


-
昆虫通过触角、口器等器官上的嗅觉感受器精准感知外界的化学信号[1]。这些被感知信号的化学本质是由昆虫同类、寄主植物、天敌生物、有机环境等生物和非生物释放的挥发性化学分子,因此嗅觉在昆虫同类交流[2]、寻找寄主[3-4]、繁殖后代[5]、栖息越冬[6]等多种行为中发挥了非常重要的作用。昆虫嗅觉感受器是一类形状多变的特化表皮结构,常见的有毛形感受器、指形感受器、腔锥形感受器等。这些感受器表面有供分子进入的细微孔道,基部有1个或多个感觉神经元。感觉神经元表面和周围分布的多种蛋白作为嗅觉感受的分子基础,在昆虫雌雄识别、寄主定位等活动中发挥了关键作用。
感觉神经元膜蛋白(Sensory neuron membrane proteins, SNMPs)是一类在昆虫嗅觉器官中大量表达的膜蛋白,在分子进化上与脊椎动物跨膜转运蛋白白细胞分化抗原36(Transmembrane transporter protein cluster of differentiation 36, CD36)家族受体同源。脊椎动物CD36家族受体包含2个分别位于氮端和碳端的跨膜区域,以及1个位于跨膜区域之间的细胞外结构域,在细胞脂肪酸转运和代谢中发挥了重要作用[7]。SNMPs分为SNMP1和SNMP2两个亚族,与脊椎动物CD36家族受体类似,也包含2个跨膜区域和1个细胞外结构域。在组织分布上,SNMPs主要在触角、口器等嗅觉器官中表达,在非嗅觉器官中表达量较低[8]。原位杂交实验表明,编码SNMP1的mRNA出现在昆虫嗅觉神经元中[9];进一步的免疫标记实验发现,SNMP1蛋白定位于嗅觉神经元细胞膜,并且与感受雌虫释放性信息素的气味受体高度关联[10]。基因敲除实验证实,SNMP1基因是昆虫性信息素感受所必需的[11]。Pregitzer等[12]在昆虫细胞系中共表达了烟芽夜蛾SNMP1和性信息素受体HR13,发现共表达SNMP1和HR13的细胞在性信息素刺激下产生的嗅觉电位反应显著高于单独表达HR13的细胞,推测SNMP1能够显著提高性信息素受体对性信息素的敏感性。SNMP2定位于嗅觉感受器支持细胞,在嗅觉神经元中不表达[9]。异源表达实验发现,SNMP2的存在与否并不影响气味受体对气味分子的识别。但是鉴于SNMP2与嗅觉功能高度关联的表达模式和细胞定位,推测该蛋白参与了气味分子被识别后的转移和清除过程[13]。
B族清道夫受体(Class b Scavenger receptors, SRb)是一类在昆虫和脊椎动物中都广泛存在的两次跨膜蛋白,与SNMPs一样也具有CD36家族受体的空间结构特征,包含2个位于细胞内的末端、2个跨膜区域和1个位于跨膜区域之间的细胞外结构域。在脊椎动物中,SRb主要作为脂类和脂蛋白受体,参与了脂类、胆固醇、类胡萝卜素等有机化合物的识别和转运[14]。对小鼠鼻腔黏膜SRbs的研究发现,这些SRbs还参与了醛类气味分子的嗅觉识别[15-16]。目前研究比较深入的昆虫SRbs主要来自果蝇、家蚕和蜜蜂;这些SRbs在头部、丝腺、体壁、脂肪体、精巢、卵巢、马氏管等多种组织和器官中分布,表达时期涵盖卵、幼虫、蛹和成虫,参与了包括物质转运、细胞免疫、生长发育在内的多种生化进程[17-20]。
杜仲梦尼夜蛾(Orthosia songi Chen et Zhang)隶属于鳞翅目(Lepidoptera)夜蛾科(Noctuidae)行军虫亚科(Hadeninae)[21]。该虫为单食性害虫,只为害我国重要的药用植物杜仲(Eucommia ulmoides Oliver)[22]。交配后的杜仲梦尼夜蛾雌蛾通常选择杜仲嫩叶进行产卵,孵化的幼虫直接以杜仲叶片为食。该虫食量大,扩散蔓延快,为害时期长,已经成为我国杜仲产业最具威胁的虫害之一[23-25]。对其他夜蛾科昆虫的研究发现,昆虫行为与嗅觉功能关系密切,干扰昆虫对异性和寄主的嗅觉识别已发展成为重要的害虫无公害防治手段[26]。SNMPs、气味受体等嗅觉蛋白在昆虫配偶识别、寄主选择等行为中发挥了重要作用,探究昆虫嗅觉蛋白的功能和表达规律可以为开发针对昆虫嗅觉的害虫防治手段提供科学依据[27]。本研究在杜仲梦尼夜蛾中鉴定得到3个CD36家族受体同源基因(OsonSNMP1、OsonSNMP2和OsonSRb1);这些基因编码的蛋白具有2个位于碳/氮端的跨膜区域和介于跨膜区域之间的胞外结构域;胞外结构域具有6~8个保守的半胱氨酸残基,并包含一个主要由反向平行β-折叠构成的桶状核心;定量PCR结果表明OsonSNMPs和OsonSRb1主要在嗅觉功能相关的器官中表达。本研究为研究杜仲梦尼夜蛾CD36家族同源基因的嗅觉功能提供了科学数据。
-
杜仲梦尼夜蛾幼虫采自湖南省张家界市江娅林场,带回实验室后置于恒温培养箱(温度27 ± 1 ℃,相对湿度70% ± 5%,光周期16L∶8D),以新鲜的杜仲叶片饲养至成虫。幼虫化蛹后,在体视镜下观察腹部生殖节判断雌雄。羽化1~3 d的成虫用于解剖获取触角、口器、胸、腹、足和翅,其中触角、口器、足和翅分别来自20头雄虫和20头雌虫,胸和腹分别来自3头雄虫和3头雌虫;所有组织在液氮中保存直至RNA提取。
-
将杜仲梦尼夜蛾组织研磨成粉末,使用柱式动物组织总RNA抽提纯化试剂盒(生工,上海)进行RNA提取,操作步骤按说明书执行;RNA浓度和纯度在核酸蛋白测定仪(Eppendorf,德国)测定。使用自带基因组DNA移除的cDNA合成试剂盒(全式金,北京)进行cDNA第一链合成,按照试剂盒说明书进行加样,样品在PCR仪(博日科技,杭州)中42 ℃反应15 min,85 ℃加热5 s终止反应;合成的cDNA在−20 ℃保存。
-
从我们前期获得的杜仲梦尼夜蛾转录组数据中筛选注解为“Sensory neuron membrane protein”和“Class b Scavenger receptor”的基因。使用Primer Premier软件将基因翻译成蛋白质序列。使用NCBI网站的BlastP服务(
http://blast.ncbi.nlm.nih.gov/ )进行相似蛋白序列搜索。采用ProtParam tool在线工具(https://web.expasy.org/protparam )对蛋白质分子量和理论等电点进行预测;蛋白跨膜区域采用DTU Health Tech网站的跨膜区域预测服务(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0 )进行预测;疏水性预测采用expasy在线工具(https://web.expasy.org/protscale/ )。蛋白质胞外结构同源建模使用Swiss-model线上服务(https://swissmodel.expasy.org/ ),预测的蛋白空间结构使用Visual Molecular Dynamics软件呈现和展示重要氨基酸残基。预测的SNMP结构与系统发育分析使用MEGA5.0软件,采用步长法构建邻接树,重复1 000次[28]。 -
