

 下载:
下载:


-
干旱是植物生长、发育和生产力的主要环境制约因素。由于林木寿命长,难以大规模灌溉,干旱极大地降低了许多森林生态系统的树木生产力和存活率[1]。因此,加快对林木抗旱机制的剖析,提高林木对干旱的适应能力,是干旱土地利用、环境可持续性和提高经济效益的迫切需要。为了适应干旱胁迫,植物已经进化出复杂的机制来感知外部信号,并通过调节基因的表达来协调代谢途径和形态特征[2]。
AP2/ERF家族转录因子调控植物的多种发育过程,在激素调控和逆境应答中发挥重要作用[3]。AP2/ERF家族转录因子在高等植物中调控多种环境胁迫响应过程,如非生物胁迫(冷、热、旱、盐、渗透胁迫)和生物胁迫(食草昆虫和微生物病原体)[4-8]。此外,许多研究表明,过表达AP2/ERF家族转录因子的转基因植物对非生物和生物胁迫的耐受性有所提高[9-11]。例如,对ERF76转基因杨树(Populus simonii × Populus nigra)转录组测序中发现有16个上调转录因子基因和45个胁迫相关基因的表达,提高了ABA(脱落酸)和GA(赤霉素)的生物合成能力,从而增强了杨树对盐胁迫的耐受性[12]。新疆杨(Populus alba var. pyramidalis Bunge) PalERF109的过表达增强了杨树的耐盐性,进一步的分析表明,PalERF109直接上调了一个高亲和力K + 转运体(HKT)基因PalHKT1;2。表明PalERF109通过直接激活PalHKT1;2增强了耐盐性,并扩展了对ERF基因在树木胁迫反应中的作用的理解[13]。在盐和干旱胁迫下转基因ERF38过表达杨树过氧化物酶(Peroxidase, POD)、超氧化物歧化酶(Superoxide dismutase, SOD)、可溶性蛋白含量和脯氨酸含量显著增加,表明ERF38基因的过表达可以提高转基因杨树的耐盐和渗透性[14]。
胡杨(Populus euphratica Oliv.)是分布于我国西北沙漠地区沙漠河岸森林的优势树种,在维持当地沙漠生态系统方面发挥着重要的作用[15]。由于其对极端恶劣环境的良好适应性,胡杨被认为是木本植物非生物胁迫耐受机制研究的典型模式物种[16]。前期通过对胡杨种子耐盐全基因组关联分析(Genome wide association study, GWAS)和转录组分析中获得胡杨PeERF1基因[17],通过亚细胞定位实验发现PeERF1为核定位蛋白,转录自激活实验证明PeERF1基因具有转录自激活活性。耐盐能力分析试验证明胡杨PeERF1基因能够增加转基因植株的耐盐性[18],但其耐旱机制中的作用尚缺乏研究。本研究为探明胡杨PeERF1基因应答干旱胁迫的表达特点,以胡杨为材料研究PeERF1基因在模拟干旱胁迫(PEG)下不同组织的时空表达模式。通过对野生型银腺杨(Populus alba × P. glandulosa ‘84k’, ‘84K’杨)、转PeERF1基因过表达和抑制表达银腺杨进行模拟干旱胁迫,观察PeERF1基因在模拟干旱胁迫下表达模式、表型和生理指标的变化,为进一步探究该基因在植物抗逆中的功能提供依据。
-
本试验以胡杨野生型、84K杨(WT)、35S::PeERF1过表达(PE)和35S::PeERF1-SRDX抑制表达(SE)转基因银腺杨为材料,35S::PeERF1是通过35S强启动子与PeERF1的cDNA融合构建的载体转化84K杨获得的转基因植株,该转化载体可以使目标基因过表达。35S::PeERF1-SRDX是由编码SRDX肽的核酸序列与PeERF1的cDNA融合构建的载体转化84K杨获得的转基因植株,该转化载体可以特异性抑制目标基因的表达,转基因植株由前期研究获得[18]。转基因组培苗保存于中国林业科学研究院林木遗传育种全国重点实验室,在含有约70 mL生根培养基的培养瓶中生长,培养条件为光照强度6 000~8 000 lx、光照/黑暗周期16 h/8 h、平均温度25 ℃、相对湿度60%~70%。
-
本试验在中国林业科学研究院科研温室中进行,84K杨组培苗在组培室生长1个月后,选择生长状态一致且健壮的组培苗移栽(花盆规格:盆高 × 盆口直径=22 cm × 20 cm),基质为草炭土∶珍珠岩=3∶1。在土培1个月后选取生长状态一致且健壮的植株进行模拟干旱处理试验。
胡杨组培苗干旱处理,将在组培室生长1个月且生长状态一致的胡杨幼苗转移到20%的PEG6000溶液中,在处理的0、12和24 h时取根、茎和叶投入液氮后保存−80 ℃冰箱备用,每个处理3次重复,每个重复3株,每个处理共9棵植株。
