

 下载:
下载:
-
核桃(Juglans regia Linnaeus)又叫胡桃,是我国重要的经济林树种。其坚果具有较高的营养价值,有良好的保健作用。我国核桃的生产与出口位居世界第一,根据FAO资料统计,在2020年,我国的核桃种植面积为48.71万公顷,核桃产量高达110万吨[1]。其中核桃的种植区主要分布在新疆、云南、四川、陕西等地,而新疆拥有独特的地理环境造就了核桃优良的品种,因此核桃产业的发展被当地高度重视。苹果蠹蛾(Cydia pomonella Linnaeus)是世界性检疫害虫,主要为害苹果(Malus pumila Mill)、桃(Prunus persica Linnaeus)、海棠(Malus spectabilis Borkh)、核桃等仁果类和坚果类果树[2]。在中国新疆和田县,于2018年首次发现苹果蠹蛾为害核桃,并造成20%~39%的产量损失,近几年来苹果蠹蛾在核桃产区不断扩散。目前该虫的防治主要依靠化学防治,这与绿色防控和生产绿色食品的原则相悖。因此有效利用植物的抗虫性对害虫的防治具有着重要意义,也符合当前绿色环保的防控策略。
植物与昆虫相互作用是生物多样性的驱动力,在漫长的协同进化过程中,植物已形成相应的防御措施,以确保植物自身的正常生长[3-4]。最直接的方式是植物通过产生多种代谢物直接作用植食性昆虫或威慑植食性昆虫来保护自身,如植物代谢物质胡桃醌可以毒杀舞毒蛾(Lymantria dispar Linnaeus)和甘蓝夜蛾(Mamestra brassicae Linnaeus)幼虫,进而提高植物对植食性昆虫的抗性[5]。另外当植食性昆虫为害时,植物可以改变自身的营养物质来抑制植食性昆虫的生长发育[6-7]。已有研究表明营养物质是影响昆虫生长发育与繁殖的重要因素,尤其是植物体内的氮和糖[8-9]。植物被害虫为害后,为了应对害虫的胁迫,植物的相关防御酶被启动,酶活性的变化则反应寄主植物的抗虫反应[10]。已有研究表明过氧化氢酶(catalase,CAT)、过氧化物酶(peroxidase,POD)、超氧化物歧化酶(superoxide dismutase,SOD)等在植食性昆虫胁迫时诱导酶活性的改变,并生成相关的防御体系[11-13]。在田间观察发现被苹果蠹蛾为害后,核桃果皮蛀孔处组织褐化,部分细胞死亡,通过室内饲喂发现与取食核仁相比取食桃外果皮的苹果蠹蛾生长发育已受到严重影响。此外,本课题组在野外调查发现核桃不同品种被蛀率之间存在着较大的差异。因此,通过研究苹果蠹蛾蛀食核桃果皮营养物质、代谢物质及酶活性的动态变化,探明核桃果皮对苹果蠹蛾取食胁迫的防御反应,为抗苹果蠹蛾核桃品种的选育提供更科学的理论依据。
-
试剂及仪器:供试药剂均是国产分析纯(苏州格锐思生物科技有限公司),人工气候培养箱RXZ-260A、水浴锅HH-M6、紫外分光光度计Each 840-37(上海俊升科学器材有限公司),高效液相色谱仪LC-100(上海伍丰科学仪器有限公司),移液枪Eppendorf 1~100 mL、50~200 µL(德国艾本德公司),离心机TLG-16、酶标仪Multiskan FC(美国ThermoFisher Scientific公司),研钵(天翼沣生物科技有限公司)。
虫源:苹果蠹蛾虫源采自于新疆维吾尔自治区和田地区和田县拉依喀乡核桃园中,将采集的老熟幼虫带回室内置于人工气候培养箱中连续饲喂2代,饲养条件为温度(25 ± 1) ℃,湿度(60 ± 5)%,光周期16∶8(L∶D),幼虫以核桃果实饲喂,成虫以10%的蜂蜜水补充能量,将幼虫饲喂至4龄备用。
供试核桃品种为‘扎343’,种植于和田县拉依喀乡,选取长势一致、无病虫害的核桃果实作为试验材料。
-
接虫:将供试苹果蠹蛾4龄幼虫接虫前进行6 h的饥饿处理备用,在野外选择进入硬核期(花后80 d)的大小一致、无病虫害核桃果实供试。以小毛笔轻轻的将备用的苹果蠹蛾4龄幼虫接于核桃果实表面,每颗核桃果实接种1头幼虫,并以网兜将果实包裹防止幼虫逃逸,以未接虫的核桃果实作对照(为害0 h)。
收样:从幼虫蛀食果实外果皮开始计时,分别接虫12、24、36、48和72 h后取样,每个时间段生物学重复3次,共计接虫核桃果实90颗。为了避免果实不同发育时期造成的影响,以不同时间接虫,同一时间收集核桃果皮。将所取的样品以纯净水将果实表面清洗干净,以手术刀切取蛀孔周围1 cm2的核桃果皮,将切取的样品在冰块上切成小块,迅速置于液氮罐中,备用于各项指标的测定。
-
可溶性糖含量的测定:采用蒽酮比色法[14]。
可溶性蛋白含量的测定:采用考马斯亮蓝G-250法[15]。
-
