备的生物炭相比更少,还可能是因为B500生物炭在淹水条件下能吸附更多的水溶性无机磷,或者是因为更多的铁被还原成氢氧化物,进而吸附土壤中更多的无机磷。在淹水条件下施加生物炭显著增加NaHCO3-Po占比,可能是因为生物炭的存在为自身提供了还原和配位体条件,铁氧化物与生物炭所携带的有机质发生络合作用,使得吸附在铁氧化物表面的NaHCO3-Po被解吸下来[21]。水溶态磷和碳酸氢钠态磷是植物可直接利用或容易利用的活性磷,因此土壤中添加生物炭主要提高了土壤有机磷含量,提高了土壤磷素的有效性,这与Zhou等人的研究结果一致[15]。由随机森林模型发现解磷菌、Fe(II)和铁还原菌对活性磷有重要影响,在厌氧条件下,Fe(III)可以还原为Fe(II)破坏闭蓄磷的氧化铁膜,释放出Fe(II)和水溶态无机磷[22]。也有研究发现随pH的升高,铁氧化物对无机磷的吸附能力降低[23],使得土壤活性磷含量上升。
非淹水状态下,施加不同温度制备的生物炭均增加了NaOH-Pi的含量,淹水处理后,NaOH-P在B500处的含量减少,这说明了不同温度制备的生物炭在厌氧情况下对磷形态的转化作用不同,B500生物炭可以促进NaOH-P转化为植物可利用的活性磷。pH和TC显著(P<0.05)影响土壤中等活性磷,并且解磷菌群落对其也有重要作用,Olander等[24]研究发现,土壤炭添加会刺激微生物对磷的需求,示踪剂P从吸附池移动到微生物池,可能因为土壤环境变化引起微生物群落结构改变,促进微生物对NaOH-P的转化[25]。HCI-Po是仅次于NaHCO3-Po为土壤供给有效磷的来源之一,本研究淹水条件下HCI-Po含量显著升高,为土壤提供大量磷源。残渣态磷通过与Ca和Fe形成络合物,具有高稳定特性,同一水分条件下各处理组之间的残渣态磷无明显差别,这和杜艳玲[26]等人研究结果一致,残渣态磷在土壤中较稳定,对生物炭添加响应不敏感。王岩等[27]研究发现,碱性条件会促进HCl-P和Residual-P等难溶性有机磷向活性有机磷的转化,本研究发现残渣态磷在淹水条件下含量减少,并且pH对稳定态磷的影响极显著(P<0.01),这说明,淹水条件下生物炭的添加使得土壤pH升高,促进Residual-P向其它形态磷转化。
-
非淹水处理下添加生物炭显著提高了土壤中亚铁离子的含量,这可能是生物炭的添加升高了土壤环境的pH值,近中性pH条件适宜铁还原菌的生长,进而促进铁氧化物的还原[28-29],可能是生物炭本身富含的醌类物质充当电子穿梭体[30-31],向Fe(III)转移更多的电子,进而促进Fe(III)还原生成Fe(II)。Xu等[32]研究发现,生物炭可提升赤铁矿的还原速率,首先是生物炭表面的醌类官能团作为电子中介体促进赤铁矿还原,其次是生物炭能够作为吸附剂在其表面富集还原生成的Fe(II)使得赤铁矿表面暴露更多的还原位点,进一步促进赤铁矿还原,另一方面有研究发现土壤微生物具有还原作用,会促进铁矿物还原溶解,释放亚铁离子[33]。在淹水条件下,Fe(II)含量大幅上升,但随生物炭添加量的增多而降低,这可能是因为原本在淹水厌氧环境中低还原电位促使土壤中大量氧化铁还原为Fe(II),但添加生物炭后升高了土壤pH值,水环境中OH-随之增多,与部分Fe(II)结合产生Fe(OH)2白色沉淀,由于Fe(OH)2很不稳定,很快就被氧化成Fe(OH)3,故降低了土壤Fe(II)含量[34]。同时有研究表明,生物炭添加会影响铁还原菌对土壤中铁矿物的还原溶出,降低亚铁离子含量[35]。
添加生物炭能够改变盐基离子、土壤有机质和微生物的聚集状态[36],而在淹水条件下,有机质的厌氧分解又影响着土壤的Eh和pH[37]。此外,土壤环境的改变还影响着厌氧微生物的丰富度和多样性,生物炭为异养型的铁还原菌提供生长所需的碳源,同时有机质为其充当络合剂并提供电子[38],所以淹水厌氧处理的铁还原菌拷贝数均大于非淹水处理组。
与非淹水条件相比,淹水条件下铁还原菌对各磷形态的直接效应增加,对中等活性磷和稳定态磷的负效应值增大,对活性磷的直接效应由负转为正。这说明淹水处理下,铁还原菌开始起比较重要的作用,其含量与稳定态磷呈负相关性。有研究表明添加生物炭可以促进铁还原菌对铁的还原,进而促进磷素的活化[39]。生物炭作为土壤改良剂是一种可高效利用的再生资源,可以提高土壤碳、氮、磷、钾等元素含量,改善土壤质量。夏丽丹等人研究发现相同水分条件下土壤pH值和生物炭添加量显著正相关[40],与本研究结果相一致。
-
本研究表明在厌氧条件下,添加生物炭能显著提高解磷菌的OTUs丰度、多样性指数和丰富度指数(P<0.05),且根据NMDS分析数据可以看出,土壤经过厌氧、添加生物炭处理后解磷菌的结构特征表现出明显不同。Grossman等[41]发现,生物炭改良过的土壤微生物群落组成更为丰度,与本研究结果一致。这可能是因为生物炭的施加改变了土壤化学性质,提高了pH值和全碳含量,为解磷菌提供充足碳源和近中性的土壤环境,进而改变了解磷菌的群落结构[42-43]。放线菌是重要的降解菌,可降解多种有机物[44],在碳源丰富的环境中能够快速生长[45],其相对丰度随生物炭的添加而上升。有研究发现土壤中添加生物炭可以增加解磷菌黄杆菌属和假单胞菌属的相对丰度[46],这与本实验结果一致。变形菌门的分布范围广、种类多、适应性强,但土壤添加B500生物炭后相对丰度下降,可能是由于500 ℃下制备的生物炭不利于变形菌门利用。朱启林等[47]研究发现,随制备温度的升高,生物炭表面烷烃基、甲基等官能团减少,生物炭结构更加稳定。