



-
作为一种典型的土壤习居丝状真菌,淡紫紫孢菌(Purpureocillium lilacinum (Thom) Luangsa-ard, Houbraken, Hywel-Jones & Samson)是最具潜力的植物寄生线虫生防真菌之一[1-2],但它也同其它生防真菌制剂一样,存在着受环境影响较大、防治效果不稳定以及自身抗逆性较差等缺点[3-4]。目前无论是研究通过对生防菌株关键基因进行改造,达到提升生防菌制剂致病性和环境稳定性的目的[5],还是研究生防菌对病原物的致病机制及它们之间的相互作用机制,都需要建立高效的遗传转化系统。
农杆菌(Agrobacterium)介导的真菌转化已经在许多丝状真菌中得到广泛应用[6-15],农杆菌介导的生防真菌转化也已有不少成功的实例,例如已成功实现了昆虫病原真菌白僵菌(Beauveria bassiana ) [10] 、绿僵菌(Metarhizium anisopliae) [12]和蜡蚧轮枝菌(Lecanicillium lecanii)[15] 等生防真菌的遗传转化。真菌菌种的不同可以导致遗传转化效率不同 [16],另外转化载体和农杆菌菌株也可以影响遗传转化效率。G418作为筛选标记建立的遗传转化体系达到了1 000~2 400个转化子/106分生孢子,遗传转化效率和抗性转化子比率偏低[17]。基于此,针对不同真菌的遗传转化体系其具体参数必须进行优化。 beta-tubulin基因是多菌灵的抗性基因,淡紫紫孢菌对多菌灵高度敏感,可用于抗性筛选。
本文在前期研究的基础上,以beta-tubulin基因为筛选标记,对影响转化体系的一些参数,如诱导培养乙酰丁香酮(AS)的浓度、诱导培养时间、农杆菌终浓度 OD660值、共培养 AS 的浓度、共培养时间和共培养温度等因子进行优化,以期建立了农杆菌介导淡紫紫孢菌高效遗传转化体系,并从中突变体库中筛选鉴定出致病力变异的突变体,从而为淡紫紫孢菌的致病机制研究及优良菌株选育奠定基础。
-
从中国林业微生物菌种保藏管理中心(CFCC)获得了强致病性菌株淡紫紫孢菌20-7和根癌农杆菌(A. tumefaciens)菌株EHA105及AGL-1。原始质粒pBI-G3C由日本京都府立大学Yasuyuki Kubo提供,带有苯菌灵的抗性基因beta-tubulin的质粒pCPXBN-1由日本宫城大学Skin Kasahara提供,该基因被克隆后用来置换pBI-G3C中的hph(Apa I/Cla I)基因。
LB培养基、基本培养基(minimal medium,MM)和诱导培养基(induction medium,IM)分别用于农杆菌划线培养、振荡培养和共培养前诱导[17]。PDA培养基用于转化子的继代筛选和转化子保存。将混和后的农杆菌和淡紫紫孢菌孢子铺在共培养培养基(0.8 mL 1.25 mol·L−1K-buffer(pH4.9),20 mL M-N,1 mL 10 mg·L−1 CaCl2·2H2O,5 mL Spore Elements,2.5 mL 200 mg·L−1NH4NO3,10 mL 500 mg·L−1甘油(glycerol),40 mL 1 mol·L−1MES(pH 5.5 用 NaOH 调节 pH 值),10 mL 200 mg·L−1葡萄糖(glucose),2 mL 100 μg·mL−1AS,加蒸馏水定容至1 L)上进行转化。链霉素(浓度 50 µg·mL−1)和卡那霉素(浓度10 µg·mL−1)用于振荡培养农杆菌;头孢噻肟钠(浓度200 µg·mL−1)和苯菌灵(浓度300 µg·mL−1)用于转化子筛选。
-
根据设计的保守引物,采用PCR方法从pCPXBN-1载体中扩增 beta-tubulin基因,胶回收后T载体过夜连接,连接产物转化大肠杆菌DH5α感受态细胞,涂在附加60 µg·mL−1氨苄青霉素(amp)的LB固体平板上筛选重组菌株。重组菌株质粒DNA碱法小量提取的步骤见文献[18]。
-
在beta-tubulin引物端对Apa I和Cla I酶切位点进行双酶切,用T4 DNA Ligase连接回收酶切后的DNA片段。连接产物转化大肠杆菌DH5α感受态细胞,构成表达载体pBI-G3C-BN。PCR检测转化后的重组子。重组菌株质粒DNA碱法小量提取的步骤同上。
-
为了确定农杆菌介导淡紫紫孢菌遗传转化体系的最佳条件,本体系是在已建立的转化淡紫紫孢菌方案[17]基础上修改而成。
-
为了评估淡紫紫孢菌对苯菌灵的敏感性,淡紫紫孢菌20-7菌株生长在PDA培养基上,该培养基添加了不同浓度的苯菌灵(0、100、200、300、400、500和1 000 µg·mL−1)。在25 ℃下培养3个独立的培养物。分别于培养后48、72、96和108 h观察菌落生长情况,经5次继代培养,筛选抗性稳定的菌株转接到PDA培养基(苯菌灵 50 µg·mL−1)斜面上保存。