使用Primer Premier软件设计特异性引物进行基因克隆(表1);PCR反应体系包含12.5 μL 2xTaq MasterMix(康为世纪,北京),上下游引物各1 μL,cDNA模板1 μL,灭菌水9.5 μL。反应程序为95 ℃预变性5 min;然后35个循环的95 ℃变性10 s,60 ℃退火30 s,72 ℃延伸90 s;最后72 ℃延伸5 min。PCR产物经1%的琼脂糖凝胶电泳检测后送生物公司测序(生工,上海)。
表 1 本研究中使用的引物
Table 1. Primers used in this study
引物名称
Primer引物序列
Sequence备注
NoteOsonSNMP1c-F ACAACGGACACCTTCAAAGAAAGCGA 基因克隆 OsonSNMP1c-R CCATACGATCGCCAAGCTCAGGAGT 基因克隆 OsonSNMP2c-F TAAAAAGTGTAAAAGTAAAGTAAAAGTGTTCA 基因克隆 OsonSNMP2c-R TTCCATCAATTGCCTTTATTAACCT 基因克隆 OsonSRb1c-F ATCAAAATGCAAGGTCGTGATAAGTGGTG 基因克隆 OsonSRb1c-R TTGACGCCTATCCTTACAATTTATCTTCTACT 基因克隆 OsonSNMP1q-F TGGCAACTGCCCTCCTAAAG 定量分析 OsonSNMP1q-R GGTCGCTGTCCAGGAAATGA 定量分析 OsonSNMP2q-F ACGGATGCTTGTTGATGGGT 定量分析 OsonSNMP2q-R CTCTGTCAGGCTTGATGCCA 定量分析 OsonSRb1q-F GCCAATACGACTCCCCGATT 定量分析 OsonSRb1q-R ATCCCCTCGACTTGCTCTCT 定量分析 OsonTUAq-F AACCTGAACCGTCTCATCGG 定量分析 OsonTUAq-R CAAGGGGAAGTGGATACGGG 定量分析 OsonNAPDHq-F CTATTTTCGATGCCGCTGCC 定量分析 OsonNAPDHq-R TAACGCGGTTGGAGTATCCG 定量分析 根据测序结果在NCBI网站中的Primer Blast服务(
https://www.ncbi.nlm.nih.gov/tools/primer-blast )中设计实时荧光定量PCR引物(表1)。实时荧光定量PCR在PikoReal荧光定量PCR仪(Thermal,美国)中进行,反应体系为10 mL,包含5 μL 2xTaq UltraSYBR Mixture(康为世纪,北京),上下游引物各0.5 μL,cDNA模板1 μL,灭菌水3 μL。反应程序为95 ℃预变性10 min;然后40个循环的95 ℃变性5 s,60 ℃退火30 s。以OsonTUA和OsonNAPDH基因作为内参基因,雌虫腹部为参考组织,采用2−△△CT法计算OsonSNMPs和OsonSRb1基因在不同组织中的相对表达水平;使用SPSS 17.0软件中的One-way analysis of variance (One-way ANOVA)方法对OsonSNMPs和OsonSRb1在杜仲梦尼夜蛾不同组织中的表达水平进行差异显著性分析。 -
我们从杜仲梦尼夜蛾中克隆得到3个隶属于CD36家族的基因(表2),包括2个SNMPs基因(OsonSNMP1和OsonSNMP2)和1个SRb亚族基因(OsonSRb1)。OsonSNMP1开放阅读框长度为1 575 bp,推导蛋白包含524个氨基酸,预测的蛋白分子量为59.08 kDa,理论等电点6.31。OsonSNMP2开放阅读框1 563 bp,预测其推导蛋白包含520个氨基酸,分子量58.58 kDa,等电点6.62。OsonSRb1开放阅读框长度为1 548 bp,编码的蛋白包含515个氨基酸,分子量58.0 kDa,等电点5.93。
表 2 OsonSNMPs和OsonSRb1序列特征
Table 2. Characteristics of OsonSNMPs and OsonSRb1 sequence
基因名称
Gene name开放阅读框
ORF /bp分子量
Mw /kDa等电点
pI长度
Length /aa推导蛋白
Protein氮端/碳端胞内序列
N-/C-termius氮端/碳端跨膜区域
TM胞外区域
ECOsonSNMP1 1 575 59.08 6.31 524 8/43 23/23 427 OsonSNMP2 1 563 58.58 6.62 520 6/26 23/23 442 OsonSRb1 1 548 58.0 5.93 515 18/38 23/23 413 -
序列一致性分析发现,OsonSNMP1与甘蓝夜蛾(Mamestra brassicae)的MbraSNMP1序列一致性较高,达到93.90%;OsonSNMP2与甜菜夜蛾(Spodoptera exigua)的SexiSNMP2序列一致性较高,达到89.62%。OsonSRb1s与其他夜蛾科昆虫的SRb家族基因序列一致性较高(72.18%~94.22%)。OsonSNMP1、OsonSNMP2和OsonSRb1三者之间序列一致性较低;OsonSNMP1与OsonSNMP2的序列一致性为31.57%,与OsonSRb1的为26.44%;OsonSNMP2与OsonSRb1的序列一致性为27.33%。在昆虫SNMPs和SRb1基因构成的系统发育树中SNMP1、SNMP2和SRb1各自聚为一支(图1),其中SNMP1分支和SNMP2分支临近。OsonSNMP1和OsonSNMP2分别位于SNMP1分支和SNMP2分支,其中OsonSNMP1与MsepSNMP1临近,步长为92%。OsonSNMP2与5个夜蛾科昆虫的SNMP2基因所在分支临近,步长为51%。OsonSRb1位于鳞翅目昆虫的SRb1基因分支,且位于分支的边缘。

图 1 昆虫CD36家族受体同源蛋白的系统发育分析
Figure 1. Phylogenetic analysis of CD36 homologs in insects
-
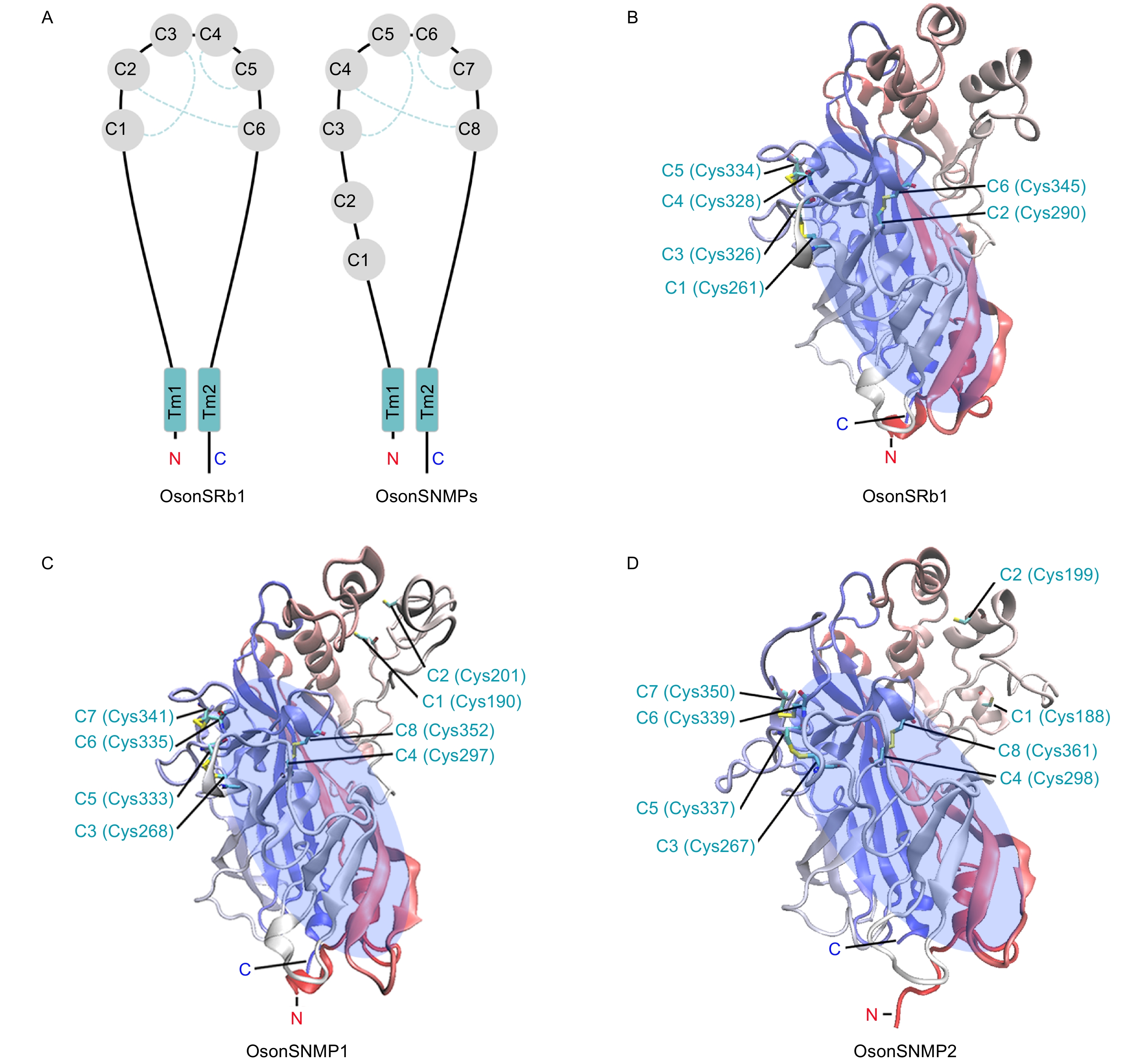