由偏最小二乘方差分析发现,淹水处理后解磷菌对磷素形态的直接效应减弱,可能是因为厌氧环境阻碍解磷菌对磷的转化,解磷菌群落为适应厌氧环境而向r对策发展,使得其多样性和丰富度指数上升,群落结构多样化。慢生根瘤菌属属于好氧或兼性厌氧革兰氏阴性杆菌,在厌氧条件下能够生长繁殖。浮霉菌门属于小门水生细菌,故淹水处理组浮霉菌门的相对丰度显著高于非淹水组,与禹桃兵[48]等对大豆品种根际微生物群落结构特征的研究结果不同,可能是植物的参与造成结果不同。解磷菌群落结构多样性对土壤中等活性磷的重要性最大,且有显著影响(P<0.05),不同门属的解磷菌群落可以分解产生酸性磷酸酶和碱性磷酸酶,进而促进中等活性磷的分解[49]。Esberg等[25]研究发现,微生物呼吸与NaOH提取态磷的变化之间存在相关性,表明微生物对该组分的获取更大,能促进NaOH提取态磷向活性磷转化。生物炭的添加使得土壤环境pH升高,而奇球菌属、阿菲波菌属的最适生长pH在7.1左右[50],有利于其生长繁殖。马赛菌属属于好氧革兰氏阴性菌,浮霉状菌属属于专性好氧菌,在厌氧条件下二者均由于缺少氧气而生长受限,导致其相对丰度下降。
-
本研究表明,淹水厌氧条件下添加生物炭能在一定程度上提高活性磷和中等活性磷含量,降低稳定态磷含量,这是由于生物炭的添加改变了土壤的化学性质,土壤pH升高使得铁还原菌数量增加,使铁氧化物结合的中等活性磷向活性磷转化;植物叶片经烧制后含有磷元素,成为土壤磷素的直接来源,进而提高土壤全磷、有效磷含量;土壤环境变化改变了解磷菌群落结构和多样性,促进闭蓄态磷向活性磷转化;淹水厌氧处理提供的厌氧环境进一步促进土壤Fe(III)的还原,并有利于厌氧微生物的生长,促进稳定态磷素分解,提高土壤磷素有效性。因此,在南方降雨充沛地区,杉木人工林施加生物炭能够改善土壤养分状况,提高土壤有效磷素含量,解决土壤有效磷素缺乏的问题,从而促进林木生长发育,为杉木人工林的可持续经营管理提供参考。
生物炭施用下亚热带红壤铁还原及磷形态转化关系研究
Relationship between Iron Reduction and Phosphorus Transformation in Subtropical Red Soil under Biochar Application
-
摘要:
目的 研究降雨丰沛,土壤水分长期饱和条件下生物炭对杉木人工林土壤铁还原的影响,分析土壤铁还原菌、解磷菌群落结构变化规律,最终明确生物炭对土壤铁还原的影响及与磷形态转化的关系。 方法 以杉木人工林红壤为供试土壤,收集林下杉木叶烧制成300℃和500℃生物炭,以0、1%、3%占比添加生物炭进行40 d的室内培养。测定土壤基本化学养分,采用修正后的Hedley方法测定土壤中不同磷素形态含量,利用高通量测序技术分析土壤解磷菌与铁还原菌群落结构。 结果 淹水处理后土壤活性磷含量增多,并且随生物炭添加量的增加而增加,其中水溶态有机磷和碳酸氢钠态无机磷占比较大;残渣态磷含量随生物炭添加量的增加而减少。淹水处理组的铁还原菌基因拷贝数高于非淹水处理组,且同一水分条件下随生物炭烧制温度的增加而增加,淹水处理组的亚铁离子含量远高于非淹水处理,且随生物炭添加量的增加而降低,淹水处理组的土壤化学性质例如pH、全碳、全磷含量高于非淹水处理组,且随生物炭添加量的增加而上升。在淹水处理中土壤解磷菌群落丰富度随烧制温度的升高而增加,并且解磷菌群落结构和多样性增强。 结论 厌氧条件促进Fe(III)还原,生物炭的添加改变了土壤化学性质,影响土壤解磷菌群落结构和多样性、铁还原菌的生长微环境,在微生物与Fe(III)还原的双重作用下,促进残渣态磷与氢氧化钠态磷向水溶态磷和碳酸氢钠态有机磷转化,增强了土壤磷素有效性。因此,在南方降雨充沛地区,杉木人工林施加生物炭能够改善土壤养分状况,为杉木生长提供足够磷素。 Abstract:Objective To study the effect of biological carbon on soil iron reduction in Chinese fir plantations under the condition of abundant rainfall and long-term soil water saturation, and analyze the community structure changes of iron-reducing bacteria and phosphorus solving bacteria in soil, for clarifying the influence of biochar on soil iron reduction and its relationship with phosphorus form transformation. Method Based on the red soil of Chinese fir plantations, Chinese fir leaves under the forest were collected and fired to produce 300℃ and 500℃ biochar. Biochar was added at 0, 1% and 3% for 40 days of indoor culture.The basic chemical nutrients of soil were analyzed. The content of different