

-
干旱严重影响植物的生长、发育和产量,是导致植物死亡的重要因素之一[1-2]。为适应生存环境,植物进化出各种响应机制。其中,双组分系统(Two-component systems,TCS)是植物响应环境信号的重要通路之一[3]。根据TCS组成成分种类以及蛋白结构域的不同,将其分为传统型和复杂型。传统型TCS主要由组氨酸激酶(Histidine kinases,HKs)和响应调节因子(Response regulators,RRs)构成,而复杂型TCS往往于两组分间增添了磷酸转移蛋白(Histidine phosphotransfer proteins,HPts)[4]。在植物等真核生物中,细胞信号传导依赖于后者。TCS的作用机制在于各组分间的磷酸基团转移。植物通过TCS感知信号并做出响应的一般流程包括:首先HKs上的感应结构域接收环境信号,催化其激酶结构域中的组氨酸残基(His)发生自磷酸化,随后磷酸基团转移至接收结构域内的天冬氨酸残基(Asp);接着HPts接受磷酸基团,并将磷酸基团进一步转移至RRs上的调节结构域,随之激活其效应结构域,从而使植物做出响应[5]。
在拟南芥(Arabidopsis thaliana(L.)Heynh.)中共鉴定到56个TCS相关基因,其中HKs及相关基因共17个,HPts基因6个,RRs基因33个[6]。拟南芥组氨酸激酶又可分为AHK、乙烯受体(Ethylene receptors)及光敏色素光感受器(Phytochrome photoreceptors)3大家族,AtHK1、AtHK2、AtHK3、AtHK4(CRE1/WOL)、AtHK5(CKI2)以及AtCKI1构成了AHK家族。其中,AtHK2、AtHK3以及AtCRE1可感知细胞分裂素信号,被称为细胞分裂素受体[7-8]。而细胞分裂素的信号传导过程与植株响应干旱胁迫和调节植株胁迫适应性存在密切联系[9]。研究表明,PEG模拟干旱胁迫条件下,拟南芥组氨酸激酶基因突变体ahk3植株根长生长量和植株鲜质量均大于野生型[10];同时ahk2、ahk3突变体株系存活率更高,抗旱性更强[11]。基因表达模式分析表明,磷酸转移蛋白基因AtHP2/3/5在干旱胁迫/盐胁迫处理后均下调表达,并且干旱胁迫下ahp2,3,5突变体表现出更高的相对含水量和更低的相对电导率,即细胞膜稳定性更强,完整度更高,更加抗旱[12]。与AtHP2/3/5相似,响应调节蛋白基因AtRR1/10/12表达水平在干旱或ABA处理后表现为下调趋势。干旱条件下,arr突变体通过维持较高含水量,进而增强抗旱能力[13]。因此,细胞分裂素信号传导通路各组分在植物响应干旱胁迫的过程中发挥重要作用。
杨树(Populus spp.)是我国重要的生态防护、绿化建设和工业用材树种,在改善生态环境及保障木材产量方面发挥重要作用。而我国干旱或半干旱地区占据国土面积50%以上,严重影响杨树的生态和经济效益[14]。因此,培育杨树抗旱新品种对于环境资源的改善与利用具有重要意义。近年来随着林木基因工程技术的推广和完善,杨树抗旱相关基因功能研究及分子育种工作取得重大进展,例如与非转基因杨树相比,转果聚糖蔗糖转移酶SacB基因银腺杨、转脱水应答元件DREB1C基因南林895杨(Populus euramericana cv.‘Nanlin 895’)和转锌指蛋白转录因子ZxZF基因欧美杨(P. × euramericana cl.‘Bofeng1’)在抗旱性能方面均有所提高[15-17]。而目前关于组氨酸激酶基因在杨树中的研究报道较少,其中在毛果杨(P. trichocarpa Torr. & Gray)基因组中有2个 AtHK3的同源基因PtHK3a和PtHK3b,并且这两个基因均在形成层中表达量较高[18]。鲁俊倩等[19-20]通过干旱、盐、高温、低温胁迫以及激素处理发现,银腺杨(P. alba × P. glandulosa ‘84k’)HK3基因(PagHK3a、PagHK3b)在赤霉素、脱落酸、细胞分裂素和水杨酸处理后表达下调,在干旱、盐、高温和低温胁迫条件下表达上调,说明PagHK3a和PagHK3b基因可能在杨树抗逆境胁迫过程中发挥作用。
本研究以2个银腺杨组氨酸激酶PagHK3a基因敲除株系C1、C2及其野生型(WT)为试验材料,通过测定干旱胁迫下各株系的生理、生化与生长等指标,对基因敲除株系的抗旱性进行评价,同时探究PagHK3a基因在杨树响应干旱胁迫过程中的相关功能,为培育抗旱基因编辑林木新种质提供参考。
-
试验材料为银腺杨组氨酸激酶基因PagHK3a基因敲除株系C1和C2及其野生型(WT)。C1和C2株系的2个Cas9靶点(sgRNA1,sgRNA2)均位于PagHK3a上的组氨酸激酶结构域(Histidine kinase domain),其中C1株系编辑类型为靶点1单碱基缺失,靶点2双碱基缺失纯合体;C2株系编辑类型为靶点1单碱基缺失,靶点2单碱基缺失纯合体(图1A)。C1、C2株系均发生移码突变,翻译提前终止(图1B)。

图 1 银腺杨及其PagHK3a基因敲除株系C1和C2靶点编辑情况
Figure 1. Schematic illustration of the sgRNA target site in wild type (WT) of P. alba × P. glandulosa and two mutant lines (C1 and C2)
-
对WT及基因敲除株系C1和C2进行组培扩繁,待生长25 d后,选取生长一致的组培苗用于胁迫处理,每株系3个生物学重复;将组培苗从固体培养基中小心取出,清水冲洗根系附着琼脂后,移至霍格兰营养液(Hoagland’s Nutrient Solution)中进行液体环境适应,每天更换一次营养液;5 d后移至含有5%PEG的霍格兰营养液中进行模拟干旱胁迫处理,胁迫3 h后,取成熟叶片,液氮速冻后用于RNA提取、cDNA反转录及相关基因表达量的测定。
-
将培养4周的组培苗移栽至上口径12 cm,下口径8 cm,高14 cm的塑料花盆中,每株系30株,栽培基质为草炭土与珍珠岩(比例为5:1),移栽前称量质量并保持一致。生长地点位于中国林业科学研究院林木遗传育种全国重点实验室温室,平均温度约为25~30 ℃,平均相对湿度为50%~60%,通风条件较好,苗木生长期间进行正常水肥及病虫害管理。
温室培养45 d后,选取生长状态一致的盆栽苗进行干旱胁迫处理,每株系处理9株。试验共分为3种水分处理方式:正常浇水、中度干旱和重度干旱,分别对应于田间持水量的70%~75%、40%~45%和25%~30%,每种处理3个生物学重复。试验采用称重控水法,于每天早晨8:00称取各盆质量,根据计算结果补充相应水分至试验设计范围内。胁迫4周后,进行生化、光合参数和生长指标的测定。
-
通过NCBI(
https://www.ncbi.nlm.nih.gov/ )Primer Blast功能设计实时定量PCR引物(表1),以Actin(Pop_A01G010149.T1)基因为内参基因,测定PagHK3a(Pop_G01G074644.T1)、PagRR2(Pop_G08G060457.T1)、PagRR15(Pop_G15G048786.T1)、PagNAC3(Pop_A01G074938.T1)、SOD4(Pop_G05G073560.T1)和POD1(Pop_A16G089950.T1)的相对表达量。表 1 实时定量PCR引物序列
Table 1. The primer sequences used for quantitative real-time PCR