转PeERF1基因84K杨干旱处理,选择转基因PeERF1基因过表达(PE1、PE2)和抑制表达(SE1、SE2)高表达的各两个株系,以及野生型(WT)试验材料进行模拟干旱胁迫试验,平均分为6组,每组每个株系3株,3组用于20% PEG6000处理7 d,另外3组为水处理作对照。处理后取叶片投入液氮后保存于−80 ℃冰箱备用,每个处理包含9棵植株。
-
处理结束时随机选取各处理各株系6株,测量植株茎基部到茎尖部分为自然株高,株高数据以cm为单位,保留一位小数。取植株地上部分,并用清水、蒸馏水、超纯水依次清洗一遍,擦干水分后,用万分之一天平测定植株鲜质量。
-
对84K杨和转PeERF1基因植株进行干旱胁迫处理7 d,取顶端第2片新鲜叶片用于叶绿素含量的测定。用乙醇法对转基因植株进行叶绿素含量测定[19]。
-
取84K杨和转PeERF1基因植株干旱处理7 d后由上而下第3、4和5片叶子测定过氧化氢(Catalase, CAT)、丙二醛(Malondialdehyde, MDA)和过氧化物酶(Peroxide dismutase, POD)含量。具体操作步骤如下:液氮研磨后,取0.1 g样品,加入1 mL提取液,进行冰浴匀浆:8 000 g,4 ℃离心10 min,取上清,置冰上待测。采用索莱宝公司过氧化氢CAT活性试剂盒[20]、丙二醛MDA活性试剂盒[21]、过氧化物酶POD活性试剂盒[22]分别进行CAT含量、 MDA含量和POD含量测定。
-
根据采用2−△△CT法获得qPCR的差异表达倍数[23]。利用Excel 2018软件将所测得的数据进行成组数据T检验统计分析。本研究中各项指标测定时,测定值重复3次后,用Excel表格和Prism8软件进行分析,组间比较用t值,当* p<0.05时,差异显著;** p<0.01时,差异极显著。
-
为了研究PeERF1在不同组织中的表达模式,我们采用qRT-PCR方法分别检测了其在叶、茎和根中的表达水平。结果表明,PeERF1在胡杨叶、茎和根等不同组织中均有表达(图1A),叶中的表达水平最高,其次是茎和根,其中叶中的表达量是茎的1.2倍,是根的1.24倍。

图 1 PeERF1的时空表达模式分析
Figure 1. Analysis of spatiotemporal expression pattern of PeERF1
为了研究PeERF1在胡杨不同组织干旱胁迫下的时空表达模式,利用qRT-PCR技术检测了20% PEG6000处理0、12和24 h后PeERF1在叶、茎和根中的表达水平,如图1B~D所示,PeERF1基因在胡杨叶片中表达量在干旱胁迫在反应初期(12 h)呈上调趋势,在处理24 h出现下调趋势。而在茎和根中则表现出先下降后上升的趋势。综上可知,当胡杨受到干旱胁迫后12 、24 h时,PeERF1基因均有不同表达,表明PeERF1基因参与植物干旱胁迫的调节过程。
-
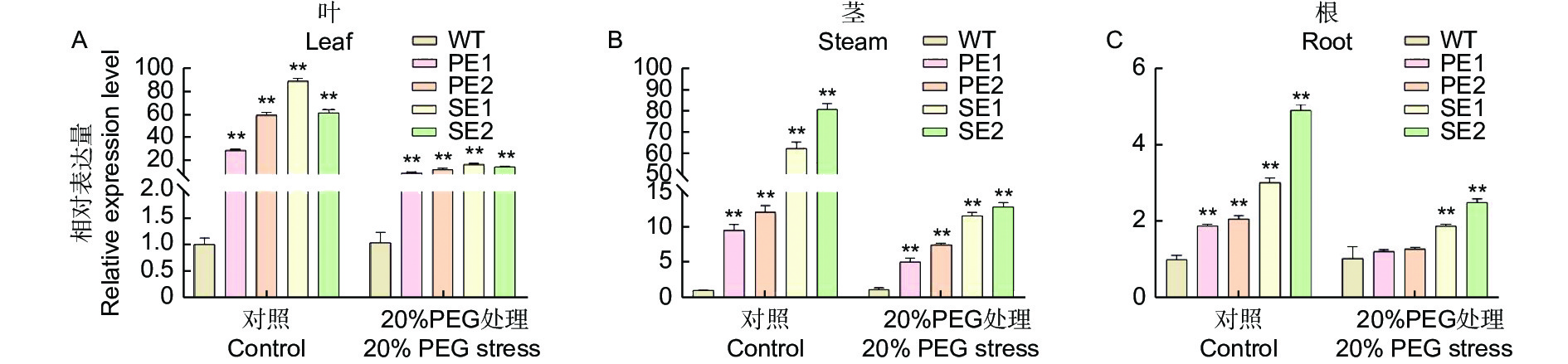
为了研究干旱胁迫下84K杨和转基因PeERF1植株在不同组织中的表达模式,利用qRT-PCR技术检测了20% PEG6000处理7 d后84K杨和转基因PeERF1植株在叶(图2A)、茎(图2B)和根(图2C)中的表达水平。结果显示,在正常状态和干旱胁迫条件下,转基因株系在根、茎和叶中表达量均高于WT,且在叶片和茎中表现差异极显著(图2)。表明PeERF1基因在一定程度上能够提高杨树的耐旱性。

图 2 干旱胁迫处理转PeERF1基因不同组织部位表达模式分析
Figure 2. Analysis on the expression patterns of different tissue sites of PeERF1 transgenic lines under drought stress
-
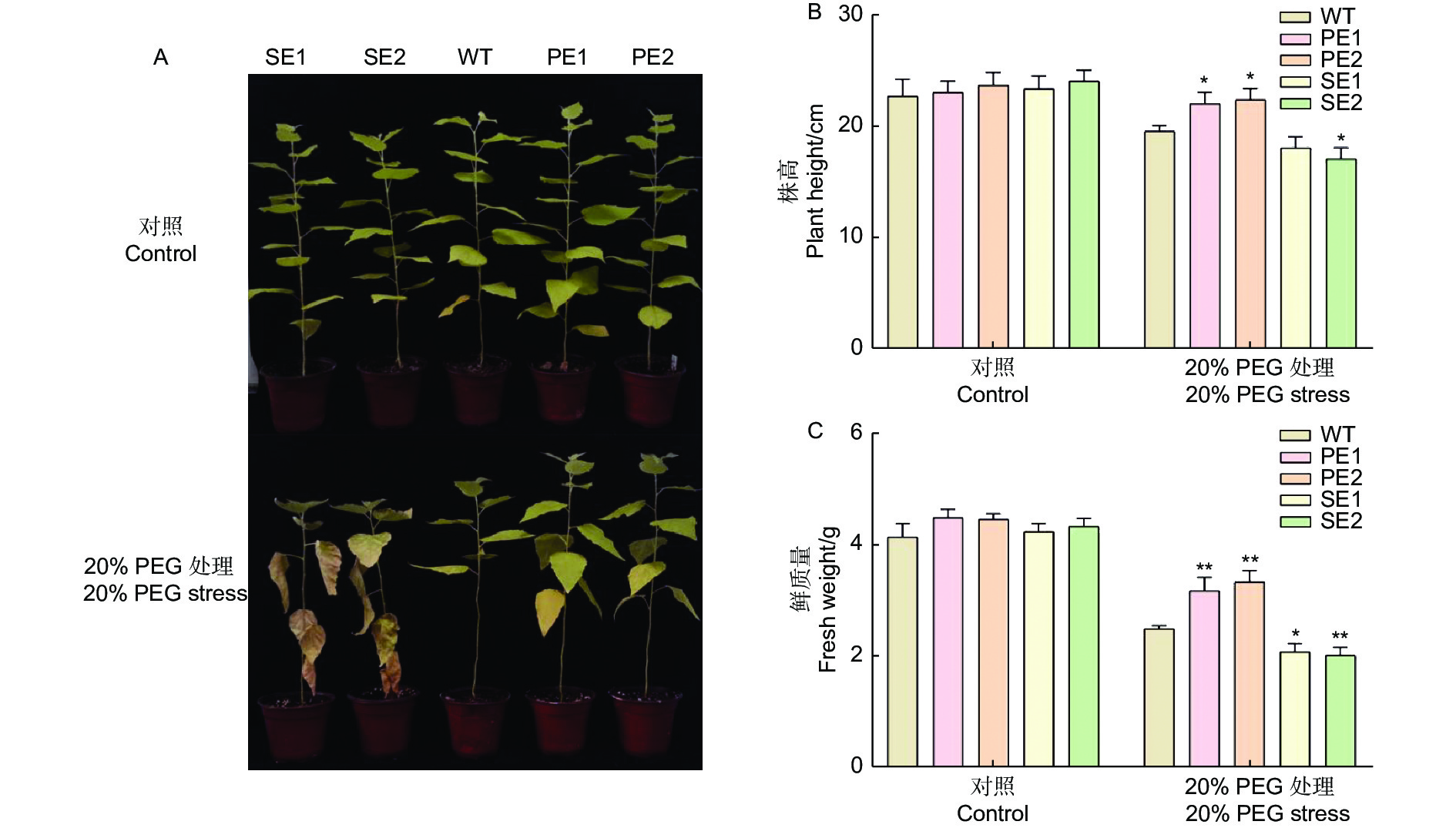
逆境胁迫严重影响着植物的生长发育,在叶片表型方面表现尤为显著。与对照组相比,在正常生长状态下,WT和转基因株系的生长形态没有表现出明显的变化(图3A)。干旱胁迫后,过表达PE转基因杨树生长状态稍好,表现出较强的耐旱性,相反WT和抑制表达转基因植株SE出现枯萎表现出不耐受,SE转基因株系叶片完全枯萎甚至死亡。

图 3 干旱胁迫对转PeERF1基因84K杨生长状态的影响
Figure 3. The effect of drought stress on the growth state of transgenic Populus alba × Populus glandulosa with PeERF1 gene