phosphorus forms in soil was determined by the modified Hedley method, and the community structure of soil phosphate solubilizing bacteria and iron reducing bacteria was analyzed by high-throughput sequencing technology. Result The content of soil active phosphorus increased with the increase of the amount of biochar, and most of them were the H2O-Po and NaHCO3-Pi. The content of Residual-P decreased with the increase of biological carbon. The copy number of genes of Fe (III)-reducing bacteria in flooded treatment group was higher than that in non-flooded treatment group, and increased with the increase of biochar firing temperature under the same water condition. The content of ferrous ions in flooded treatment group was much higher than that in non-flooded treatment group, and decreased with the increase of biochar addition. The soil chemical properties such as pH, TC and TP contents in the flooded group were higher than those in the non-flooded treatment group, and increased with the increase of biochar addition amount. The richness of soil phosphorus solubilizing bacteria community increased with increasing firing temperature, and the community structure and diversity of phosphorus solubilizing bacteria also increased with increasing firing temperature. Conclusion The anaerobic conditions promote the reduction of Fe (III). The addition of biochar changes the chemical properties of soil, affects the community structure and diversity of phosphorus solving bacteria and the growth microenvironment of iron reducing bacteria. Under the double action of microorganism and Fe (III) reduction, the conversion of residual phosphorus and sodium hydroxide phosphorus into aqueous phosphorus and sodium bicarbonate phosphorus are promoted, and the availability of soil phosphorus is enhanced. Therefore, in the southern regions with abundant rainfall, the application of biological carbon in Chinese fir plantation can improve the soil nutrient status and provide enough phosphorus for the growth of Chinese fir. -
图 1 不同处理土壤有机磷、无机磷组分特征(平均值 ± 标准误)
Figure 1. Soil organic and inorganic phosphorus fractions under different treatments(mean ± SE)
图 2 不同处理土壤pH、亚铁离子含量和土壤铁还原菌基因拷贝数(平均值 ± 标准误)
Figure 2. Soil pH,ferrous ion content of soil under and the copy number of soil iron-reducing bacteria under different treatments(mean ± SE)