OsonSNMP1、OsonSNMP2和OsonSRb1推导蛋白为包含2个跨膜区域的膜蛋白;氮末端和碳末端位于细胞内,2个疏水的跨膜区域分别靠近氮端和碳端,跨膜区域之间是一个大的细胞外结构(图2A)。OsonSNMP1、OsonSNMP2和OsonSRb1氮末端细胞内结构分别包含8、6和18个氨基酸残基;碳末端细胞内结构分别为43、26和38个氨基酸残基;3个蛋白氮端、碳端跨膜区域均包含23个氨基酸残基;胞外结构分别为427、442和413个氨基酸残基(表2)。

图 2 OsonSNMPs和OsonSRb1蛋白结构和保守半胱氨酸残基在胞外结构域的空间分布
Figure 2. Protein structure of OsonSRb1 and OsonSNMPs and distribution of conserved cysteine residues in extracellular domain
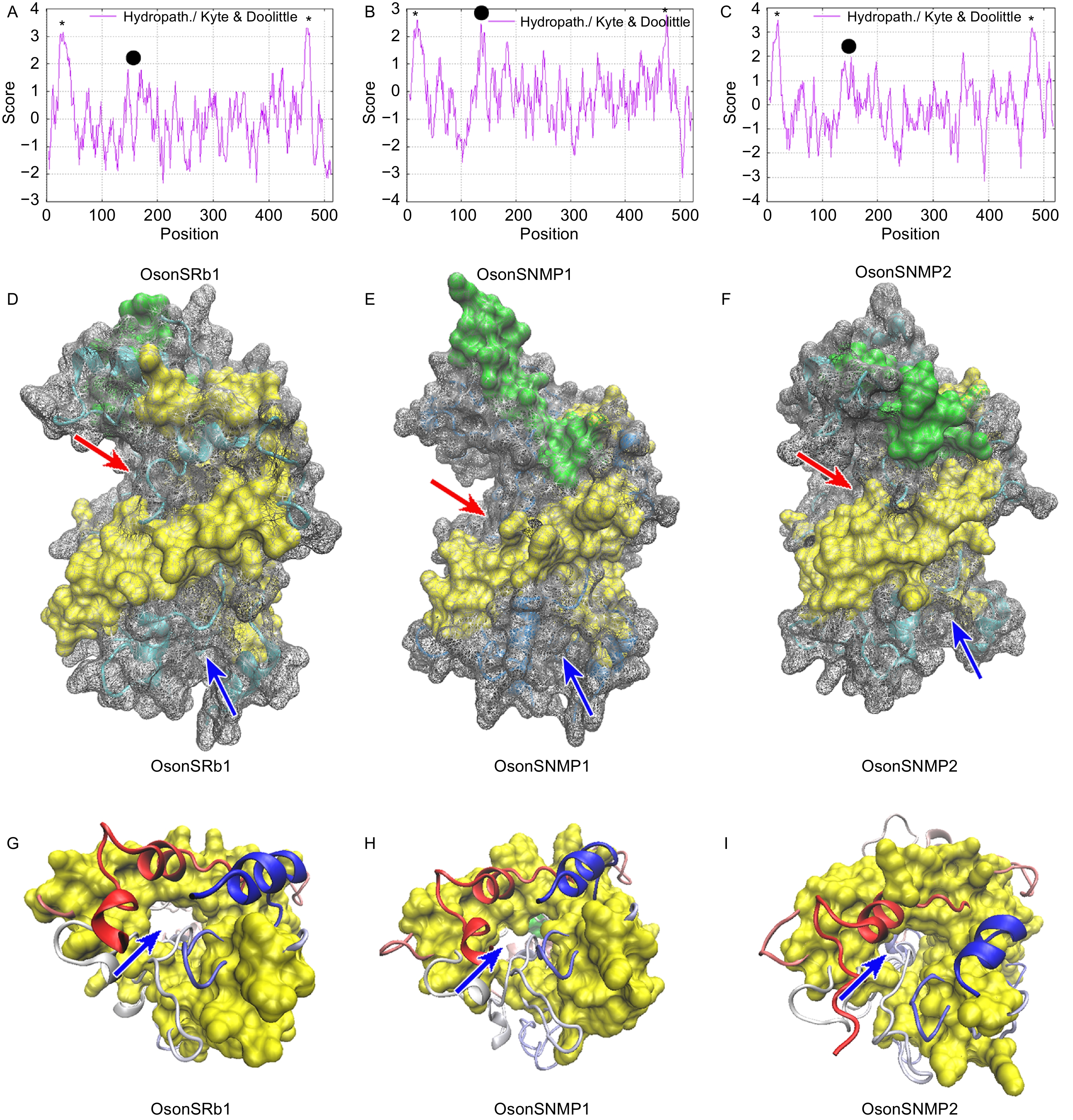
胞外结构域空间结构预测表明,OsonSRb1和OsonSNMP2具有1个由反向平行β-折叠构成的桶状核心和多个位于该核心外围的α-螺旋。OsonSRb1的细胞外结构域上有6个保守的半胱氨酸残基(C1~C6)(图2A),空间结构模型显示这些半胱氨酸残基参与了3个二硫键键的形成(C1~C3、C2-C6和C4~C5)(图2B)。OsonSNMP1和OsonSNMP2的胞外结构域上存在8个保守的半胱氨酸残基(C1~C8)(图2A),其中C3~C8与OsonSRb1的C1~C6同源,可能参与了3个二硫键键的形成(C3~C5、C4~C8和C6~C7),另外OsonSNMP1和OsonSNMP2的C1和C2在空间上临近,但当前模型中并未形成二硫键(图2C, D)。疏水性分析发现,OsonSNMP1、OsonSNMP2和OsonSRb1包含多个疏水性区域(图3A~C),其中碳/氮端跨膜区域疏水值最高;在胞外结构域也存在1个疏水性区域,该区域为一段富含疏水性氨基酸残基的α-螺旋。OsonSRb1、OsonSNMP1和OsonSNMP2胞外结构富含疏水性氨基酸残基的α-螺旋位于蛋白的端部(图3D~F);疏水性氨基酸残基占比分别为55.5%、58.6%和58.6%。OsonSNMP1、OsonSNMP2和OsonSRb1胞外结构的β-折叠核心区域形成了1个中空的桶状结构,该结构与其他二级结构一起形成了一个贯穿整个蛋白的通道,通道入口靠近蛋白顶端,出口朝向细胞膜(图3D~I)。

图 3 OsonSNMPs和OsonSRb1疏水性分析和胞外结构域空间结构
Figure 3. Hydrophobicity analysis and extracellular domain stereo view of OsonSRb1 and OsonSNMPs
-
通过荧光定量PCR实验结果表明,杜仲梦尼夜蛾OsonSNMPs和OsonSR1均在触角中大量表达(图4)。OsonSNMP1在雌雄虫的触角、翅和雌虫足中表达水平较高,且在雄虫触角和雌虫翅中的表达水平显著高于其他组织(图4A)。OsonSNMP2在雌虫触角和足中表达水平较高;在雌虫足中的表达水平低于雌虫触角,但显著高于其他组织(图4B)。OsonSR1在雄虫触角和口器,以及雌虫口器中的表达水平显著高于其他组织(图4C)。

图 4 OsonSNMPs和OsonSRb1在不同组织中的表达
Figure 4. Expression of OsonSNMPs and OsonSRb1 in different tissues
-
CD36家族受体是一类在脂肪细胞、肝细胞、血细胞、嗅觉上皮细胞等多种细胞中广泛表达的受体蛋白[29],在脊椎动物免疫、代谢、血管形成和行为中发挥了重要作用[30]。昆虫SNMPs和SRbs与脊椎动物CD36家族基因同源,在化学感受和脂类物质转运过程中发挥了重要作用[8, 14]。本研究在杜仲单食性害虫杜仲梦尼夜蛾中克隆得到2个SNMPs基因和1个SRbs基因。这些基因编码的蛋白具有CD36家族受体的典型特征,包含2个分别位于氮端和碳端的跨膜区域和1个包含保守半胱氨酸残基的细胞外结构。蛋白结构预测发现,昆虫OsonSRb1和OsonSNMPs均包含一个由β-折叠构成的桶状结构,6个保守半胱氨酸残基参与形成了3个二硫键。序列一致性分析发现,这3个基因编码的蛋白与各自亚族蛋白序列一致性较高,但相互之间序列一致性较低:OsonSNMP1与OsonSNMP2为31.57%,与OsonSRb1为26.44%,OsonSNMP2与OsonSRb1为27.33%。OsonSNMP1和OsonSNMP2在雌雄蛾的触角中大量表达;OsonSRb1在雌雄蛾的触角和口器大量表达;此外,这些基因在翅上也有一定表达。我们的研究结果表明OsonSNMP1、OsonSNMP2和OsonSRbs属于CD36家族,这些基因可能在杜仲梦尼夜蛾感觉器官中发挥了重要功能。
昆虫SRbs蛋白结构域与脊椎动物CD36家族受体类似,包含2个靠近碳/氮末端的跨膜区域和1个胞外结构域,以及位于细胞内的碳/氮末端。CD36家族受体的胞外结构域在配体结合和转移中起到了重要作用[7]。人类CD36胞外结构域具有1个主要由β-折叠构成的通道和1个位于顶端α-螺旋区域的疏水性区域[31]。该通道包含1~2个靠近蛋白顶端的入口和1个朝向细胞膜的出口,脂溶性化合物到达疏水性区域后沿通道滑向细胞膜[32]。本研究预测了OsonSRb1的蛋白结构,发现该蛋白符合CD36家族受体的结构特征,位于胞外结构核心区域的β-折叠形成了一个中空的桶状结构,与其他二级结构一起形成了一条贯穿蛋白的通道,此外OsonSRb1端部具有1个富含疏水性氨基酸残基的α-螺旋,我们推测OsonSRb1能够捕获细胞外的脂溶性化合物,并通过蛋白内的通道将这些化合物向下游传递。在脊椎动物中,血脂主要以脂蛋白的形式进行运输,CD36家族受体通过胞外结构顶端的丙氨酸残基与脂蛋白结合,并借助核心区域的通道转运疏水性的长链脂肪酸[32]。昆虫CD36家族受体也能够结合长链脂肪酸等脂溶性物质,这些膜蛋白在昆虫脂肪酸代谢、免疫、感觉等生理活动中发挥了重要作用。在家蚕中,BmorSRbs家族受体与脂蛋白相互作用,促进固醇类化合物的摄入和转运[33];BmorSCRB8可以直接与病原关联因子结合,提高机体对病原的清除效力[18]。果蝇SRbs受体ninaD参与了细胞对类胡萝卜素的摄入,由于类胡萝卜素是视觉发色团合成的重要原料,缺失ninaD基因会导致果蝇失明[20, 34]。Yang和Zhang[35]对果蝇嗅觉系统的蛋白互作分析发现,果蝇SRbs与嗅觉功能高度关联,这些蛋白可能在嗅觉系统中参与了脂类转运和废弃代谢物清理。
