





-
金银花学名忍冬(Lonicera japonica Thunb.),是一种重要的药用植物,具有广泛的药理作用,如抗菌、抗炎、抗病毒、抗内毒素、减脂、解热等[1-2]。金银花在国内分布十分广泛,其种植区域主要集中在山东、河北和河南等省份,其中河北省巨鹿县为全国金银花三大主产地(河北巨鹿、河南封丘、山东平邑)之首,成为当地的支柱产业,获得“中国金银花之乡”的称号[3-4]。
金银花叶部病害是田间生产的巨大障碍,主要有褐斑病(Cercospora rhamni Fuckel)、白粉病(Microsphaera linicerae Wint. inRabenh)等。其中,金银花褐斑病对金银花的产量与质量有严重影响。此前,有报道认为金银花褐斑病的病原菌是鼠李尾孢(C. rhamni)[5-6],也有研究认为是多主棒孢霉(Corynespora cassiicola (Berk. & M.A. Curtis) C.T. Wei)[7-8];2014年,de Miranda等人的研究表明假尾孢属真菌(Pseudocercospora lonicerigena U. Braun & Crous)是引起巴西金银花褐斑病的病原菌[9];2016年张永信等人的研究发现拟茎点霉属真菌(Phomopsis sp.)也可以使金银花产生褐斑病 [10]。
2014年到2018年期间,笔者在研究河北金银花褐斑病病原过程中,总能分离到一种产棕黄色或红色色素的真菌,其分出率约为17%。利用离体叶片致病性接种时,该菌产生的病斑与褐斑病不符,并依据ITS序列和形态特征将其初步鉴定为匍柄霉(Stemphylium sp.)。针对这种金银花新型叶斑病,本文依照科赫氏法则对其病原菌进行了致病性测定,并结合形态学和多基因片段分析对其进行了分类鉴定,并将该病害命名为金银花黑斑病,以期为该病害的有效防控提供理论基础。
-
本项研究所用金银花病叶样采集自河北省巨鹿县和廊坊市。样本利用自封袋包装并按野外样本采集要求记录采集时间、地点、采集人、环境条件等要素后,带回实验室用于病原菌分离鉴定。
病原菌的分离采用常规组织分离法。首先将采集的叶片用无菌水冲洗干净,用剪刀将病叶的病健交界处剪为4 mm × 4 mm的小组织块,于超净工作台上用75%酒精浸泡30 s;随后用无菌水淋洗,再用0.1%升汞溶液浸泡2 min,无菌水洗3次;用无菌滤纸吸去叶组织块表面的水分后,再将其摆放至PDA培养基(马铃薯200 g、琼脂20 g、葡萄糖20 g、蒸馏水1 L)平板上进行分离培养。每个平板均匀摆放6个组织块,于25 ℃下恒温培养24 h后,挑取已长出菌落的边缘菌丝进行菌株的纯化,从而获得纯菌株。