基因
Gene引物序列
Primer sequence(5′→3′)产物长度
Product length/bpPagHK3a F: ACCGCACAAGACAGTGGAAA 100 R: ACTGCATTGCCTCAAGCTCA PagRR2 F: CAAACCCTCAAACTGCCCTC 103 R: TCAGCTAGTCCAATGCCTCC PagRR15 F: GCGAAGTTTGGCGGATGAAG 161 R: TGCTCCCACTCTCTACAACAG PagNAC3 F: TCAACTCGGTGCCTCAACTC 161 R: AGCTCCACTTTGAACCTCCC SOD4 F: CAGTGGAGGTGCATTGGAAG 138 R: GCGAAGGGTTGCCATTGTTA POD1 F: TAGCAACATCCCTTCCCCAG 128 R: TGGTACATCTTGCTTGCCCT Actin F: TGCGTATGTTGCCCTTGACT 157 R: TCCGATGAGAGAAGGCTGGA PEG处理3 h后,分别取WT及基因敲除株系组培苗成熟叶片0.1 g,利用RNAprep Pure多糖多酚植物总RNA提取试剂盒(天根,北京)提取叶片总RNA,经1%琼脂糖凝胶电泳检测RNA提取质量,NanoDrop One(Thermo Fisher Scientific,美国)测定RNA浓度及纯度后,利用PrimeScriptTM RT Master Mix(Perfect Real Time)试剂盒(Takara,日本)进行cDNA合成。将反转录得到的cDNA稀释10倍后作为实时定量PCR的模板,按照TB Green TM Premix Ex TaqTM II(TliRNaseH Plus)试剂盒(Takara,日本)说明配制反应体系:TB Green Premix TaqII(TliRNaseH Plus,2 × )10 μL,Primer-F/R各0.8 μL,cDNA模板2 μL,ddH2O补足至20 μL。利用LightCycler480 System(Roche,瑞士)进行实时定量PCR反应,反应条件如下:预变性95 ℃ 30 s,扩增阶段95 ℃ 5 s,60 ℃ 30 s,45个循环;熔解阶段95 ℃ 5 s,60 ℃ 1 min,95 ℃;降温阶段50 ℃ 30 s。每个株系3次技术性重复,最后通过2−ΔΔCT 法计算各基因的相对表达量[21]。
-
采用苏州科铭生物技术有限公司相关试剂盒测定功能叶片丙二醛(MDA)含量、过氧化氢(H2O2)含量、超氧化物歧化酶(SOD)活性和过氧化物酶(POD)活性。测定方法为微量法,测定仪器为SpectraMax 190(Molecular Devices,美国),具体操作流程参考各试剂盒说明书。
-
于胁迫处理最后一天上午9:00—11:30,选取自茎尖向下第4~6片完全展开的功能叶片进行光合参数测定。光合参数测定采用LI-6400XT(Li-COR,美国)便携式光合作用测量系统,测定指标为净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)以及蒸腾速率(Tr);仪器光合有效辐射设定为1 000 μmol· m−2s−1,CO2浓度设定为400 μmol·mol−1。
-
在胁迫处理前后,分别利用钢尺及游标卡尺测定植株株高与地径,株高或地径生长量为胁迫前后株高或地径的差值;胁迫处理第28 d时,将植株根系从盆中取出,清理表面基质,自来水冲洗干净,吸水纸吸干后置于干净信封中,烘箱72 ℃烘干48 h后测定根系干质量。
-
分别采用Excel 2019进行数据整理与表格制作,通过SPSS 21.0软件进行数据的单因素方差分析(one-way ANOVA)与多重比较(Duncan新复极差法),利用Graphpad Prism 9.0.0进行统计图的绘制。
-
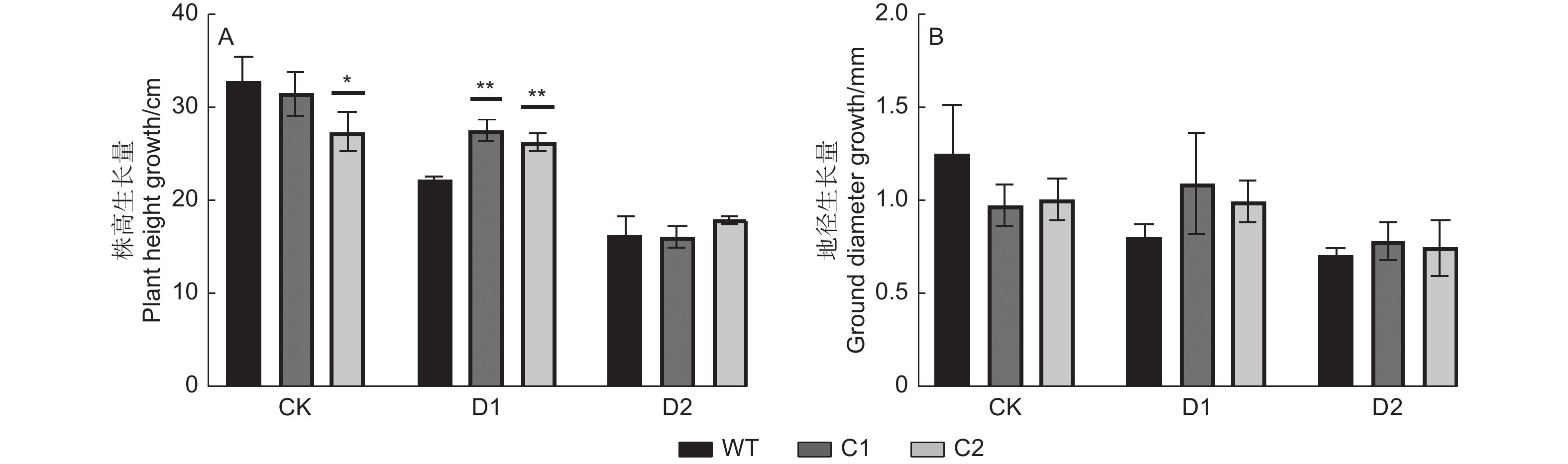
水分亏缺条件下,植物生长指标的变化能够直接反映其抗旱能力的强弱。本试验中,分别测定了基因敲除株系C1、C2及WT株系胁迫前后的株高与地径,结果显示:正常浇水条件下,PagHK3a基因敲除株系C1和C2株高生长量均低于WT,其中C2株系与WT相比差异显著;随着干旱胁迫程度的加深,C1、C2及WT株系的株高生长量均呈现出下降趋势,其中WT株系下降幅度较大,与正常浇水相比,中度干旱(D1)条件下WT株系株高生长量减少10.6 cm,而基因敲除株系C1减少3.9 cm,C2株系减少1.1 cm。在重度干旱(D2)条件下,WT株系株高生长量减少16.6 cm,而C1株系减少15.4 cm,C2株系减少9.5 cm。并且在中度干旱条件下,基因敲除株系平均株高生长量显著大于WT,与WT相比,C1株系株高生长量高出5.3 cm,C2株系高出4.0 cm;而在重度干旱处理中,基因敲除株系C1、C2与WT相比差异不显著。另外,正常浇水及干旱条件下,基因敲除株系的地径生长量与WT株系差异均不显著(图2)。

图 2 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT之间株高和地径生长量的差异
Figure 2. Differences in plant height and ground diameter growth between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
对根干质量测定结果进行差异显著性分析(表2)发现,与正常浇水相比,干旱胁迫处理后,WT及基因敲除株系的根系均变得更加发达,根干质量相应增加,尤其是在重度干旱条件下,基因敲除株系C1与C2以及WT的根干质量均显著增加。然而,无论是在正常浇水还是在干旱胁迫下,两个基因敲除株系的根干质量与WT相比差异均不显著。
表 2 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT之间根干质量差异
Table 2. Differences in root dry weight between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
株系
Lines根干质量/g
Root dry weightCK D1 D2
PWT 0.50 ± 0.17 aB 0.64 ± 0.03 aAB 0.84 ± 0.13 aA 0.043 C1 0.48 ± 0.05 aC 0.67 ± 0.12 aB 0.92 ± 0.09 aA 0.003 C2 0.56 ± 0.05 aB 0.71 ± 0.03 aA 0.77 ± 0.09 aA 0.014 P 0.687 0.567 0.276 -- 注:表格中数据表现格式为均值 ± 标准差,其后大写字母表示为同一株系在不同处理条件下的差异显著性,小写字母表示为同一种处理方式下不同种株系之间的差异显著性,字母相同表示差异不显著,字母不同表示差异显著,显著性水平为0.05 Notes: The data in the table are presented in the format of mean ± standard deviation, followed by capital letters indicating the significance of the difference between the same strain under different treatment conditions, and lowercase letters indicating the significance of the difference between different strains under the same treatment, with the same letter indicating that the differences are not significant, and different letters indicating that the differences are significant, with a significance level of 0.05 -
