

-
短枝木麻黄(Casuarina equisetifolia L.)为木麻黄科(Casuarinaceae)木麻黄属(Casuarina)常绿乔木,原产于澳大利亚、太平洋诸岛以及亚洲东南部,喜高温多湿气候,适生于海岸的疏松沙地[1-2]。木麻黄生长迅速、耐干旱,并具有防风固沙、耐盐碱和生物固氮等特殊功能,被广泛用于热带和亚热带沿海防护林的建设,是我国东南和华南沿海地区不可替代的沿海防护林树种和当家树种,同时也是重要的经济林和园林绿化树种[2-6]。近年来,由于抗病品种缺乏、栽培经营措施不当以及自然灾害等因素的影响,木麻黄病虫害的发生也日趋严重,其中,青枯病是发生规模最大、危害最严重的病害[7]。木麻黄青枯病是由劳尔氏菌(Ralstonia solanacearum)引起的一种系统性维管束病害,被称为木麻黄的“癌症”或“绝症”,青枯病也是目前世界范围内传播广泛、危害严重、最难防治的重大细菌性病害之一,可危害54个科的450余种植物[2, 8]。木麻黄青枯病的大面积发生,严重制约着沿海木麻黄防护林的健康发展,对我国东南沿海的生态环境和生态安全构成了严重威胁,目前尚无有效的防控措施[4, 9-10]。
根际微生物是受植物影响最大的土壤微生物类群,蕴含着极其丰富的微生物资源[11]。根际土壤中的有益微生物可通过直接或间接的方式促进植物生长、防治由病原菌引起的多种植物病害[12-13]。因此,根际微生物是重要生物活性产物的来源,探究和开发根际微生物为生物防治病虫害问题以及提高农林业的经济价值提供了广阔前景[14]。放线菌是根际微生物中一类重要的微生物资源,目前基于微生物源活性先导化合物而研发的抗生素、免疫抑制剂、抗真菌、抗肿瘤以及抑制炎症等药物绝大多数来源于放线菌[15-16]。Elshafie等[17]从金合欢(Vachellia farnesiana (L.) Wight & Arn.)和油橄榄(Canarium oleosum (Lam.) Engl.)中分离到2株根际放线菌可以很好的保护番茄(Solanum lycopersicum L.)免受核盘菌(Sclerotinia sclerotiorum)的侵染;方正等[18]从金钗石斛(Dendrobium nobile Lindl.)根际土壤中分离出164株放线菌,其中,部分放线菌对多种供试细菌均表现出抑菌活性;叶景静等[19]从红树林根际土壤中分离出88株放线菌,其中,7株放线菌对至少一种供试细菌具有抗菌活性;吴佳等[20]从缬草(Valeriana officinalis L.)根际土壤中共分离出118株放线菌,其中,33株放线菌对病原菌有较好的抑制效果;Kumari等[21]发现,植物根际放线菌的粗提物对金黄色葡萄球菌(Staphylococcus aureus)、变形杆菌(Proteus vulgaris)以及枯草芽孢杆菌(Bacillus subtilis)表现出明显的抑制作用。放线菌可产生次生代谢产物抑制某些病原菌生长,促进植物生长发育,一些放线菌还可以侵染植物诱导其产生大量的活性次生代谢产物[22-23]。木麻黄是一种特殊类型的共生营养型植物,对木麻黄共生固氮菌和菌根菌的研究一直是木麻黄研究的热点[24]。Gunasekera等[25]从木麻黄固氮根瘤中分离到Frankia sp.、Micromonospora sp.和新的共生放线菌链霉菌属(Streptomyces sp.),其中,链霉菌属放线菌能明显促进木麻黄侧根的生长。而目前对于木麻黄根际土壤放线菌的报道较少,Abhijit等[26]从印度不同地区的木麻黄根际土壤中共分离到14株链霉菌属放线菌,其中部分放线菌对3种供试细菌(Bacillus subtilis、Escherichia coli和Xanthomonas citri)中的1种或2种表现出抑菌活性,所有放线菌对供试植物病原真菌(Fusarium oxysporum)都表现出抑制活性。本研究通过采集短枝木麻黄根际土壤,分离鉴定根际土壤中的放线菌,并测定短枝木麻黄根际放线菌次生代谢产物的抗细菌活性,从中筛选出抗劳尔氏菌的活性菌株,进一步采用HPLC分析活性菌株次生代谢产物的情况,以期为木麻黄青枯病的生物防治以及放线菌资源的综合开发和利用提供理论依据。
-
短枝木麻黄根际土壤于2018年6月采自雷州半岛的广东省湛江市雷州市,选取8个不同地点采集样品,分别为A(20°58′43″N;110°7′52″E)、B(20°48′60″N;110°20′2″E)、C(20°49′3″N;110°20′49″E)、D(20°49′4″N;110°20′49″E)、E(20°47′32″N;110°23′10″E)、F(20°47′32″N;110°23′10″E)、G(20°47′37″N;110°23′8″E)和H(20°47′35″N;110°23′8″E)。采集时选取健康的短枝木麻黄植株,在主干1.5 m范围内从4个不同的方向挖取距离地面15~30 cm的根系,将根系挖出后采用抖根法收集根系掉落的土壤,将4个不同方向采集的土壤合并在一起用自封袋密封后立即带回实验室,4 ℃保存备用。
-
采用热击稀释法[27]分离短枝木麻黄根际土壤中的放线菌。首先将采集的土壤充分混匀,去除杂质后取1 g土壤置于100 mL的旋蒸瓶内,加入10 mL无菌水,在50 ℃无压条件下采用OSB-2100旋转蒸发器(东京理化器械株式会社)旋转15 min,使其充分混匀;而后取1 mL混匀后的土壤悬浊液于15 mL离心管内,并向其加入9 mL无菌水,充分混匀后采用同样的方法依次稀释成浓度为10−1、10−2和10−3的土壤溶液,备用。分别吸取100 µL各浓度的土壤溶液,置于改良的M1固体培养基(未添加人工海盐)[28]平板上,用无菌的涂布棒涂布均匀,每个浓度重复3次;然后将培养基平板放在28 ℃培养箱内暗培养,根据菌落生长情况以及菌落特征挑选肉眼可辨别的放线菌,接种于M1培养基纯化培养。将纯化后的放线菌菌株接种于M1培养基上,28 ℃恒温暗培养3~7 d,观察记录菌落形态和颜色,合并相同的菌株并拍照。将纯化后的菌株接种在2.5 mL冻存管里(含0.5 mL无菌的甘油和0.5 mL M1液体培养基),−20 ℃保存备用。
-