-
为了确定最佳共培养条件,根据20-7菌株的不同生长时期(4 ℃ 保藏30 d和新鲜菌株)、农杆菌菌株转化效率(EHA105和AGL-1)、农杆菌悬浮液浓度(OD660值为0.1、0.2、0.3、0.4、0.5、0.6 和 0.7)、乙酰丁香酮浓度(100,200,500和1 000 µg·mL−1)、20-7菌株与农杆菌菌株的共培养时间(24、48、72 和 96 h)、培养温度(22、25、28和31 ℃)和pH(5.0、5.5 和 6.0)来进行筛选,每种条件设置3个重复。采用两种共培养方式:A. 等体积混合EHA105菌液和20-7孢子悬浮液(106个·mL−1),于28 ℃摇床振荡(140 rmp)培养14 h后,将混合物涂布在含有200 µg·mL−1的头孢噻肟钠和300 µg·mL−1的苯菌灵PDA培养基上,用于筛选转化子。置于25 ℃培养箱中培养3~5 d;B. 将等体积混合的EHA105菌液和20-7孢子悬浮液(106个·mL−1)涂布在共培养平板上的无菌滤纸条上,在25 ℃下避光培养72 h。揭下滤纸条,反铺到PDA培养基(含200 µg·mL−1头孢噻肟钠和300 µg·mL−1苯菌灵)上,25 ℃ 培养48 h。对每个平板中的真菌菌落进行计数和统计分析。
-
挑取抗性单菌落,至PDA平板(含200 µg·mL−1头孢噻肟钠和300 µg·mL−1苯菌灵)上,25 ℃下恒温培养,再进行筛选。7 d后,单孢分离有孢子产生的转化子,单孢转接到PDA(含苯菌灵 300 µg·mL−1和头孢噻肟钠 200 µg·mL−1)培养基上进行筛选;具体筛选步骤参考王曦茁等 [17],经5次筛选的抗性菌株转接到PDA斜面培养基(含300 µg·mL−1苯菌灵)上保存。
-
转化子总DNA的提取及农杆菌Ti质粒DNA的碱法小量提取步骤参照《植物基因工程原理与技术》[18]。随机挑选100个转化子进行PCR验证。引物序列为Tub-FWD(GGGCCCTCGAGAGGGGGCCTTCCACCCTTC)和Tub-REV(ATCGATTAGAGGATCCCCGGGGGATCC--AGA),其中下划线表示Apa I和Cla I酶切位点。扩增反应程序参考王曦茁等文献[17],根据扩增反应结果来确定T-DNA是否插入到淡紫紫孢菌基因组内。
-
取PCR阳性的转化子参考《分子克隆实验指南》第三版的方法[19]提取基因组DNA,用Hind III酶切,37 ℃过夜,pCPXBN-1质粒DNA用Hind III酶切作为阳性对照。酶切后的DNA在0.8%的琼脂糖凝胶上进行电泳分离。DNA印迹转膜、洗涤、预杂交、洗膜、显影以及探针标记等参考文献[20]。
-
随机选取20株转化子菌丝到含有苯菌灵的PDA培养基上,25 ℃下培养7 d后,再连续转接5次。观察和记录转化子的菌落形态和生长状况等表型特征,分析确定其遗传稳定性。
-
按照常规方法获得南方根结线虫(M. incognita)虫卵[21]。在直径6 cm的培养皿中加入300 µL卵悬液(1 000 个·mL−1),然后分别加入3 mL的20个随机选取转化子的孢子悬浮液(1.0 × 106 个·mL−1),并以原始菌株20-7作为对照。4 d后统计卵的寄生和未寄生数目,计算转化子对南方根结线虫虫卵的寄生率。
-
构建质粒pBI-G3C-BN的示意图见图1,将原有质粒pBI-G3C中的hph基因置换为beta-tubulin基因,beta-tubulin基因从质粒pCPXBN-1上扩增获得。

图 1 载体构建示意图
Figure 1. Schematic diagram of carrier construction
-
淡紫紫孢菌菌株20-7菌落生长与PDA培养基中苯菌灵浓度密切相关,浓度越大,生长越受抑制。当苯菌灵浓度达到300 µg·mL−1以上时,完全抑制菌落生长(表1)。
表 1 苯菌灵浓度对淡紫紫孢菌20-7菌落生长的影响
Table 1. Effects of different concentrations of benomyl on the growth of P. lilacinum 20-7
浓度
Concentration/
(µg·mL−1)20-7菌落直径/cm
The colony diameter of P. lilacinum /cm48 h 72 h 96 h 108 h 0 2.11 5.13 7.57 9.12 100 0.34 0.83 1.46 1.94 200 0.01 0.03 0.04 0.06 300 0 0 0 0 400 0 0 0 0 500 0 0 0 0 1 000 0 0 0 0 -