图 3 不同处理下土壤解磷菌在门、属水平上的相对丰度变化(平均值 ± 标准误、相对丰度>1%)
Figure 3. Changes in relative abundance of soil phosphorus solubilizing bacteria at the phylum and genus level under different treatments (mean ± SE,relative abundance>1%)
表 1 不同处理下土壤解磷菌群落OTUs、α多样性和β多样性指数(平均值 ± 标准误)
Table 1. OTUs, α diversity and β diversity indices of soil phosphorus solubilizing bacteria communities under different treatments(mean ± SE)
处理组
Treatment group分类操作单元
OTUsα多样性指数
Alpha exponentβ多样性指数
Beta exponentChao1指数
Chao1 indexShannon指数
Shannon indexNMDS1指数
NMDS1 indexNMDS2指数
NMDS2 indexN-CK 1 918 ± 31.4 c 2 713.5 ± 48.9 a 7.2 ± 0.0 d −0.6 ± 0.2 −0.7 ± 0.1 N-B300 1903.3 ± 36.6 c 2 492.2 ± 39.8 ab 7.6 ± 0.0 c −0.6 ± 0.1 0.8 ± 0.1 N-B500 1 982.7 ± 36.4 abc 2 638.9 ± 47.3 ab 7.2 ± 0.2 d −1.1 ± 0.2 0.2 ± 0.1 D-CK 1 939 ± 47.2 bc 2 450.1 ± 148.4 b 7.7 ± 0.1 bc 0.5 ± 0.2 −0.8 ± 0.2 D-B300 2 053.3 ± 35.5 ab 2 604.3 ± 51.1 ab 8.0 ± 0.0 ab 1.1 ± 0.16 0.5 ± 0.2 D-B500 2 061.3 ± 36.8 a 2 601.5 ± 35.0 ab 8.0 ± 0.1 a 0.6 ± 0.1 0.1 ± 0.1  下载: 导出CSV
下载: 导出CSV
-
[1] 国家林业和草原局. 中国森林资源报告(2014-2018)[M]. 北京: 中国林业出版社. 2019, 28-29. [2] 刘 丽, 段争虎, 汪思龙, 等. 不同发育阶段杉木人工林对土壤微生物群落结构的影响[J]. 生态学杂志, 2009, 28(12):2417-2423. doi: 10.13292/j.1000-4890.2009.0421 [3] LUAN J, XIANG C, LIU S,et al. Assessments of the impacts of Chinese fir plantation and natural regenerated forest on soil organic matter quality at Longmen mountain, Sichuan, China[J]. Geoderma, 2010, 156(3): 228-236. [4] 韦宜慧, 陈嘉琪, 董玉红, 等. 杉木人工林土壤溶磷细菌筛选及培养条件优化[J]. 林业科学研究, 2020, 33(4):83-91. doi: 10.13275/j.cnki.lykxyj.2020.04.011 [5] 王永壮, 陈 欣, 史 奕. 农田土壤中磷素有效性及影响因素[J]. 应用生态学报, 2013, 24(1):260-268. doi: 10.13287/j.1001-9332.2013.0147 [6] 关诗洋, 王佳琪, 于 贺, 等. 减水减肥对设施黑土菜田土壤无机磷形态及分布的影响[J]. 中国农学通报, 2021, 37(23):89-93. doi: 10.11924/j.issn.1000-6850.casb2021-0106 [7] 张 虹, 于姣妲, 李海洋, 等. 不同栽植代数杉木人工林土壤磷素特征研究[J]. 林业科学研究, 2021, 34(1):10-18. doi: 10.13275/j.cnki.lykxyj.2021.01.002 [8] LI Q, BI S, JI G. Determination of strongly reducing substances in sediment[J]. Environmental Science & Technology, 2003, 37(24): 5727-5731. [9] SMOLDERS E, BAETENS E, VERBEECK M,et al. Internal loading and redox cycling of sediment iron explain reactive phosphorus concentrations in lowland rivers[J]. Environmental Science & Technology, 2017, 51(5): 2584-2592. [10] 周垂帆, 林静雯, 李 莹, 等. 磷与草甘膦在酸性土壤中吸附解吸交互作用机制[J]. 