干旱胁迫处理一周后分别对处理组和对照组植株的株高(图3B)和鲜质量(图3C)进行测量后发现,与对照处理相比,胁迫后对照(WT)、过表达(PE1、PE2)和抑制表达(SE1、SE2)植株的株高和鲜质量均下降,各株系株高显著下降,分别为对照组的86.0%、95.6%、94.3%、77.1%和70.8%,植株鲜质量也有相同趋势,下降呈极显著。由此可知,干旱胁迫会对植株生长量的增加、生物量的积累产生不利影响。与对照WT植株相比,转基因植株(SE)各株系在干旱胁迫下受到的不利影响更大,未处理前转基因植株株高和鲜质量变化不明显,将PeERF1基因抑制后会导致植株的株高和鲜质量均显著下降,其中株高下降8.3%和14.7%,鲜质量下降20.3%和24.1%,差异均达到极显著水平。以上实验结果表明PeERF1基因在耐旱性中起着至关重要的作用。
-
为了研究干旱胁迫对转PeERF1基因植株叶片叶绿素含量的影响,对过表达和抑制表达转基因株系进行叶绿素含量测定(图4A)。结果显示,在正常状态下,WT和转基因株系叶绿素含量差异不显著;在干旱胁迫处理后,过表达转基因株系叶绿素含量高于WT叶绿素含量,差异极显著;抑制表达转基因株系叶绿素含量低于WT,且差异极显著。

图 4 干旱胁迫对转PeERF1基因84K杨叶片叶绿素含量和过氧化氢酶(CAT)含量的影响
Figure 4. Effect of drought stress on chlorophyll content and the content of catalase (CAT) in leaves of transgenic lines
为了研究干旱胁迫对转PeERF1基因84K杨叶片CAT含量的影响,对过表达和抑制表达转基因株系进行过氧化氢(CAT)含量测定(图4B)。结果显示,在正常状态下,野生型和转基因株系差异不显著;在干旱胁迫处理后,过表达转基因株系高于野生型,差异极显著;抑制表达转基因株系低于野生型,且差异极显著。
-
为了研究干旱胁迫对转PeERF1基因叶片丙二醛(MDA)含量的影响,对过表达和抑制表达转基因株系进行MDA含量测定(图5A)。结果显示,在正常状态下,WT和转基因株系MDA含量差异不显著;在干旱胁迫处理后,过表达转基因株系MDA含量低于WT;抑制表达转基因株系MDA含量高于WT,且差异极显著。

图 5 干旱胁迫对转PeERF1基因叶片丙二醛(MDA)含量和过氧化物酶(POD)含量的影响
Figure 5. Effects of drought stress on malondialdehyde (MDA) content and peroxidase (POD) content in transgenic PeERF1 gene lines
为了研究干旱胁迫对转PeERF1基因叶片POD含量的影响,对过表达和抑制表达转基因株系进行过氧化物酶(POD)含量测定(图5B)。结果显示,在正常状态下,WT和转基因株系POD含量差异不显著;在干旱胁迫处理后,过表达转基因株系POD含量高于WT,差异极显著;抑制表达转基因株系POD含量低于WT,且差异极显著。
-
植物的生理和生化变化在某种程度上反映了植物应对逆境的能力[24]。非生物胁迫总是在胁迫暴露的特定阶段导致活性氧(Reactive oxygen species, ROS)水平增加,但ROS对植物细胞具有细胞毒性[25]。因此,提高对ROS的清除能力可能有利于植物对非生物胁迫的耐受。干旱胁迫常常导致植物体内ROS的过度积累并导致氧化损伤[26]。严重的干旱胁迫会损害细胞离子、转运蛋白和膜相关酶的功能,从而导致ROS的产生[27]。在本研究中,由于干旱胁迫,所有株系叶绿素含量都显著下降。可能是由于干旱诱导的叶绿素降解酶—叶绿素酶活性的增加[28]。但是过表达转基因株系表现出比WT更高的叶绿素含量,表明在干旱胁迫下叶片中的叶绿素含量受到保护,这可能是由于其具有较高的抗氧化酶活性,从而阻止叶片中叶绿素的降解。本研究中对POD和CAT含量测定结果也证实了这一情况。氧自由基引起膜脂质过氧化,降低膜的流动性和选择性。以丙二醛含量(MDA)衡量的脂质过氧化被认为是应激引起的氧化损伤的指标[29]。前期对PeERF1基因进行耐盐实验表型和生理指标分析结果发现PeERF1基因可以提高盐胁迫下84K杨清除活性氧的能力和脯氨酸的积累,增强其耐盐能力[18]。本研究对转PeERF1基因杨树进行干旱胁迫能力分析,发现正常条件下转基因和非转基因MDA含量变化不明显,干旱胁迫后PE丙二醛含量低于WT,SE则高于WT,这一结果表明,PeERF1基因具有诱导植物抵御干旱胁迫引起的氧化损伤的能力。