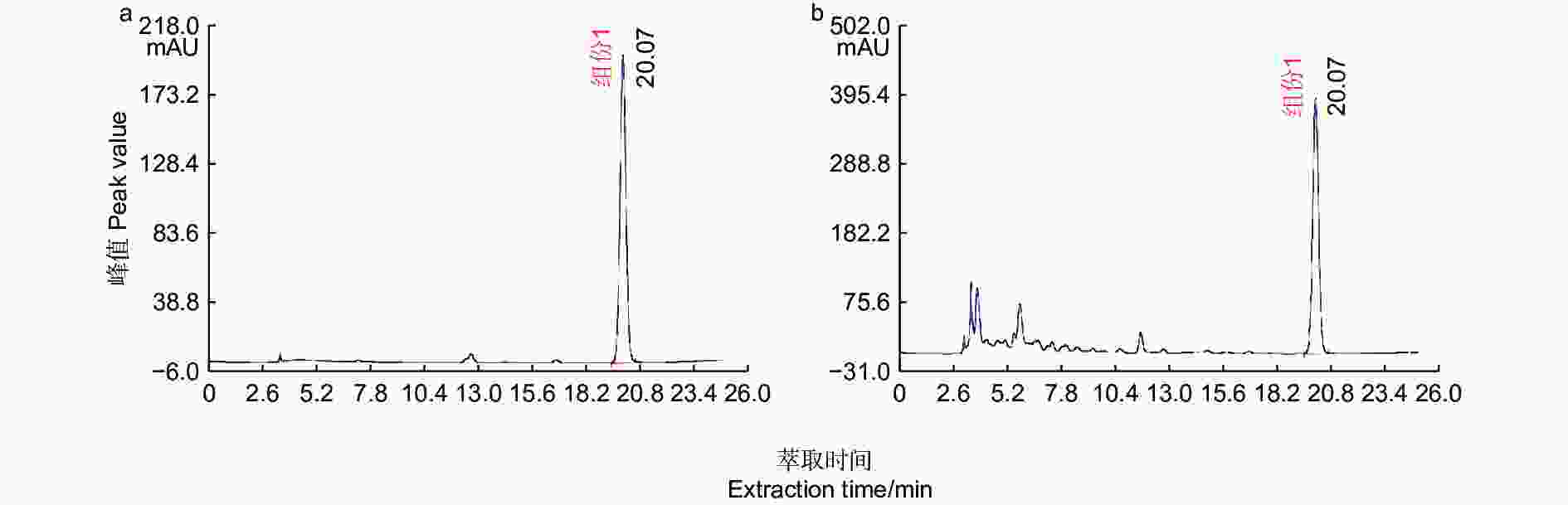
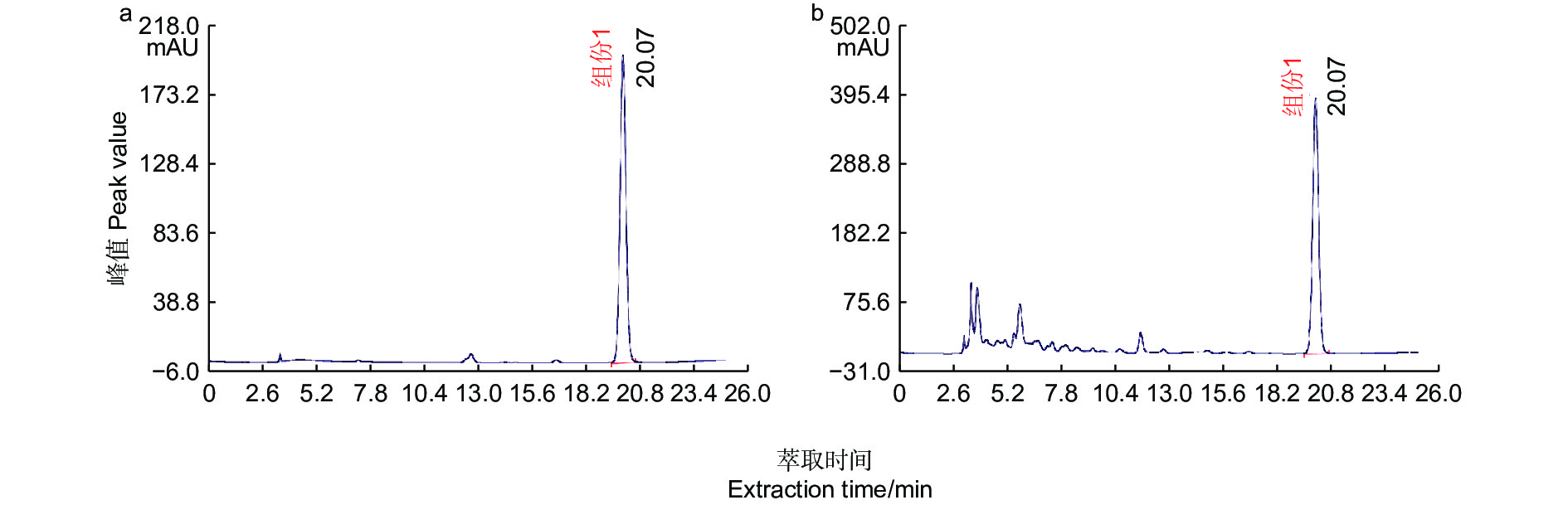
图 1 胡桃醌标准品(a)与核桃果皮(b)的HPLC
Figure 1. HPLC of Juglone standard (a) and walnut husk (b)
单宁含量的测定:采用试剂盒测定,试剂盒购自苏州格锐思生物科技有限公司。称取约0.1 g核桃果皮鲜样,加入1 mL蒸馏水,以研钵充分匀浆后转移至离心管中,用80 ℃水浴锅水浴提取1 h,之后以12 000 r·min−1、4 ℃的条件下离心10 min。取上清液10 µL与130 µL的蒸馏水加入离心管中,加入试剂一40 µL,试剂二60 µL,用蒸馏水做标准曲线,以酶标仪在波长650 nm下检测单宁的含量。
总黄酮含量的测定:采用NaNO2-Al(NO3)3-NaOH显色法测定黄酮总含量,以苏州格锐思生物科技有限公司的试剂盒进行测定。称取约0.1 g核桃果皮鲜样,加入1.5 mL的60%乙醇研磨,60 ℃振荡提取2 h(若蒸发用60%乙醇定容至1.5 mL),在12 000 r·min−1、25 ℃的条件下离心10 min,将离心管中加入50 µL的上清液与15 µL的试剂一、30 µL试剂二和105 µL试剂三迅速混匀。用蒸馏水做标准曲线,以酶标仪在波长510 nm下检测总黄酮含量。
-
酶液的提取:称取约0.1 g核桃果皮鲜样,放入带有液氮的研钵中研磨后加入1 mL提取液,用离心机在12 000 r·min−1、4 ℃条件下离心10 min,上清液则为酶液提取液。提取液中含有50 mmol·L−1(pH 7.8)的磷酸缓冲液和1%的聚乙烯吡咯烷酮,所用试剂盒购自苏州格锐思生物科技有限公司。
过氧化物酶(POD)活性的测定:采用愈创木酚法[17]。
过氧化氢酶(CAT)活性的测定:采用钼酸铵法[18]。
超氧化物歧化酶(SOD)活性的测定:采用黄嘌呤氧化酶法[19]。
-
采用Microsoft Excel 2013进行数据汇总,采用SPSS 22统计分析软件对数据进行方差显著性分析,采用Duncan’s多重比较进行检验相关指标的差异性,采用origin 2018软件对数据进行处理与绘图。
-
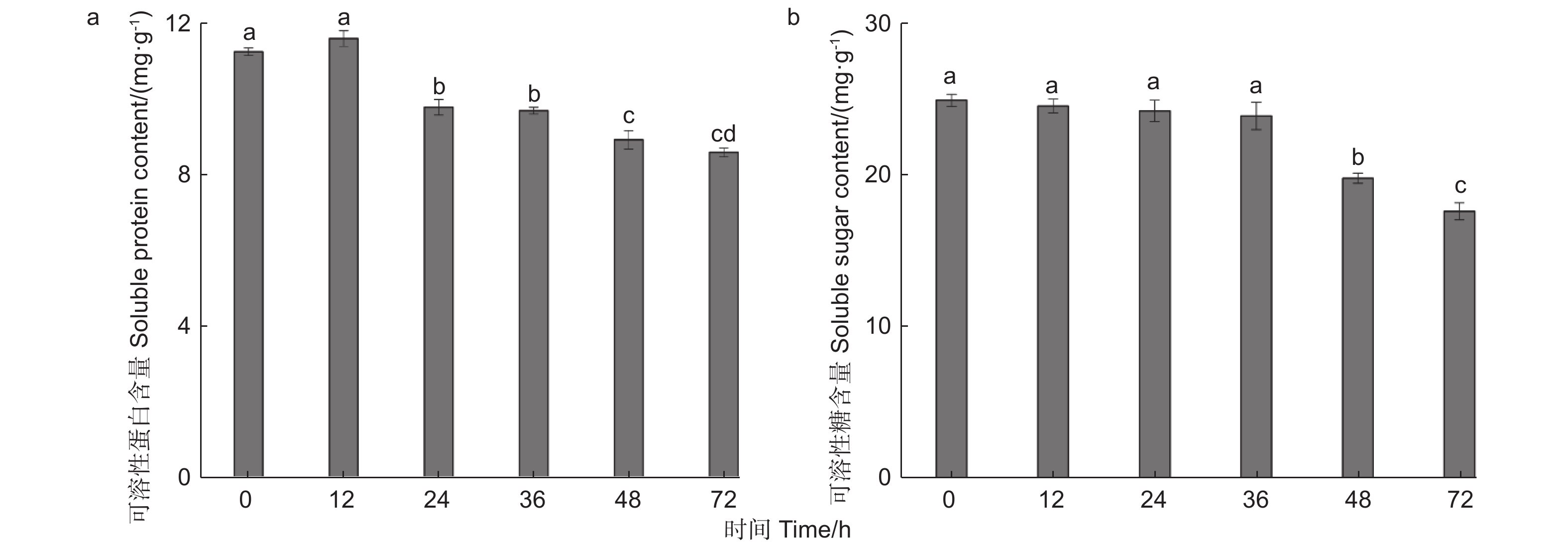
由图2a可知,核桃果皮可溶性蛋白的含量随苹果蠹蛾蛀食时间的推移存在较大差异。苹果蠹蛾为害12 h时与对照相比可溶性蛋白含量增加,增加量为3.15%,但无显著差异(P0.05)。然而,在为害24、36、48、72 h时可溶性蛋白含量持续降低,与对照相比分别低于13.02%、13.81%、20.67%、23.64%,差异水平均显著(P<0.05)。可见在为害24 h时,可溶性蛋白含量显著降低,核桃果皮可能开始产生防御反应。
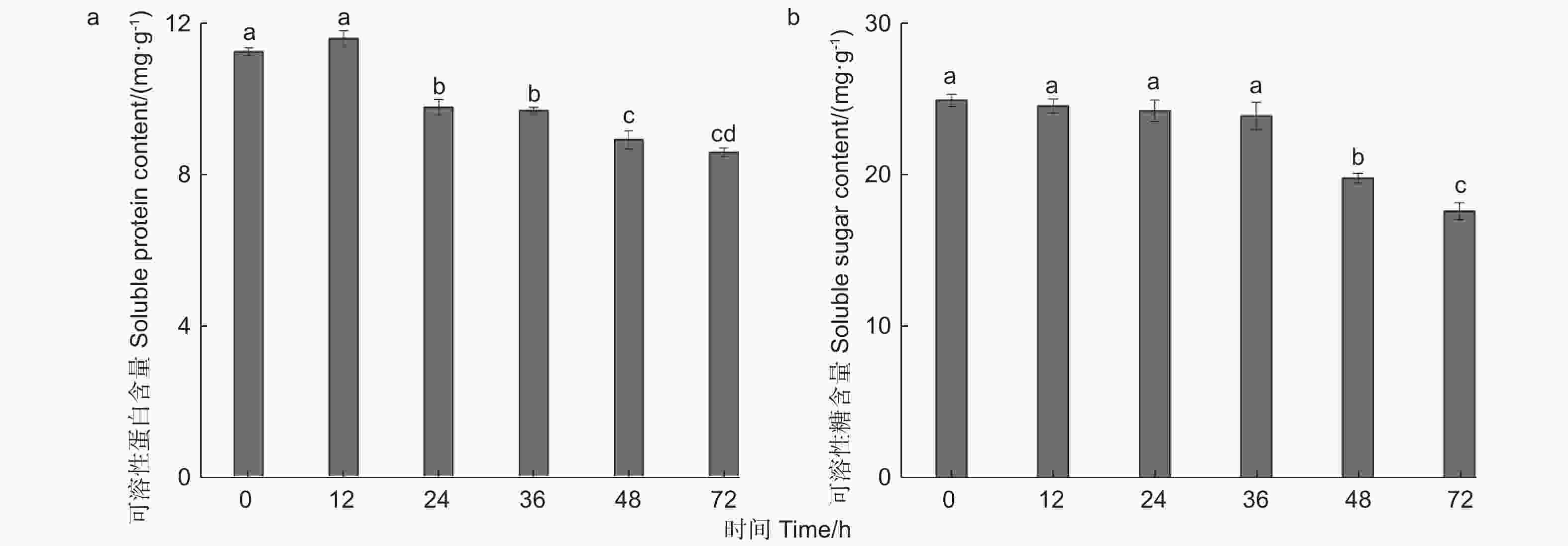
图 2 苹果蠹蛾为害后核桃果皮营养物质含量的变化
Figure 2. Changes of nutrient content in Walnut husk after feeding by Cydia pomonella
-
由图2b可知,苹果蠹蛾为害不同时间段后,核桃果皮可溶性糖逐渐下降。为害12、24、36、48、72 h时可溶性糖含量分别比对照低1.51%、2.78%、4.15%、20.66%、29.42%。其中,在苹果蠹蛾为害12~36 h内与对照相比均无显著差异(P>0.05),在为害48 h时与对照相比差异水平显著(P<0.05)。可见在危害48 h时,可溶性糖含量显著降低,进而可能抵御苹果蠹蛾的为害。
综上所述,核桃果皮营养物质的含量随着苹果蠹蛾为害时间的延长而逐渐降低,由此推测核桃果皮可能通过改变营养物质进而影响苹果蠹蛾的生长发育,其中不同营养物质所改变的时间有所差异。
-
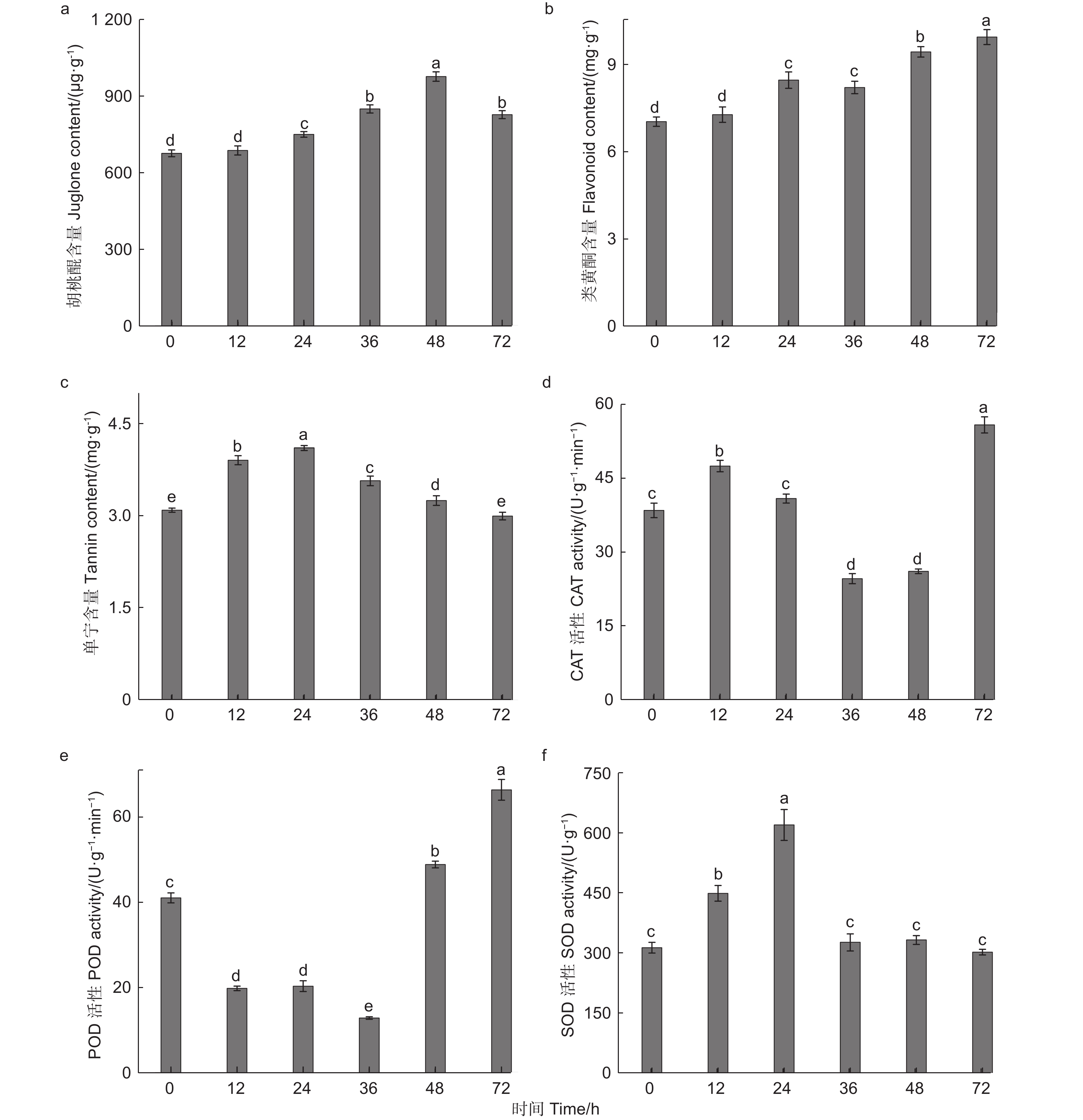
根据图3a可知,随着苹果蠹蛾为害时间的延长,核桃果皮中胡桃醌的含量与对照相比均增加,其含量呈先上升后下降的趋势。在苹果蠹蛾为害24~72 h时与对照相比胡桃醌的含量均有显著差异(P<0.05),其中在为害48 h时胡桃醌含量达到最高,为976.68 µg·g−1,与对照相比增加44.48%。为害36 h和72 h与对照相比胡桃醌含量变化较小,分别增加25.60%和22.32%。由此可见在为害48 h时,苹果蠹蛾幼虫的取食可能受到影响。