昆虫SNMPs结构符合CD36家族受体的结构特征,也具有主要由β-折叠构成的通道和位于胞外结构顶端的疏水性区域。Gomez-Diaz等[36]以哺乳动物CD36家族受体为模板分析果蝇SNMP1空间结构,发现果蝇SNMP1的胞外结构也具有1个足以容纳性信息素的通道,氨基酸定点突变阻塞这个通道后会降低果蝇对性信息素的嗅觉电位反应,而异源表达小鼠CD36家族受体能够恢复果蝇SNMP1突变体对性信息素的嗅觉电位反应,表明昆虫SNMPs可能具有与脊椎动物CD36家族受体类似的功能。本研究预测了OsonSNMPs胞外区域的蛋白结构,位于蛋白核心区域的β-折叠与其他二级结构一起形成了一条贯穿蛋白的通道,此外在OsonSNMP1和OsonSNMP2胞外结构端部均有1个富含疏水性氨基酸残基的α-螺旋,我们推测OsonSNMPs与脊椎动物CD36家族受体和其他昆虫SNMPs类似,能够捕获细胞外的疏水性气味化合物分子,并通过蛋白内的通道将这些化合物向气味受体传递。CD36家族受体结合的配体包括各种氧化脂蛋白、乙酰化脂蛋白和各种天然脂蛋白,在CD36存在的条件下,长链脂肪酸从血清蛋白或磷酸双分子层上分离可以发生在毫秒之内[37]。在果蝇中,SNMP1能极大提高感觉神经元对性信息素的敏感性,这一现象可能是由于SNMP1能够从气味结合蛋白上卸载了气味分子,并通过胞外结构的通道将气味分子输送到气味受体进行识别[11, 36, 38]。
在脊椎动物嗅觉上皮细胞中分布的CD36家族受体参与了油酸、醛类的嗅觉感受[39-41]。昆虫CD36家族受体也在嗅觉功能中发挥了重要作用;这些嗅觉蛋白的组织分布主要包括触角、下颚须、下唇须等感觉器官[42-44]。沙漠蝗SgreSNMP1和SgreSNMP2在触角中大量表达,组织定位发现SgreSNMP1在感觉神经元树突和支持细胞绒毛上分布,SgreSNMP2只在支持细胞绒毛分布,以上组织分布情况表明示SgreSNMP1可能参与了气味分子的接收和嗅觉淋巴液内气味分子清除,而SgreSNMP2的功能仅限于嗅觉淋巴液内气味分子清除[45-46]。在果蝇中,SNMP1被证实定位于触角感受器基部的感觉神经元,并参与了气味受体对性信息素的识别过程[11, 41]。本研究发现,OsonSNMP1和OsonSNMP2在杜仲梦尼夜蛾触角中大量表达,OsonSRb1在雌雄蛾的触角和口器大量表达;与嗅觉器官高度关联的表达模式和与脊椎动物类似的蛋白结构暗示这些基因在杜仲梦尼夜蛾嗅觉系统中发挥了功能。此外,OsonSNMP1在运动相关的组织足和翅中也大量表达。但是这些蛋白的具体嗅觉功能还有待进一步细胞定位和蛋白功能实验来验证。
-
综上,OsonSNMP1、OsonSNMP2和OsonSRb1具有脊椎动物CD36家族受体类似的空间结构,包含2个跨膜区域和1个细胞外结构域;胞外结构域具有1个主要由反向平行β-折叠构成的桶状核心和1个富含疏水性氨基酸残基的α-螺旋。OsonSNMP1和OsonSNMP2在杜仲梦尼夜蛾触角中大量表达,OsonSRb1在雌雄蛾的触角和口器大量表达。与嗅觉器官高度关联的表达模式和与脊椎动物类似的蛋白结构表明这些基因可能在杜仲梦尼夜蛾嗅觉系统中发挥了功能。
杜仲梦尼夜蛾CD36家族同源基因克隆与组织分布研究
Cloning and Tissue Distribution of Homologous Genes of CD36 Family in Orthosia songi Chen et Zhang (Lepidoptera: Noctuidae)
-
摘要:
目的 揭示杜仲梦尼夜蛾CD36家族同源基因神经元膜蛋白(SNMPs)和b族清道夫受体(SRb)的序列特征和组织分布情况。 方法 设计特异性引物克隆杜仲梦尼夜蛾SNMPs和SRb基因,对这些基因编码的蛋白进行同源建模;通过荧光定量PCR分析这些基因在不同组织的表达情况。 结果 在杜仲梦尼夜蛾中鉴定得到3个CD36家族同源基因(OsonSNMP1、OsonSNMP2和OsonSRb1);这些基因编码的蛋白具有2个跨膜区域和1个细胞外结构域。空间结构预测发现,这些蛋白的胞外结构域包含1个主要由反向平行β-折叠构成的桶状核心和1个富含疏水性氨基酸残基的α-螺旋;荧光定量PCR结果表明,OsonSNMP1、OsonSNMP2和OsonSRb1均在触角中大量表达;OsonSNMP1在雄虫触角中的表达水平显著高于雌虫触角,OsonSNMP2在雌虫触角的表达水平显著高于其他组织,OsonSRb1在雄虫触角中大量表达外,还在雌雄虫的口器中大量表达。 结论 本研究发现OsonSNMP1、OsonSNMP2和OsonSRb1编码的蛋白具有与脊椎动物CD36家族受体类似的空间结构,与嗅觉器官高度关联的表达模式表明这些基因可能在杜仲梦尼夜蛾嗅觉系统中发挥了功能,以上结果为探究杜仲梦尼夜蛾CD36家族同源基因的嗅觉功能提供了数据。 Abstract:Objective To reveal the sequence characteristics and expression profiles of CD36 homolog genes, sensory neuron membrane proteins (SNMPs) and class b scavenger receptors (SRb) in Orthosia songi Chen et Zhang. Method Specific primers were used to clone the SNMP and SRb genes. The protein encoded by these genes were constructed by homo-modeling, and the gene expression profiles were determined by quantitative fluorescent real-time PCR. Result Three CD36 homolog genes (OsonSNMP1, OsonSNMP2 and OsonSRb1) were identified from O. songi. The proteins encoded by these genes consisted of two transmembrane domains and an extracellular domain. The extracellular domain contained an antiparallel β-barrel core and a hydrophobic domain near the top. OsonSNMP1 was mainly expressed in the antennae and wings of male and female moth, and