转录因子是一种调节蛋白,可以与下游基因启动子中的特定顺式元件结合,改变其表达水平。ERF可以与一些顺式元件结合,例如致病相关基因启动子中的GCC框和脱水基因启动子中的DRE基序[30]。拟南芥的AtERF1可以在不同的胁迫下与不同类型的顺式元件结合[31]。因此,ERF基因的过表达对改善生物胁迫和非生物胁迫都具有优势。在本研究中,PeERF1基因已经被证明能够与GCC-BOX、DRE、TGG1和TGG2元件特异性结合[18]。PeERF1可能与参与盐分和干旱胁迫反应的下游基因的不同顺式元件结合,因此修饰单个PeERF1基因可以提高植物耐旱性和耐盐性。
本试验的结果表明胡杨PeERF1基因作为抗旱相关的转录因子,参与了杨树对干旱胁迫的调控,相关研究为后续胡杨抗旱机制的研究奠定了基础,抗旱新基因的挖掘可为植物基因工程育种提供具有重大育种价值的基因资源。
-
本研究结果显示PeERF1基因在胡杨叶片中表达量最高,干旱胁迫下,转PeERF1基因植株生长状态及相关生理指标均发生显著变化。PE比WT表现出更好的生长状态,PE的叶绿素含量、CAT和POD含量高于WT,MDA含量低于WT;而SE则表现出相反的性状。表明PeERF1基因可以提高杨树的抗旱能力。
胡杨PeERF1基因提高转基因银腺杨84K耐旱性研究
Improvement of Drought Tolerance of PeERF1 Transgenic Populus alba × Populus glandulosa ‘84K’
-
摘要:
目的 分析PeERF1基因在胡杨干旱胁迫下的作用和PeERF1转基因植株抗旱的生理适应机制,为进一步研究该基因在木本植物中的抗旱调控机制奠定基础。 方法 以胡杨为材料进行20% PEG6000模拟干旱(0、12和24 h)处理,对胡杨PeERF1基因进行时空表达模式分析。以非转基因(WT)、过表达35S::PeERF1转基因植株(PE)、显性抑制35S::PeERF1-SRDX转基因植株(SE)为试验材料,采用不同浓度PEG6000(对照组,20%)处理WT、PE和SE模拟干旱胁迫,并对其进行表达模式、生长性状和生理指标分析。 结果 研究结果表明PeERF1基因在胡杨叶中的表达水平最高,其次是茎和根。在正常状态下,转基因植株和WT生长性状、叶绿素含量、过氧化氢酶(CAT)、丙二醛(MDA)和过氧化物酶(POD)含量变化不大。在20%PEG6000处理后,PE转基因植株比WT表现出更好的生长状态,PE转基因植株的叶绿素含量、CAT和POD含量高于WT,PE转基因植株MDA含量低于WT。而SE转基因植株则表现出相反的性状。 结论 本研究结果初步显示干旱胁迫下,PeERF1基因转基因植株生长状态、叶绿素含量、过氧化氢酶、丙二醛和过氧化物酶等相关生理指标均发现显著变化。PeERF1对转基因杨树响应干旱起到了正向调控的作用。 Abstract:Objective This study aimed to analyze the role of PeERF1 gene under drought stress in Populus euphratica, and evaluate the physiological adaptation mechanism of PeERF1 transgenic ‘ 84k’ (Populus alba × Populus glandulosa ‘84k’) to drought resistance for providing insights into further study of the drought resistance regulation mechanism of this gene in woody plants. Methods The drought (0, 12 and 24 h) treatment simulated with 20% PEG6000 was carried out on P. euphratica to analyze the temporal and spatial expression pattern of the PeERF1. Based on non-transgenic (WT), overexpressing 35S::PeERF1 transgenic plants (PE), and dominantly suppressed 35S::PeERF1-SRDX transgenic plants (SE), WT, PE and SE were treated with different concentrations of PEG-6000 (Control and 20%) to