淡紫紫孢菌孢子新鲜程度不同,对其遗传转化率有所影响。当AS浓度200 µg·mL−1,苯菌灵的筛选浓度为300 µg·mL−1时,新鲜孢子的转化率比保存一个月后的孢子转化率约增长15倍(表2)。
表 2 淡紫紫孢菌20-7菌株孢子新鲜程度对转化效率的影响
Table 2. Effects of different degree of spore freshness on transformation efficiency of P. lilacinum 20-7
孢子新鲜程度
Spores freshness转化子个数
Transformants /(×10−6)保存一个月的孢子
Spores saved for a month187 新鲜孢子
Fresh spores3 019 -
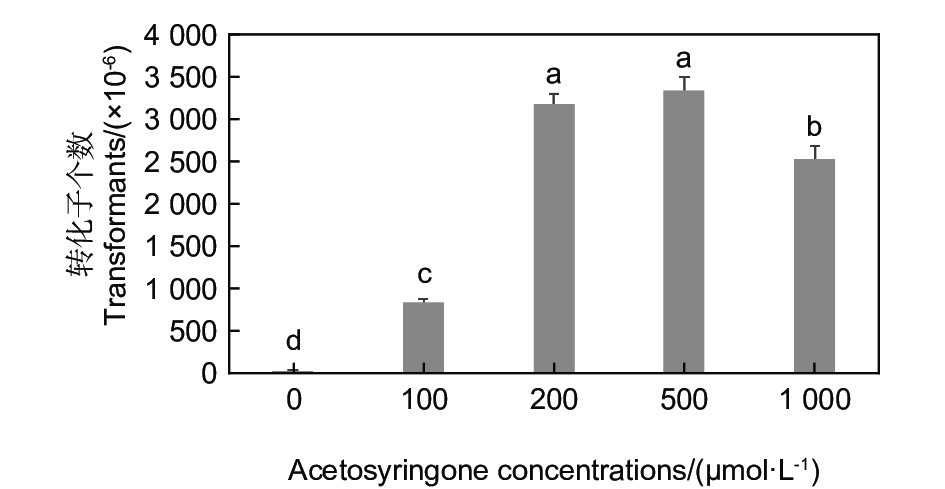
乙酰丁香酮AS浓度对淡紫紫孢菌转化效率影响明显,当AS浓度为200 µg·mL−1到1 000 µg·mL−1时,转化效率均较高(图2)。从节约成本出发,选用AS浓度在200 µg·mL−1到500 µg·mL−1为宜。

图 2 乙酰丁香酮浓度对淡紫紫孢菌20-7转化效率的影响
Figure 2. Effects of different acetosyringone concentrations on the transformation efficiency of P. lilacinum 20-7
-
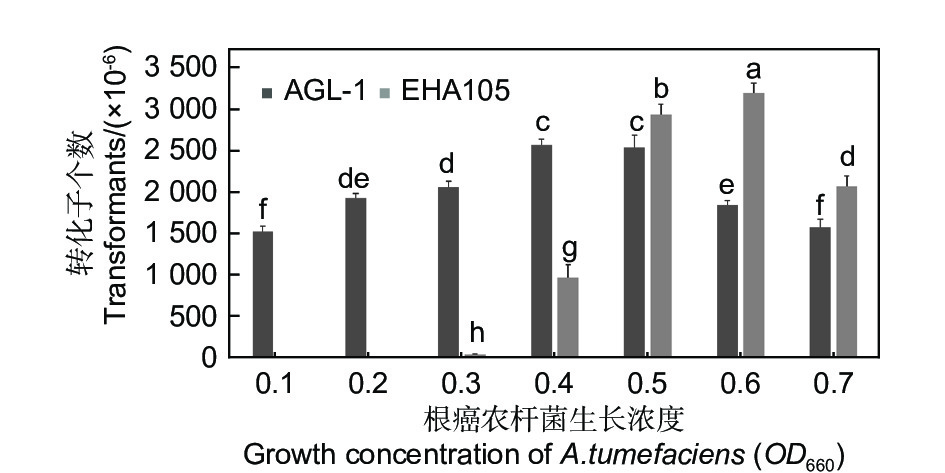
农杆菌菌株EHA105和AGL-1在相同的试验条件下,对淡紫紫孢菌20-7菌株进行遗传转化的效率存在较大差异。农杆菌菌株AGL-1的所有生长浓度(OD660)均能对淡紫紫孢菌进行有效转化,而农杆菌菌株EHA105的生长浓度(OD660)在0.4以上时,遗传转化率较高。因此,农杆菌菌株及浓度的不同均能影响淡紫紫孢菌遗传转化率。EHA105生长浓度0.6时,转化效率最高,明显高于AGL-1的转化率(图3)。

图 3 农杆菌菌株浓度对淡紫紫孢菌20-7转化率的影响
Figure 3. Effects of different Agrobacterium strains on the conversion rate of P. lilacinum 20-7
-
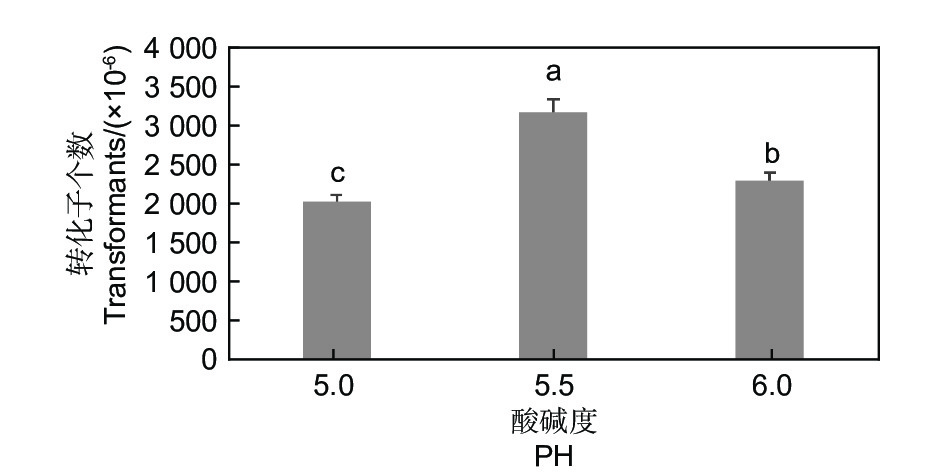
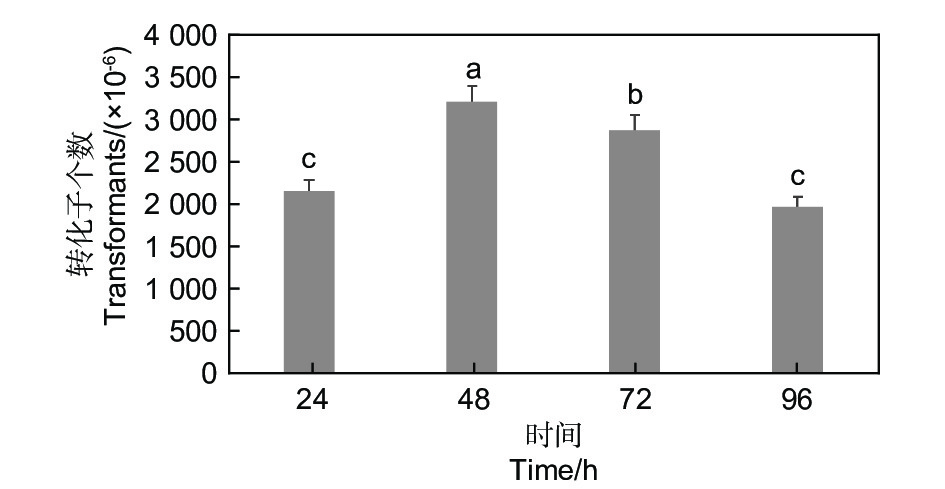
在相同的试验条件下,共培养培养基的酸碱度、共培养时间和温度都对转化效率有影响。酸碱度、时间和温度分别为pH5.5、48 h和25 ℃时转化效率最高(图4、图5和图6)。

图 4 pH对淡紫紫孢菌20-7转化效率的影响
Figure 4. Effect of pH on transformation efficiency of P. lilacinum 20-7

图 5 时间对淡紫紫孢菌20-7转化效率的影响
Figure 5. Effect of time on transformation efficiency of P. lilacinum 20-7

图 6 温度对淡紫紫孢菌20-7菌株转化效率的影响
Figure 6. Effect of temperature on transformation efficiency of P. lilacinum 20-7
-
不同的共培养方式,转化效率不同, EHA105菌液和20-7孢子悬浮液混合振荡培养的转化效率约是混合液直接涂在滤纸上转化效率的1.6倍(表3)。
表 3 共培养方法对淡紫紫孢菌20-7转化效率的影响
Table 3. Effects of different co-culture methods on transformation efficiency of P. lilacinum 20-7
共培养方法
Co-culture method转化子个数
Transformants/ (×10−6)振荡培养 Shake the medium 3 208 涂于滤纸片上 Coat on filter paper 1 948 -
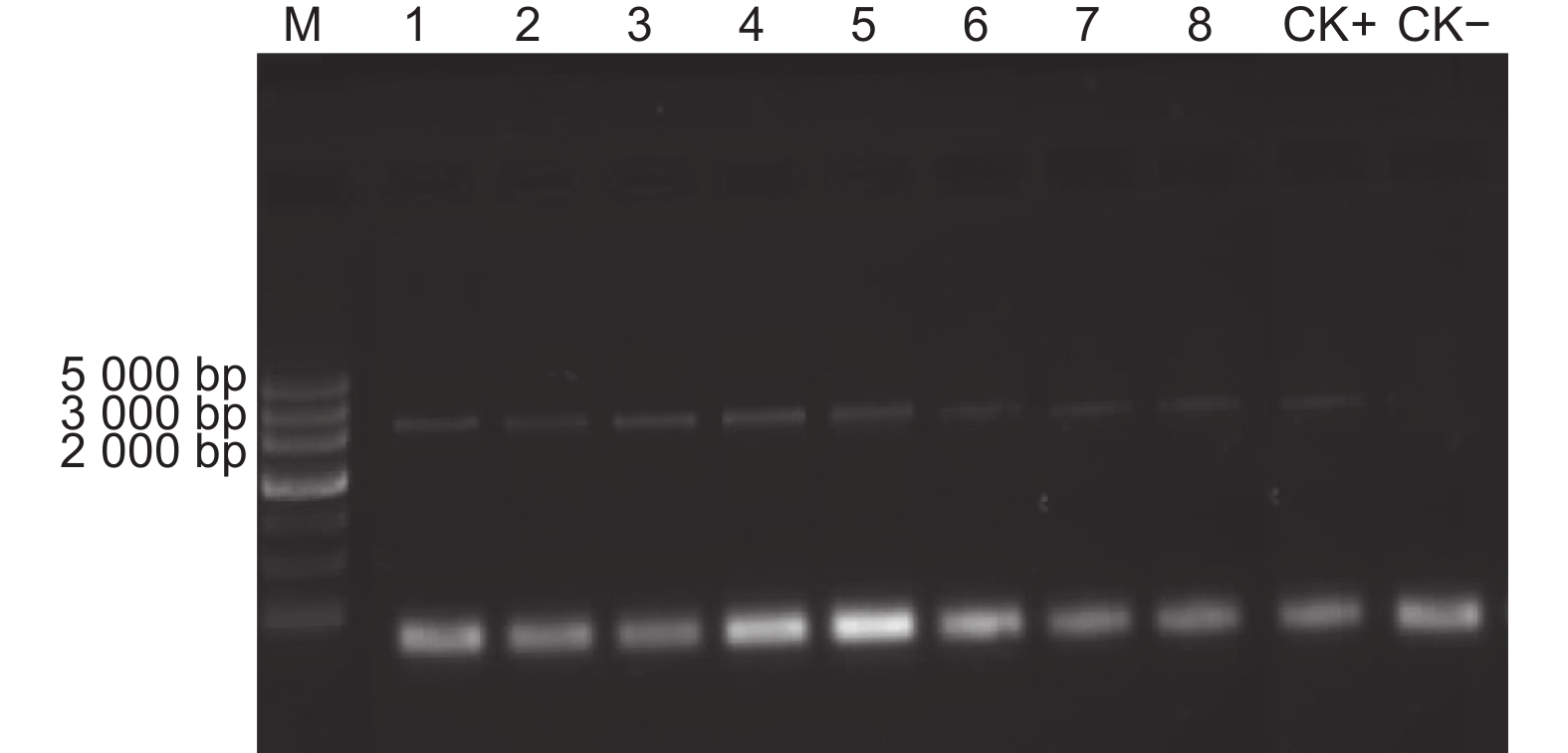
提取随机选择100个的转化子、质粒pBI-G3C-BN和原始菌株基因组DNA,并将质粒pBI-G3C-BN和原始菌株基因组DNA用作PCR研究的参考。使用beta-tubulin基因特异性引物进行扩增,从96个转化子和阳性对照中获得了约2.2 kb的序列,证明苯菌灵基因已整合到20-7转化子的基因组中;而同时观察到4个转化子没有条带,可能为假阳性,图7为随机选取的8个T20-7菌落PCR结果。