-
将纯化后的菌株在PDA培养基上培养4 d后,分别转接到新的PDA和PCA(马铃薯和胡萝卜各100 g, 葡萄糖20 g,琼脂20 g,蒸馏水1 L)培养基上,于25 ℃下恒温培养7 d,观察并记录菌落的形状、颜色、光泽度、质地等培养特征。分别采用番茄快速诱导产孢法和双玻片插片诱导产孢法诱导该菌株产孢[11-12],菌株的显微特征主要包括菌株分生孢子的大小、形状,分生孢子梗的形状等特征,并拍照。
番茄快速诱导产孢法:将接种于PDA上的匍柄霉菌在恒温培养箱中按照12 h光照12 h黑暗光周期在温度25 ℃下培养至菌丝长满培养皿;取菌丝生长边缘的匍柄霉菌片接种到经过催芽处理新长出的番茄叶片上,在温度25 ℃、相对湿度60%~80%与12小时光照12小时黑暗光周期的条件下培养3 d,挑取匍柄霉菌片表面在显微镜下观察。
双玻片插片诱导产孢法:于25 ± 1 ℃培养箱中恒温培养3~4 d后,取3~5个平板于菌落边缘1~2 cm处以30°~45°角斜插入洁净的双层重叠的盖玻片,然后放置 25 ± 1 ℃培养箱恒温培养3~4周左右。用镊子取出诱导培养3~4周后的双层玻片,并轻轻分成2个单层玻片,单层玻片的正面载有菌体,在显微镜下观察。
-
所获得的纯菌株于PDA培养基上培养10 d后,刮取少量菌丝置于2 mL离心管中,采用改良的CTAB法提取本研究所用菌株的DNA[13],利用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)/ITS4(5′-TCCTCCGCTTATTGATATGC-3′)、引物gpd1(5′-CAACGGCTTCGGTCGCATTG-3′)/gpd2(5′-GCCAAGCAGTTGGTTGTGC-3′)和引物EF446F(5′-TCACTTGATCTACAAGTGCGGTGG-3′)/EF1473R(5′-CGATCTTGTAGA CAT CCTGGAGG-3′)分别扩增核糖体内转录间隔区(ITS)、3-磷酸甘油脱氢酶(gpd)和蛋白延伸因子(EF-1α)基因片段[14-16]。ITS基因片段反应体系为(25 μL):12.5 μL 2 × Taq PCR Master Mix(天根生化科技(北京)有限公司),9.5 μL ddH2O, 1 μL ITS1, 1 μL ITS4和1 μL DNA 模板。