simulate drought stress, and analyzed for growth traits and physiological indicators. Results The results showed that the expression level of PeERF1 gene was the highest in leaves of P. euphratica, followed by stems and roots. Under normal conditions, the transgenic plants and the WT showed little change in growth traits, chlorophyll content, catalase (CAT), malondialdehyde (MDA) and peroxide dismutase (POD) content. Under 20% PEG6000 treatment, the PE transgenic plants showed better growth status, and higher chlorophyll content, CAT and POD content than WT, and the PE transgenic plants showed lower MDA content than WT, while the SE transgenic plants showed opposite traits. Conclusion Under drought dress, significant changes are found in the growth status, chlorophyll content, catalase, malondialdehyde and peroxidase related physiological indicators of transgenic plants. PeERF1 plays a positive regulatory role on transgenic P. alba × P. glandulosa ‘84k’ in response to drought. -
图 2 干旱胁迫处理转PeERF1基因不同组织部位表达模式分析
Figure 2. Analysis on the expression patterns of different tissue sites of PeERF1 transgenic lines under drought stress
图 3 干旱胁迫对转PeERF1基因84K杨生长状态的影响
Figure 3. The effect of drought stress on the growth state of transgenic Populus alba × Populus glandulosa with PeERF1 gene
图 4 干旱胁迫对转PeERF1基因84K杨叶片叶绿素含量和过氧化氢酶(CAT)含量的影响
Figure 4. Effect of drought stress on chlorophyll content and the content of catalase (CAT) in leaves of transgenic lines
-
[1] WILLIAMS A P, ALLEN C D, MACALADY A K, et al. Temperature as a potent driver of regional forest drought stress and tree mortality[J]. Nature Climate Change, 2012, 3(3): 292-297. [2] SHINOZAKI K, YAMAGUCHI-SHINOZAKI K, et al. Gene networks involved in drought stress response and tolerance[J]. Journal of Experimental Botany, 2006, 58(2): 221-227. doi: 10.1093/jxb/erl164 [3] FENG K, HOU X L, XING G M, et al. Advances in AP2/ERF super-family transcription factors in plant[J]. Critical Reviews in Biotechnology, 