将纯化后的放线菌接种于ISP-2液体培养基[29]中,28 ℃ 150 rpm的条件下震荡培养1 d,吸取适量的放线菌菌丝,DNA提取方法按照Ezup柱式细菌基因DNA抽提试剂盒(生工生物工程(上海)股份有限公司)提取步骤进行;采用原核生物通用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-TACGGCTACCTTGTTACGACTT-3′)进行PCR扩增[2]。PCR反应体系为30 µL:12 µL ddH2O,上下游引物各0.5 µL,15 µL 2 × Taq PCR MasterMix,2 µL模板DNA。PCR扩增条件为:95 ℃预变性5 min;94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸1 min,循环35次;最后72 ℃延伸10 min,4 ℃保温。所得序列均使用DNAMAN软件进行互补拼接,通过在NCBI网站上进行BLAST,将测序结果在GenBank数据库中进行同源性检索,下载与其相似性较高的序列及其近似属的序列,使用MAFTT version 7进行序列处理后,采用邻接法(Neighbor-joining),用MEGA 7.0.26软件构建系统发育树,其中,Bootstrap method中重复抽样次数设置1 000,模式为Maximum Composite Likelihood。将最终的鉴定结果和所得的序列提交到NCBI数据库(
http://www.ncbi.nlm.nih.gov ),并获得其登录号。 -
采用无菌打孔器从培养好的放线菌M1培养基平板上打取4块菌饼,接种到装有250 mL M1液体培养基的三角瓶中,在28 ℃ 150 rpm条件下培养14 d。发酵完成后采用4层纱布将菌丝残渣和发酵液分开,然后往发酵液中加入等体积的乙酸乙酯,室温下连续萃取3次,合并萃取液经减压浓缩后即得放线菌乙酸乙酯层次生代谢产物,4 ℃条件下保存,备用。
-
供试细菌分别为木麻黄青枯病菌(Ralstonia solanacearum,G−)、大肠杆菌(Escherichia coli,G−)、枯草芽孢杆菌(Bacillus subtilis,G+)和溶血葡萄球菌(Staphylococcus haemolyticus,G+)。采用TLC-MTT-生物自显影法[30]测定短枝木麻黄根际放线菌次生代谢产物对不同供试细菌的抑制活性。先将各根际放线菌次生代谢产物溶解,用直径0.5 mm的毛细管在薄层层析板上点样,点样量为5 μL。采用二氯甲烷:甲醇=20:1的展开系统进行薄层层析,在ZF-2型三用紫外仪下观察化合物的薄层层析情况。薄层层析后在薄层板的一侧点样原点处点5 μL 0.2 mg·mL−1硫酸链霉素作为阳性对照。向灭菌的LB半固体培养基(琼脂质量浓度为5 g·L−1)[30]中加入一定量的供试细菌菌液(45 mL LB + 5 mL菌液),调至约108 CFU·mL−1。用移液枪将制备好的菌悬液均匀喷洒到层析后的薄层板上,待培养基冷却后,将薄层板置于28 ℃下黑暗保湿培养,12 h后在薄层板上均匀喷洒噻唑蓝(MTT),约10 min后观察试验结果。有抗菌活性成分处,供试细菌由于受到活性成分的抑制而出现白色抑菌斑;无抗菌活性成分处,供试细菌正常生长,与MTT反应显蓝色。根据抑菌斑直径的大小和多少来初步评价活性化合物的抑菌活性和数量,通过抑菌斑的迁移率(Rf)来初步判断样品中抗菌化合物的极性。
$ R_{\mathrm{f}}=\frac{\text { 抑菌斑与点样原点之间的距离 }}{\text { 展开剂前沿与点样原点之间的距离 }} $
-
为进一步阐明不同放线菌次生代谢产物的情况,采用高效液相色谱LC-16(岛津仪器(苏州)有限公司)对不同放线菌乙酸乙酯层提取物进行分析。分别称取20 mg浓缩后的提取物,加入1 mL色谱甲醇,超声溶解,再用0.22 μm的有机滤膜过滤,配置成浓度为20 mg·mL−1的溶液,备用。流动相为乙腈和水,流速为1 mL·min−1,WondaSil C18色谱柱(4.6 mm × 250 mm,5 μm,岛津仪器(苏州)有限公司),柱温为40 ℃,进样量为5 μL。色谱分析条件为:0~2 min,10%的乙腈等度洗脱;1~30 min,乙腈浓度由10%线性递增到100%;30~36 min,100%的乙腈等度洗脱;36~37 min,乙腈浓度由100%线性递减到10%;37~45 min,使用10%的乙腈对色谱柱进行再平衡;采用SPD-M20A二极管阵列检测器(岛津仪器(苏州)有限公司)进行全波长扫描检测。
-
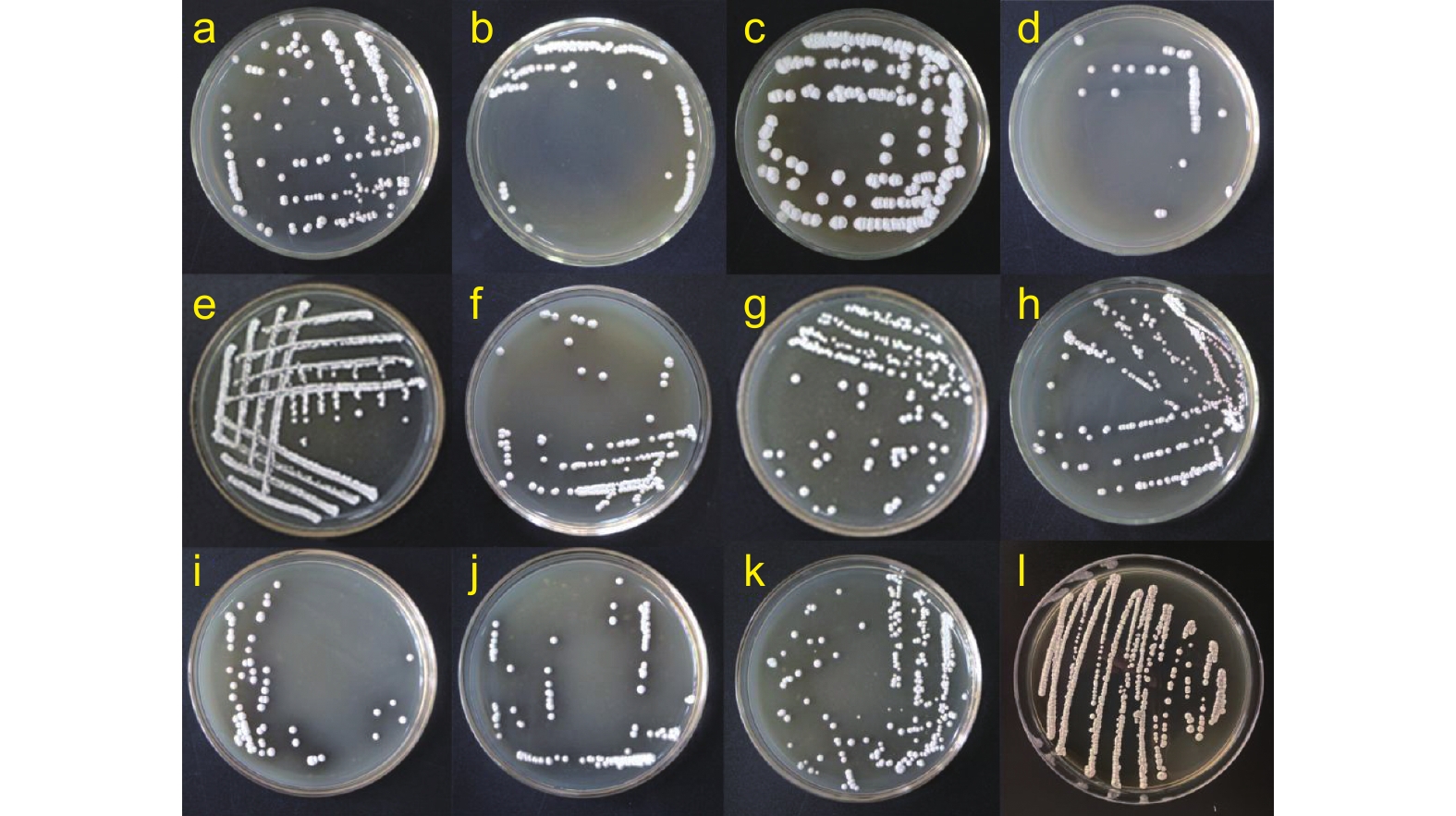
通过菌落形态观察,对从短枝木麻黄根际土壤中分离到的放线菌进行初步合并,最终从木麻黄根际土壤中共分离得到12株不同的放线菌,其菌落形态见图1。从图1中可看出:在相同的生长时间内,大多数放线菌单菌落为白色、圆形,但菌落的大小有所差别,菌株Ceaf-6的单菌落最大,生长速度较快;菌株Ceaf-9、Ceaf-19和Ceaf-23的单菌落相对较小,且菌株Ceaf-23后期培养基颜色会变深。

图 1 短枝木麻黄根际土壤分离到的放线菌
Figure 1. Actinomycetes isolated from the rhizosphere soil of Casuarina equisetifolia
-
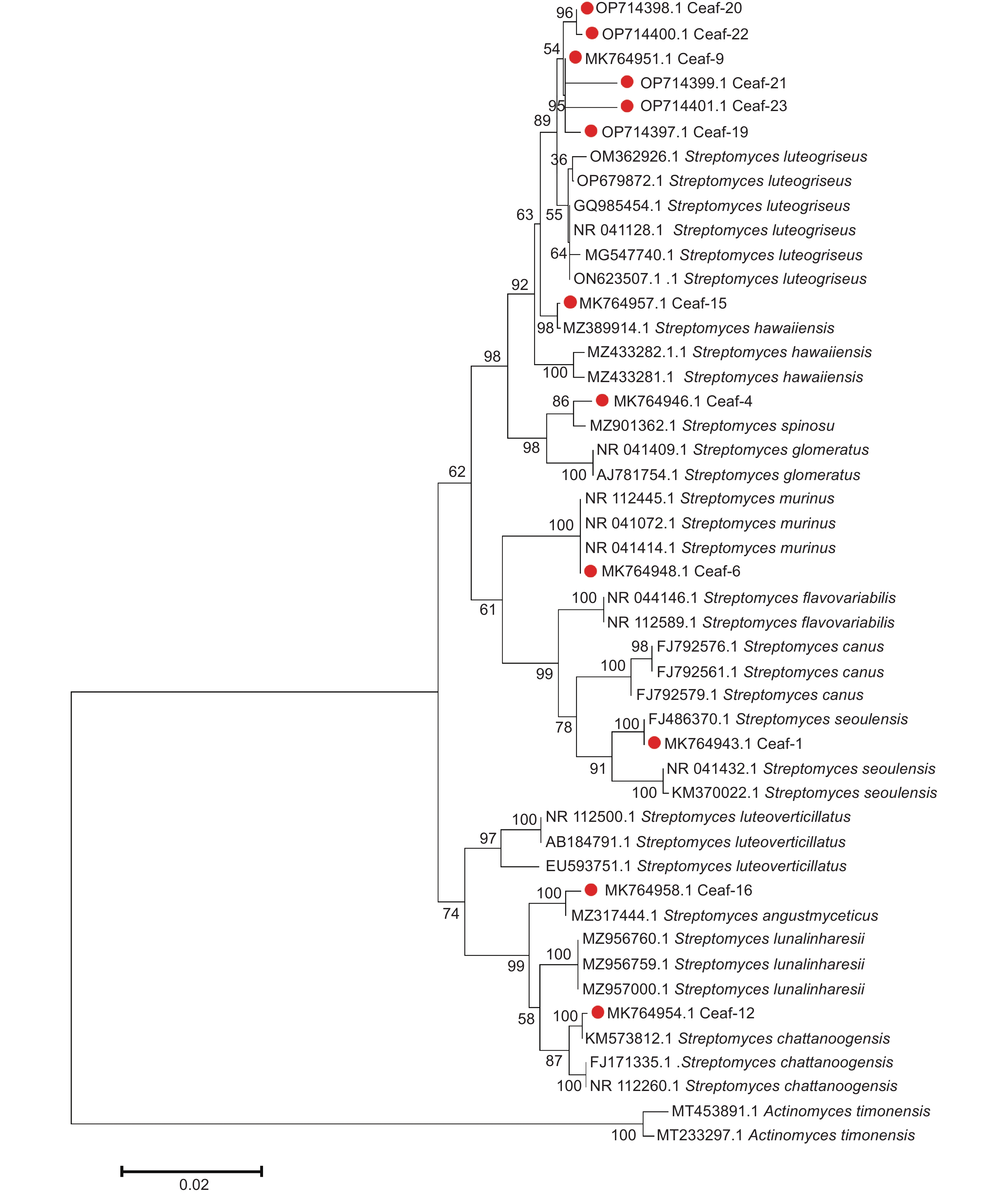
基于短枝木麻黄根际放线菌16S rRNA基因序列构建的系统发育树(图2)表明:菌株Ceaf-9、Ceaf-19、Ceaf-20、Ceaf-21、Ceaf-22和Ceaf-23与菌株S. luteogriseus聚在同一大支上,亲缘关系较近,但自展值只有89,且遗传距离也不相同,进一步通过序列比对发现并不是同一菌株,因此,以上菌株未鉴定到种。菌株Ceaf-15与S. hawaiiensis(登录号为MZ389914.1)聚在同一支上,且自展值为98,最大相似度为99.65%,因此,菌株Ceaf-15最终鉴定为S. hawaiiensis(登录号为MK764957)。按照同样的分类依据,综合菌落形态以及构建的系统发育树,短枝木麻黄根际放线菌的最终鉴定结果见表1。BLAST结果显示:所有菌株与最大相似菌株的相似性均在97%以上。分离到的12株短枝木麻黄根际放线菌均为链霉菌属(Streptomyces sp.)放线菌,说明链霉菌属放线菌为短枝木麻黄根际土壤中的优势菌群。