农业环境科学学报, 2016, 35(12):2367-2376. doi: 10.11654/jaes.2016-0862 [11] FERNANDES A P, NUNES T C, PAQUETE C M,et al. Interaction studies between periplasmic cytochromes provide insights into extracellular electron transfer pathways of Geobacter sulfurreducens[J]. Biochemical Journal, 2017, 474(5): 797-808. doi: 10.1042/BCJ20161022 [12] WU P, WANG G, FAROOQ T H,et al. Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: does neighboring root activity aggravate P nutrient deficiency?[J]. Journal of Soils and Sediments, 2017, 17(12): 2775-2785. doi: 10.1007/s11368-017-1852-8 [13] YU J, TANG L, PANG Y,et al. Magnetic nitrogen-doped sludge-derived biochar catalysts for persulfate activation: Internal electron transfer mechanism[J]. Chemical Engineering Journal, 2019, 364: 146-159. doi: 10.1016/j.cej.2019.01.163 [14] BANINAJARIAN S, SHIRVANI M. Use of biochar as a possible means of minimizing phosphate fixation and external P requirement of acidic soil[J]. Journal of Plant Nutrition, 2021, 44(1): 59-73. doi: 10.1080/01904167.2020.1792491 [15] ZHOU C, HEAL K, TIGABU M,et al. Biochar addition to forest plantation soil enhances phosphorus availability and soil bacterial community diversity[J]. Forest Ecology and Management, 2020, 455: 117635. doi: 10.1016/j.foreco.2019.117635 [16] 令狐荣云, 余炜敏, 王荣萍, 等. 铁还原菌Shewanella oneidensis MR-1对铁磷复合物中铁、磷释放规律的影响[J]. 生态环境学报, 2017, 26(10):1704-1709. [17] JIAN-FEN G, YU-SHENG Y, GUANG-SHUI C,et al. Dissolved organic carbon and nitrogen in precipitation, throughfall and stemflow from Schima superba and Cunninghamia lanceolata plantations in subtropical China[J]. Journal of Forestry Research, 2005, 16(1): 19-22. doi: 10.1007/BF02856847 [18] 中国科学院南京土壤研究所. 土壤理化分析[M]. 上海: 上海科学技术出版社, 1978. [19] ZHANG Y, LI Y, WANG S,et al. Soil phosphorus fractionation and its association with soil phosphate-solubilizing bacteria in a chronosequence of vegetation restoration[J]. Ecological Engineering, 2021, 164: 106208. doi: 10.1016/j.ecoleng.2021.106208 [20] HALE S E, ALLING V, MARTINSEN V,et al. The sorption and desorption of phosphate-P, ammonium-N and nitrate-N in cacao shell and corn cob biochars[J]. Chemosphere, 2013, 91(11): 1612-1619. doi: 10.1016/j.chemosphere.2012.12.057 [21] 谷丽丽. 长期定位施肥及水田连作对农田土壤中磷赋存形态的影响[D]. 