2020, 40(6): 750-776. doi: 10.1080/07388551.2020.1768509 [4] DIETZ K J, VOGEL M O, VIEHHAUSER A. AP2/EREBP transcription factors are part of gene regulatory networks and integrate metabolic, hormonal and environmental signals in stress acclimation and retrograde signalling[J]. Protoplasma, 2010, 245(1): 3-14. [5] ZHOU M Q, SHEN C, WU L H, et al. CBF-dependent signaling pathway: a key responder to low temperature stress in plants[J]. Critical Reviews in Biotechnology, 2011, 31(2): 186-192. doi: 10.3109/07388551.2010.505910 [6] YU Y, LIU A, DUAN X, et al. GsERF6, an ethylene-responsive factor from Glycine soja, mediates the regulation of plant bicarbonate tolerance in Arabidopsis[J]. Planta: An International Journal of Plant Biology, 2016, 244(3): 681-698. [7] XIE Z, NOLAN T, JIANG H, et al. The AP2/ERF transcription factor TINY modulates brassinosteroid-regulated plant growth and drought responses in Arabidopsis[J]. The Plant Cell, 2019, 31(8): 1788-1806. doi: 10.1105/tpc.18.00918 [8] FENG W, LI J, LONG S, et al. A DREB1 gene from zoysiagrass enhances Arabidopsis tolerance to temperature stresses without growth inhibition[J]. Plant Science, 2018, 278: 20-31. [9] ZHU J K. Salt and drought stress signal transduction in plants[J]. Annual Review of Plant Biology, 2002, 53(1): 247-273. doi: 10.1146/annurev.arplant.53.091401.143329 [10] HWANG J E, CHAN J L, CHEN H, et al. Overexpression of Arabidopsis dehydration-responsive element-binding protein 2C confers tolerance to oxidative stress[J]. Molecules and Cells, 2012, 33(2): 135-140. doi: 10.1007/s10059-012-2188-2 [11] JIN Y, PAN W, ZHENG X, et al. OsERF101, an ERF family transcription factor, regulates drought stress response in reproductive tissues[J]. Plant Molecular Biology, 2018, 98(1-2): 51-56. doi: 10.1007/s11103-018-0762-5 [12] YAO W, WANG S, ZHOU B, et al. Transgenic poplar overexpressing the endogenous transcription factor ERF76 gene improves salinity tolerance[J]. Tree Physiology, 2016, 36(7): 