

-
全球气温的持续上升会改变陆地生态系统水循环的强度和过程,导致全球降雨格局发生变化[1],降雨量的改变势必会影响土壤水分和养分的有效性,从而引起植物对养分的利用与分配策略发生改变,尤其在半干旱、干旱地区这种水资源严重缺乏的地区更加显著。碳(C)、氮(N)、磷(P)和钾(K)作为植物生长发育的基本化学营养元素,可以调节植物光合速率、蒸腾速率以及繁殖生长等重要的生理生态过程[2-4],这些元素含量及其生态化学计量特征可以反映植物器官的内稳性以及各元素在不同器官中的分配比例和相互关系[5-6],同时又可以判断植物生长过程中的限制性元素和养分利用效率的高低[7-10]。通过对植物器官生态化学计量学特征的研究,可以掌握植物生长繁殖与更新修复过程中养分的循环利用状况。前期研究结果表明,在个体水平间植物根、茎、叶的生态化学计量特征存在一定的关联性[11-12]。国内外学者从不同尺度研究植物生态化学计量对环境因子的响应结果表明,植物化学元素含量及其生态化学计量比在各器官间的协同变化和耦合关系[8-10],有助于维持植物的稳定代谢和生长发育,从而反映了植物对环境变化的响应和适应[13-14]。因此,通过对植物碳、氮、磷、钾生态化学计量学特征的研究,有助于理解植物的生长调节机制和生存策略,探讨植物各器官与不同生境下降雨量差异对植物的影响。
沙冬青(Ammopiptanthus mongolicus (Maxim. ex Kom.) Cheng f.)作为第三纪孑遗种是国家二级重点保护的珍稀濒危植物,主要分布于内蒙古中西部、宁夏北部和甘肃北部,其中最集中且适宜的区域是西鄂尔多斯—东阿拉善地区的沙漠边缘和山麓洪积砾质坡地[15-16]。沙冬青是豆科植物,作为我国干旱荒漠区唯一常绿阔叶灌木,分类地位特殊,抗旱耐热性极强,在恶劣贫瘠的荒漠区可以持续发挥防风固沙作用。沙冬青的价值主要包括园林绿化观赏、根瘤固氮作用、叶和茎入药能祛风除湿和活血化瘀等、各器官内均含有多种物质可开发利用于生物工程[17]。国内外学者从器官、物种、群落以及生态系统等角度对植物化学计量特征进行了研究,并结合气候、地形、纬度等因子对生态系统化学计量特征的影响进行了讨论。研究发现水分差异会引起植物生长状况差异且影响土壤养分的有效性。因此,水分供应对植物的影响也会导致植物C、N、P和K元素含量及其计量比的差异,进一步反映了植物对不利环境的防御和适应策略[14]。植物不同器官具有各自特定的功能、生长和周转速率,从而导致植物体养分含量也存在器官间差异[8-9] ,而干旱荒漠生态系统由于蒸发强烈且降雨稀少,造成土壤干旱可能会通过改变植物体内化学元素含量而影响植物生长。受全球气候变化影响,中国西北地区干旱加剧,荒漠化扩大,植物对荒漠化环境的生态适应对策成为关键科学问题。目前植物器官生态化学计量学研究主要聚焦在叶片的碳氮磷元素含量及其化学计量比的研究,关于根系生态化学计量学特征的研究报道日渐增多,但相对于具有支撑输导作用的茎的化学计量特征研究知之甚少。各器官作为植物组成的构件应该视为一个整体,不同植物器官间的化学元素分配比例以及各器官间化学计量特征的关联性还缺乏认知。鉴于此,本研究分析沿自然降雨梯度条件下沙冬青根、茎、叶的C、N、P、K 元素含量及其计量比特征,从植物器官生态化学计量学角度理解植物的养分分配和利用策略,进一步了解沙冬青对环境的响应和适应机制,为揭示其濒危机制提供科学依据。
-
根据沙冬青在我国的分布情况,沿降雨梯度选择6个代表性的自然分布区,包括内蒙古中西部和甘肃北部等地区,地理分布范围为37.51°~41.94° N,102.51°~107.98° E,海拔范围为1 039.3~1 799.9 m,各分布区年平均气温在4.9~8.8 ℃之间,年均日照时数在2 725.7~3 368.4 h之间,年均降水量在53.5~269.8 mm之间,气候具有典型的雨热同期特征。自然分布区的土壤类型以荒漠风沙土、草原风沙土和石质土为主。在不同的降雨梯度下设置沙冬青群落的典型样地,用GPS定位记录采样点的经度(Longitude)、纬度(Latitude)和海拔(Altitude)信息(见表1)。
表 1 研究区位置与环境特征
Table 1. Sample location and environmental characteristics
站点各样地
Sites经纬度
Longitude, Latitude海拔
Altitude/m年平均降雨量
Mean annual
precipitation/mm年平均气温
Mean annual
temperature/℃日照时数
Mean annual sunshine
duration/h生境类型
Habitat
type鄂托克旗 107.98° E,39.11° N 1 380.3 269.8 7.2 2 846.1 荒漠草原区 阿拉善左旗 105.67° E,38.83° N 1 561.4 211.2 7.9 3 099.2 荒漠草原区 景泰 103.83° E,37.51° N 1 799.9 185.1 8.8 2 725.7 典型荒漠区 磴口 106.42° E,40.18° N 1 039.3 142.2 8.0 3 211.7 典型荒漠区 乌拉特后旗 106.36° E,41.74° N 1 126.3 101.3 4.9 2 968.4 典型荒漠区 额济纳旗 102.51° E,41.94° N 1 051.9 53.5 8.7 3 368.4 极端荒漠区 -
于2021年7月中旬采集沙冬青各器官样品包括叶、茎、根。沿自然降雨梯度(53.5~269.8 mm),降雨量每增加50 mm左右设置1个采样区,共计6个采样地点,每个采样区选取10棵长势较好的沙冬青,按不同器官进行取样。根系利用跟踪法进行采集,首先找到沙冬青主根,然后顺着主根的方向逐渐向下挖掘,用枝剪剪下足够的完整根系,将每株沙冬青的根系样品装入自封袋内做好标记。茎按其东南西北4个方向各取一枝生长良好的枝条。叶选择枝条顶端完全伸展、发育成熟且没有病虫害的叶片,用剪刀剪下后混合装入纸袋,将采集的所有样品带回实验室置于85 ℃杀青30 min,65 ℃烘干48 h,取烘干的植物器官样品(>1 g)用杯式粉碎机进行粗粉碎,然后用冷冻混合球磨仪进行细粉碎,过100目筛,装袋标号,用于实验分析。采用重铬酸钾外加热法测定碳含量;经H2SO4-H2O2法消煮后,用凯氏定氮法测定全氮含量,钼锑抗比色法测定全磷含量,火焰光度计法测全钾含量。
-