图 7 淡紫紫孢菌20-7转化子的PCR检测
Figure 7. PCR detection of Transformants of P. lilacinum 20-7
-
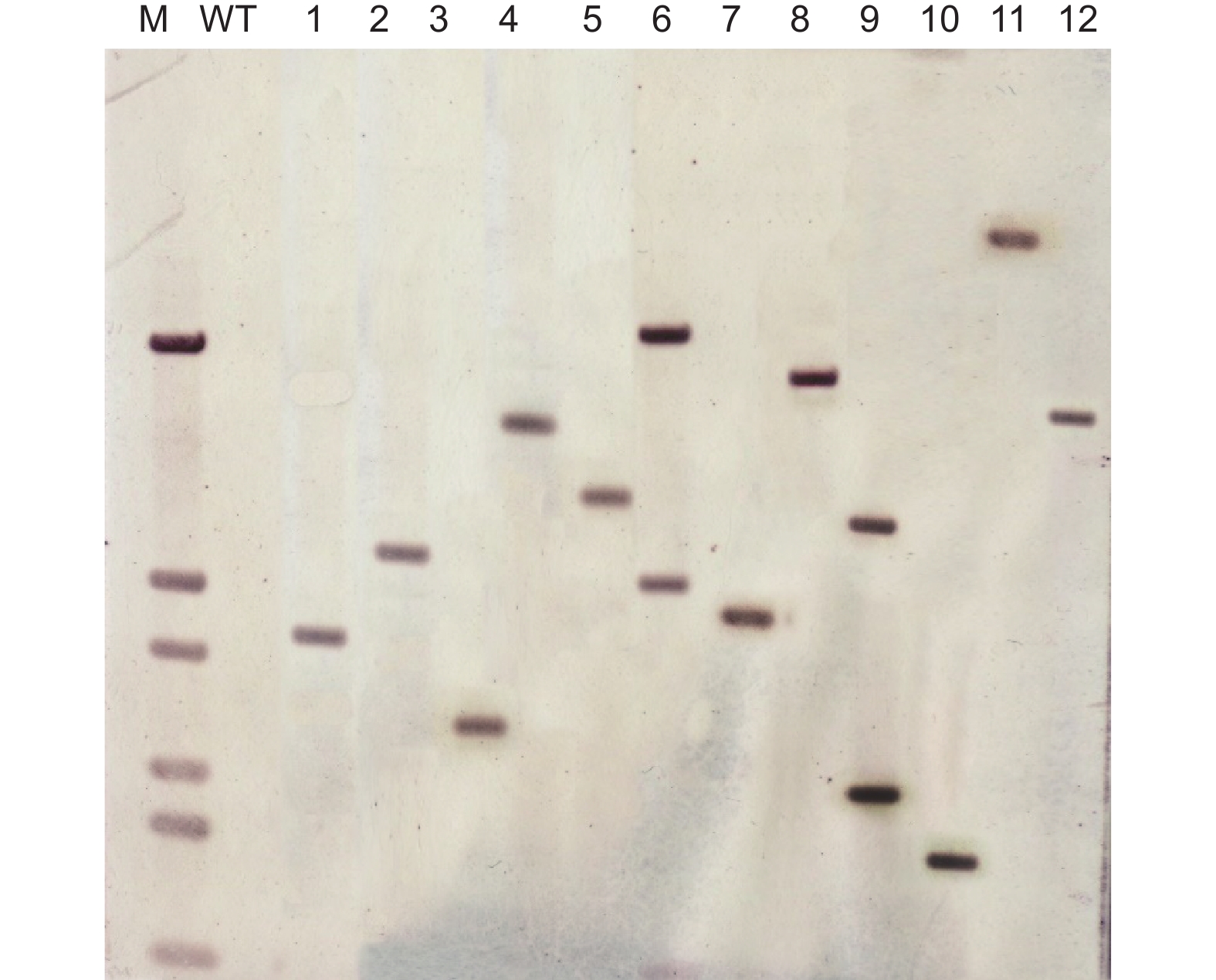
从根癌农杆菌介导转化体系突变体库中随机选取的12个转化子中,每个转化子都有不一致的Southern杂交带,说明T-DNA是随机整合的。有10个转化子是单一杂交带,表明这10个转化子T-DNA插入是单拷贝的,占83.3%。另外2个转化子(6号和9号)有两条杂交带,说明这2个转化子其T-DNA插入为多拷贝,占16.7%(图8)。

图 8 转化子Southern杂交验证T-DNA插入位点及拷贝数
Figure 8. Southern blotting of randomly selected transformants for the T-DNA insertion
-
在PDA(苯菌灵 300 µg·mL−1)平板上接种原始菌株20-7和转化子72 h后,转化子T20-7菌落扩展2 cm左右,而20-7菌落没有扩展,被苯菌灵抑制。T20-7连续培养共5代,仍然不被苯菌灵抑制,从而表明转化子T20-7对苯菌灵的抗性稳定遗传。
-
随机选取的20个转化子对南方根结线虫卵寄生率测定表明:仅1个转化子对南方根结线虫卵的寄生率显著增大,卵寄生率为86.67%,比野生型菌株20-7的卵寄生率67.64%提高了19.03%;而15个突变体侵染卵的效果明显下降,转化子最低寄生率为20.12%,与野生型菌株20-7相比下降了47.52%,结果最为显著(P=0.05);仅有4个突变体的致病力与20-7相比并无显著性差异(图9)。

图 9 不同转化子对南方根结线虫卵寄生率
Figure 9. Parasitic rate of transformants of P. lilacinum against M. incognita eggs
-
根癌农杆菌介导转化法具有转化效率高、遗传稳定、适用范围广等诸多优点,已成为真菌遗传转化研究中的强有力手段,在真菌基因资源开发、真菌性疾病研究和外源蛋白表达研究中发挥巨大作用[7],到2020年,已经对100余种真菌进行了遗传转化[22],但此种方法仍有不足之处,在真菌中的应用不完全成熟,如农杆菌种类、共培养温度及时间等众多因素都会对转化效率产生影响,不同菌株甚至同一菌株不同受体材料之间转化效率差异较大,限制了该技术的推广使用[23]。在本研究中,淡紫紫孢菌萌发的孢子与农杆菌EHA105在25 ℃共振荡培养48 h时,且在共培养阶段当乙酰丁香酮浓度为 200 µg·mL−1(pH5.5)时转化效率最高。对于根癌农杆菌介导淡紫紫孢菌遗传转化,筛选标记对抗性菌株的比例有很大影响。以膦丝菌素乙酸转移酶( bar) 基因作为选择标记建立的淡紫拟青霉的遗传转化体系[24],57% 的抗性转化子PCR 检测为阳性;以大肠杆菌转座子Tn903 编码的氨基糖苷磷酸转移酶( G-418 ) 基因作为筛选标记,79%抗性转化子的PCR 为阳性[17],而本研究以beta-tubulin基因为筛选标记,96%抗性转化子的PCR为阳性,通过比较说明选择合适的标记对转化子的筛选尤为重要,本研究证明,beta-tubulin基因作为选择标记适合淡紫拟青霉的遗传转化。
淡紫紫孢菌的遗传改良主要是以提高生防效率为目标,本研究在利用根癌农杆菌介导转化体系成功转化的淡紫紫孢菌转化子库中,成功筛选到了致病力发生变化的转化子,而高致病力强的菌株有可能作为优良菌株用于实际生产;由于其带有beta-tubulin基因,对杀菌剂苯菌灵具有抗性,提高了淡紫紫孢菌的生态适应性和应用性,对协调生物防治与化学防治,以及提高淡紫紫孢菌杀线虫剂的应用效果均具有重要的科学意义和应用价值。
-
本研究构建了农杆菌介导的、以基因beta-tubulin为选择标记的淡紫紫孢菌高效遗传转化体系,成功将携带 beta-tubulin基因的T-DNA整合到淡紫紫孢菌基因组中,转化菌株对苯菌灵的抗性可正常表达且稳定遗传,还获得了致病力变异的插入转化子,为淡紫紫孢菌的致病机制研究及优良菌株选育奠定了基础。
以beta-tubulin基因为选择标记的淡紫紫孢菌遗传转化
Efficient Transformation System of Agrobacterium tumefaciens Mediated Transformation of Purpureocillium lilacinum by Using beta-tubulin as Selectable Marker
-
摘要:
目的 建立稳定的食线虫真菌淡紫紫孢菌遗传转化体系,并获得插入突变体。 方法 介导的方法,以淡紫紫孢菌20-7的分生孢子为受体,将新构建的携带beta tubulin基因的质粒转化进入淡紫紫孢菌的细胞中,通过优化诱导乙酰丁香酮(AS)的浓度、诱导培养时间、农杆菌终浓度 OD660值、共培养 AS 的浓度、共培养时间和共培养温度等因子,建立高效遗传转化体系,获得致病力不同的突变体。 结果 共培养过程中使用萌发孢子是成功建立淡紫紫孢菌遗传转化体系的必要条件;淡紫紫孢菌萌发的孢子与农杆菌EHA105在25 ℃共振荡培养48 h时,且在共培养阶段当乙酰丁香酮浓度为 200 µg·mL−1(pH5.5)时转化效率最高,转化效率为1 200~3 200个转化子/106分生孢子, 阳性抗性转化子比率为96%;转化子PCR表明,T-DNA已整合到淡紫紫孢菌的基因组中;Southern 杂交验证表明,83.3% 的转化子为T-DNA 单拷贝插入;成功建立了可靠的淡紫紫孢菌的遗传转化体系,并从20个转化子中筛选到16个致病力变异的突变体。 结论 本研究成功构建了农杆菌介导的、以beta-tubulin基因为选择标记的淡紫紫孢菌高效遗传转化体系,并获得致病力变异的插入突变体,为淡紫紫孢菌的基因功能、致病机制研究及优良菌株选育奠定了基础。 -
关键词:
- 淡紫紫孢菌
- / 根癌农杆菌
- / beta-tubulin
- / 插入突变
- / 致病力
Abstract:Purpose To establish an efficient transformation system of the nematopathogenic fungus Purpureocillium lilacinum and obtain its insertional mutagenesis. Methods The benomyl resistance gene beta-tubulin being as the selective marker, Agrobacterium tumefaciens-mediated transformation technique was developed to screen different pathogenicity mutants in P. lilacinum. PCR amplification and Southern hybridization were used to verify the transformation events, and Southern blotting of beta-tubulin gene and cloning of transforming DNA (T-DNA) flanking sequences were used to determine insert number and site of T-DNA in the fungal genome, respectively. Results A reliable transformation method was established for P. lilacinum. Specifically, pre-germinating spores of P. lilacinum used at co-cultivated period was a prerequisite.P. lilacinum germinating spores co-cultivated with A. tumefaciens EHA105 at 25 ℃ for 48 h achieved the highest transformation efficiency, which was 1 200-3 200 transformants per 106 spores, and the ratio of positive resistant transformants was 96%. The transformants were cultivated up to 5 generations on beta-tubulin-containing medium and confirmed by PCR and those genetic traits remained stable. Southern hybridization showed that 83.3% of the transformants were single copy insertions of T-DNA, and 16 mutants with virulence variants were screened from 20 transformants. Conclusion This study successfully