896-908. doi: 10.1093/treephys/tpw004 [13] CHEN N, TONG S, TANG H, et al. The PalERF109 transcription factor positively regulates salt tolerance via PalHKT1;2 in Populus alba var. pyramidalis[J]. Tree Physiology, 2020, 40(6): 717-730. doi: 10.1093/treephys/tpaa018 [14] CHENG Z, ZHANG X, ZHAO K, et al. Over-expression of ERF38 gene enhances salt and osmotic tolerance in transgenic poplar[J]. Frontiers in Plant Science, 2019, 10: 1375. doi: 10.3389/fpls.2019.01375 [15] 张 肖, 王 旭, 焦培培, 等. 胡杨(Populus euphratica)种子萌发及胚生长对盐旱胁迫的响应[J]. 中国沙漠, 2016, 36(6):1597-1605. doi: 10.7522/j.issn.1000-694X.2015.00173 [16] 马玉花. 植物耐盐分子机理研究进展[J]. 湖北农业科学, 2013, 52(2):255-257,261. doi: 10.3969/j.issn.0439-8114.2013.02.002 [17] JIA H, LIU G, LI J, et al. Genome resequencing reveals demographic history and genetic architecture of seed salinity tolerance in Populus euphratica[J]. Journal of Experimental Botany, 2020, 71(14): 4308-4320. doi: 10.1093/jxb/eraa172 [18] GE X L, ZHANG L, DU J J, et al. Transcriptome analysis of Populus euphratica under salt treatment and PeERF1 gene enhances salt tolerance in transgenic Populus alba × Populus glandulosa[J]. International Journal of Molecular Sciences, 2022, 23(7): 3727. doi: 10.3390/ijms23073727 [19] 张淑霞. 果树叶片中叶绿素、糖和淀粉的联合测定[J]. 河北果树, 1998, 10(2):18-19. doi: 10.19440/j.cnki.1006-9402.1998.02.012 [20] ZHANG Z Y, LIU H H, et al. A C2H2 zinc-finger protein OsZFP213 interacts with OsMAPK3 to enhance salt tolerance in rice[J]. Journal of Plant Physiology, 2018, 229: 100-110. doi: 10.1016/j.jplph.2018.07.003 [21] PENG Q Y, WANG Y M, et al. Inhibiting the CD38/cADPR pathway protected rats against sepsis associated brain injury[J]. Brain Research, 2018, 1678: 56-63. doi: 10.1016/j.brainres.2017.09.029 [22] DOERGE D R, DIVI R L, CHURCHWELL M I. Identification of the colored guaiacol oxidation product produced by peroxidases[J]. Analytical Biochemistry, 1997, 250(1): 10-17. doi: 10.1006/abio.1997.2191 [23] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method[J]. Methods, 