通过测定每个器官(根、茎和叶)的元素含量,计算沙冬青各器官元素含量及其化学计量比值的权重,按每个器官的比例加权:灌木的加权平均值=叶的数值×叶的比例 + 茎的数值×茎的比例 + 根的数值×根的比例,得到整株灌木C、N、P、K、C∶N、C∶P、C∶K、N∶P、N∶K和K∶P的加权平均值;对沙冬青整株和各器官的元素含量及其化学计量比进行描述性分析(Descriptive analysis),通过单因素方差分析(One-way ANOVA)检验沙冬青化学计量特征在器官间和不同降雨量间是否存在显著差异;再通过相关分析(Correlation analysis)探讨沙冬青各器官元素及其化学计量比间的相互关系;采用 GLM( General Linear Model) 模型分析器官和降雨量对沙冬青化学计量特征的主因子效应和交互作用。数据的处理作图和统计分析利用Excel 2016、SPSS23.0和R软件。
-
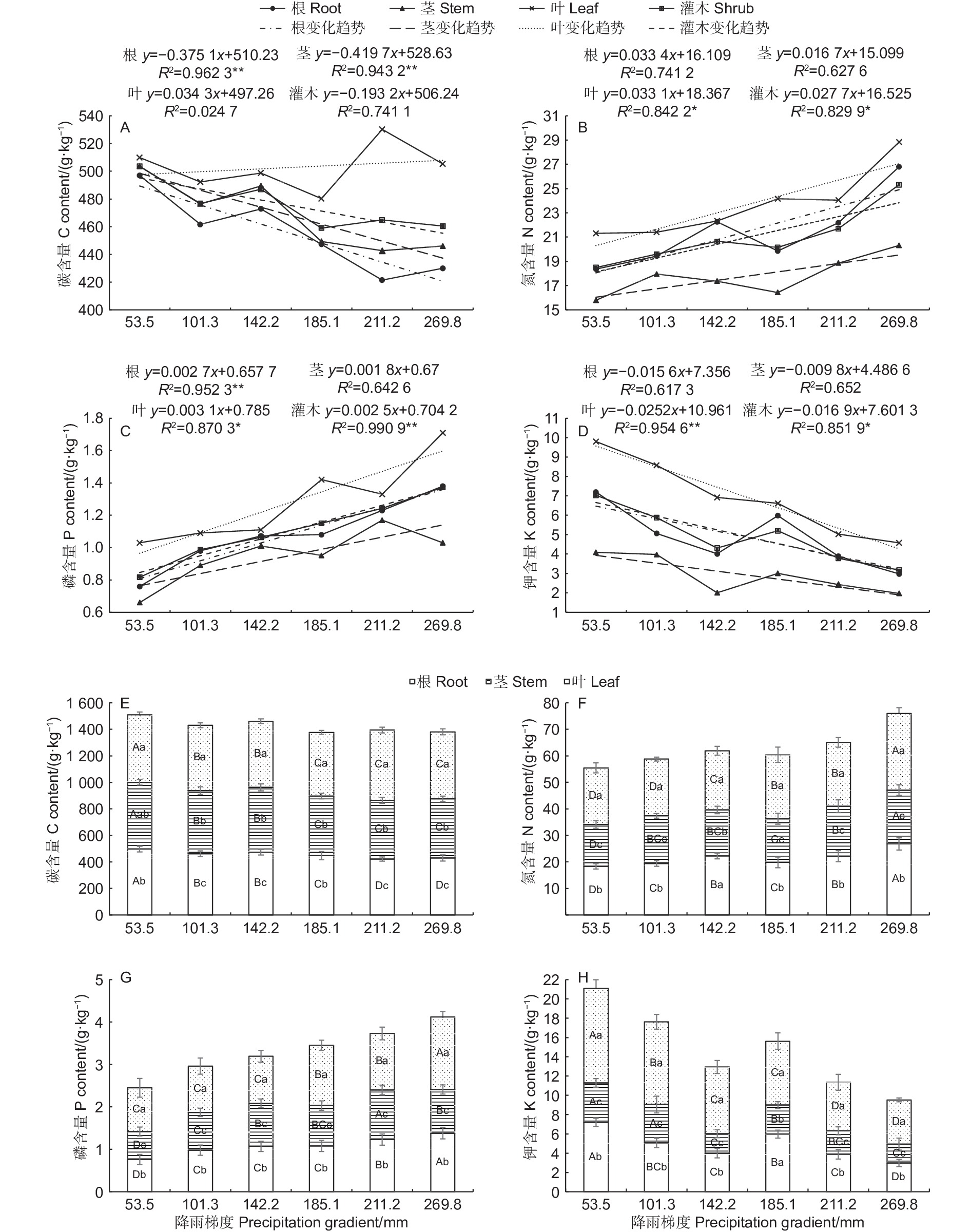
沿降雨梯度的增加,沙冬青各器官的N、P均呈上升趋势,而K呈下降趋势,其中叶和整株灌木中的N、P以及根中的P均呈显著的线性上升趋势,叶和整株灌木中的K均呈显著线性下降趋势,然而叶中的C含量却不随降雨梯度的变化而变化,但根和茎中的C呈极显著线性下降趋势。沿降雨梯度,叶和根的N、P、K的变化速度要快于茎的变化速度(图1ABCD)。不同降雨条件下沙冬青各器官 C、N、P、K 含量变化特征存在一定的差异性(图1EFGH),沙冬青C在各器官中的含量表现为:叶>茎>根,除53.5 mm和185.1 mm外,其余降雨梯度下的C 含量在各器官中差异均显著(p<0.05)。不同降雨条件下元素N、P、K含量的分配情况均为:叶>根>茎,其中N、P、K元素在不同器官中的含量,除了142.2 mm降雨梯度下的N和185.1 mm降雨梯度下的K外,均差异明显(p<0.05),在整个自然降雨梯度上,叶片的C、N、P、K普遍地高于根和茎。沙冬青器官C含量在不同降雨量地区间的变异较小,而N、P、K的变异系数相对较大,其中K的变异程度最大。不同降雨条件下沙冬青各器官营养元素累积量间存在差异,但各元素在器官间分配比例较均衡,其中沙冬青地下部分(根)C、N、P、K含量分别占全株植物元素含量的32.18%、34.18%、32.74%和32.53%,地上部分(叶和茎) C、N、P、K含量分别占全株植物元素含量的68.65%、65.98%、67.48%和65.91%,说明地上部分元素含量高于地下部分。

图 1 不同降雨梯度下沙冬青各器官 C、N、P、K 元素含量(ABCD)及分配比例(EFGH)
Figure 1. C,N,P,K concentrations and its proportion in the organs of A.mongolicus in different precipitation gradients
-
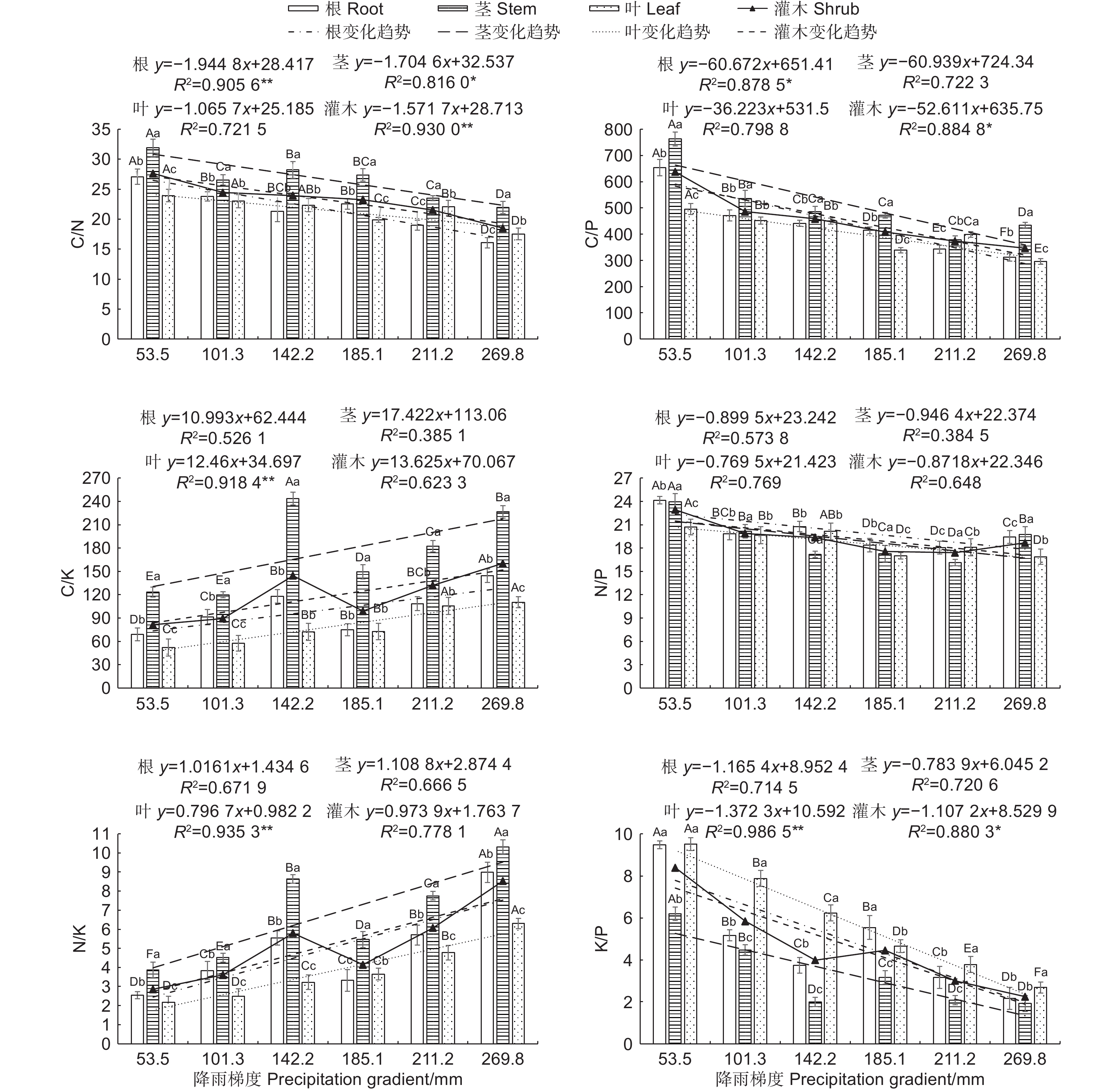
在不同降雨梯度下,沙冬青各器官C∶N、C∶P、C∶K、N∶P、N∶K和K∶P的数值大小的描述性统计值可见图2,C∶N在茎中最大,且显著高于叶和根(p<0.05),C∶K、N∶K在各器官中的变化规律与C∶N相同均表现为:茎显著高于叶和根 (p<0.05)。除211.2 mm外,其余降雨梯度下的C∶P 在茎中最大,且显著高于叶和根(p<0.05),除101.3 mm和142.2 mm外,其余降雨梯度下的C∶P 在各器官中差异均显著(p<0.05)。各器官中N∶P比值均大于16。K∶P 在各器官中的变化范围为3.30~5.79,除53.5 mm和269.8 mm外,其余降雨梯度下的K∶P 在各器官中差异均显著(p<0.05)。器官C、N、P、K的化学计量比值在不同降雨量地区间存在差异,沿降雨梯度的增加,沙冬青整株灌木和各器官的C∶N、C∶P、N∶P、K∶P均呈下降趋势,而C∶K、N∶K呈上升趋势,其中根的C∶N、C∶P比值、茎的C∶N比值、叶的K∶P比值以及整株灌木的C∶N、C∶P、K∶P均呈显著的线性下降趋势,然而叶的C∶K、N∶K呈显著线性上升趋势。各器官中的N∶P的比值在不同降雨梯度间相对比较稳定。

图 2 不同降雨梯度下沙冬青各器官元素化学计量比
Figure 2. Elemental stoichiometry in the organs of A.mongolicus in different precipitation gradients
-
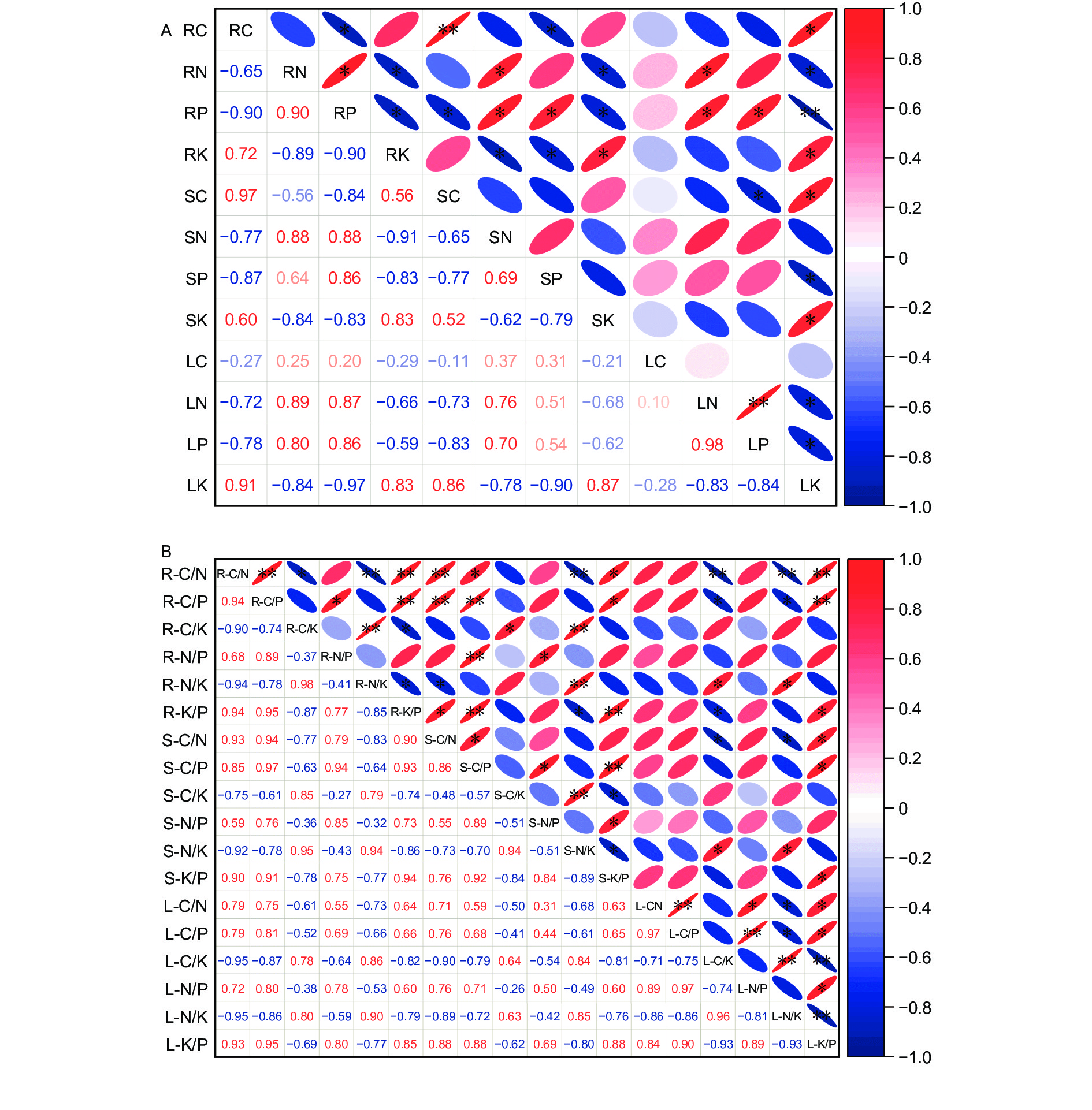
沙冬青各器官元素含量相关分析结果表明(图3A),根的C含量与根和茎的P含量呈显著的负相关关系(p<0.05),与茎的C含量和叶的K含量呈显著的正相关关系(p<0.05),根的N含量与根、茎、叶的K含量呈显著的负相关关系(p<0.05),与根的P含量和茎、叶的N含量呈显著的正相关关系(p<0.05),根的P含量与根、茎的K含量和茎的C含量呈显著的负相关关系(p<0.05),与叶的K含量呈极显著的负相关关系(p<0.01),与茎、叶的N、P含量呈显著的正相关关系(p<0.05),根的K含量与茎的N、P含量呈显著的负相关关系(p<0.05),与茎、叶的K含量呈显著的正相关关系(p<0.05),叶的K含量与茎的P含量和叶的N、P含量呈显著的负相关关系(p<0.05),与茎的C、K含量呈显著的正相关关系(p<0.05),茎的C含量与叶的P含量呈显著的负相关关系(p<0.05)。沙冬青根、茎、叶中的4种元素在器官间和器官内的66个元素组对中,有29对达到显著水平,占43.94%,其中,根与茎的组对中有5对正相关和6对负相关,根与叶的组对中有5对正相关和2对负相关,茎与叶的组对中有2对正相关和2对负相关,根与根的组对中有1对正相关和3对负相关,叶与叶的组对中有1对正相关和2对负相关,茎与茎的组对中均无统计学意义上的相关性(p>0.05),这说明在降雨量变化的背景下,沙冬青器官间的养分协同性远高于器官内部。

图 3 沙冬青各器官元素含量(A)和化学计量学比(B)的相关性
Figure 3. Correlation analysis of element contents and stoichiometric ratio in organs of A.mongolicus
各器官元素化学计量比的相关分析结果表明(图3B),沙冬青6种化学计量比在器官间和器官内的153个化学计量比组对中,有61对达到显著水平,占39.87%,其中,根与茎的组对中有14对正相关和3对负相关,根与叶的组对中有5对正相关和5对负相关,茎与叶的组对中有5对正相关和2对负相关,根与根的组对中有5对正相关和4对负相关,茎与茎的组对中有5对正相关和2对负相关,叶与叶的组对中有7对正相关和4对负相关,这说明在降雨量变化的背景下,沙冬青器官间的化学计量比协同性略高于器官内部。