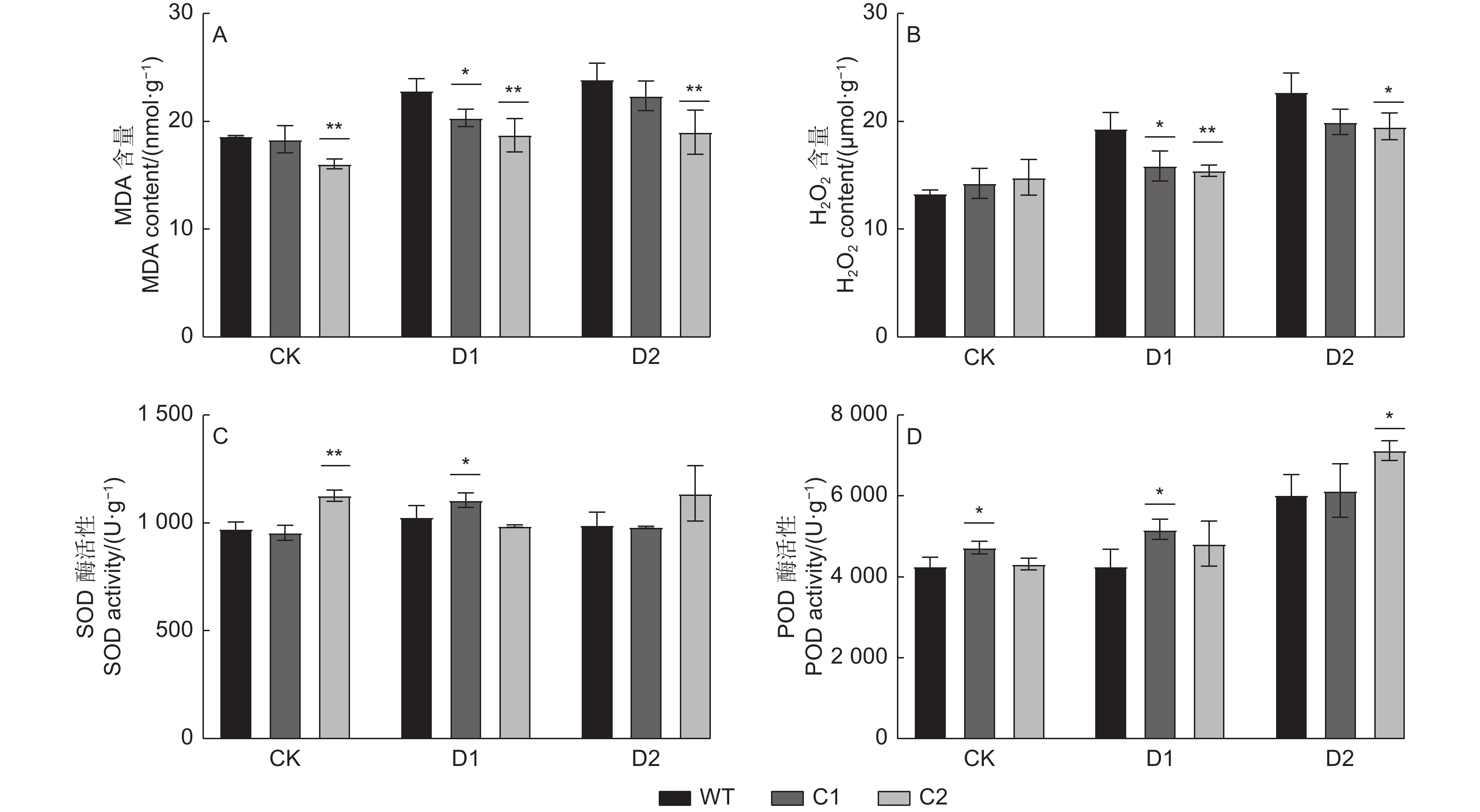
胁迫处理后,植物体内氧化还原平衡出现紊乱,活性氧ROS过度积累,进而使得植物防御系统受损,最终造成一定伤害[22-23]。本试验中,对正常浇水及干旱胁迫条件下基因敲除株系C1、C2与WT株系叶片MDA、H2O2含量以及SOD、POD酶活性进行测定分析,结果如图3所示:与正常浇水相比,干旱胁迫下,各株系MDA含量均表现为增加的趋势。另外在正常浇水条件下,C2株系的MDA含量显著低于WT;在中度干旱条件下,C1、C2株系MDA含量均显著低于WT,即中度干旱胁迫下,WT细胞膜受损程度更加严重;而在重度干旱条件下,仅C2株系MDA含量极显著低于WT,C1株系与WT相比差异不显著(图3A)。同时结果显示,与正常浇水相比,WT株系与2种基因敲除株系的H2O2含量在干旱胁迫后也有所增加,但对比发现,胁迫处理后,WT株系变化幅度更大,其中在正常浇水条件下,C1、C2株系H2O2含量与WT株系差异不显著;在中度干旱条件下,C1、C2株系H2O2含量均显著低于WT,而在重度干旱组中,仅C2株系显著低于WT(图3B)。即在中度干旱胁迫下,基因敲除株系受H2O2影响显著小于WT。SOD酶活性测定结果(图3C)显示,在正常浇水条件下,C2株系显著高于WT;在中度干旱条件下,C1株系显著高于WT;而在重度干旱胁迫下,C1、C2株系SOD酶活性与WT相比均差异不显著。POD活性(图3D)测定结果显示,与正常浇水相比,干旱胁迫后,植株POD活性呈不断上升的变化趋势,在重度干旱处理组中各株系POD酶活性最高。另外,在正常浇水及中度干旱条件下,C1、C2株系POD酶活性均值高于WT,但在重度干旱条件下,仅C2株系显著高于WT。

图 3 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT生化指标差异
Figure 3. Differences in biochemical indexes between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
-
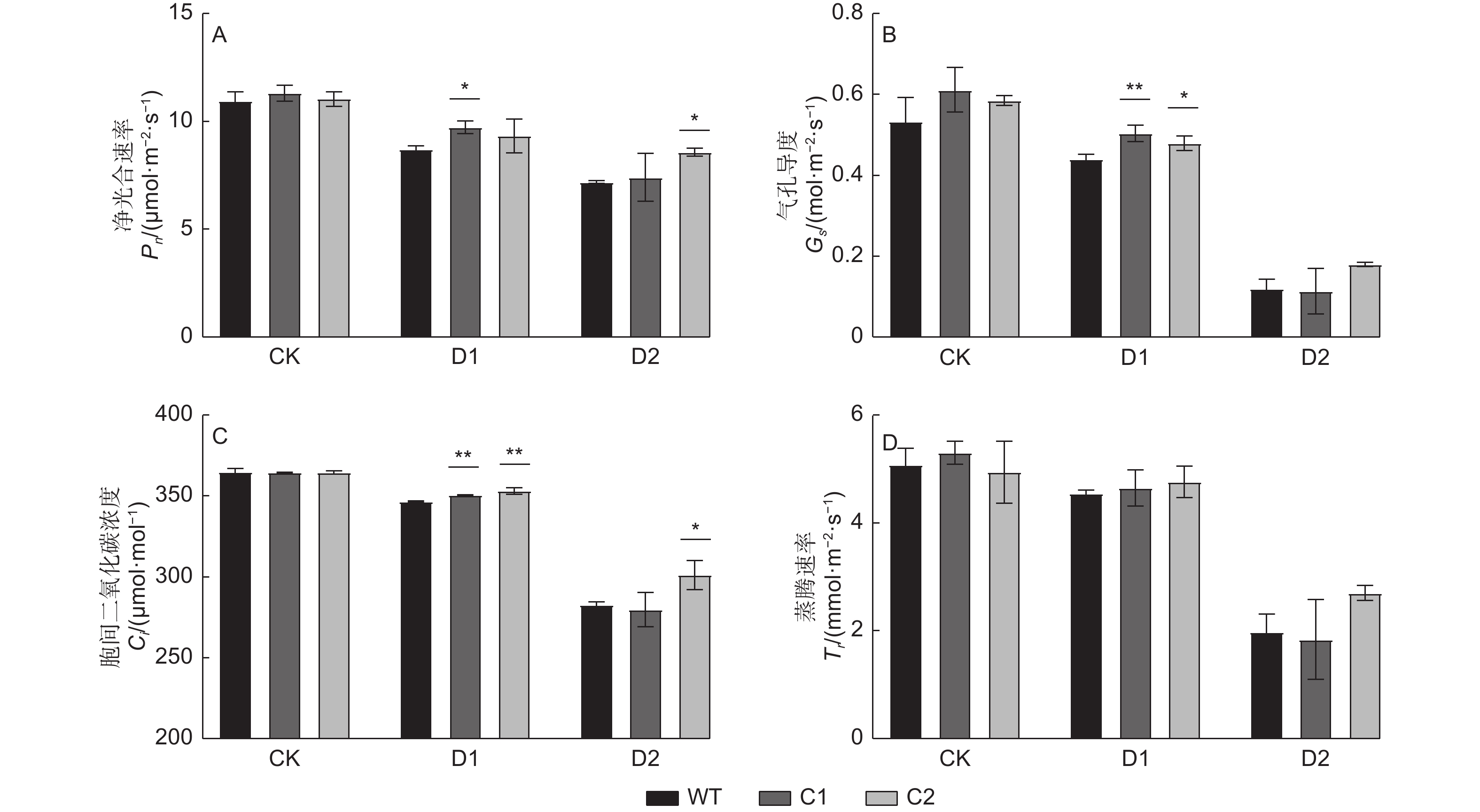
光合作用是植物生长发育所依赖的基本代谢过程。由于土壤含水量的降低,植株光合相关指标发生变化,从而影响植株生长[24]。胁迫处理末期,测定各株系光合参数(图4)发现:与正常浇水条件相比,干旱胁迫下各株系光合参数下降,并且随着干旱胁迫程度的加深,参数降低幅度逐渐增大。另外,在正常浇水条件下,与WT株系相比,基因敲除株系的4种光合参数均无显著差异;而在干旱胁迫条件下,基因敲除株系与WT相比有所差异。其中在中度干旱胁迫下,C1株系净光合速率显著大于WT株系,C2株系与WT相比差异不显著;在重度干旱条件下,C2株系净光合速率显著大于WT株系,C1株系与WT之间差异不显著(图4A)。气孔导度测定结果显示,在中度干旱条件下,与WT相比,C1株系显著高出15%,C2株系显著高出9%,即基因敲除株系均显著大于WT;而重度干旱条件下,基因敲除株系C1、C2与WT差异不显著(图4B)。比较分析胞间CO2浓度(图4C)发现,中度干旱条件下,C1、C2株系显著大于WT,而重度干旱条件下,仅C2株系显著大于WT。为减少水分过度损失,随着土壤含水量的降低,各株系蒸腾速率(图4D)均降低,但在干旱胁迫下,基因敲除株系的蒸腾速率与WT株系相比差异不显著。

图 4 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT之间光合参数差异
Figure 4. Differences in photosynthetic parameters between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
-
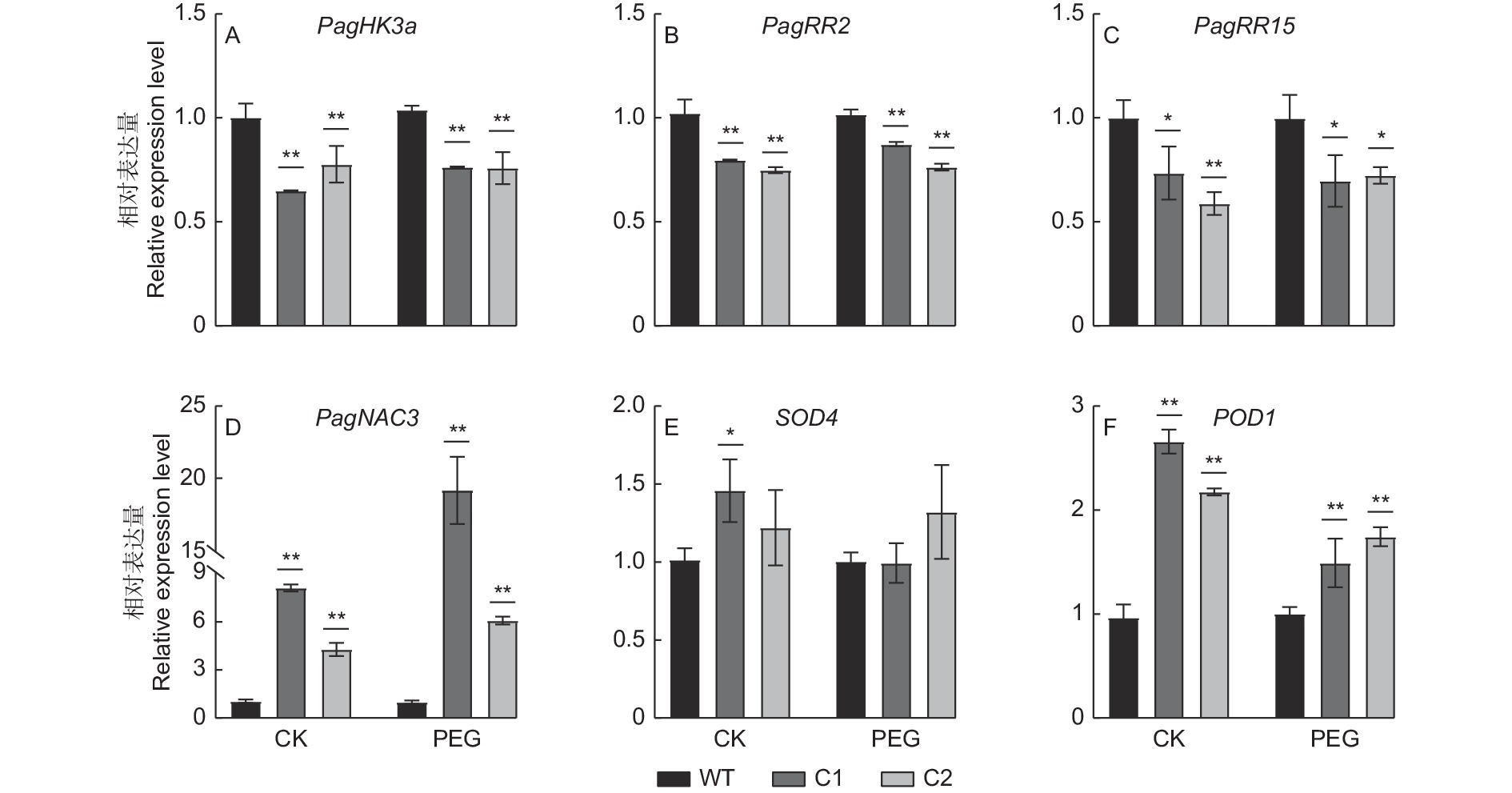
为比较干旱胁迫下PagHK3a基因敲除株系C1和C2及野生型株系WT间叶片中PagHK3a基因及其下游基因、胁迫相关基因、抗氧化相关基因表达差异,我们利用5%PEG处理各株系组培苗。实时定量PCR结果(图5)显示:与WT相比,基因敲除株系中PagHK3a及其下游响应调节因子PagRR2和PagRR15基因表达显著下降,而胁迫相关基因PagNAC3和POD1基因表达显著上升。其中,在非胁迫处理条件下,基因敲除株系PagHK3a基因表达水平均显著低于WT,其中C1株系与WT相比降低35.2%,C2株系与WT相比降低22.4%;在PEG胁迫处理条件下,与WT相比,C1株系PagHK3a表达水平降低26.5%,C2株系降低26.9%。同时,各株系下游响应调节因子PagRR2和PagRR15基因表达与PagHK3a基因相似,非胁迫处理及胁迫处理条件下,基因敲除株系基因表达水平均显著低于WT。胁迫响应相关基因PagNAC3的定量结果显示,在非胁迫处理条件下,C1株系PagNAC3基因表达水平为WT的7.7倍,C2株系PagNAC3基因表达量为WT的4.1倍;在PEG胁迫处理条件下,C1株系PagNAC3基因表达水平为WT的19.3倍,C2株系为WT的6.1倍(图5D)。超氧化物歧化酶合成基因SOD4表达结果显示,在非胁迫条件下,仅C1株系SOD4基因表达量显著高于WT,在PEG胁迫条件下,C1、C2株系与WT相比均差异不显著。而对过氧化物酶合成基因POD1表达分析发现,与WT相比,基因敲除株系C1、C2在PEG胁迫与非胁迫处理条件下均具有更高的表达水平,且差异显著。

图 5 PEG模拟干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2及WT叶片中相关基因表达水平
Figure 5. Expression levels of genes in leaves of PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa in Hoagland’s nutrient solution with 5% PEG
-