2001, 25(4): 402-408. doi: 10.1006/meth.2001.1262 [24] JAMBUNATHAN N. Determination and detection of reactive oxygen species (ROS), lipid peroxidation, and electrolyte leakage in plants[J]. Methods Molecular Biology, 2010, 639: 292-298. [25] HASANUZZAMAN, NAHAR, HOSSAIN, et al. Coordinated actions of glyoxalase and antioxidant defense systems in conferring abiotic stress tolerance in plants[J]. International Journal of Molecular Sciences, 2017, 18(1): 200. doi: 10.3390/ijms18010200 [26] VERSLUES P E, AGARWAL M, KATIYAR-AGARWAL S, et al. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status[J]. Plant Journal, 2006, 45(4): 523-539. doi: 10.1111/j.1365-313X.2005.02593.x [27] AAYUDH D, MOUSTAFA E, BIMAL P, et al. Leaf proteome analysis reveals prospective drought and heat stress response mechanisms in Soybean[J]. Biomed Research International, 2016, 2016(8): 6021047. [28] NOREEN Z, ASHRAF M. Changes in antioxidant enzymes and some key metabolites in some genetically diverse cultivars of radish (Raphanus sativus L. )[J]. Environmental & Experimental Botany, 2009, 67(2): 395-402. [29] HAN D, HAN J, XU T, et al. Overexpression of MbERF12, an ERF gene from Malus baccata (L. ) Borkh, increases cold and salt tolerance in Arabidopsis thaliana associated with ROS scavenging through ethylene signal transduction[J]. In Vitro Cellular & Developmental Biology-Plant, 2021, 57(5): 760-770. [30] ZHU X, LIN Q, XIN L, et al. The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses[J]. Plant Physiology, 2014, 164(3): 1499-1514. doi: 10.1104/pp.113.229575 [31] CHENG M C, LIAO P M, KUO W W, et al. The Arabidopsis ETHYLENE RESPONSE FACTOR1 regulates abiotic stress-responsive gene expression by binding to different cis-acting elements in response to different stress signals[J]. Plant Physiology, 2013, 162(3): 1566-1882. doi: 10.1104/pp.113.221911 -

 点击查看大图
点击查看大图

图(5)
计量
- 文章访问数: 2735
- HTML全文浏览量: 1173
- PDF下载量: 103
- 被引次数: 0