-
沙冬青C、N、P、K元素含量及其化学计量比受器官和降雨量单因素及双因素的交互影响程度各不相同(表2) 。器官和降雨量对4种化学元素含量的影响均达到了极显著水平(p<0.01),且降雨量对这4种元素含量的影响更大。器官对C∶K的影响达到了显著水平(p<0.05),且对其它元素化学计量比均达到了极显著水平(p<0.01),降雨量对N∶P的影响不显著(p>0.05),但对其它元素化学计量比均达到了极显著水平(p<0.01),其中降雨量对C∶N、C∶K、N∶K的影响更大,而C∶P、N∶P、K∶P主要受器官的影响更大。生境和器官的交互作用对C、C∶N、K∶P的影响均达到了极显著水平(p<0.01),且对N的影响达到了显著水平(p<0.05),但对P、K、C∶P、C∶K、N∶P、N∶K的影响均不显著(p>0.05)。
表 2 碳、氮、磷、钾元素含量及其化学计量特征影响的 GLM 分析
Table 2. GLM analysis of C, N, P,K contents and their stoichiometric characteristics
自变量
Variables器官
Organ降雨量
Precipitation交互作用
Interaction碳含量 C content 22.78** 90.75** 10.03** 氮含量 N content 30.04** 94.53** 2.54* 磷含量 P content 12.39** 17.77** 1.39 钾含量 K content 7.98** 31.57** 0.82 C/N 76.00** 139.00** 6.41** C/P 20.78** 11.15** 1.78 C/K 2.83* 13.35** 0.62 N/P 10.36** 2.55 1.47 N/K 6.77** 8.31** 0.52 K/P 91.66** 58.38** 5.32** -
水分是限制干旱区植物生长与生存的重要因素,在植物整个生长过程中对于体内的物质的运输与代谢、优化自身资源配置等方面起着十分重要的作用。在长期的自然选择中,植物会通过改变自身各器官对养分吸收差异来适应环境的变化和不同环境带来的胁迫,从而影响植物器官化学计量特征[18]。本研究中,不同降雨量条件下沙冬青叶C、N平均含量为502.76 ± 16.99 g·kg−1、23.68 ± 2.81 g·kg−1,大于全球陆生植物叶片C(464.00 g·kg−1、20.60 g·kg−1)[19]、中国东部南北样带(480.10 g·kg−1、18.30 g·kg−1)[20]、阿拉善 (379.01 g·kg−1、10.65 g·kg−1 )[21]、东阿拉善-西鄂尔多斯( 435.07 g·kg−1、23.40 g·kg−1)[22]等地区,且叶N比中国陆地植物(19.72 g·kg−1)[23]高20.08%,但比中国草地生态系统(27.60 g·kg−1)[24]、黄土高原(24.10 g·kg−1)[25]、北方典型荒漠(24.40 g·kg−1)[26] 分别低14.20%、1.74%、2.95%。沙冬青茎C平均含量为467.92 ± 25.55 g·kg−1,比东阿拉善-西鄂尔多斯(460.90 g·kg−1)[22] 高1.52%;根系C平均含量为455.02 g·kg−1,比黄土丘陵(285.16 g·kg−1)[27]、东阿拉善-西鄂尔多斯(418.96 g·kg−1)[22]分别高59.57%、8.61%。茎N平均含量为17.78 ± 1.65 g·kg−1,比东阿拉善-西鄂尔多斯(12.99 g·kg−1)[22] 高36.87%,根系N平均含量为21.48 ± 3.03 g·kg−1,比中国陆生植物(7.55 g·kg−1)[23]、黄土丘陵(5.79 g·kg−1)[27]、东阿拉善-西鄂尔多斯(15.07 g·kg−1)[22]分别高184.50%、270.98%、42.53%。沙冬青叶片P、K平均含量分别为1.28 ± 0.26 g·kg−1、6.92 ± 2.01 g·kg−1,均低于中国陆生植物叶片P(1.51 g·kg−1)和K(15.09 g·kg−1)的平均水平[23],其中叶P均值低于全球陆地植物(1.99 g·kg−1)[19]、中国东部南北样带(2.00 g·kg−1)[20]、黄土高原(1.60 g·kg−1)[25] 、北方典型荒漠(1.74 g·kg−1)[26]等地区,分别低35.68%、36.00%、20.00%、26.44%,而比阿拉善(1.04 g·kg−1)[21]、东阿拉善-西鄂尔多斯(1.18 g·kg−1)[22]、全国常绿木本植物(0.96 g·kg−1)[28]高23.08%、8.47%、33.33%;叶片K平均含量6.92 g·kg−1低于全国阔叶树(8.95 g·kg−1)和灌丛(8.38 g·kg−1)的平均水平[29]。茎、根的P平均含量分别为0.95 ± 0.17 g·kg−1、1.08 ± 0.21 g·kg−1,比东阿拉善-西鄂尔多斯(1.03 g·kg−1、1.10 g·kg−1)[22] 低7.77%、1.82%,其中根系P的平均含量比中国陆生植物(0.80 g·kg−1)[23]高35.00%。与其它研究区相比,沙冬青各器官C、N含量较高,而P、K含量偏低。元素C主要在叶中富集,通常叶片C含量越高,光合速率越低,生长速率越慢,对外界恶劣环境的防御能力越强[30],沙冬青作为荒漠区常绿阔叶树种,其生长相对较缓慢,通过光合作用为自己制造足够的有机物质,其作为豆科植物,根系具有固氮作用,可以满足植物的生长发育与更新修复,然而磷和钾的含量低可能是导致沙冬青种群数量小的原因之一。豆科植物主要对 N 元素有固定和富集的作用,而对 P元素的影响较小,但是不同降雨梯度下沙冬青各器官的 P含量比N含量的变异程度大,植物器官化学计量特征之间的权衡关系反映了植物在不同生境下获取资源与分配养分的调控策略。沿自然降雨梯度,沙冬青的根、茎、叶之间主要表现为协同关系,氮和磷含量在各器官中均随着降雨量的增大而增加,这表明干旱胁迫减弱,伴随着土壤含水量的增加,这使得可被植物利用的养分比例提高[31],因此沙冬青的根对水分和养分的获取能力增强,然后通过茎运输到叶片,提高了植物的光合速率,同时叶片中较高的氮磷含量可以更好的维持植物的光合作用,从而可以保障植物根和茎生长的物质能源,表明沙冬青根茎叶之间存在着同时投资的生态策略。
-