干旱胁迫严重影响植物的生长发育及生物量的积累[25]。研究表明,株高、地径等生长指标可以用于直观地反映胁迫条件下植株的生长状况[26]。本试验中,基因敲除株系C1和C2在株高生长层面表现出相对优势,其中,与WT相比,C1和C2株系的株高生长量在中度干旱条件下均显著增加。同时水是植物进行光合作用的原料之一,在水分亏缺条件下,植株光合作用减弱[27]。本试验结果显示,干旱处理后各株系4种光合参数均下降,并且重度干旱条件下各指标达到最低值。其中在中度干旱条件下,C1和C2株系的气孔导度及胞间CO2浓度均显著大于WT,表明基因敲除株系具有更强的气体交换能力,进而保障植株CO2同化率[28]。另外,干旱胁迫能够导致植株体内ROS的产生及消除之间的动态平衡紊乱,而ROS的过多积累使得植物发生脂质过氧化反应,导致植物细胞膜受损[22]。并且研究发现,MDA是细胞膜系统脂质过氧化反应过程中的重要产物,可用于反映植物细胞膜受损程度[29]。本研究结果显示,在中度干旱条件下,C1和C2株系的H2O2及MDA含量显著低于WT株系,即说明基因敲除株系受H2O2影响较小,细胞膜损伤程度更低。同时研究表明,细胞分裂素信号传导与植物响应干旱胁迫存在密切联系[9]。细胞分裂素信号传导依赖于TCS系统中的多级磷酸基团转移机制,例如在拟南芥中,组氨酸激酶AtHK3作为细胞分裂素受体,感知并接收细胞分裂素信号,通过下游HPts以及RRs之间的磷酸基团转移,实现信号的传递[30]。另外,RRs分为A、B两种类型,B型RRs接收来自HPts的磷酸基团,同时也可作为转录因子,调控A型RRs的转录[31]。本研究发现基因敲除株系中PagHK3a基因表达显著下调的同时,PagRR2(B型RRs)与PagRR15(A型RRs)基因表达也显著下调,因此推测PagHK3a下调表达引起了通路下游PagRR2表达量下降,进而影响了PagRR15的表达。与该结果相似,拟南芥ahk2ahk3cre1三突变体AtRR15的表达下调,同时突变体拟南芥表现得更加抗旱[11];而拟南芥ahk3功能获得型突变体ore12-1中,AtRR15则表达上调[32]。转录组测序结果表明,在干旱胁迫条件下,ahk2ahk3突变体、ahp2,3,5突变体中胁迫相关基因AtNAC3上调表达,并且AtNAC3基因过表达株系抗旱性增强[12,31,33]。与该结果类似,本试验的两个PagHK3a基因敲除株系PagNAC3基因的表达水平显著上调。因此推测,干旱胁迫下,PagHK3a基因敲除株系通过细胞分裂素信号传导途径RRs基因下调表达,及抗逆相关基因如NAC3、抗氧化基因POD1等的上调表达,降低了植株细胞膜氧化损伤,增强了植株气体交换能力,使植株具有更强的抗旱能力。
测序结果发现,本研究的两个PagHK3a基因敲除株系C1和C2,其靶点1编辑情况相同,均为纯合单碱基缺失,蛋白翻译均在2个靶点间提前终止,即靶点2的编辑类型差异不会影响其蛋白翻译结果,理论上这两个株系各项指标测定结果类似。然而,干旱胁迫结果发现,C1、C2株系间存在一定差异。例如:PEG模拟干旱胁迫下C1株系PagNAC3基因表达水平明显高于C2株系;温室重度干旱条件下,C2株系MDA含量、H2O2含量、POD酶活性、净光合速率以及胞间CO2浓度与WT相比均差异显著,而C1株系差异不显著。分析其中原因可能在于:一,在基因编辑操作中,发生在与目标编辑区域序列相似的位点上的非特异性编辑被称作“脱靶编辑”[34],尽管研究认为脱靶编辑在植物中发生频率较低[35-36],但由于脱靶效应造成的非预期编辑结果对基于基因编辑技术进行植物基因功能研究或分子育种研究可能产生的影响仍不可忽视。研究显示,由于脱靶效应,编辑拟南芥AtETC2基因的同时,AtCPC、AtTRY基因也被编辑,导致植株叶片较小且表面呈现簇状的毛状体等脱靶性状[37]。二,在植物组织培养过程中,由于继代次数、培养条件等原因易造成植物体细胞突变事件的发生[34,38],从而可能造成株系间存在差异。我们推测本试验中两个基因编辑株系的差异很可能是由脱靶效应或体细胞突变产生的,其产生的确切原因仍需进一步深入研究。
-
银腺杨PagHK3a基因敲除株系中细胞分裂素信号传导通路中响应调节蛋白编码基因PagRR2和PagRR15显著下调表达,诱导胁迫响应基因PagNAC3和抗氧化基因POD1显著上调表达;同时在中度干旱条件下,基因敲除株系C1和C2表现为膜系统氧化损伤更轻,H2O2含量更低,气体交换能力更强,株高生长量更大,进而比WT具有更强的抗旱能力。因此通过编辑杨树组氨酸激酶基因能够在一定程度上增强杨树抗旱性。
PagHK3a基因敲除对银腺杨抗旱性的影响
Effects of PagHK3a Gene Knockout on Drought Resistance of Populus alba × P. glandulosa
-
摘要:
目的 :对银腺杨(Populus alba × P. glandulosa‘84k’)组氨酸激酶PagHK3a基因敲除株系的抗旱性进行评价,同时探究PagHK3a基因在杨树响应干旱胁迫过程中的分子调节机制。 方法 :利用5%PEG模拟干旱胁迫处理继代25 d的野生型银腺杨(WT)及其PagHK3a基因敲除株系(C1和C2)组培苗,处理3 h后采用实时定量PCR(Quantitative real-time PCR)方法测定各株系叶片PagHK3a基因、PagHK3a下游基因、干旱胁迫响应基因及抗氧化基因的表达水平;利用称重控水法对各株系盆栽苗进行3个梯度的温室干旱胁迫处理,包括正常浇水(土壤相对含水量:70%~75%)、中度干旱(40%~45%)以及重度干旱(25%~30%),干旱胁迫4周后测定WT及C1、C2株系的瞬时光合参数、过氧化氢(H2O2)及丙二醛(MDA)含量、过氧化物酶(POD)及超氧化物歧化酶(SOD)酶活性、株高和地径等生理生化及生长指标。 结果 :在温室中度干旱条件下,C1和C2株系株高生长量均显著高于WT,而在重度干旱条件下,这两个株系的株高生长量与WT相比均无显著差异。分析光合参数发现,在中度干旱条件下,基因敲除株系C1和C2气孔导度及胞间CO2浓度均显著高于WT,但在重度干旱胁迫条件下只有C2株系胞间CO2浓度显著高于WT;在中度干旱胁迫条件下C1株系瞬时净光合速率显著高于WT,在重度干旱胁迫条件下,C2株系瞬时净光合速率显著高于WT。同时生化指标测定结果显示,在中度干旱条件下基因敲除株系C1和C2中MDA及H2O2含量显著低于WT,而在重度干旱胁迫时,仅C2株系显著低于WT;对比分析各株系抗氧化酶活性发现,正常供水条件下,C1株系叶片POD酶活性以及C2株系SOD酶活性显著高于WT,在中度干旱胁迫条件下,C1株系这两种保护酶活性均显著高于WT,而C2株系仅在重度干旱胁迫下显著高于WT。另外,在PEG模拟干旱胁迫处理下,基因敲除株系C1和C2叶片组氨酸激酶基因PagHK3a及其下游响应调节蛋白PagRR2、PagRR15基因表达量与WT相比均显著下调;而干旱胁迫响应基因PagNAC3以及过氧化物酶合成基因POD1的表达则显著上调,超氧化物歧化酶合成基因SOD4的表达与WT相比无显著差异。 结论 :在PagHK3a基因敲除株系中,PagHK3a基因及细胞分裂素信号传导途径中响应调节因子基因PagRR2、PagRR15表达均较WT显著降低,而胁迫响应基因PagNAC3及POD合成基因POD1表达显著升高;同时,在中度干旱条件下PagHK3a基因敲除株系的气体交换能力更强,MDA以及H2O2含量更低,株高生长量更大,从而具有更强的抗旱能力。 -
关键词:
- 银腺杨
- / 组氨酸激酶基因PagHK3a
- / 基因敲除
- / 抗旱性