图 2 基于16S rRNA基因序列构建的短枝木麻黄根际放线菌系统进化树
Figure 2. Phylogenetic tree of rhizospheric actinomycetesin Casuarina equisetifolia based on gene sequences of 16S rRNA
表 1 短枝木麻黄根际放线菌鉴定结果
Table 1. Identification results of rhizospheric actinomycetes isolated from Casuarina equisetifolia
菌株
Strains登录号
Accession number鉴定结果
Identification results最大相似菌株
Closest related species相似度
Similarity/%Ceaf-1 MK764943 S. seoulensis FJ486370.1 S. seoulensis 99.93 Ceaf-4 MK764946 S. spinosus MZ901362.1 S. spinosus 99.44 Ceaf-6 MK764948 S. murinus NR041414.1 S. murinus 99.72 Ceaf-9 MK764951 Streptomyces sp. MG547740.1 S. luteogriseus 99.37 Ceaf-12 MK764954 S. chattanoogensis KM573812.1 S. chattanoogensis 99.93 Ceaf-15 MK764957 S. hawaiiensis MZ389914.1 S. hawaiiensis 99.65 Ceaf-16 MK764958 S. angustmyceticus MZ317444.1 S. angustmyceticus 99.86 Ceaf-19 OP714397 Streptomyces sp. ON623507.1 S. luteogriseus 99.16 Ceaf-20 OP714398 Streptomyces sp. GQ985454.1 S. luteogriseus 99.36 Ceaf-21 OP714399 Streptomyces sp. OP679872.1 S. luteogriseus 98.66 Ceaf-22 OP714400 Streptomyces sp. OM362926.1 S. luteogriseus 99.44 Ceaf-23 OP714401 Streptomyces sp. NR041128.1 S. luteogriseus 97.90 -
短枝木麻黄根际放线菌次生代谢产物对4种供试细菌的抑制活性见表2。4种供试细菌中,木麻黄青枯病菌和大肠杆菌为革兰氏阴性菌,枯草芽孢杆菌和溶血葡萄球菌为革兰氏阳性菌。由表2可知:不同放线菌次生代谢产物对4种供试细菌的抑制活性差异较大,但对革兰氏阳性菌和革兰氏阴性菌的抑制活性未表现出明显差异。菌株Ceaf-9和Ceaf-23提取物均未表现出任何抑菌活性,其他提取物均表现出一定的抑菌活性,但对不同供试细菌的抑菌活性差异较大;菌株S. spinosus Ceaf-4提取物对木麻黄青枯病菌、大肠杆菌及溶血葡萄球菌的抑菌斑直径均大于10 mm,表现出较强的抑菌活性;菌株S. chattanoogensis Ceaf-12对木麻黄青枯病菌的抑菌斑直径也大于10 mm,但菌株Ceaf-4抑菌斑的Rf值为0.30~0.57,而菌株Ceaf-12抑菌斑的Rf值为0.45~0.58,说明菌株Ceaf-4对青枯病菌的抑制活性更强;Rf越小,化合物的极性越大,多数放线菌抑菌斑的Rf值为0.30~0.50,说明具有活性的化合物多为极性中等偏大的化合物。综上所述,菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12对木麻黄青枯病菌表现出较强的抑制活性。
表 2 短枝木麻黄根际放线菌提取物对不同供试细菌的抑制活性
Table 2. Antibacterial activity of the crude extract from the rhizospheric actinomycetes of C. equisetifolia against different tested bacteria
供试样品
Tested samplesRf值(抑菌斑直径)
Rfvalue (Inhibition spot diameter)木麻黄青枯病菌
R.solanacearum枯草芽孢杆菌
B. subtilis大肠杆菌
E. coli溶血葡萄球菌
S.haemolyticusCeaf-1 — 0.48~0.52+ 0.52~0.53++ 0.22~0.27+; 0.32~0.37+ Ceaf-4 0.30~0.57+++ 0.40~0.52++ 0.37~0.53+++ 0.00~0.53+++ Ceaf-6 0.18~0.38++; 0.48~0.50++ — 0.53~0.55++ — Ceaf-9 — — — — Ceaf-12 0.45~0.58+++ 0.15~0.23+; 0.28~0.37++ 0.32~0.43++ 0.25~0.37++ Ceaf-15 0.47~0.50+ 0.32~0.37+ — — Ceaf-16 — 0.32~0.35+ — — Ceaf-19 0.33~0.45+ — — — Ceaf-20 0.45~0.55++ 0.42~0.52++ 0.47~0.50++ 0.33~0.53+++ Ceaf-21 0.45~0.58++ 0.40~0.45++ 0.48~0.53++ 0.33~0.52++ Ceaf-22 0.38~0.52+ 0.23~0.43++ 0.28~0.42++ — Ceaf-23 — — — — 硫酸链霉素 +++ +++ +++ +++ 注:“—”表示无抗菌活性; “+”表示抑菌斑最大直径d<5 mm,“++ ”表示5 mm≤d<10 mm,“+++”表示d ≥10 mm;阳性对照硫酸链霉素仅在原位点样。