氮、磷、钾是影响植物生长与发育的限制性元素,其生态化学计量比特征对植物生长发育和养分供给状况具有指示作用,其中叶片C∶N 和C∶P的比值越大,意味着植物吸收营养化学元素时所能同化C的能力越强,反映了植物对营养元素的吸收与利用效率,大量研究表明元素化学计量特征与植物生长速率有很强的相关性[32],基于此产生了生长速率理论,一般认为生长速率较快的植物和新陈代谢速率快的器官具有低的C∶P和N∶P,而P的含量呈增加趋势[33-34]。本研究中沙冬青不同器官,叶片的C∶N、C∶P、C∶K、N∶P和N∶K的比值均显著低于根和茎,表明根、茎对 N、P、K 的利用率高于叶片,来维持基本代谢活动,而叶片中的N、P、K含量均显著高于根和茎,这是因为我们在植物生长旺盛期对沙冬青的各器官样品进行了采集测试,此时为了保证叶片较快的新陈代谢速率,植物将体内更多的养分分配给叶片,表明了不同器官元素含量及其化学计量比特征在一定程度上符合生长速率假说。王绍强等[35]研究表明在低营养环境下,植物生长相对较缓慢,但其对营养元素的利用效率较高,因此沿降雨梯度的增加,伴随着干旱胁迫的减轻,沙冬青为了适应环境的变化和维持自身生长,其整株灌木和各器官的C∶N、C∶P、N∶P、K∶P均呈下降趋势,而C∶K、N∶K呈上升趋势。在沙冬青的生长季叶片生长较快,养分供给充足,而根和茎与叶相比处于低营养环境中,根和茎的生长较慢,所以对养分利用的利用效率高于叶片。N∶P阈值假说[3,36-37]认为叶片中的N∶P比值可以指示植物生长活动受限制的元素,一般植物N∶P比值小于14倾向于受氮限制,而 N∶P比值大于16则倾向于受磷限制,介于14和16之间受N和P的共同限制。2003年Olde等[37]提出当 N∶P小于14.5、N∶K小于2.1时,植物倾向于受氮限制,当N∶P大于14.5、K∶P大于3.4 时,植物倾向于受磷限制或磷和氮共同限制,当N∶K大于2.1、K∶P小于3.4时,植物倾向于受钾限制或钾和氮共同限制。在不同的降雨量条件下,沙冬青叶的N∶P、N∶K、K∶P比值范围分别为16.86~20.69、2.17~4.78、3.78~9.51,其N∶P比值均大于16临界值,N∶K比值均大于2.1临界值,K∶P比值均大于3.4临界值,说明沙冬青生长过程中主要受磷的限制。
-
植物各器官结构功能和生长环境的差异性导致C、N、P、K元素含量及其化学计量比变化显著[38-40]。本研究中,沙冬青各器官间的化学元素含量差异显著,干旱胁迫的生境也会影响植物的生长发育、生理代谢以及光合和呼吸作用,进而对植物的生物量积累、养分吸收与利用产生影响[41-43]。沙冬青通过叶进行光合作用获取足够的碳元素,各生境下水分因子的差异会导致植物的光合速率和吸收利用养分的能力发生变化,故降雨对沙冬青元素含量的影响较大。沙冬青各器官中的C、N 含量均较高,首先C含量升高表明其光合速率较低,植物生长相对缓慢,但对外界恶劣环境的防御能力增强,其次在干旱荒漠区植物的生长易受水分胁迫,为了保持细胞渗透平衡,需要增加游离氨基酸[39-40],提高叶片 N 含量,从而可以能更好地适应干旱环境。沙冬青各器官中的 P、K 含量普遍偏低,一方面是是因为沙冬青自然分布区风蚀作用强烈,致使土壤中黏粒和粉粒物质会被大量吹失,造成有机质及其养分会大量流失,植物根系可以吸收和利用的养分不足;另一方面K是干旱气候条件下植物抵抗逆境的重要营养元素,沿降雨梯度,随着降雨量的增加,各器官中的K含量呈下降趋势,即在干旱胁迫下沙冬青通过提高K含量,调控植物水分传导,可以显著提高叶片的光合和蒸腾速率,维持渗透平衡,增强酶活性,增强抵抗力[44],沙冬青靠较强的K吸收能力来抵御干旱环境的伤害。最后干旱胁迫可能导致植物产生栓塞现象[45],造成P、K的运输通道堵塞,从而植物整体P、K含量偏低,沿降雨梯度,随着干旱胁迫的加剧,沙冬青通过减少各器官中的N、P含量,增加K含量提升植物的抗旱性,来维持基本代谢活动。不同降雨地区间沙冬青的根茎叶N∶P比值均大于16,N∶K比值均大于2.1,表明其生长是受到磷的限制,在这个生长条件下,沙冬青要依靠自身遗传特性来调节器官内氮、磷、钾含量及其化学计量比的分配比例,以适应不同的生存条件。叶片和根系分别是植物地上部分和地下部分代谢活性较强的器官,因此对环境的变化最为敏感[46]。植物叶片通常比茎和根具有更高的养分含量,用以维持叶片高的生理生态活性[26, 47],叶片是植物的同化和储藏养分的主要器官,而茎是水分和养分的运输通道,主要负责将营养元素从根部运输到叶片,所以储存养分最少,根是吸收和转运营养元素的器官,贮存的养分向上运输主要用于支持植物的生长发育[48],因此沙冬青N、P、K元素含量在根茎叶中的顺序均表现为叶>根>茎。不同干旱胁迫条件下沙冬青的同一器官间的 C 含量变异程度均较小,这主要由于C在各器官中的含量均很高,作为植物的结构性物质在植物体内主要起骨架作用,且不直接参与植物生长和繁殖等各项生产活动,在植物体内有较好的稳定性。而沙冬青根茎叶中的N、P、K 含量在不同降雨量条件下差异性显著,其中 N、P、K含量均在叶片中最高,根次之,最后是茎,说明N、P、K等大量元素主要在叶中富集,这主要是因为采样时间为植物生长旺盛季,成熟叶片的光合与蒸腾作用强烈,需要大量的营养元素合成植物所需的叶绿素、蛋白质、核酸和酶[49] ,因此叶片的 N、P、K 元素得到优先分配,由根部吸收通过茎向上运输至叶片,使 N、P、K 在叶片出现富集。这种养分在各器官的差异分配,与植物器官内部结构以及组织功能分化有密切的关系,同时反映资源分配模式和植物生长策略[2,50],而降雨量的改变会潜在的调节和影响这种分配过程。
-
植物器官生态化学计量特征为探索有机体的元素组成差异、器官功能的内在机制以及器官响应环境变化的权衡策略提供了研究方法。沙冬青各器官元素含量及其化学计量比在荒漠生境中存在复杂的相互作用,植物各器官对氮、磷、钾的吸收与利用的比例是不同的,主要受自身遗传生理功能的影响,植物将更多的养分分配给生长旺盛的器官,沙冬青各器官的N、P、K元素含量分配情况均为:叶>根>茎,这种3大营养元素在叶片中的富集,可以提高沙冬青的光合速率,为植物的生长与繁殖提供充分的物质保障。与其它研究区相比,沙冬青C、N含量较高,而P、K含量偏低,地上部分元素含量高于地下部分,且地上器官叶和茎与地下器官根养分分配具有一致性,功能联系密切,叶与根的N、P、K 以及茎与根的C、N、P、K均表现为显著正相关。根与茎之间的生长速率和养分利用效率是相互联系、相互影响的,具有储存、运输和起机械支撑作用的根与茎器官之间的C∶N、C∶P、C∶K、N∶P、N∶K、K∶P均呈显著正相关。不同降雨量地区间沙冬青各器官的N:P比值均大于16则生长主要受P限制,在养分供应不足的情况下,植物自身也通过养分在体内的转移满足代谢活跃器官的生长需要,这是植物维持生长与发育,高效合理利用养分的一种重要权衡机制。沿自然降雨梯度,沙冬青的根、茎、叶之间主要表现为协同关系,存在着同时投资的生态策略,N和P含量在各器官中均随着降雨量的增加呈上升趋势,而K均呈下降趋势。因此,本研究从化学计量学的角度了解该物种的生长特征和养分利用策略,揭示植物在适应干旱环境及自身生长发育需要的过程中的有机元素分配格局,为探讨其濒危机制提供科学依据。