Abstract:Objective : This study aimed to evaluate the drought resistance of two histidine kinase PagHK3a gene knockout lines of poplar '84k' (Populus alba × P. glandulosa '84k'), and explore the molecular regulatory mechanism of PagHK3a gene in poplar response to drought stress. Method : Plantlets of wild-type (WT) and gene knockout strains (C1 and C2) sub-cultured for 25 days were treated with 5% PEG and the mature leaves of the plantlets were gathered 3 hours later. Quantitative real-time PCR (qPCR) was used to detect the expression levels of PagHK3a gene, PagHK3a downstream related genes, drought stress response genes and antioxidant related genes in leaves of the three lines. Meanwhile, the drought stress treatment was carried out using the potted plants of each line by weighing method with three levels: normal watering greenhouse (soil relative water content: 70%-75%), moderate drought (40%-45%) and severe drought (25%-30%). After four weeks of drought stress treatment, transient photosynthetic parameters, contents of hydrogen peroxide (H2O2) and malondialdehyde (MDA), activities of peroxidase (POD) and superoxide dismutase (SOD), plant height and ground diameter of WT and C1, C2 were measured. Result : Under moderate drought conditions, the height growth of both C1 and C2 lines was significantly higher than that of WT, but under severe drought conditions, the height growth of both lines was not significantly different from that of WT. Analysis of photosynthetic parameters revealed that the stomatal conductance and intercellular carbon dioxide concentration of C1 and C2 were significantly higher than those of WT under moderate drought condition, but only C2 was significantly higher than that of WT under severe drought stress condition. Under moderate drought stress, the instantaneous net photosynthetic rate of C1 was significantly higher than that of WT, and under severe drought stress, the instantaneous net photosynthetic rate of C2 was significantly higher than that of WT. At the same time, the results of biochemical indexes showed that MDA and H2O2 contents in knockout lines C1 and C2 were significantly lower than those in WT under moderate drought stress, but only C2 was significantly lower than that in WT under severe drought stress. Comparative analysis of the antioxidant enzyme activity of each line found that, under normal water supply conditions, the POD enzyme activity of C1 strain and the SOD activity of C2 strain were significantly higher than that of WT. And under moderate drought stress, the two protective enzyme activities of C1 strain were significantly higher than that of the control, while C2 was significantly higher than that of WT only under severe drought stress. In addition, under 5% PEG treatment, the expression levels of histidine kinase gene PagHK3a and its downstream response regulatory proteins PagRR2 and PagRR15 in the leaves of gene knockout lines C1 and C2 were significantly down-regulated compared with those of WT. The expression levels of drought stress response gene PagNAC3 and peroxidase synthesis gene POD1 were significantly up-regulated, while the expression level of superoxide dismutase synthesis gene SOD4 was not significantly different from that of WT. Conclusion : In PagHK3a knockout lines, expression of PagHK3a was down-regulated, expression of its downstream genes PagRR2 and PagRR15 in the cytokinin signaling pathway was also significant down regulated and the expression of stress response gene PagNAC3 and POD synthesis gene POD1 were significantly up regulated; meanwhile the PagHK3a knockout lines had stronger gas exchange ability, lower MDA and H2O2 content and larger plant height growth under moderate drought conditions, therefore had stronger drought stress resistance than WT. -