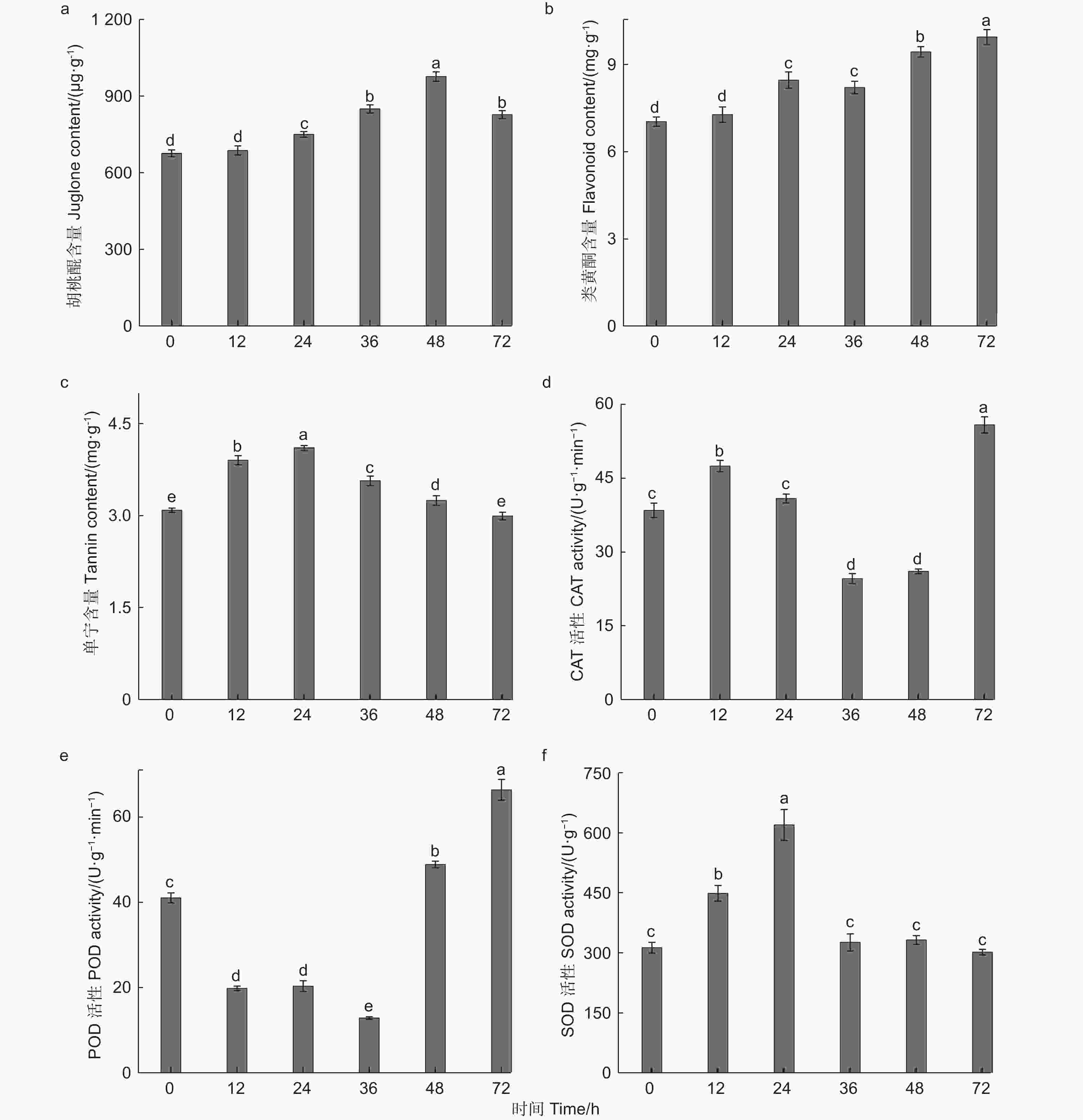
图 3 苹果蠹蛾为害后核桃果皮次生代谢物质含量与防御酶活性的变化
Figure 3. Changes of secondary metabolite content and defense enzyme activities in walnut husk after feeding by Cydia pomonella
-
由图3b可知,苹果蠹蛾为害不同时间,与对照相比每个时间点的类黄酮的含量均增加,其含量的表达趋势逐渐上升。在取食24 h时与对照相比类黄酮含量差异显著(P<0.05),其含量为8.47 mg·g−1,是对照的1.21倍,在为害72 h时类黄酮含量达到最高为9.95 mg·g−1,是对照的1.41倍。由此可知苹果蠹蛾为害不同时间后类黄酮的大量合成,可能是因为类黄酮参与对苹果蠹蛾的防御所导致。
-
由图3c可知,苹果蠹蛾为害时间不同,核桃果皮单宁含量不同,其单宁含量呈先增加后降低的趋势。在为害12 h时,与对照相比差异显著(P<0.05),单宁含量为对照的1.26倍,在为害24 h时,单宁含量达到最高,为4.11 mg·g−1,是对照的1.33倍,进一步说明在为害24 h时,核桃果皮可能已产生抗虫性能。在为害12~48 h内,单宁的含量均显著高于对照(P<0.05),但在为害72 h时,与对照相比单宁含量无显著差异(P>0.05),且含量是对照的97%。
综上所述,苹果蠹蛾为害不同时间点,核桃果皮次生代谢物的含量变化趋势不同,可见核桃果皮中的不同次生代谢物在苹果蠹蛾胁迫时均产生相应的抗性。
-
通过对核桃果皮CAT活性的测定,由图3d可知,随着取食时间的推移CAT活性呈先增加后降低再增加的趋势。由于苹果蠹蛾的取食诱导,核桃果皮CAT的活性迅速增加,在为害12 h时,与对照相比CAT活性差异显著(P<0.05),在为害24 h时,虽然CAT活性增加,但是与对照相比无显著差异(Pc0.05),在为害36~48 h内,与对照相比CAT活性均显著降低(P<0.05),在为害36 h时,CAT活性达最低值(24.65 U·min−1·g−1),在为害72 h时,CAT活性达峰值为55.97 U·min−1·g−1,是对照的1.45倍,差异水平达到显著(P<0.05)。
-
根据图3e所示,苹果蠹蛾为害不同时间后,核桃果皮中POD的活性具有显著影响。在为害0~36 h内迅速降低,在为害36 h时,POD活性降到最低值为12.89 U·min−1·g−1,差异水平显著(P<0.05),在为害48~72 h内,POD活性迅速增加,均高于对照,与对照相比均差异显著(P<0.05),在为害72 h时,POD活性达到峰值为66.44 U·min−1·g−1,是对照的1.62倍。
-
通过对核桃果皮中SOD活性的测定,根据图3f可知,核桃果皮应对苹果蠹蛾不同时间的取食胁迫时,体内的SOD活性迅速提升,在为害24 h时达到峰值623.69 U·g−1,是对照的1.98倍,之后SOD的活性有所回落,在取食36 h(329.22 U·g−1)和48 h(335.15 U·g−1)时,核桃果皮中SOD活性虽高于对照,但与对照相比均无显著差异(P>0.05),在取食72 h时,核桃果皮中SOD活性低于对照,但与对照相比SOD活性无显著差异(P>0.05)。结果表明,苹果蠹蛾为害核桃果皮时,核桃果皮防御酶会迅速应对,由此可见核桃果皮对苹果蠹蛾的为害产生明显的诱导防御反应。
-
对于植食性昆虫而言,取食是唯一获取生长发育所需营养的必须途径,因此植物在面对植食性昆虫损害与入侵的时候,可通过对营养物质含量的调节,进而影响植食性昆虫的生长发育与繁殖,降低了昆虫的存活率而获得抗虫性[20-22]。在李传明[22]等人的研究中发现辣椒(Capsicum annuum Linnaeus)被烟粉虱(Bemisia tabaci Gennadius)为害时自身的可溶性糖含量增加,可溶性蛋白含量降低,进而对烟粉虱产生抗性;李胜[23]等人研究表明在颈盲蝽(Pachypeltis Sp)为害不同时间后,薇甘菊(Mikania micrantha Kunth)可溶性糖含量相比对照均显著上升,可溶性蛋白含量逐渐下降。本研究发现,苹果蠹蛾为害后,可溶性糖含量与可溶性蛋白含量均降低,其中可溶蛋白含量在为害24 h是与对照相比差异显著,可溶性糖含量与对照相比在为害48 h时差异显著。但是对于这些营养物质含量与抗虫的相关性,目前无绝对定论[24]。也有研究表明一些刺吸式的昆虫如蝽类,此昆虫的唾液酶中包含一些淀粉酶、蛋白酶等,有助于将食物进行体外消化,将淀粉转化为麦芽糖最终分解为葡萄糖[25]。因此对于苹果蠹蛾为害后核桃果皮中可溶性糖与可溶性蛋白含量的变化是植物自身调节还是由于某些物质的分解所导致,有待进一步研究。