武汉: 华中农业大学, 2017. [22] PETTICREW E, AROCENA J. Evaluation of iron-phosphate as a source of internal lake phosphorus loadings[J]. The Science of The Total Environment, 2001, 266(1-3): 87-93. doi: 10.1016/S0048-9697(00)00756-7 [23] 郭智俐, 李 苓, 刘晓月, 等. 两种铁氧化物对无机磷的吸附特征分析[J]. 中国海洋大学学报(自然科学版), 2021, 51(8):42-48. doi: 10.16441/j.cnki.hdxb.20200271 [24] OLANDER L P, VITOUSEK P M. Biological and geochemical sinks for phosphorus in soil from a wet tropical forest[J]. Ecosystems, 2004, 7(4): 404-419. [25] ESBERG C, DU TOIT B, OLSSON R,et al. Microbial responses to P addition in six South African forest soils[J]. Plant and Soil, 2010, 329(1-2): 209-225. doi: 10.1007/s11104-009-0146-3 [26] 杜艳玲, 周怀平, 杨振兴, 等. 长期不同秸秆还田方式对褐土磷素组分的影响[J]. 山西农业科学, 2019, 47(11):1947-1954 + 1959. doi: 10.3969/j.issn.1002-2481.2019.11.20 [27] 王 岩, 张志勇, 秦红杰, 等. 种养凤眼莲条件下pH值对底泥中不同形态磷释放的影响[J]. 南京农业大学学报, 2017, 40(4):681-689. doi: 10.7685/jnau.201611021 [28] 索慧慧, 林 颖, 赵苗苗, 等. 生物炭对淹水土壤中溶解性有机质含量及组成特征的影响[J]. 水土保持学报, 2019, 33(2):155-161,271. doi: 10.13870/j.cnki.stbcxb.2019.02.025 [29] BLÖTHE M, AKOB D M, KOSTKA J E,et al. pH gradient-induced heterogeneity of Fe(III)-reducing microorganisms in coal mining-associated lake sediments[J]. Applied and Environmental Microbiology, 2008, 74(4): 1019-1029. doi: 10.1128/AEM.01194-07 [30] KLÜPFEL L, KEILUWEIT M, KLEBER M,et al. Redox properties of plant biomass-derived black carbon (biochar)[J]. Environmental Science & Technology, 2014, 48(10): 5601-5611. [31] SHI L, DONG H, REGUERA G,et al. Extracellular electron transfer mechanisms between microorganisms and minerals[J]. Nature Reviews Microbiology, 2016, 14(10): 651-662. doi: 10.1038/nrmicro.2016.93 [32] XU S, ADHIKARI D, HUANG R,et al. Biochar-facilitated microbial reduction of hematite[J]. Environ Sci Technol, 2016, 50: 2389-2395. doi: 10.1021/acs.est.5b05517 [33] AYYASAMY P M, CHUN S, LEE S. Desorption and dissolution of heavy metals from contaminated soil using Shewanella sp. (HN-41) amended with various carbon sources and synthetic soil organic matters[J]. Journal of Hazardous Materials, 2009, 161(2): 1095-1102. [34] 文帅龙, 刘静静, 戴家如, 等. 铁(氢)氧化物介导的溶解性有机质、无机磷的固定及相互作用研究进展[J]. 湖泊科学, 2022, 34(5):1428-1440. doi: 10.18307/2022.0502 [35] 张又弛, 李会丹. 生物炭对土壤中铁生物还原作用和重金属分布的影响[J]. 环境污染与防治, 2019, 41(4):377-381. doi: 10.15985/j.cnki.1001-3865.2019.04.001 [36] 包明琢, 曲雪铭, 高倩倩, 等. 磷肥和生物炭配施对杉木林地土壤微生物的影响[J]. 西北林学院学报, 2022, 37(2):10-19. doi: 10.3969/j.issn.1001-7461.2022.02.02 [37] 曲 植, 李丽娜, 贾 蓉. 