the expression level in male antennae was significantly higher than that in other tissues. OsonSNMP2 was discriminatorily expressed in male and female antennae and female wings, and its expression level in female antennae was significantly higher than that in other tissues. OsonSRb1 were specifically expressed in the antennae and mouthparts of male and female. Conclusion The proteins encoded by OsonSNMP1, OsonSNMP2 and OsonSRb1 have similar structures with CD36 receptors in vertebrates. Their expression profiles highly associated with olfactory organs indicate the potential roles in olfactory systems. These results provide experimental data for further study on the olfactory function of CD36 homolog genes in Orthosia songi. -
Key words:
- Orthosia songi
- / olfactory
- / CD36 family
- / Sensory neuron membrane protein
- / Class b scavenger receptor
-
图 2 OsonSNMPs和OsonSRb1蛋白结构和保守半胱氨酸残基在胞外结构域的空间分布
Figure 2. Protein structure of OsonSRb1 and OsonSNMPs and distribution of conserved cysteine residues in extracellular domain
图 3 OsonSNMPs和OsonSRb1疏水性分析和胞外结构域空间结构
Figure 3. Hydrophobicity analysis and extracellular domain stereo view of OsonSRb1 and OsonSNMPs
图 4 OsonSNMPs和OsonSRb1在不同组织中的表达
Figure 4. Expression of OsonSNMPs and OsonSRb1 in different tissues
表 1 本研究中使用的引物
Table 1. Primers used in this study
引物名称
Primer引物序列
Sequence备注
NoteOsonSNMP1c-F ACAACGGACACCTTCAAAGAAAGCGA 基因克隆 OsonSNMP1c-R CCATACGATCGCCAAGCTCAGGAGT 基因克隆 OsonSNMP2c-F TAAAAAGTGTAAAAGTAAAGTAAAAGTGTTCA 基因克隆 OsonSNMP2c-R TTCCATCAATTGCCTTTATTAACCT 基因克隆 OsonSRb1c-F ATCAAAATGCAAGGTCGTGATAAGTGGTG 基因克隆 OsonSRb1c-R TTGACGCCTATCCTTACAATTTATCTTCTACT 基因克隆 OsonSNMP1q-F TGGCAACTGCCCTCCTAAAG 定量分析 OsonSNMP1q-R GGTCGCTGTCCAGGAAATGA 定量分析 OsonSNMP2q-F ACGGATGCTTGTTGATGGGT 定量分析 OsonSNMP2q-R CTCTGTCAGGCTTGATGCCA 定量分析 OsonSRb1q-F GCCAATACGACTCCCCGATT 定量分析 OsonSRb1q-R ATCCCCTCGACTTGCTCTCT 定量分析 OsonTUAq-F AACCTGAACCGTCTCATCGG 定量分析 OsonTUAq-R CAAGGGGAAGTGGATACGGG 定量分析 OsonNAPDHq-F CTATTTTCGATGCCGCTGCC 定量分析 OsonNAPDHq-R TAACGCGGTTGGAGTATCCG 定量分析  下载: 导出CSV
下载: 导出CSV
表 2 OsonSNMPs和OsonSRb1序列特征
Table 2. Characteristics of OsonSNMPs and OsonSRb1 sequence
基因名称
Gene name开放阅读框
ORF /bp分子量
Mw /kDa等电点
pI长度
Length /aa推导蛋白
Protein氮端/碳端胞内序列
N-/C-termius氮端/碳端跨膜区域
TM胞外区域
ECOsonSNMP1 1 575 59.08 6.31 524 8/43 23/23 427 OsonSNMP2 1 563 58.58 6.62 520 6/26 23/23 442 OsonSRb1 1 548 58.0 5.93 515 18/38 23/23 413
下载: 导出CSV
-
[1] HALLEM E A, DAHANUKAR A, CARLSON J R. Insect odor and taste receptors[J]. Annu Rev Entomol., 2006, 51: 113-135. doi: 10.1146/annurev.ento.51.051705.113646 [2] TANAKA K, UDA Y, ONO Y,et al. Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile[J]. Curr Biol., 2009, 19(11): 881-90. doi: 10.1016/j.cub.2009.04.035 [3] KASANG G. Uptake of the sex pheromone 3H-bombykol and related compounds by male and female Bombyx antennae[J]. J Insect Physiol., 1974, 20(12): 2407-2422. doi: 10.1016/0022-1910(74)90027-4 [4] MATSUMOTO S, HULL J J, OHNISHI A,et al. Molecular mechanisms underlying sex pheromone production in the silkmoth, Bombyx mori: characterization of the molecular components involved in bombykol biosynthesis[J]. J Insect Physiol., 2007, 53(8): 752-729. doi: 10.1016/j.jinsphys.2007.02.014 [5] GEGEAR R J. Exploring the role of cognition in the annual fall migration of the monarch butterfly (Danaus plexippus)[J]. Insects, 2021, 12(8): 760. doi: 10.3390/insects12080760 [6] GREENSTEIN L, STEELE C, TAYLOR C M. Host plant specificity of the monarch butterfly Danaus plexippus: A systematic review and meta-analysis[J]. PLoS One, 2022, 17(6): e0269701. doi: 10.1371/journal.pone.0269701 [7] PEPINO M Y, KUDA O, SAMOVSKI D,et al. Structure-function of CD36 and importance of fatty acid signal transduction in fat metabolism[J]. Annu Rev Nutr., 2014, 34(1): 281-303. doi: 10.1146/annurev-nutr-071812-161220 [8] 宋月芹, 白小军, 陈庆霄, 等. 红脊长蝽感觉神经元膜蛋白基因克隆及组织表达谱分析[J]. 林业科学研究, 2021, 34(5):135-141. doi: 10.13275/j.cnki.lykxyj.2021.005.016 [9] ZHANG J, LIU Y, WALKER W B,et al. Identification and localization of two sensory neuron membrane proteins from Spodoptera litura (Lepidoptera: Noctuidae)[J]. Insect Sci., 2015, 22(3): 399-408. doi: 10.1111/1744-7917.12131 [10] SHAN S, WANG S N, SONG X,et al. Molecular characterization and expression of sensory neuron membrane proteins in the parasitoid Microplitis mediator (Hymenoptera: Braconidae)[J]. Insect Sci., 2020, 27(3): 425-439. doi: 10.1111/1744-7917.12667 [11] BENTON R, VANNICE KS, VOSSHALL LB. An essential role for a CD36-related receptor in pheromone detection in Drosophila[J]. Nature, 2007, 450(7167): 289-293. doi: 10.1038/nature06328 [12] PREGITZER P, GRESCHISTA M, BREER H,et al. The sensory neurone membrane protein SNMP1 contributes to the sensitivity of a pheromone detection system[J]. Insect Mol Biol., 2014, 23(6): 733-742. doi: 10.1111/imb.12119 [13] CASSAU S, KRIEGER J. The role of SNMPs in insect olfaction[J]. Cell Tissue Res., 2021, 383(1): 21-33. doi: 10.1007/s00441-020-03336-0 [14] MA B T, JIA J, WANG X B, ZHANG R,et al. Differential roles of Scavenger receptor class B type I: A protective molecule and a facilitator of atherosclerosis[J]. Mol Med Rep., 2020, 22(4): 2599-2604. doi: 10.3892/mmr.2020.11383 [15] LEE S, EGUCHI A, TSUZUKI S,et al. Expression of CD36 by olfactory receptor cells and its abundance on the epithelial surface in mice[J]. PLoS One, 2015, 10(7): e0133412. doi: 10.1371/journal.pone.0133412 [16] TSUZUKI S, KIMOTO Y, LEE S,et al. A novel role for scavenger receptor B1 as a contributor to the capture of specific volatile odorants in the nasal cavity[J]. Biomed Res., 2018, 39(3): 117-129. doi: 10.2220/biomedres.39.117 [17] HERBOSO L, TALAMILLO A, PÉREZ C,et al. Expression of the scavenger receptor class B type I (SR-BI) family in Drosophila melanogaster[J]. Int J Dev Biol., 2011, 55(6): 603-611. doi: 10.1387/ijdb.103254lh [18] ZHANG K, HU X, ZHAO Y,et al. Scavenger receptor B8 improves survivability by mediating innate immunity in silkworm, Bombyx mori[J]. Dev Comp Immunol., 2021, 116: 103917. doi: 10.1016/j.dci.2020.103917 [19] ZHOU LZ, WANG RJ, YAN Y Y,et al. Scavenger receptor B1 mediates phagocytosis and the antimicrobial peptide pathway in the endoparasitic wasp Micropilits mediator[J]. Dev Comp Immunol., 2021, 119: 104039. doi: 10.1016/j.dci.2021.104039 [20] VOOLSTRA O, KIEFER C, HOEHNE M,et al. The Drosophila class B scavenger receptor NinaD-I is a cell surface receptor mediating carotenoid transport for visual chromophore synthesis[J]. Biochemistry, 2006, 45(45): 13429-13437. doi: 10.1021/bi060701u [21] 赵 阳, 朱景乐, 李芳东, 等. 杜仲梦尼夜蛾发育历期及取食行为研究[J]. 植物保护, 2015, 41(2):75-79. doi: 10.3969/j.issn.0529-1542.2015.02.013 [22] 赵 阳, 朱景乐, 李芳东, 等. 杜仲梦妮夜蛾幼虫的空间分布型及抽样技术[J]. 环境昆虫学报, 2014, 36(4):629-634. [23] 周云龙, 张声堂, 刘湘银, 等. 杜仲梦尼夜蛾生物学特性及防治研究[J]. 西北林学院学报, 1996(2):66-70 + 81. [24] 孙志强, 赵 阳, 马志刚, 等. 杜仲梦尼夜蛾危害的风险分级[J]. 林业科学, 2016, 52(7):78-86. [25] 李剑豪, 李东平, 黄祖惠, 等. 杜仲梦尼夜蛾生物学特性初步研究[J]. 森林病虫通讯, 1997(4):19-21. [26] 王留洋, 杨超霞, 郭兵博, 等. 昆虫性信息素研究进展与应用前景[J]. 农药学学报, 2022, 24(5):997-1016. doi: 10.16801/j.issn.1008-7303.2022.0086 [27] CLARK J T, RAY A. Olfactory mechanisms for discovery of odorants to reduce insect-host contact[J]. J Chem Ecol., 2016, 42(9): 919-930. doi: 10.1007/s10886-016-0770-3 [28] TAMURA K, PETERSON D, PETERSON N,et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Mol Biol Evol., 2011, 28(10): 2731-2739. doi: 10.1093/molbev/msr121 [29] YANG R, LIU Q, ZHANG M. The past and present lives of the intraocular transmembrane protein CD36[J]. Cells, 2023, 12(1): 171. doi: 10.3390/CELLS12010171 [30] SILVERSTEIN R L, FEBBRAIO M. CD36, a scavenger receptor involved in immunity, metabolism, angiogenesis, and behavior[J]. Sci Signal., 2009, 2(72): re3. doi: 10.1126/scisignal.272re3 [31] NECULAI D, SCHWAKE M, RAVICHANDRAN M,et al. Structure of LIMP-2 provides functional insights with implications for SR-BI and CD36[J]. Nature, 2013, 504(7478): 172-176. doi: 10.1038/nature12684 [32] HSIEH F L, TURNER L, BOLLA J R,et al. The structural basis for CD36 binding by the malaria parasite[J]. Nat Commun., 2016, 7(1): 12837. doi: 10.1038/ncomms12837 [33] TAKESHIMA M, OGIHARA M H, KATAOKA H. Characterization and functional analysis of BmSR-B1 for phytosterol uptake[J]. Steroids, 2022, 184: 109039. doi: 10.1016/j.steroids.2022.109039 [34] KIEFER C, SUMSER E, WERNET M F,et al. A class B scavenger receptor mediates the cellular uptake of carotenoids in Drosophila[J]. Proc Natl Acad Sci U S A., 2002, 99(16): 10581-10586. doi: 10.1073/pnas.162182899 [35] YANG S, ZHANG W J. Systematic analysis of olfactory protein-protein interactions network of