在植物体内含有一类结构复杂、种类较多的次生代谢物质,这类物质不能直接参与植物的生长发育,但是在植食性昆虫为害时,可有效抑制、干扰昆虫的一些交配、取食行为,同时也可以吸引天敌,进而诱杀。因此在植物防御植食性昆虫为害时发挥重要作用,也是衡量植物抗性的重要指标[26-27]。在前人的研究中发现刚竹毒蛾(Pantana phyllostachysae Chao)为害毛竹(Phyllostachys edulis Houz)叶片时,单宁与类黄酮的含量均显著上升;马尾松毛虫(Dendrolimus punctatus Walker)为害马尾松(Pinus massoniana Lamb)后,其叶片中的单宁含量显著升高;当甜槠(Castanopsis eyrei Totch)被红腹柄天牛(Aphredisinm faldermannii rufiventris Gressitt)为害后,其植物体内的类黄酮含量显著升高[28-30]。本研究发现在苹果蠹蛾为害不同时间后单宁含量在12~48 h均显著高于对照,其中在为害24 h时单宁含量达到峰值,由此可以推断此时间点核桃果皮可能产生抗性,抵御苹果蠹蛾的为害;为害不同时间类黄酮的含量均持续升高并差异显著,说明苹果蠹蛾为害可诱导核桃果皮中类黄酮含量的升高,进而提高核桃果皮对苹果蠹蛾的抗性。胡桃醌是核桃果皮中主要的次生代谢物之一,其具有抗氧化、抑菌、杀虫的作用[31]。王雨萌[32]等人研究表明胡桃醌对小菜蛾(Plutella xylostella Linnaeus)、蚜虫具有较好的触杀作用,对白粉虱(Trialeurodes vaporariorum Westwood)成虫具有胃毒作用。Piskorski[33]等人研究表明当苹果蠹蛾取食含有胡桃醌的饲料时与对照相比,幼虫发育历期延长,体重减少,当胡桃醌的含量是核桃果皮的两倍时对幼虫有毒杀的作用。在本研究中,苹果蠹蛾取食核桃果皮,因此胡桃醌含量的高低与核桃果皮对苹果蠹蛾的抗性有关,故胡桃醌的含量可以作为评价核桃种或品种抗苹果蠹蛾的指标之一。可见在苹果蠹蛾为害不同时间后,次生代谢物含量均有变化,其在24~48 h之间,从单宁、类黄酮和胡桃醌含量的变化趋势推测这3种物质可能相互协同,因此本研究从物质含量变化的角度揭示核桃果皮受虫害胁迫后其防御响应的机制,但对于更深层次的防御机制需进一步的研究。
植物与病虫害对抗时,防御酶起着非常重要的作用,能作用昆虫使昆虫中毒或者营养失衡[34]。其中CAT、POD、SOD等防御酶是植物在逆境胁迫下最为关键的酶[35]。CAT的主要功能是将植物体内的H2O2分解,维持植物相关组织活性;POD与植物体内活性氧代谢有关,主要维持植物体内自由基平衡,同时参与木质素合成,在植物抗病虫害中起着重要作用;SOD的主要功能是植物在逆境条件下清除自由基,延缓植物衰老[36]。陈鹏[37]等人研究表明当二斑叶螨(Tetranychus cinnabarinus Koch)为害草莓(Fragaria ananassa Duch)叶片不同时间时,SOD活性呈先上升后降低的趋势;母小庆[38]等人研究梨小食心虫(Grapholita molesta Busck)与苹果蠹蛾对苹果果实防御酶活性的影响中表明,在遭受害虫为害后,SOD活性随时间的推移而呈上升趋势;李胜[23]等人研究表明颈盲蝽为害薇甘菊后,随着时间的推移,POD与CAT活性呈先上升后下降的趋势,而SOD呈先降低后上升再降低的趋势。可见寄主植物与植食性昆虫不同,防御酶活性趋势也有较大差异。本研究发现苹果蠹蛾为害核桃果皮后,SOD活性先上升,在为害24 h时达到峰值,之后活性逐渐降低;CAT活性呈先上升后降低再上升的趋势;POD活性呈先降低后升高的趋势,CAT与POD均在在为害72 h是达到峰值。由此可见苹果蠹蛾为害胁迫会引起核桃果皮酶促反应中相关酶活性的变化,进而抵御苹果蠹蛾的为害。从防御酶活性变化趋势推测SOD、CAT与POD可能存在拮抗作用,但对于防御酶相互作用的分子机制需进一步研究。
近年来农业产业发展结构的不断调整与果园管理模式的改变,苹果蠹蛾已在我国各大果区严重发生,尤其在核桃园区中扩散面积呈上升趋势。因此本研究明确苹果蠹蛾为害后,可诱导核桃果皮营养物质与次生代谢物质的含量、防御酶活性的变化,进而从物质含量变化的角度揭示核桃果皮受苹果蠹蛾为害后其防御的响应机制。由于植物自身在应对生物与非生物胁迫时,相关防御体系、代谢途径形式多样化,因此在今后的研究中从微观代谢途径的变化进一步研究,探究植物应对虫害胁迫时所产生的防御机制,为苹果蠹蛾的绿色防控奠定基础。