constructed an efficient genetic transformation system mediated by A. tumefaciens with beta-tubulin gene as a selective marker, and obtained an insertion mutant with pathogenicity variation, which was P. lilacinum. It provides insights into studying gene function, pathogenic mechanism and breeding excellent strains. -
图 2 乙酰丁香酮浓度对淡紫紫孢菌20-7转化效率的影响
Figure 2. Effects of different acetosyringone concentrations on the transformation efficiency of P. lilacinum 20-7
图 3 农杆菌菌株浓度对淡紫紫孢菌20-7转化率的影响
Figure 3. Effects of different Agrobacterium strains on the conversion rate of P. lilacinum 20-7
图 4 pH对淡紫紫孢菌20-7转化效率的影响
Figure 4. Effect of pH on transformation efficiency of P. lilacinum 20-7
图 5 时间对淡紫紫孢菌20-7转化效率的影响
Figure 5. Effect of time on transformation efficiency of P. lilacinum 20-7
图 6 温度对淡紫紫孢菌20-7菌株转化效率的影响
Figure 6. Effect of temperature on transformation efficiency of P. lilacinum 20-7
图 8 转化子Southern杂交验证T-DNA插入位点及拷贝数
Figure 8. Southern blotting of randomly selected transformants for the T-DNA insertion
图 9 不同转化子对南方根结线虫卵寄生率
Figure 9. Parasitic rate of transformants of P. lilacinum against M. incognita eggs
表 1 苯菌灵浓度对淡紫紫孢菌20-7菌落生长的影响
Table 1. Effects of different concentrations of benomyl on the growth of P. lilacinum 20-7
浓度
Concentration/
(µg·mL−1)20-7菌落直径/cm
The colony diameter of P. lilacinum /cm48 h 72 h 96 h 108 h 0 2.11 5.13 7.57 9.12 100 0.34 0.83 1.46 1.94 200 0.01 0.03 0.04 0.06 300 0 0 0 0 400 0 0 0 0 500 0 0 0 0 1 000 0 0 0 0  下载: 导出CSV
下载: 导出CSV
表 2 淡紫紫孢菌20-7菌株孢子新鲜程度对转化效率的影响
Table 2. Effects of different degree of spore freshness on transformation efficiency of P. lilacinum 20-7
孢子新鲜程度
Spores freshness转化子个数
Transformants /(×10−6)保存一个月的孢子
Spores saved for a month187 新鲜孢子
Fresh spores3 019
下载: 导出CSV
表 3 共培养方法对淡紫紫孢菌20-7转化效率的影响
Table 3. Effects of different co-culture methods on transformation efficiency of P. lilacinum 20-7
共培养方法
Co-culture method转化子个数
Transformants/ (×10−6)振荡培养 Shake the medium 3 208 涂于滤纸片上 Coat on filter paper 1 948
下载: 导出CSV
-