图 1 银腺杨及其PagHK3a基因敲除株系C1和C2靶点编辑情况
Figure 1. Schematic illustration of the sgRNA target site in wild type (WT) of P. alba × P. glandulosa and two mutant lines (C1 and C2)
图 2 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT之间株高和地径生长量的差异
Figure 2. Differences in plant height and ground diameter growth between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
图 3 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT生化指标差异
Figure 3. Differences in biochemical indexes between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
图 4 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT之间光合参数差异
Figure 4. Differences in photosynthetic parameters between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
图 5 PEG模拟干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2及WT叶片中相关基因表达水平
Figure 5. Expression levels of genes in leaves of PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa in Hoagland’s nutrient solution with 5% PEG
表 1 实时定量PCR引物序列
Table 1. The primer sequences used for quantitative real-time PCR
基因
Gene引物序列
Primer sequence(5′→3′)产物长度
Product length/bpPagHK3a F: ACCGCACAAGACAGTGGAAA 100 R: ACTGCATTGCCTCAAGCTCA PagRR2 F: CAAACCCTCAAACTGCCCTC 103 R: TCAGCTAGTCCAATGCCTCC PagRR15 F: GCGAAGTTTGGCGGATGAAG 161 R: TGCTCCCACTCTCTACAACAG PagNAC3 F: TCAACTCGGTGCCTCAACTC 161 R: AGCTCCACTTTGAACCTCCC SOD4 F: CAGTGGAGGTGCATTGGAAG 138 R: GCGAAGGGTTGCCATTGTTA POD1 F: TAGCAACATCCCTTCCCCAG 128 R: TGGTACATCTTGCTTGCCCT Actin F: TGCGTATGTTGCCCTTGACT 157 R: TCCGATGAGAGAAGGCTGGA  下载: 导出CSV
下载: 导出CSV
表 2 干旱胁迫下PagHK3a基因敲除银腺杨株系C1、C2与WT之间根干质量差异
Table 2. Differences in root dry weight between PagHK3a gene knockout lines C1, C2 and wild-type of P. alba × P. glandulosa under drought stress
株系
Lines根干质量/g
Root dry weightCK D1 D2
PWT 0.50 ± 0.17 aB 0.64 ± 0.03 aAB 0.84 ± 0.13 aA 0.043 C1 0.48 ± 0.05 aC 0.67 ± 0.12 aB 0.92 ± 0.09 aA 0.003 C2 0.56 ± 0.05 aB 0.71 ± 0.03 aA 0.77 ± 0.09 aA 0.014 P 0.687 0.567 0.276 -- 注:表格中数据表现格式为均值 ± 标准差,其后大写字母表示为同一株系在不同处理条件下的差异显著性,小写字母表示为同一种处理方式下不同种株系之间的差异显著性,字母相同表示差异不显著,字母不同表示差异显著,显著性水平为0.05 Notes: The data in the table are presented in the format of mean ± standard deviation, followed by capital letters indicating the significance of the difference between the same strain under different treatment conditions, and lowercase letters indicating the significance of the difference between different strains under the same treatment, with the same letter indicating that the differences are not significant, and different letters indicating that the differences are significant, with a significance level of 0.05
下载: 导出CSV
-