-
本研究发现苹果蠹蛾为害诱导核桃果皮在营养物质、次生代谢物质以及防御酶活性方面产生防御响应。其中单宁含量与CAT、POD、SOD活性在苹果蠹蛾为害12 h时较对照差异显著(P<0.05);可溶性蛋白、胡桃醌以及类黄酮含量在苹果蠹蛾为害24 h时较对照差异显著(P<0.05);可溶性糖含量在苹果蠹蛾为害48 h时较对照差异显著(P<0.05)。为之后核桃抗虫品种的选育提供科学的理论依据。
核桃果皮对苹果蠹蛾为害的防御响应
Defensive Response of Walnut Husk to Cydia Pomonella
-
摘要:
目的 研究苹果蠹蛾为害后诱导核桃果皮产生的防御应答响应。 方法 采用紫外分光光度法与酶标仪微量法分析核桃果皮营养物质、次生代谢物质的含量,防御酶的活性以及苹果蠹蛾与核桃的互作关系。 结果 核桃果皮被苹果蠹蛾蛀食后可溶性糖与可溶性蛋白的含量随着时间推移逐渐降低,在为害24 h时可溶性蛋白含量与对照相比差异显著(P<0.05);在为害48 h时可溶性糖与对照相比差异显著(P<0.05)。次生代谢物质胡桃醌、单宁的含量随时间的推移呈先上升后下降的趋势,其中胡桃醌在为害48 h时含量达到最高,为976.68 µg·g−1,是对照的1.44倍;单宁含量在24 h时达到高峰,为4.11 mg·g−1,是对照的1.33倍;类黄酮的含量呈逐渐上升趋势,在24 h时与对照相比差异显著(P<0.05)。在为害12 h时,CAT活性与对照相比差异显著(P<0.05),在72 h时CAT活性达峰值,为55.97 U·min−1·g−1,是对照的1.45倍;POD活性呈先下降后上升的趋势,在72 h时达最高值,是对照的1.62倍;SOD活性呈先上升后下降的趋势,在24 h时达到峰值,为623.69 U·g−1,是对照的1.98倍。 结论 核桃果皮主要通过调节体内的营养物质、次生代谢物质以及防御酶活性的变化,对苹果蠹蛾的为害产生应激反应,进而发挥防御作用。 Abstract:Objective To Investigate the defensive response to walnut husk after Cydia pomonella infestation. Method UV Spectrophotometry and Microplate Reader were used to analyze nutrient substances, secondary metabolites content, defense enzyme activity and interaction between Cydia pomonella and walnut Result The content of soluble sugar and soluble protein decreased gradually over time after the walnut husk was eaten by C. pomonella. The soluble protein content at 24 h of damage was significantly different from the control (P<0.05); There was a significant difference between the soluble sugar and the control at 48 hours of damage (P<0.05). The contents of secondary metabolites juglone and tannin increased first and then decreased. Among them, the content of juglone quinone reached the highest at 48 h, which was 976.68 µg·g−1 FW, and was 1.44 times that of the control; The tannin content reached a peak at 24 h at 4.11 mg·g−1, which was 1.33 times that of the control; The content of flavonoids showed a gradual upward