Notes: —: Inhibition spot was not observed;+: Maximum inhibition spot diameter (d)<5 mm;++ : 5 mm≤ d <10 mm;+++: d ≥10 mm;Thepositive control streptomycin sulfate was only sampled on TLC plate. -
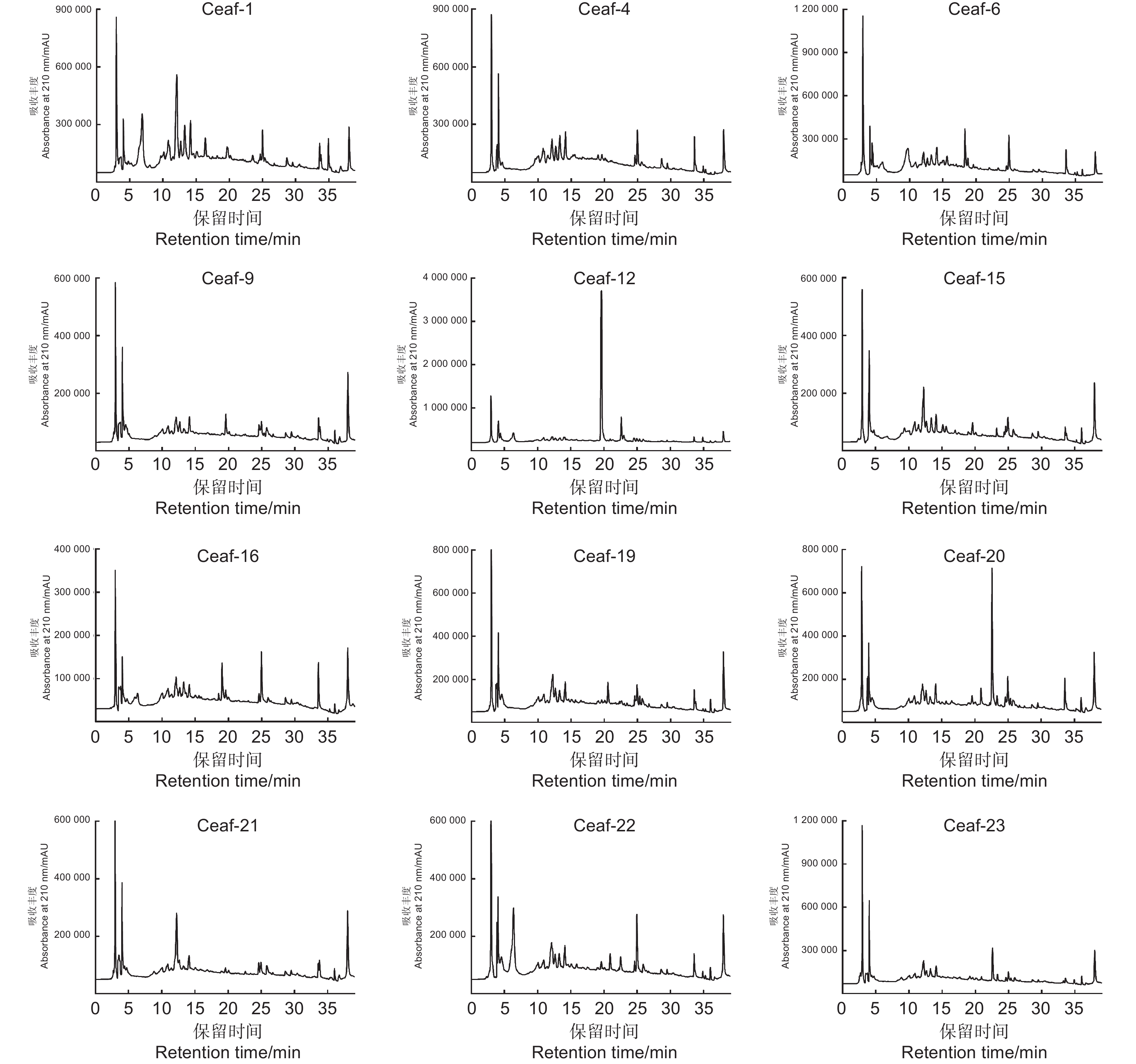
12株短枝木麻黄根际放线菌乙酸乙酯层次生代谢产物的HPLC-UV色谱图见图3。从图3可看出:12株放线菌均含有一定数量的次生代谢产物,但不同化合物的相对含量(峰高)差别较大。抗细菌活性结果(表2)表明,菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12对木麻黄青枯病菌表现出较强的抑制活性,但图3表明这2株放线菌的次生代谢产物具有明显的区别,S. spinosus Ceaf-4中的各个次生代谢产物相对含量差别不大,保留时间(Rt)集中在10~15 min,通过紫外吸收分析发现为结构类似的化合物;而S. chattanoogensis Ceaf-12在Rt = 20 min左右有一个明显的吸收峰,说明此化合物的含量较高。此外菌株Ceaf-1、Ceaf-6和Ceaf-20也含有丰富的次生代谢产物,但抗菌活性一般。综合考虑抗菌活性和次生代谢产物的情况,菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12可作为候选菌株进一步分离其中的活性成分。

图 3 短枝木麻黄根际放线菌提取物的HPLC色谱图
Figure 3. HPLC chromatograms for crude extracts of rhizospheric actinomycetes isolated from C. equisetifolia
-
木麻黄是我国南方尤其是华南地区重要的经济林和生态林树种,而青枯病作为木麻黄的“癌症”,严重影响其种植和生长,影响我国的生态安全以及沿海防护林的建设[8, 31]。根际微生物作为植物的第二套基因组,在植物保护中起着重要作用[32]。近年来,随着生物信息学技术的不断成熟,木麻黄根际微生物的研究也逐渐成为研究的热点。木麻黄根际土壤中特征微生物的含量差异明显,细菌分布量最大,其次是真菌和放线菌。随着栽植代数的增加,细菌含量减少,而真菌含量增加[33]。同时,木麻黄根系分泌物-酚酸对根际土壤微生物群落具有特异选择性,引起根际微生物群落结构失衡,进而导致连栽障碍问题[34]。本研究采用热击稀释法从短枝木麻黄根际土壤中共分离出12株放线菌,均为链霉菌属放线菌,说明链霉菌属放线菌为短枝木麻黄根际土壤的优势菌株,这与Abhijit等[26]的研究结果一致。根际微生物的组成受到植物种类、土壤类型、季节变化、环境条件和栽培管理制度等各种因素的影响[35]。木麻黄作为我国华南和东南沿海地区重要的生态防护林和用材林树种[36],其生长环境土壤贫瘠、含盐量高、有机质含量少,且植被资源匮乏,物种单一,是造成放线菌种类相对单一的重要原因。此外,并不是所有根际放线菌都能在人工培养基上培养,因此,分离到的这12株放线菌只是短枝木麻黄根际放线菌的一部分。前期的研究表明,从海芒果(Cerbera manghas L.)、红豆杉(Taxus wallichiana var. chinensis (Pilg.) Florin)等植物根际土壤中分离出的链霉菌属放线菌比例均比较高[37]。
目前,放线菌已广泛应用于农业、生物和医药等多个领域。放线菌是产生抗生素最多的一类生物群体,而链霉菌属又是产生抗生素最多的一类放线菌[16, 38]。根际放线菌种类丰富,代谢途径各异,可产生一系列的次生代谢产物从而抑制病原菌在植物根际的生长和繁殖[22]。近年来,木麻黄青枯病的大面积发生严重影响我国沿海的生态安全,从木麻黄根际土壤中寻找新的活性菌株以及次生代谢产物,为病害的生物防治提供了一个新的思路。本研究的抗菌活性结果表明,菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12对木麻黄青枯病菌均表现出较好的抑制活性,其抑菌斑直径均大于10 mm。HPLC结果表明,菌株S. spinosus Ceaf-4的提取物中含有丰富的次生代谢产物,菌株S. chattanoogensis Ceaf-12在Rt = 20 min左右时存在一个紫外吸收较高的化合物,2株放线菌中的抗菌活性成分是否与这些化合物有关值得进一步研究。本研究只采用了M1培养基对不同放线菌进行发酵,在后续研究中可改变培养基或培养条件,有效激活沉默基因的表达,可诱发菌株产生新的活性化合物,同时利用LC-MS/MS构建不同菌株次生代谢产物的可视化分子网络,快速探寻新型的化合物及其同系物[39]。随着现代科学技术的发展,高通量测序技术已广泛应用于根际微生物的研究中,探究开发根部有益微生物为病虫害的防治提供了广阔的前景[14]。在后续的研究中可结合高通量测序技术研究木麻黄根际放线菌的生物多样性,同时尝试采用不同的培养基,分离得到尽可能多的放线菌菌株,并比较不同培养基中放线菌的分布和生长情况。
-
通过对华南沿海短枝木麻黄根际土壤样品中放线菌的分离培养,共分离得到12株不同的链霉菌属放线菌,链霉菌属放线菌为短枝木麻黄根际土壤中的优势放线菌。进一步对筛选出的菌株进行发酵及抗菌活性研究,筛选出一些抗菌活性较好的菌株,其中,菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12对木麻黄青枯病菌表现出较好的抑制活性。HPLC分析发现,菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12的次生代谢产物类型丰富,值得进一步对其化合物进行分离制备,可作为候选抗菌活性菌株进一步开发和利用。