[1] JIA H, ZHANG J, LI J,et al. Genome-wide transcriptomic analysis of a desert willow, Salix psammophila, reveals the function of hub genes SpMDP1 and SpWRKY33 in drought tolerance[J]. BMC Plant Biology, 2019, 19(1): 356. doi: 10.1186/s12870-019-1900-1 [2] MCDOWELL N, ALLEN C D, ANDERSON‐TEIXEIRA K,et al. Drivers and mechanisms of tree mortality in moist tropical forests[J]. New Phytologist, 2018, 219(3): 851-869. doi: 10.1111/nph.15027 [3] SINGH D, SINGLA-PAREEK S, PAREEK A. Two-component signaling system in plants: interaction network and specificity in response to stress and hormones[J]. Plant Cell Reports, 2021, 40(11): 2037-2046. doi: 10.1007/s00299-021-02727-z [4] HWANG I, CHEN H C, SHEEN J. Two-component signal transduction pathways in Arabidopsis[J]. Plant Physiology, 2002, 129(2): 500-515. doi: 10.1104/pp.005504 [5] WEST A H, STOCK A M. Histidine kinases and response regulator proteins in two-component signaling systems[J]. Trends in Biochemical Sciences, 2001, 26(6): 369-376. doi: 10.1016/S0968-0004(01)01852-7 [6] SCHALLER G E, KIEBER J J, SHIU S H. Two-component signaling elements and histidyl-aspartyl phosphorelays[J]. The Arabidopsis Book, 2008, 2008(6): 112. [7] HWANG I, SHEEN J. Two-component circuitry in Arabidopsis cytokinin signal transduction[J]. Nature, 2001, 413(6854): 383-389. doi: 10.1038/35096500 [8] INOUE T, HIGUCHI M, HASHIMOTO Y,et al. Identification of CRE1 as a cytokinin receptor from Arabidopsis[J]. Nature, 2001, 409(6823): 1060-1063. doi: 10.1038/35059117 [9] PAVLŮ J, NOVÁK J, KOUKALOVÁ V,et al. Cytokinin at the crossroads of abiotic stress signalling pathways[J]. International Journal of Molecular Sciences, 2018, 19(8): 2450. doi: 10.3390/ijms19082450 [10] KUMAR M N, VERSLUES P E. Stress physiology functions of the Arabidopsis histidine kinase cytokinin receptors[J]. Physiologia Plantarum, 2015, 154(3): 369-380. doi: 10.1111/ppl.12290 [11] KANG N Y, CHO C, KIM N Y,et al. Cytokinin receptor-dependent and receptor-independent pathways in the dehydration response of Arabidopsis thaliana[J]. Journal of Plant Physiology, 2012, 169(14): 1382-1391. doi: 10.1016/j.jplph.2012.05.007 [12] NISHIYAMA R, WATANABE Y, LEYVA-GONZALEZ M A,et al. Arabidopsis AHP2, AHP3, and AHP5 histidine phosphotransfer proteins function as redundant negative regulators of drought stress response[J]. Proceedings of the National Academy of Sciences, 2013, 110(12): 4840-4845. doi: 10.1073/pnas.1302265110 [13] NGUYEN K H, HA C V, NISHIYAMA R,et al. Arabidopsis type B cytokinin response regulators ARR1, ARR10, and ARR12 negatively regulate plant responses to drought[J]. Proceedings of the National Academy of Sciences, 2016, 113(11): 3090-3095. doi: 10.1073/pnas.1600399113 [14] 主楚杰, 王爱云. 基因工程技术在杨树抗逆境方面的研究进展[J]. 江苏农业科学, 2015, 43(2):10-13. [15] 李义良, 苏晓华, 张冰玉, 等. 外源SacB基因在银腺杂种杨基因组中的表达及抗旱性分析[J]. 北京林业大学学报, 2007, 29(2):1-6. [16] 杨春霞, 李火根, 程 强, 等. 南林 895 杨抗旱耐盐基因DREB1C的转化[J]. 林业科学, 2009, 45(2):17-21. [17] 张伟溪, 刘浡洋, 丁昌俊, 等. 欧美杨锌指蛋白转录因子基因(ZxZF)的遗传转化及抗旱性初步分析[J]. 林业科学, 2014, 50(3):31-37. [18] NIEMINEN K, IMMANEN J, LAXELL M,et al. Cytokinin signaling regulates cambial development in poplar[J]. Proceedings of the National Academy of Sciences, 2008, 105(50): 20032-20037. doi: 10.1073/pnas.0805617106 [19] 鲁俊倩, 武 舒, 钟姗辰, 等. ‘84K’杨组氨酸激酶基因 PaHK3a 的表达及功能分析[J]. 北京林业大学学报, 2021, 43(2):46-53. [20] 鲁俊倩, 武 舒, 钟姗辰, 等. ‘84K’杨组氨酸激酶基因 PaHK3b 的克隆及功能分析[J]. 