trend, and the difference was significant compared with the control at 24 h (P<0.05). At 12 h of damage, there was a significant difference in CAT activity compared with the control (P<0.05). At 72 h, the CAT activity peaked at 55.97 U·min−1·g−1, which was 1.45 times that of the control; The POD activity showed a trend of first decreasing and then increasing. At 72 h, the POD activity reached the highest value, which was 1.62 times that of the control; The SOD activity increased firts and then decreased. At 24 h, the SOD activity reached a peak value of 623.69 U·g−1, which was 1.98 times that of the control. Conclusion The walnut husk mainly produces a stress response to the damage of C. pomonella by regulating the changes of nutrients, secondary metabolites and defense enzyme activities in the body, and then plays a defensive role. -
Key words:
- Cydia pomonella
- / walnut husk
- / nutrients
- / metabolites
- / defense enzyme activity
-
图 2 苹果蠹蛾为害后核桃果皮营养物质含量的变化
Figure 2. Changes of nutrient content in Walnut husk after feeding by Cydia pomonella
-
[1] Food and Agriculture Organization of the United Nations[DB/OL]. [2022-4-18]https://www.fao.org/home/en. [2] KADOIĆ BALAŠKO M, BAŽOK R, MIKAC K M, et al. Pest management challenges and control practices in codling moth: a review[J]. Insects, 2020, 11(1): 38. doi: 10.3390/insects11010038 [3] CHEN C Y, MAO Y B. Research advances in plant–insect molecular interaction[J]. F 1000 Research, 2020, 9(198): 1-11. [4] 高微微, 佟建明, 郭顺星. 植物次生代谢产物的生态学功能研究进展[J]. 中国药学杂志, 2006(13):961-964. doi: 10.3321/j.issn:1001-2494.2006.13.001 [5] 孙墨珑, 宋湛谦, 方桂珍. 核桃楸叶乙醇提取物杀虫活性及活性成分[J]. 应用生态学报, 2007(12):2910-2914. [6] 王国迪, 赵 丽, 尉吉乾, 等. 植物次生代谢产物对烟粉虱影响的研究进展[J]. 河南科技学院学报(自然科学版), 2018, 46(3):8-14. [7] HOWE G A, JANDER G. Plant immunity to insect herbivores[J]. Annual Review of Plant Biology, 2008, 59(1): 41-66. doi: 10.1146/annurev.arplant.59.032607.092825 [8] VISSER J H. Differential sensory perceptions of plant compounds by insects[J]. Plant Resistance to Insects, 1983, 208(208): 215-230. [9] 黄炜东. 