干旱区降雨梯度对沙冬青根-茎-叶生态化学计量特征的影响
Effects of Precipitation Gradient on Ecological Stoichiometric Characteristics in Root-Stem-Leaf of Ammopiptanthus mongolicus in Arid Region
-
摘要:
目的 水分是限制干旱区植物生长与生存的重要因素,为探明降水变化对荒漠植物化学计量特征的影响, 以及C、N、P和K元素在植物各器官中的分配规律。 方法 以国家二级重点保护的珍稀濒危植物且是荒漠区唯一常绿阔叶灌木沙冬青为研究对象,分析沿自然降雨梯度下,沙冬青根-茎-叶的元素含量及其化学计量比的变化特征,了解其生长的适应机制和生存策略。 结果 沙冬青各器官C含量表现为叶>茎>根,而N、P、K营养元素含量均表现为叶>根>茎,N、P、K三大营养元素在叶片中的富集,可以提高沙冬青光合速率,为植物的生长与繁殖提供充分的物质保障,反映了资源分配模式和植物生长策略。沿自然降雨梯度,沙冬青的根、茎、叶之间主要表现为协同关系且存在着同时投资的生态策略,N和P含量在各器官中均随着降雨量的增加呈上升趋势,而K均呈下降趋势,沙冬青通过提高各器官K含量和降低N、P含量来应对干旱胁迫的环境。器官间的养分协同性远高于器官内部,各器官中的N:P在不同降雨梯度间相对比较稳定,说明沙冬青以保守的养分利用方式抵消元素含量随环境的变化,且N:P比值均大于16则生长主要受P限制。 结论 各器官N、P、K 化学计量特征沿降雨梯度具有相同趋势,但在不同器官间呈现显著差异,而化学计量的灵活性是实现植物自我调节的重要方式,有助于植物在不同环境条件影响下维持自身正常的生命活动。 Abstract:Objective Water is an important factor limiting the growth and survival of plants in arid areas. The effects of precipitation changes on the desert Phytochemistry metrological characteristics, and the distribution of C, N, P and K elements in various plant organs are explored. Method Based on the second-class of rare-endangered plants in China, Ammopiptanthus mongolicus, the only evergreen broad-leaved shrub in desert area, this paper analyzed the ecological stoichiometric characteristics of roots, stems and leaves of A. mongolicus under different precipitation gradients. Results The results showed that the C content in the leaves was the most, followed by stems and roots, the nutrient contents of N、P and K in the leaves were also the most, followed by the roots and stems. The enrichment of the three nutrient elements N, P and K in the leaves could improve the photosynthetic rate of A. mongolicus and provide enough material support for plant growth and reproduction, reflecting the resource allocation pattern and plant growth strategy. Along the natural precipitation gradient, the roots, stems and leaves of A. mongolicus mainly showed a synergetic relationship. The content of N and P in each organ showed an upward trend with the increase of precipitation, while K showed a downward trend. A. mongolicus responded to the drought stress environment by increasing the content of K in each organ and reducing the content of N and P, indicating that there is an ecological strategy of simultaneous investment between the roots, stems and leaves of A. mongolicus. The ratio of N: P in each organ was relatively stable among different precipitation gradients. A. mongolicus counteracts the changes of element content with the environment by coordinating nutrient conservation strategies in a conservative way. The nutrient synergy between organs was much higher