林业科学研究, 2021, 34(1):26-34. [21] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method[J]. Methods, 2001, 25(4): 402-408. doi: 10.1006/meth.2001.1262 [22] QURESHI M K, MUNIR S, SHAHZAD A N,et al. Role of reactive oxygen species and contribution of new players in defense mechanism under drought stress in rice[J]. Int J Agric Biol, 2018, 20(6): 1339-1352. [23] FOYER C H, NOCTOR G. Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses[J]. The Plant Cell, 2005, 17(7): 1866-1875. doi: 10.1105/tpc.105.033589 [24] 张永强, 毛学森, 孙宏勇, 等. 干旱胁迫对冬小麦叶绿素荧光的影响[J]. 中国生态农业学报, 2002, 10(4):13-15. [25] SHAKEEL A A, XIAO-YU X, LONG-CHANG W,et al. Morphological, physiological and biochemical responses of plants to drought stress[J]. African Journal of Agricultural Research, 2011, 6(9): 2026-2032. [26] 杨佳骏, 吴永波, 张燕红. 高温与干旱胁迫对‘南林895杨’扦插苗生长和超微结构的影响[J]. 林业科学, 2020, 56(5):176-183. doi: 10.11707/j.1001-7488.20200520 [27] AHMED U, RAO M J, QI C,et al. Expression profiling of flavonoid biosynthesis genes and secondary metabolites accumulation in populus under drought stress[J]. Molecules, 2021, 26(18): 5546. doi: 10.3390/molecules26185546 [28] MEDRANO H, ESCALONA J M, BOTA J,et al. Regulation of photosynthesis of C3 plants in response to progressive drought: stomatal conductance as a reference parameter[J]. Annals of Botany, 2002, 89(7): 895-905. doi: 10.1093/aob/mcf079 [29] 温 琦, 赵文博, 张幽静, 等. 植物干旱胁迫响应的研究进展[J]. 江苏农业科学, 2020, 48(12):11-15. doi: 10.15889/j.issn.1002-1302.2020.12.003 [30] MULLER B, SHEEN J. Arabidopsis cytokinin signaling pathway[J]. Science’s STKE, 2007, 2007(407): 1-4. [31] TRAN L S P, URAO T, QIN F,et al. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis[J]. Proceedings of the National Academy of Sciences, 2007, 104(51): 20623-20628. doi: 10.1073/pnas.0706547105 [32] KIM H J, RYU H, HONG S H,et al. Cytokinin-mediated control of leaf longevity by AHK3 through phosphorylation of ARR2 in Arabidopsis[J]. Proceedings of the National Academy of Sciences, 2006, 103(3): 814-819. doi: 10.1073/pnas.0505150103 [33] TRAN L S P, NAKASHIMA K, SAKUMA Y,et al. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter[J]. The Plant Cell, 2004, 16(9): 2481-2498. doi: 10.1105/tpc.104.022699 [34] GRAHAM N, PATIL G B, BUBECK D M,et al. Plant genome editing and the relevance of off-target changes[J]. Plant Physiology, 2020, 183(4): 1453-1471. doi: 10.1104/pp.19.01194 [35] BELHAJ K, CHAPARRO-GARCIA A, KAMOUN S,et al. Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system[J]. Plant Methods, 2013, 9(1): 39. doi: 10.1186/1746-4811-9-39 [36] ZHANG H, ZHANG J, WEI P,et al. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation[J]. Plant Biotechnology Journal, 2014, 12(6): 797-807. doi: 10.1111/pbi.12200 [37] ZHANG Q, XING H L, WANG Z P,et al. Potential high-frequency off-target mutagenesis induced by CRISPR/Cas9 in Arabidopsis and its prevention[J]. Plant Molecular Biology, 2018, 96(4/5): 445-456. [38] NEELAKANDAN A K, WANG K. Recent progress in the understanding of tissue culture-induced genome level changes in plants and potential applications[J]. Plant Cell Reports, 2012, 31(4): 597-620. doi: 10.1007/s00299-011-1202-z -

 点击查看大图
点击查看大图

计量
- 文章访问数: 2431
- HTML全文浏览量: 1203
- PDF下载量: 102
- 被引次数: 0