gpd基因片段反应体系(25 μL):12.5 μL 2 × Taq PCR Master Mix,9.5 μL ddH2O, 1 μL gpd1, 1 μL gpd2和1 μL DNA模板。EF-1α基因片段反应体系为(25 μL):12.5 μL 2 × Taq PCR Master Mix,9 μL ddH2O, 1 μL EF446F, 1 μL EF1473R和1.5 μL DNA模板。ITS序列PCR反应程序为:94 ℃预变性5 min,94 ℃变性30 s,54 ℃退火 30 s,72 ℃延伸 30 s,35个循环,最后72 ℃延伸10 min,4 ℃保存。gpd序列PCR反应程序为:94 ℃预变性3 min,94 ℃变性30 s,57 ℃退火 1 min,72 ℃延伸 1.5 min,35个循环,最后72 ℃延伸10 min,4 ℃保存。EF-1α序列反应程序为:94 ℃预变性1 min,94 ℃变性30 s,60 ℃退火 1 min,72 ℃延伸 3 min,35个循环,最后72 ℃延伸5 min,4 ℃保存。PCR扩增产物于1.2%琼脂糖凝胶电泳检测后,送至北京擎科生物公司进行测序。
测序所获得的拼接序列在NCBI中进行比对,确定菌株分类地位,并从GenBank中下载相似性高的序列或近缘物种序列作为参考序列(表1)。使用MEGA 10.2.2软件[17]进行序列对齐和剪切,以ITS-gpd-EF-1α的顺序进行序列串联,并分别利用最大似然法(ML)、贝叶斯法(BI)和最大简约法(MP)进行系统发育树的构建,外群为互隔链格孢菌Alternaria alternata EGS34-016。
表 1 用于分子系统发育分析的参考菌株信息
Table 1. The information of strains used for phylogenetic analysis
种名
Species菌株
Strains登录号 GenBank Accession No. ITS gpd EF-1α Pleospora herbarum EGS 36-138.2 AF442785 AF443884 P. gigaspora EGS37-016 AY329174 AY316975 AY324678 P. tarda EGS 04-118c AF442782 AF443881 P. tarda