than that within organs. The N:P ratios of organs were all greater than 16, indicating that the growth of A. mongolicus was mainly limited by P. Conclusion The stoichiometric characteristics of N, P, K in each organ have the same trend along the rainfall gradient, but show significant differences among different organs. The flexibility of stoichiometry is an important way to achieve plant self-regulation, which helps plants maintain their normal life activities under different environmental conditions. -
图 1 不同降雨梯度下沙冬青各器官 C、N、P、K 元素含量(ABCD)及分配比例(EFGH)
Figure 1. C,N,P,K concentrations and its proportion in the organs of A.mongolicus in different precipitation gradients
图 2 不同降雨梯度下沙冬青各器官元素化学计量比
Figure 2. Elemental stoichiometry in the organs of A.mongolicus in different precipitation gradients
图 3 沙冬青各器官元素含量(A)和化学计量学比(B)的相关性
Figure 3. Correlation analysis of element contents and stoichiometric ratio in organs of A.mongolicus
表 1 研究区位置与环境特征
Table 1. Sample location and environmental characteristics
站点各样地
Sites经纬度
Longitude, Latitude海拔
Altitude/m年平均降雨量
Mean annual
precipitation/mm年平均气温
Mean annual
temperature/℃日照时数
Mean annual sunshine
duration/h生境类型
Habitat
type鄂托克旗 107.98° E,39.11° N 1 380.3 269.8 7.2 2 846.1 荒漠草原区 阿拉善左旗 105.67° E,38.83° N 1 561.4 211.2 7.9 3 099.2 荒漠草原区 景泰 103.83° E,37.51° N 1 799.9 185.1 8.8 2 725.7 典型荒漠区 磴口 106.42° E,40.18° N 1 039.3 142.2 8.0 3 211.7 典型荒漠区 乌拉特后旗 106.36° E,41.74° N 1 126.3 101.3 4.9 2 968.4 典型荒漠区 额济纳旗 102.51° E,41.94° N 1 051.9 53.5 8.7 3 368.4 极端荒漠区  下载: 导出CSV
下载: 导出CSV
表 2 碳、氮、磷、钾元素含量及其化学计量特征影响的 GLM 分析
Table 2. GLM analysis of C, N, P,K contents and their stoichiometric characteristics
自变量
Variables器官
Organ降雨量
Precipitation交互作用
Interaction碳含量 C content 22.78** 90.75** 10.03** 氮含量 N content 30.04** 94.53** 2.54* 磷含量 P content 12.39** 17.77** 1.39 钾含量 K content 7.98** 31.57** 0.82 C/N 76.00** 139.00** 6.41** C/P 20.78** 11.15** 1.78 C/K 2.83* 13.35** 0.62 N/P 10.36** 2.55 1.47 N/K 6.77** 8.31** 0.52 K/P 91.66** 58.38** 5.32**
下载: 导出CSV
-
[1] MASSON-DELMOTTE VALÉRIE, ZHAI P, PIRANI A. Climate change 2021: The physical science basis: summary for policymakers: working group I contribution to the sixth assessment report of the intergovernmental panel on climate change[M]. IPCC: Geneva, Switzerland, 2021. [2] CHAPIN F S. The mineral nutrition of wild plants[J]. Annual review of ecology and systematics, 1980, 11(1): 233-260. doi: 10.1146/annurev.es.11.110180.001313 [3] GÜSEWELL S. N: P ratios in terrestrial plants: variation and functional significance[J]. New Phytologist, 2004, 164(2): 243-266. doi: 10.1111/j.1469-8137.2004.01192.x [4] ELSER J J, STERNER R W, GOROKHOVA E,et al. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2000, 3(6): 540-550. doi: 10.1046/j.1461-0248.2000.00185.x [5] TORSTEN I. Mineral nutrient requirements of pinus silvestris and picea abies seedlings[J]. Physiologia Plantarum, 1979, 45(4): 373-380. doi: 10.1111/j.1399-3054.1979.tb02599.x [6] ARONSSON A, ELOWSON S. Effects of irrigation and fertilization on mineral nutrients in Scots pine needles[J]. Ecological Bulletins, 1980, 32(4): 219-228. [7] 王 飞, 郭树江, 张卫星, 等. 民勤荒漠区主要灌木叶片C、N、P化学计量特征的季节变化[J]. 西北植物学报, 2020, 40(1):121-129. [8] 陈 婵, 王光军, 赵 月, 等. 会同杉木器官间 C、N、P 化学计量比的季节动态与异速生长关系[J]. 生态学报, 2016, 36(23):7614-7623. [9] 张天霖, 邱治军, 吴仲民, 等. 粤北针阔混交林不同器官碳氮磷钾的生态化学计量特征[J]. 林业科学研究, 2021, 34(2):149-157. doi: 10.13275/j.cnki.lykxyj.2021.02.016 [10] SARDANS J, RIVAS-UBACH A, PEñUELAS J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: a review and perspectives[J]. Biogeochemistry, 2011, 111(1-3): 1-39. [11] 周 鹏, 耿 燕, 马文红, 等. 温带草地主要优势植物不同器官间功能性状的关联[J]. 植物生态学报, 2010, 34(1):7-16. doi: 10.3773/j.issn.1005-264x.2010.01.003 [12] ZHANG J, HE N, LIU C,et al. Allocation strategies for nitrogen and phosphorus in forest plants[J]. Oikos, 2018, 127(10): 1506-1514. doi: 10.1111/oik.05517 [13] 戚燕强, 郑 听, 张子雄, 等. 白芨不同器官干物质积累及氮、磷、钾化学计量特征[J]. 江苏农业科学, 2022, 50(3):157-162. [14] 王 凯, 高 爽, 刘焕彬, 等. 施氮与增水对杨树幼苗不同器官碳氮磷化学计量的影响[J]. 生态学杂志, 2021, 40(12):3870-3880. doi: 10.13292/j.1000-4890.202112.020 [15] 段义忠, 王 驰, 王海涛, 等. 不同气候条件下沙冬青属植物在我国的潜在分布—基于生态位模型预测[J]. 生态学报, 2020, 40(21):7668-7680. [16] 牛若恺, 高润红, 侯艳青, 等. 气候变化下沙冬青适宜分布区预测[J]. 西北林学院学报, 2021, 36(1):102-107. doi: 10.3969/j.issn.1001-7461.2021.01.14 [17] 李昌龙, 尉秋实, 李爱德. 孑遗植物沙冬青的研究进展与展望[J]. 中国野生植物资源, 2004,23(5):21-23. [18] 陈佳瑞, 王国梁, 孟 敏, 等. 干旱胁迫3种灌木不同器官化学计量特征的影响[J]. 应用生态学报, 2021, 32(1):73-81. [19] ELSER J J, FAGAN W F, DENNO R F,et al. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature, 2000, 408(6812): 578-580. doi: 10.1038/35046058 [20] 任书杰, 于贵瑞, 姜春明, 等. 中国东部南北样带森林生态系统102 个优势种叶片碳氮磷化学计量学统计特征[J]. 应用生态学报, 2012, 23(3):581-586. [21] 张 珂, 何明珠, 李新荣, 等. 阿拉善荒漠典型植物叶片碳、氮、磷化学计量特征[J]. 生态学报, 2014, 34(22):6538-6547. [22] 刘 珮, 马 慧, 智颖飙, 等. 9种典型荒漠植物生态化学计量学特征分析[J]. 干旱区研究, 2018, 35(1):207-216. doi: 10.13866/j.azr.2018.01.26 [23] HAN W X, FANG J Y, GUO D L,et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2): 377-385. doi: 10.1111/j.1469-8137.2005.01530.x [24] HE J S, FANG J, WANG Z,et al. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 2006, 149(1): 115-122. doi: 10.1007/s00442-006-0425-0 [25] ZHENG S X, SHANGGUAN Z P. Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China[J]. Trees, 2007, 21(3): 357-370. doi: 10.1007/s00468-007-0129-z [26] 李玉霖, 毛 伟, 赵学勇, 等. 北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究[J]. 环境科学, 2010, 31(8):1716-1725. [27] 张海东, 汝海丽, 焦 峰, 等. 黄土丘陵区退耕时间序列梯度上草本植被群落与土壤C、N、P、K 化学计量学特征[J]. 环境科学, 2016, 37(3):1129-1138. [28] 张 剑, 齐璇璇, 刘 冬, 等. 干旱区湿地芦苇各器官生态化学计量对土壤因子的响应[J]. 生态学杂志, 2021, 40(3):701-711. doi: 10.13292/j.1000-4890.202103.010 [29] 秦 海, 李俊祥, 高三平, 等. 中国660种陆生植物叶片8种元素含量特征[J]. 生态学报, 2010, 30(5):1247-1257. [30] 王凯博, 上官周平. 