两色绿刺蛾为害对毛竹营养物质的影响[J]. 竹子研究汇刊, 2006(4):24-27. [10] 杨金睿, 肖关丽. 植物抗虫生理研究进展[J]. 中国农学通报, 2021, 37(6):130-136. [11] 冯建雄, 董晓亮, 杨博慧, 等. 油菜叶片营养物质含量和防御酶活性与其对黄宽条跳甲抗性的关系[J]. 植物保护, 2019, 45(3):49-54. [12] KESSLER A, BALDWIN I T. Plant responses to insect herbivory: the emerging molecular analysis[J]. Annual Review of Plant Biology, 2002, 53(1): 299-328. doi: 10.1146/annurev.arplant.53.100301.135207 [13] TSCHARNTKE T, VIDAL S, HAWKINS B A. Parasitoids of grass-feeding chalcid wasps: a comparison of German and British communities[J]. Oecologia, 2001, 129(3): 445-451. doi: 10.1007/s004420100733 [14] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000. [15] BRADFORD M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry, 1976, 72(1-2): 248-254. doi: 10.1016/0003-2697(76)90527-3 [16] 侯 栋, 高 哲, 何童森, 等. HPLC测定核桃青皮中胡桃醌的含量[J]. 中国实验方剂学杂志, 2012, 18(15):70-72. doi: 10.3969/j.issn.1005-9903.2012.15.019 [17] 李忠光, 龚 明. 愈创木酚法测定植物过氧化物酶活性的改进[J]. 植物生理学通讯, 2008(2):323-324. [18] 彭 建, 王丹英, 徐春梅, 等. 钼酸铵法测定水稻过氧化氢酶活性[J]. 中国农学通报, 2009, 25(16):61-64. [19] 徐 东, 赵 建, 黄汉昌, 等. 改良的黄嘌呤氧化酶法测定动植物组织中SOD比活力[J]. 食品科学, 2011, 32(6):237-241. [20] BAGHBAN A, SENDI J J, KHOSRAVI R, et al. Effect of heavy metals (Cd, Cu, and Zn) on feeding indices and energy reserves of the cotton boll worm Helicoverpa armigera Hübner (Lepidoptera: Noctuidae)[J]. Journal of Plant Protection Research, 2014, 54(4): 367-373. [21] SOLANKI R, DHANKHAR R. Zinc and copper induced changes in physiological characteristics of Vigna mungo (L. )[J]. Journal of Environmental Biology, 2011, 32(6): 747. [22] 李传明, 何 菁, 顾爱祥, 等. 烟粉虱取食对不同抗虫性辣椒品种营养物质和抗性物质的影响[J]. 中国生态农业学报, 2017, 25(10):1456-1462. [23] 李 胜, 郑端靖, 王 尹, 等. 颈盲蝽取食对薇甘菊叶片营养物质和防御酶的影响[J]. 生物安全学报, 2018, 27(1):45-49. doi: 10.3969/j.issn.2095-1787.2018.01.007 [24] 王紫薇, 徐华潮, 汪云珍, 等. 树皮内含物对光肩星天牛取食与刻槽产卵量的影响[J]. 