NO 537 AF443879 AF442780 P. triglochinicola EGS 36-118 AF443901 AF442802 AY324753 Stemphylium vesicarium EGS29-089 AY329229 AY317033 AY324711 S. vesicarium EGS 37-067 AF442803 AF443902 AY324708 S. majusculum EGS 29-094 AF442792 AF443891 S. alfalfae EGS 36-088 AF442775 AF443874 S. majusculum EGS16-068 AY329228 AY317032 AY324710 S. alfalfae EGS 39-127 AF442774 AF443873 S. astragali EGS29-062 AY329181 AY316984 AY324684 S. gracilariae EGS37-073 AY329217 AY317021 AY324709 S. astragali EGS 27-194.1 AF442777 AF443876 S. eturmiunum EGS29-099 AY329230 AY317034 AY324712 S. subglobuliferum HSAUP XF0140 AY751454 AY751459 S. xanthosomatis EGS 17-137 AF442804 AF443903 AY324758 S. xanthosomatis EGS47-197 AY329223 AY317027 AY324764 S. lycopersici EGS 46-001 AF442790 AF443889 S. lycopersici EGS45-036 AY329225 AY317029 AY324766 S. lycopersici EGS49-043 AY329256 AY317060 AY324770 S. callistephi EEB 1055 AF229482 AY278822 JQ672393 S. callistephi NO 536 AF442783 AF443882 AY324750 S. solani EGS 42-027 AF442797 AF443896 S. solani EGS41-135 AY329214 AY317018 