水稻土中水溶性有机碳对铁还原过程的贡献[J]. 植物营养与肥料学报, 2018, 24(2):346-356. [38] CUI X, FANG S, YAO Y,et al. Potential mechanisms of cadmium removal from aqueous solution by Canna indica derived biochar[J]. Science of The Total Environment, 2016, 562: 517-525. doi: 10.1016/j.scitotenv.2016.03.248 [39] DING X, ZHANG S, WANG R,et al. Exogenous labile C application enhances Fe-P utilization for mycorrhizal plants through iron-reducing bacteria in subtropical soil[J]. Journal of soil science and plant nutrition, 2014, 14(4): 803-817. [40] 夏丽丹, 曹 升, 张 虹, 等. 不同水分条件下生物炭对红壤磷素形态及磷酸酶活性的影响[J]. 农业环境科学学报, 2019, 38(5):1101-1111. doi: 10.11654/jaes.2018-1171 [41] GROSSMAN J M, O’NEILL B E, TSAI S M,et al. Amazonian anthrosols support similar microbial communities that differ distinctly from those extant in adjacent, unmodified soils of the same mineralogy[J]. Microbial Ecology, 2010, 60(1): 192-205. doi: 10.1007/s00248-010-9689-3 [42] SIMARANI K, AZLAN HALMI M F, ABDULLAH R. Short-term effects of biochar amendment on soil microbial community in humid tropics[J]. Archives of Agronomy and Soil Science, Taylor & Francis, 2018, 64(13): 1847-1860. [43] 张燕林, 黄彩凤, 包明琢, 等. 生物炭及其老化对杉木林土壤养分含量和微生物群落组成影响的室内模拟[J]. 林业科学, 2021, 57(6):169-179. doi: 10.11707/j.1001-7488.20210619 [44] 陈倩倩, 刘 波, 王阶平, 等. 基于宏基因组方法分析养猪发酵床微生物组季节性变化[J]. 农业环境科学学报, 2018, 37(6):1240-1247. doi: 10.11654/jaes.2017-1330 [45] ZENG J, LIU X, SONG L,et al. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition[J]. Soil Biology and Biochemistry, 2016, 92: 41-49. doi: 10.1016/j.soilbio.2015.09.018 [46] LIU S, MENG J, JIANG L,et al. Rice husk biochar impacts soil phosphorous availability, phosphatase activities and bacterial community characteristics in three different soil types[J]. Applied Soil Ecology, 2017, 116: 12-22. doi: 10.1016/j.apsoil.2017.03.020 [47] 朱启林, 曹 明, 张雪彬, 等. 不同热解温度下禾本科植物生物炭理化特性分析[J]. 生物质化学工程, 2021, 55(4):21-28. doi: 10.3969/j.issn.1673-5854.2021.04.004 [48] 禹桃兵, 石琪晗, 年 海, 等. 涝害对不同大豆品种根际微生物群落结构特征的影响[J]. 作物学报, 2021, 47(9):1690-1702. [49] 王兵爽, 李淑君, 张舒桓, 等. 西瓜根系分泌酸性磷酸酶对有机肥营养的响应[J]. 土壤学报, 2019, 56(2):454-465. doi: 10.11766/trxb201807090318 [50] 刘梦葭, 杨 粟, 程凯莹, 等. 奇球菌属的最新研究进展及其应用[J]. 核农学报, 2017, 31(9):1723-1729. doi: 10.11869/j.issn.100-8551.2017.09.1723 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3156
- HTML全文浏览量: 1410
- PDF下载量: 101
- 被引次数: 0