[1] KERRY B R. An assessment of progress towards microbial control of plant parasitic nematodes[J]. Journal of Nematology, 1990, 22: 621-631. [2] ZHANG S W, GAN Y T, XU B L. Biocontrol potential of a native species of Trichoderma longibrachiatum against Meloidogyne incognita[J]. Applied Soil Ecology, 2015, 94: 21-29. doi: 10.1016/j.apsoil.2015.04.010 [3] GINE A, SORRIBAS F J. Effect of plant resistance and BioAct WG (Purpureocillium lilacinum strain 251) on Meloidogyne incognita in a tomato-cucumber rotation in a greenhouse[J]. Pest Management Science, 2017, 73(5): 880-887. doi: 10.1002/ps.4357 [4] KOOLIYOTTIL R, DANDURAND L M, KNUDSEN G R. Prospecting fungal parasites of the potato cyst nematode Globodera pallida using a rapid screening technique[J]. Journal of Basic Microbiology, 2017, 5(57): 386-392. [5] 柯心如, 刘登艳, 谭建彬, 等. 一株淡紫紫孢菌的分离、鉴定及生物学特性研究[J]. 广东农业科学, 2022, 49(6):108-117. [6] GROOT M D, BUNDOCK P, HOOYKAAS P, et al. Agrobacterium tumefaciens-mediated transformation of filamentous fungi[J]. Nature Biotechnology, 1998, 16(9): 839-42. doi: 10.1038/nbt0998-839 [7] FURLANETO M C, PAIÃO F G, PINTO F G S, et al. Transformation of the entomopathogenic fungus Metarhizium flavoviride to high resistance to benomyl[J]. Canadian Journal of Microbiology, 1999, 45: 875-878. doi: 10.1139/w99-074 [8] MULLINS E D, CHEN X, ROMAINE P, et al. Agrobacterium-mediated transformation of Fusarium oxysporum: an efficient tool for insertional mutagenesis and gene transfer[J]. Phytopathology, 2001, 91: 173-180. doi: 10.1094/PHYTO.2001.91.2.173 [9] ROGERS C W, CHALLEN M P, GREEN J R, et al. Use of REMI and Agrobacterium-mediated transformation to identify pathogenicity mutants of the biocontrol fungus Coniothyrium minitans[J]. FEMS Microbiology Letters, 2004, 241: 207-214. doi: 10.1016/j.femsle.2004.10.022 [10] REIS M C, PELEGRINELLI F M H, DELGADO R T, et al. Agrobacterium tumefaciens-mediated genetic transformation of the entomopathogenic fungus Beauveria bassiana[J]. Journal of Microbiological Methods, 2004, 58: 197-202. doi: 10.1016/j.mimet.2004.03.012 [11] LI M