短枝木麻黄根际放线菌及其抗细菌活性
Antibacterial Activities of Rhizospheric Actinomycetes Associated with Casuarina equisetifolia
-
摘要:
目的 分离并鉴定短枝木麻黄根际土壤中的放线菌,测定短枝木麻黄根际放线菌的抗细菌活性,并从中筛选出活性放线菌菌株。 方法 采用热击稀释法分离短枝木麻黄根际土壤中的放线菌,通过形态学观察和分子生物学相结合的方法对分离得到的放线菌菌株进行鉴定,采用薄层层析(TLC)-生物自显影法测定短枝木麻黄根际放线菌次生代谢产物对不同供试细菌的抑制活性,进一步采用高效液相色谱(HPLC)分析根际放线菌乙酸乙酯层提取物中次生代谢产物的情况。 结果 从短枝木麻黄根际土壤中共分离鉴定得到12株放线菌,均为链霉菌属(Streptomyces sp.)放线菌,最大相似度均在97%以上。活性测定的结果表明:菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12对木麻黄青枯病菌(Ralstonia solanacearum)表现出较强的抑制活性,抑菌斑的直径均在10 mm以上;菌株S. spinosus Ceaf-4对大肠杆菌(Escherichia coli)和溶血葡萄球菌(Staphylococcus haemolyticus)的抑菌斑直径也大于10 mm。高效液相色谱分析的结果表明,菌株S.spinosus Ceaf-4和S. chattanoogensis Ceaf-12均含有丰富的次生代谢产物,但次生代谢产物的类型和相对含量均存在明显差异。 结论 短枝木麻黄根际土壤中存在以链霉菌属为优势种群的放线菌,菌株S. spinosus Ceaf-4和S. chattanoogensis Ceaf-12对木麻黄青枯病菌表现出较强的抑制活性且含有丰富的次生代谢产物,可作为候选菌株进一步发掘其抗菌活性成分。 Abstract:Objective This study aimed to isolate and identify actinomycetes in rhizospheric soil of Casuarina equisetifolia, determine the antibacterial activity of the rhizospheric actinomycetes, and screen out the actinomycetes isolates with antimicrobial activities. Methods The actinomycetes in the rhizospheric soil of C. equisetifolia were isolated by heat strike dilution method. The actinomycetes isolates were further identified by combining morphological observation with molecular biology method. The antibacterial activities of secondary metabolites produced by the rhizospheric actinomycetes against different bacteria were determined using TLC-bioautography assay. The secondary metabolites in the ethyl acetate layer of rhizospheric actinomycetes were analyzed by HPLC (high performance liquid chromatography). Results A total of twelve actinomycetes strains were isolated and identified from the rhizospheric soil of C. equisetifolia, all of which were the genus of Streptomyces sp., and the maximum similarities were all above 97%. S. spinosus Ceaf-4 and S. chattanoogensis Ceaf-12 had stronger antibacterial activity against Ralstonia solanacearum, and the diameters of antibacterial spots were more than 10 mm. The antibacterial spot diameters of strain S. spinosus Ceaf-4 against Escherichia coli and Staphylococcus haemolyticus were also greater than 10 mm. In addition, S. spinosus Ceaf-4 and S. chattanoogensis Ceaf-12 both contained abundant secondary metabolites, but there were significant difference in the types and relative contents of secondary metabolites. Conclusion There are actinomycetes with Streptomyces sp. as the dominant population in the rhizospheric soil of C. equisetifolia. S. spinosus Ceaf-4 and S. chattanoogensis Ceaf-12 show the stronger inhibitory activity against R. solanacearum and contain abundant secondary metabolites. They can be used as candidate active strains to further explore their antibacterial active components. -