fruitfly, Drosophila melanogaster[J]. Arch Insect Biochem Physiol., 2022, 110(2): e21882. doi: 10.1002/ARCH.21882 [36] GOMEZ-DIAZ C, BARGETON B, ABUIN L,et al. A CD36 ectodomain mediates insect pheromone detection via a putative tunnelling mechanism[J]. Nat Commun., 2016, 7: 11866. doi: 10.1038/ncomms11866 [37] JAY A G, CHEN A N, PAZ M A,et al. CD36 binds oxidized low density lipoprotein (LDL) in a mechanism dependent upon fatty acid binding[J]. J Biol Chem., 2015, 290(8): 4590-4603. doi: 10.1074/jbc.M114.627026 [38] LI Z, NI J D, HUANG J,et al. Requirement for Drosophila SNMP1 for rapid activation and termination of pheromone-induced activity[J]. PLoS Genet., 2014, 10(9): e1004600. doi: 10.1371/journal.pgen.1004600 [39] OBERLAND S, ACKELS T, GAAB S,et al. CD36 is involved in oleic acid detection by the murine olfactory system[J]. Front Cell Neurosci., 2015, 9: 366. doi: 10.3389/fncel.2015.00366 [40] LEE S, TSUZUKI S, AMITSUKA T,et al. CD36 involvement in the olfactory perception of oleic aldehyde, an odour-active volatile compound, in mice[J]. Biomed Res., 2017, 38(3): 207-213. doi: 10.2220/biomedres.38.207 [41] TSUZUKI S, KIMOTO Y, MARUI K,et al. Application of a novel fluorescence intensity assay: identification of distinct fatty acetates as volatile compounds that bind specifically to amino acid region 149-168 of a transmembrane receptor CD36[J]. Biosci Biotechnol Biochem., 2022, 86(4): 509-518. doi: 10.1093/bbb/zbac018 [42] JIN X, HA T S, SMITH D P. SNMP is a signaling component required for pheromone sensitivity in Drosophila[J]. Proc Natl Acad Sci U S A., 2008, 105(31): 10996-11001. doi: 10.1073/pnas.0803309105 [43] LEMKE R S, PREGITZER P, EICHHORN A S,et al. SNMP1 and odorant receptors are co-expressed in olfactory neurons of the labial and maxillary palps from the desert locust Schistocerca gregaria (Orthoptera: Acrididae)[J]. Cell Tissue Res., 2020, 379(2): 275-289. doi: 10.1007/s00441-019-03083-x [44] GUO M, CHEN Q, LIU Y,et al. Chemoreception of mouthparts: sensilla morphology and discovery of chemosensory genes in proboscis and labial palps of adult Helicoverpa armigera (Lepidoptera: Noctuidae)[J]. Front Physiol., 2018, 9: 970. doi: 10.3389/fphys.2018.00970 [45] JIANG X, PREGITZER P, GROSSE-WILDE E,et al. Identification and characterization of two "sensory neuron membrane proteins" (SNMPs) of the desert locust, Schistocerca gregaria (Orthoptera: Acrididae)[J]. J Insect Sci., 2016, 16(1): 33. doi: 10.1093/jisesa/iew015 [46] CASSAU S, SANDER D, KARCHER T,et al. The sensilla-specific expression and subcellular localization of SNMP1 and SNMP2 reveal novel insights into their roles in the antenna of the desert locust Schistocerca gregaria[J]. Insects, 2022, 13(7): 579. doi: 10.3390/insects13070579 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 2201
- HTML全文浏览量: 994
- PDF下载量: 86
- 被引次数: 0