X, GONG X Y, ZHENG J, et al. Transformation of Coniothyrium minitans, a parasite of Sclerotinia sclerotiorum, with Agrobacterium tumefaciens[J]. FEMS Microbiology Letters, 2005, 243: 323-329. doi: 10.1016/j.femsle.2004.12.033 [12] FANG W, PEI Y, BIDOCHKA M J. Transformation of Metarhizium anisopliae mediated by Agrobacterium tumefaciens[J]. Canadian Journal of Microbiology, 2006, 52(7): 623-626. doi: 10.1139/w06-014 [13] DUARTE R T D, STAATS C C, FUNGARO M H P, et al. Development of a simple and rapid Agrobacterium tumefaciens-mediated transformation system for the entomopathogenic fungus Metarhizium anisopliae var. acridum[J]. Letters in Applied Microbiology, 2007, 44: 248-254. doi: 10.1111/j.1472-765X.2006.02092.x [14] 孙文良, 胡晓璐, 吴萌章, 等. 根癌农杆菌介导的深绿木霉菌T23遗传转化研究[J]. 上海交通大学学报:农业科学版, 2009, 27(5):489-493. [15] 赵津津. 蜡蚧轮枝菌“Lecanicillium lecanii (Zimmerman)Viegas”苯菌灵抗性基因转化的研究[D]. 北京: 中国农业科学院, 2011. [16] DNURM A, BAILEY A M, CAIRNS T C, et al. A silver bullet in a golden age of functional genomics: the impact of Agrobacterium-mediated transformation of fungi[J]. Fungal Biology and Biotechnology, 2017, 4(1): 6. doi: 10.1186/s40694-017-0035-0 [17] 王曦茁, 朴春根, 李 虹, 等. 根癌农杆菌介导的淡紫拟青霉遗传转化体系的建立[J]. 林业科学, 2010, 46(10):95-102. [18] 王关林. 植物基因工程原理与技术[M]. 北京: 科学出版社, 1998. [19] 黄培堂, 王嘉玺, 朱厚础. 分子克隆实验指南[M]. 北京: 科学出版社, 2002. [20] AUSUBEL F, BRENT R, KINGSTON R E, et al. Short Protocols in Molecular Biology. 3rd edition[M]. John Wiley & Sons, 1995: 228-244. [21] 汪来发, 杨宝君, 李传道. 寄生真菌对根结线虫的致病力评价[J]. 林业科学, 1999, 35(3):41-47. [22] MICHIELSE C B, HOOYKAAS P, HONDEL C, et al. Agrobacterium-mediated transformation as a tool for functional genomics in fungi[J]. Current Genetics, 2005, 48: 1-17. doi: 10.1007/s00294-005-0578-0 [23] 胡 懋, 曾杨璇, 苗华彪, 等. 根癌农杆菌介导真菌遗传转化的研究及应用[J]. 微生物学通报, 2021, 48(11):4344-4363. doi: 10.13344/j.microbiol.china.210156 [24] 赵培静, 任文彬, 缪承杜, 等. 2007. 根癌农杆菌介导的淡紫拟青霉遗传转化[C]//中国植物病理学会. 中国植物病理学会2007年学术年会论文集. 北京: 391-397. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3265
- HTML全文浏览量: 929
- PDF下载量: 62
- 被引次数: 0