图 1 短枝木麻黄根际土壤分离到的放线菌
Figure 1. Actinomycetes isolated from the rhizosphere soil of Casuarina equisetifolia
图 2 基于16S rRNA基因序列构建的短枝木麻黄根际放线菌系统进化树
Figure 2. Phylogenetic tree of rhizospheric actinomycetesin Casuarina equisetifolia based on gene sequences of 16S rRNA
图 3 短枝木麻黄根际放线菌提取物的HPLC色谱图
Figure 3. HPLC chromatograms for crude extracts of rhizospheric actinomycetes isolated from C. equisetifolia
表 1 短枝木麻黄根际放线菌鉴定结果
Table 1. Identification results of rhizospheric actinomycetes isolated from Casuarina equisetifolia
菌株
Strains登录号
Accession number鉴定结果
Identification results最大相似菌株
Closest related species相似度
Similarity/%Ceaf-1 MK764943 S. seoulensis FJ486370.1 S. seoulensis 99.93 Ceaf-4 MK764946 S. spinosus MZ901362.1 S. spinosus 99.44 Ceaf-6 MK764948 S. murinus NR041414.1 S. murinus 99.72 Ceaf-9 MK764951 Streptomyces sp. MG547740.1 S. luteogriseus 99.37 Ceaf-12 MK764954 S. chattanoogensis KM573812.1 S. chattanoogensis 99.93 Ceaf-15 MK764957 S. hawaiiensis MZ389914.1 S. hawaiiensis 99.65 Ceaf-16 MK764958 S. angustmyceticus MZ317444.1 S. angustmyceticus 99.86 Ceaf-19 OP714397 Streptomyces sp. ON623507.1 S. luteogriseus 99.16 Ceaf-20 OP714398 Streptomyces sp. GQ985454.1 S. luteogriseus 99.36 Ceaf-21 OP714399 Streptomyces sp. OP679872.1 S. luteogriseus 98.66 Ceaf-22 OP714400 Streptomyces sp. OM362926.1 S. luteogriseus 99.44 Ceaf-23 OP714401 Streptomyces sp. NR041128.1 S. luteogriseus 97.90  下载: 导出CSV
下载: 导出CSV
表 2 短枝木麻黄根际放线菌提取物对不同供试细菌的抑制活性
Table 2. Antibacterial activity of the crude extract from the rhizospheric actinomycetes of C. equisetifolia against different tested bacteria
供试样品
Tested samplesRf值(抑菌斑直径)
Rfvalue (Inhibition spot diameter)木麻黄青枯病菌
R.solanacearum枯草芽孢杆菌
B. subtilis大肠杆菌
E. coli溶血葡萄球菌
S.haemolyticusCeaf-1 — 0.48~0.52+ 0.52~0.53++ 0.22~0.27+; 0.32~0.37+ Ceaf-4 0.30~0.57+++ 0.40~0.52++ 0.37~0.53+++ 0.00~0.53+++ Ceaf-6 0.18~0.38++; 0.48~0.50++ — 0.53~0.55++ — Ceaf-9 — — — — Ceaf-12 0.45~0.58+++ 0.15~0.23+; 0.28~0.37++ 0.32~0.43++ 0.25~0.37++ Ceaf-15 0.47~0.50+ 0.32~0.37+ — — Ceaf-16 — 0.32~0.35+ — — Ceaf-19 0.33~0.45+ — — — Ceaf-20 0.45~0.55++ 0.42~0.52++ 0.47~0.50++ 0.33~0.53+++ Ceaf-21 0.45~0.58++ 0.40~0.45++ 0.48~0.53++ 0.33~0.52++ Ceaf-22 0.38~0.52+ 0.23~0.43++ 0.28~0.42++ — Ceaf-23 — — — — 硫酸链霉素 +++ +++ +++ +++ 注:“—”表示无抗菌活性; “+”表示抑菌斑最大直径d<5 mm,“++ ”表示5 mm≤d<10 mm,“+++”表示d ≥10 mm;阳性对照硫酸链霉素仅在原位点样。