AY324759 S. lancipes CBS 719.68 GQ395367 GQ395373 S. lancipes EGS46-182 AY329203 AY317007 AY324742 S. paludiscirpi EGS31-016 AY329231 AY317035 AY324769 S. trifolii EGS12-142 AY329218 AY317022 S. trifolii NO 712 AF442800 AF443899 S. drummondii CBS 346.83 GQ395365 KU850687 S. loti NO 1364 AF442788 AF443887 AY324776 S. loti NO 770 AF442789 AF443888 AY324775 S. sarciniforme EGS29-188 AY329221 AY317025 AY324746 S. sarciniforme EGS 38-121 AF442793 AF443892 AY324743 S. botryosum EGS08-069 AY329168 AY316968 AY324671 Alternaria alternata EGS34-016 AY751456 AY278808 AH013339 -
本研究分别对金银花和番茄的叶片进行致病性接种实验。
金银花叶片接种:采用刺伤接种菌丝块的方法对健康的金银花活体叶片进行致病性测定。首先将纯菌株培养4~5 d后打取菌落边缘直径为5 mm的菌饼;用75%酒精对叶片进行表面消毒,用接种针刺伤接种部位,菌饼接种于刺伤处,以无菌PDA琼脂块接种作为对照,每组处理设10次重复。接种后于25 ℃下恒温放置9 d,观察并记录发病状况。随后,通过常规组织分离法,取病健交界处进行病原菌的再次分离和纯化;通过菌株菌落形态结合ITS序列,确定菌株是否与接种菌株相同。
番茄叶片接种:采用刺伤接种菌丝块的方法对健康的番茄离体叶片进行致病性测定,具体步骤同上。接种后于25 ℃下恒温放置48 h,观察并记录发病情况,并进行病原菌的再次分离、纯化和复核鉴定。
-
在田间调查和实验中,该病害主要危害金银花的叶片,发病初期(5—8月份),通常在叶片边缘产生芝麻粒大小的灰黑色小斑点。9月份进入发病后期,黑色病斑逐渐扩大,有时多个小斑扩大后融合在一起,呈不规则状。不同于褐斑病,该病斑能横跨叶脉,后期数个小斑融合在一起,发病严重时,叶片枯黄脱落(图1)。