环境昆虫学报, 2016, 38(5):942-949. [25] AZEVEDO D O, ZANUNCIO J C, ZANUNCIO JR J S, et al. Biochemical and morphological aspects of salivary glands of the predator Brontocoris tabidus (Heteroptera: Pentatomidae)[J]. Brazilian Archives of Biology and Technology, 2007, 50: 469-477. doi: 10.1590/S1516-89132007000300013 [26] DUČAIOVÁ Z, SAJKO M, MIHALIČOVÁ S, et al. Dynamics of accumulation of coumarin-related compounds in leaves of Matricaria chamomilla after methyl jasmonate elicitation[J]. Plant Growth Regulation, 2016, 79(1): 81-94. doi: 10.1007/s10725-015-0114-2 [27] SHAHABINEJAD M, SHOJAADDINI M, MASRETI B, et al. Exogenous application of methyl jasmonate and salicylic acid increases antioxidant activity in the leaves of pistachio (Pistacia vera L. ) trees and reduces the performance of the phloem-feeding psyllid Agonoscena pistaciae[J]. Arthropod-Plant Interactions, 2014, 8(6): 525-530. doi: 10.1007/s11829-014-9334-z [28] 张潮巨. 毛竹受害后化学物质含量的变化及其对刚竹毒蛾种群参数的影响[J]. 华东昆虫学报, 2006(4):281-283. [29] 任琴. 马尾松快速诱导抗性及化学信号物质的研究[D]. 北京: 北京林业大学, 2006. [30] 刘鹏程, 陈顺立, 童应华, 等. 红腹柄天牛为害对甜槠黄酮、单宁含量的影响[J]. 福建林学院学报, 2006(4):314-317. doi: 10.3969/j.issn.1001-389X.2006.04.006 [31] 张宏露, 罗兴平. 核桃青皮主要成分胡桃醌的生物活性研究进展[J]. 当代化工研究, 2021(9):164-165. [32] 王雨萌. 植物源杀虫活性成分分离鉴定及杀虫活性研究[D]. 长春: 长春师范大学, 2015. [33] PISKORSKI R, DORN S. How the oligophage codling moth Cydia pomonella survives on walnut despite its secondary metabolite juglone[J]. Journal of Insect, 2011, 57(6): 744-750. [34] 李旭颖. 植食性昆虫与寄主植物之间的相互关系和影响[J]. 科技创新导报, 2011(11):130. [35] 郭祖国, 王梦馨, 崔 林, 等. 6种防御酶调控植物体应答虫害胁迫机制的研究进展[J]. 应用生态学报, 2018, 29(12):4248-4258. [36] 陈 磊, 王宇飞, 李海平, 等. 荞麦叶片生化物质含量和防御酶活性与其对西伯利亚龟象抗性的关系[J]. 应用昆虫学报, 2022, 59(4):888-899. doi: 10.7679/j.issn.2095-1353.2022.093 [37] 陈 鹏, 刘奇志. 二斑叶螨为害对草莓叶片H2O2、MDA含量以及部分防御酶活性的影响[J]. 环境昆虫学报, 2022, 44(3):697-703. doi: 10.3969/j.issn.1674-0858.2022.03.21 [38] 母小庆, 林 萍, 李 贞, 等. 梨小食心虫与苹果蠹蛾为害对苹果果实防御酶活性的影响[J]. 应用昆虫学报, 2020, 57(2):348-354. -

 点击查看大图
点击查看大图
图(3)
计量
- 文章访问数: 1546
- HTML全文浏览量: 828
- PDF下载量: 34
- 被引次数: 0