Notes: —: Inhibition spot was not observed;+: Maximum inhibition spot diameter (d)<5 mm;++ : 5 mm≤ d <10 mm;+++: d ≥10 mm;Thepositive control streptomycin sulfate was only sampled on TLC plate.
下载: 导出CSV
-
[1] 白 莹, 周柳婷, 张 晨, 等. 外源微生物对木麻黄幼苗生长和土壤微生物群落的影响[J]. 应用生态学报, 2021, 32(8):2939-2948. [2] 单体江, 秦 楷, 谢银燕, 等. 木麻黄内生真菌次生代谢产物及生物活性[J]. 华南农业大学学报, 2019, 40(3):67-74. [3] 韩 强, 仲崇禄, 张 勇, 等. 山地木麻黄种源在海南临高的遗传变异及选择[J]. 林业科学研究, 2017, 30(4):595-603. [4] 许秀玉, 徐 斌, 甘先华, 等. 木麻黄青枯病菌的分离及强致病菌株的筛选[J]. 林业科学研究, 2017, 30(3):409-416. [5] MODHUMITA G D, RADHA V, KARPAGA R S B. Characterization of genes expressed in Casuarina equisetifolia in response to elicitation by cell wall components of Trichosporium vesiculosum[J]. Silvae Genetica, 2017, 62(4): 161-173. [6] GOPICHAND C V, ANJANI A, HIMAJA A, et al. Phytochemical evaluation of Lantana camara, Casuarina equisetifolia, Michella nilagirica[J]. International Journal of Research in Pharmacy and Biosciences, 2015, 3(6): 461-463. [7] 孙 战, 李 明, 魏永成, 等. 木麻黄青枯病发生与土壤五种元素含量分析[J]. 分子植物育种, 2023, 21(4):1313-1321. [8] 谢银燕, 王 松, 吴春银, 等. 木麻黄病虫害及其防治的最新进展[J]. 江苏农业科学, 2019, 47(20):36-41. [9] SU Y, XU Y, LI Q, et al. The essential genome of Ralstonia solanacearum[J]. Microbiological Research, 2020, 238: 126500. doi: 10.1016/j.micres.2020.126500 [10] ZHANG Y, LI G, QIAO L, et al. Identification and characterization of virulence-attenuated mutants in Ralstonia solanacearum as potential biocontrol agents against bacterial wilt of Pogostemon cablin[J]. Microbial Pathogenesis, 2020, 147: 104418. doi: 10.1016/j.micpath.2020.104418 [11] 史芳芳, 李向泉. 葡萄根际土壤真菌群落多样性分析[J]. 中国农业科技导报, 2019, 21(7):47-58. [12] KAUR R, KAUR S, KAUR G. Molecular and physiological manipulations in rhizospheric bacteria[J]. Acta Physiologiae Plantarum, 2021, 43(5): 77. doi: 10.1007/s11738-021-03251-z [13] 蒋靖怡, 王铁霖, 池秀莲, 等. 基于高通量测序的紫花丹参与白花丹参根际细菌群落结构研究[J]. 中国中药杂志, 2019, 44(8):1545-1551. doi: 10.19540/j.cnki.cjcmm.20190301.015 [14] 王 端, 姚香梅, 叶 健. 根际微生物-植物-病毒-介体昆虫多元互作研究进展[J]. 生物技术通报, 2018, 34(2):54-65. [15] SELIM M S M, ABDELHAMID S A, MOHAMED S S. Secondary metabolites and biodiversity of actinomycetes[J]. Journal of Genetic Engineering and Biotechnology, 2021, 19(1): 72. [16] XIE C L, XIA J M, WANG J S, et al. Metabolomic investigations on Nesterenkonia flava revealed significant differences between marine and terrestrial actinomycetes[J]. Marine Drugs, 2018, 16(10): 356. doi: 10.3390/md16100356 [17] ELSHAFIE H S, CAMELE I. Rhizospheric actinomycetes revealed antifungal and plant-growth-promoting activities under controlled environment[J]. Plants-Basel, 2022, 11(14): 1872. doi: 10.3390/plants11141872 [18] 方 正, 詹 伟, 吴庆珊, 等. 金钗石斛根际土壤放线菌的分离及抑菌活性筛选[J]. 贵州师范大学学报(自然科学版), 2018, 36(6):47-53. [19] 叶景静, 郑红芸, 吴 越, 等. 