黄土丘陵区燕沟流域典型植物叶片C、N、P化学计量特征季节变化[J]. 生态学报, 2011, 31(17):4985-4991. [31] 崔宁洁, 刘小兵, 张丹桔, 等. 不同林龄马尾松(Pinus massoniana)人工林碳氮磷分配格局及化学计量特征[J]. 生态环境学报, 2014, 23(2):188-195. [32] ÅGREN G I. The C: N: P stoichiometry of autotrophs theory and observations[J]. Ecology Letters, 2004, 7(3): 185-191. doi: 10.1111/j.1461-0248.2004.00567.x [33] STERNER R W, ELSER J J. Ecological stoichiometry: The biology of elements from molecules to the biosphere[M]. Princeton University Press: Princeton, 2002. [34] NIKLAS K J, COBB E D. N, P, and C stoichiometry of eranthis hyemalis (Ranunculaceae) and the allometry of Plant Growth[J]. American Journal of Botany, 2005, 92(8): 1256-1263. doi: 10.3732/ajb.92.8.1256 [35] 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征[J]. 生态学报, 2008(8):3937-3947. [36] PÉREZ-HARGUINDEGUY N, DÍAZ S, GARNIER E,et al. New handbook for standardized measurement of plant functional traits worldwide[J]. Australian Journal of Botany, 2013, 61: 167-234. doi: 10.1071/BT12225 [37] OLDE VENTERINK H, WASSEN M J, VERKROOST A W,et al. Species richness–productivity patterns differ between N-, P-, and K-limited wetlands[J]. Ecology, 2003, 84(8): 2191-2199. doi: 10.1890/01-0639 [38] 李从娟, 徐新文, 孙永强, 等. 不同生境下三种荒漠植物叶片及土壤C、N、P的化学计量特征[J]. 干旱区地理, 2014, 37(5):996-1004. [39] ELSER J J, FAGAN W F, KERKHOFF A J,et al. Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change[J]. New Phytologist, 2010, 186(3): 593-608. doi: 10.1111/j.1469-8137.2010.03214.x [40] 余 洋, 张志浩, 杨建明, 等. 疏叶骆驼刺叶、根生态化学计量特征对水氮添加的响应[J]. 干旱区研究, 2022, 39(2):551-559. [41] 黄树荣, 谢燕燕, 陈双林, 等. 毛竹林叶片碳氮磷化学计量特征的海拔梯度效应[J]. 竹子学报, 2020, 39(1):73-78. [42] 陈晓萍, 郭炳桥, 钟全林, 等. 武夷山不同海拔黄山松细根碳、氮、磷化学计量特征对土壤养分的适应[J]. 生态学报, 2018, 38(1):273-281. [43] 杨 蕾, 孙 晗, 樊艳文, 等. 长白山木本植物叶片氮磷含量的海拔梯度格局及影响因子[J]. 植物生态学报, 2017, 41(12):1228-238. [44] SARDANS J, PEÑUELAS J. Potassium: a neglected nutrient in global change[J]. Global Ecology and Biogeography, 2015, 24(3): 261-275. doi: 10.1111/geb.12259 [45] 李 荣, 党 维, 蔡 靖, 等. 6 个耐旱树种木质部结构与栓塞脆弱性的关系[J]. 植物生态学报, 2016, 40(3):255-263. [46] SCHREEG L A, SANTIAGO L S, WRIGHT S J,et al. Stem, root, and older leaf N: P ratios are more responsive indicators of soil nutrient availability than new foliage[J]. Ecology, 2014, 95(8): 2062-2068. doi: 10.1890/13-1671.1 [47] 罗 艳, 贡 璐. 塔里木盆地南缘不同生境下芦苇生态化学计量特征[J]. 生态学杂志, 2016, 35(3):684-691. [48] 李合生. 现代植物生理学[M]. 北京: 高等教育出版社, 2002:96-100. [49] KERKHOFF A J, FAGAN W F, ELSER J J, et al. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants[J/OL]. The American Naturalist, 2006, 168(4): E103-E122. [50] FORTUNEL C, FINE P V, BARALOTO C. Leaf, stem and root tissue strategies across 758 neotropical tree species[J]. Functional Ecology, 2012, 26(5): 1153-1161. doi: 10.1111/j.1365-2435.2012.02020.x -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2459
- HTML全文浏览量: 1027
- PDF下载量: 88
- 被引次数: 0