广西茅尾海红树林植物根际土壤放线菌多样性及抗菌活性研究[J]. 中国病原生物学杂志, 2018, 13(11):1221-1226,1231. [20] 吴 佳, 李晓霞, 舒伟学, 等. 缬草内生菌和根际放线菌的分离及安莎类抗生素的筛选[J]. 西北农林科技大学学报, 2019, 47(6):107-114. [21] KUMARI N, MENGHANI E, MITHAL R. Bioactive compounds characterization and antibacterial potentials of actinomycetes isolated from rhizospheric soil[J]. Journal of Scientific & Industrial Research, 2019, 78(11): 793-798. [22] 杜用玺, 蒋靖怡, 王铁霖, 等. 拮抗丹参根腐病病原菌的丹参根际放线菌筛选研究[J]. 中国植保导刊, 2020, 40(9):17-22. [23] 段佳丽, 薛泉宏, 舒志明, 等. 放线菌Act12与腐植酸钾配施对丹参生长及其根域微生态的影响[J]. 生态学报, 2015, 35(6):1807-1819. [24] 张 昕, 沈爱华, 林永敬, 等. Frankia菌侵染木麻黄与其共生结瘤程序[J]. 浙江农林大学学报, 2013, 30(5):669-673. [25] GUNASEKERA D T W, GUNAWARDANA D, JAYASINGHEARACHCHI H. Diversity of actinomycetes in nitrogen fixing root nodules of Casuarina equisetifolia and its impact on plant growth[J]. International Journal of Multidisciplinary Studies, 2016, 3(1): 17-24. doi: 10.4038/ijms.v3i1.79 [26] ABHIJIT K. Colony characters and antagonism of Streptomyces against bacteral and fungal pathogens[J]. Bioinfolet, 2010, 7(2): 153-155. [27] 王 鹤, 彭 楚, 来航线, 等. 太白山土壤放线菌分离方法及生态分布规律[J]. 干旱地区农业研究, 2017, 35(5):107-114. [28] 李莎莎, 李 琴, 李翊铭, 等. 海洋来源链霉菌IMB18-531与枝孢菌IMB19-099共培养代谢产物研究[J]. 药学学报, 2023, 58(4): 967-974. [29] 田浩杰, 李豆南, 邱树毅, 等. 酱香大曲中高温放线菌的筛选及基因组解析[J]. 食品科学, 2023 网络首发尚未正式发表 [30] MAO Z, ZHANG W, WU C, et al. Diversity and antibacterial activity of fungal endophytes from Eucalyptus exserta[J]. BMC Microbiology, 2021, 21(1): 155. doi: 10.1186/s12866-021-02229-8 [31] 谢银燕. 木麻黄内生真菌和根际微生物及其抗细菌活性[D]. 广州: 华南农业大学, 2019. [32] MULLER D B, VOGEL C, BAI Y, et al. The plant microbiota: systems-level insights and perspectives[J]. Annual Review of Genetics, 2016, 50(1): 211-234. doi: 10.1146/annurev-genet-120215-034952 [33] 李建鹃, 罗 扬, 周柳婷, 等. 连栽木麻黄根际微生物群落结构和功能特征[J]. 森林与环境学报, 2020, 40(1):9-15. [34] 周柳婷, 罗 扬, 李建鹃, 等. 酚酸介导下连栽木麻黄根际微生物变化[J]. 生态学杂志, 2021, 40(4):1021-1028. [35] 曹曼曼, 王 飞, 周北海, 等. 铁尾矿芦苇根际微生物和根内生菌群落分布及其限制性因子解析[J]. 环境科学, 2021, 42(10):4998-5009. [36] 张 勇, 仲崇禄, 陈 羽, 等. 海南木麻黄无性系生长过程研究[J]. 林业科学研究, 2017, 30(4):588-594. [37] 袁丽杰, 章广玲, 张玉琴, 等. 药用植物根际放线菌的种群多样性及生物活性初步研究[J]. 中国抗生素杂志, 2009, 34(8):463-466,504. [38] WU H, LIU B, OU X, et al. Streptomyces thermoalkaliphilus sp. nov. an alkaline cellulase producing thermophilic actinomycete isolated from tropical rainforest soil[J]. Antonie van Leeuwenhoek, 2017, 111(3): 413-422. [39] KHUSHI S, NAHAR L, SALIM A, et al. Cacolides: Sesterterpene butenolides from a southern australian marine sponge, Cacospongia sp.[J]. Marine Drugs, 2018, 16(11): 456-467. doi: 10.3390/md16110456 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2273
- HTML全文浏览量: 1561
- PDF下载量: 54
- 被引次数: 0