图 1 金银花黑斑病不同时期的危害症状
Figure 1. Symptoms of the black spot disease in different periods on Lonicera japonica
-
对采集到的金银花叶片样品进行分离培养,分离纯化后得到的纯菌株在PDA培养基上呈现出相同的菌落,经过ITS序列比对,初步确定所分离得到的菌株属于同一种。挑选其中1株(YP4)作为代表菌株进行后续形态学观察和系统发育学分析。
-
供试菌株YP4在PDA培养基上培养7 d后,形成表面为灰白色或黄白色、棉絮状的菌落。菌落呈同心环状,中心为灰白色,外围白色发黄,并在PDA培养基上产生棕黄色至深红色色素。经过番茄快速诱导产孢法和双玻片插片诱导产孢法诱导后,在显微镜下未观察到其分生孢子及分生孢子梗,仅观察到其菌丝。该菌株的菌丝光滑,直或弯曲,有分隔,具叉状分枝,直径约为1.8~2.5 μm(图2)。
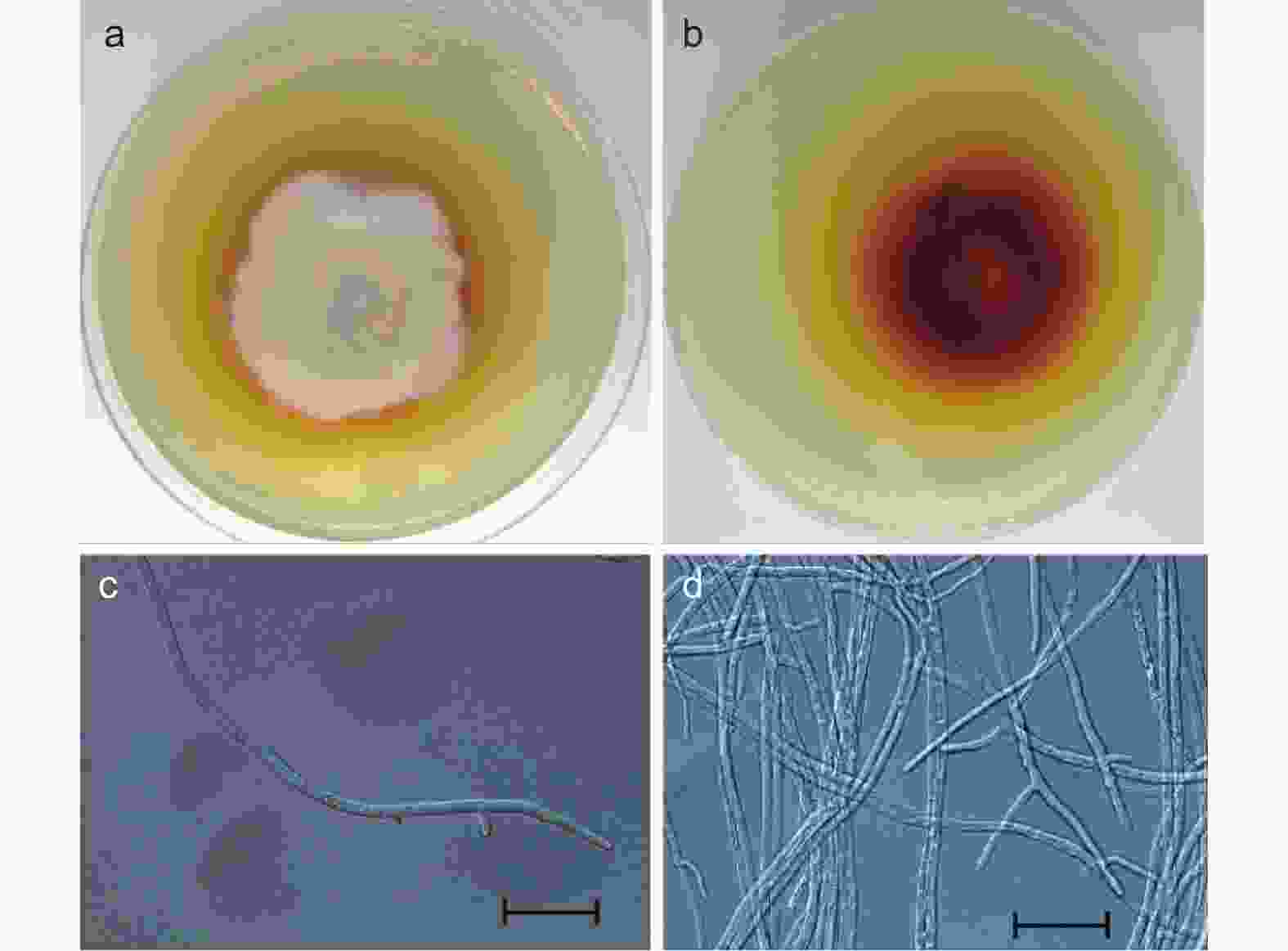
图 2 菌株YP4的形态特征
Figure 2. Morphological characteristics of the strain YP4
-
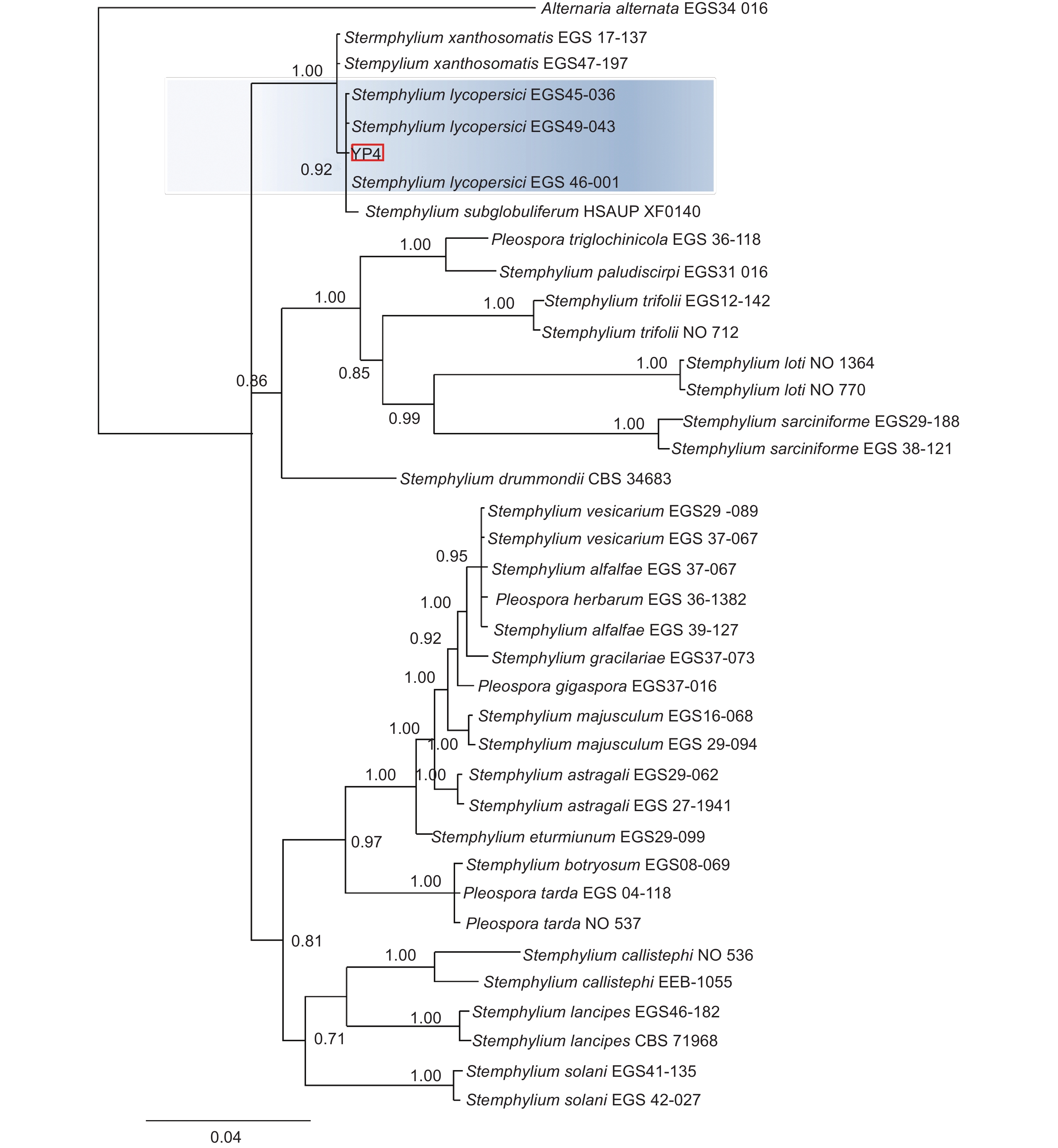
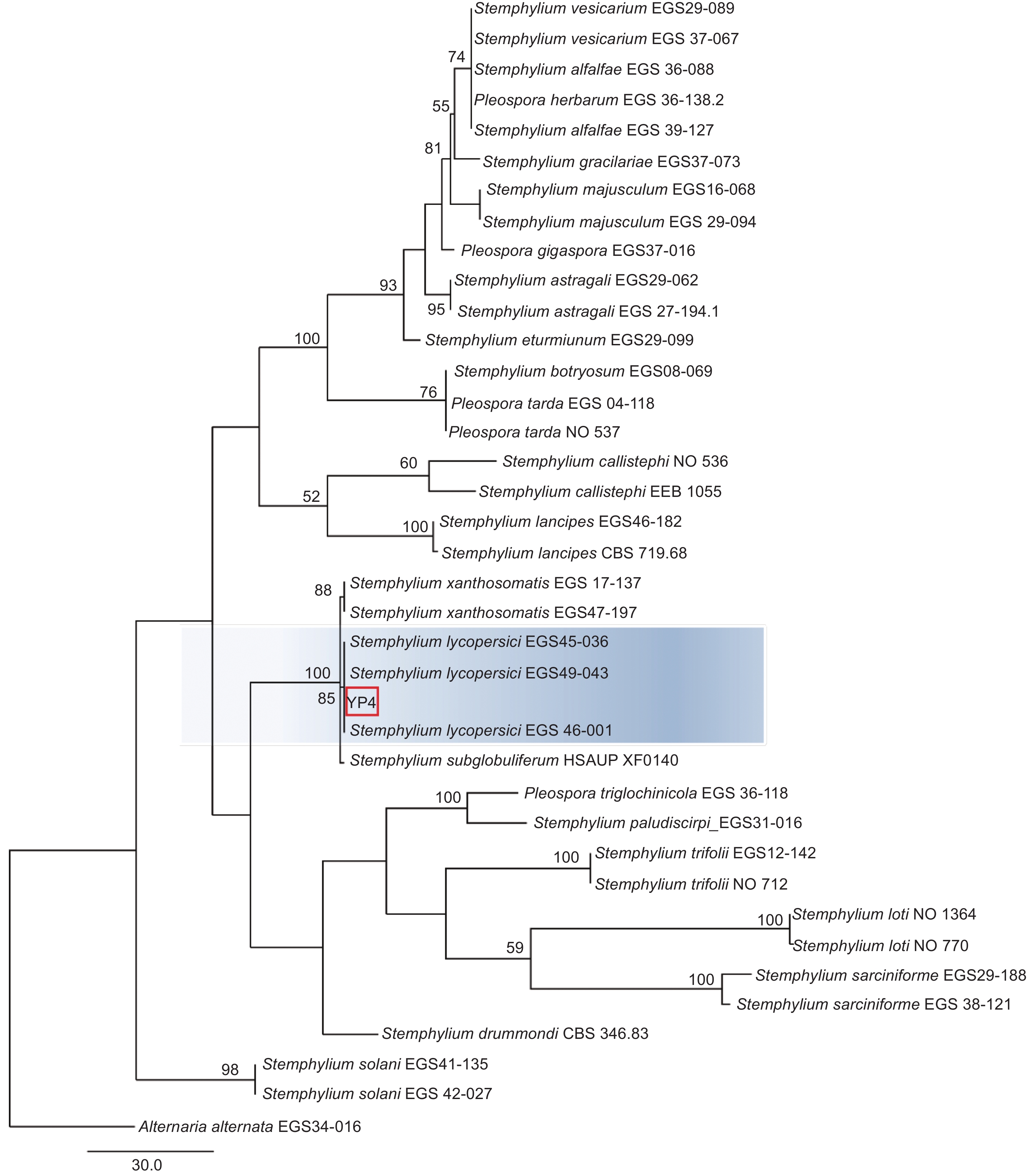
本研究基于ITS、gpd和EF-1α 3个基因片段利用最大似然法(ML)、贝叶斯法(BI)和最大简约法(MP)分别构建了匍柄霉属的系统发育树(图3~5),3个树中包含1个供试菌株YP4和36个从Genbank库中下载的参考菌株,以及1个外类群菌株Alternaria alternata。ITS、gpd和EF-1α 3个基因获得扩增片段大小分别为530、533和674 bp(GenBank登录号OQ157590、OQ186451和OQ185385),串联后总长度为1737 bp,菌株YP4与番茄匍柄霉Stemphylium lycopersici (EGS45-036、EGS45-043) 的参考序列进行多重比对时的一致性均为100%。并在3种方法构建的系统发育树(图3~5)中,菌株YP4均与番茄匍柄霉S. lycopersici同源性最高,在ML树中以86%的支持率聚为一支,在BI树中以0.92的支持率聚为一支,在MP树中以85%的支持率聚为一支。
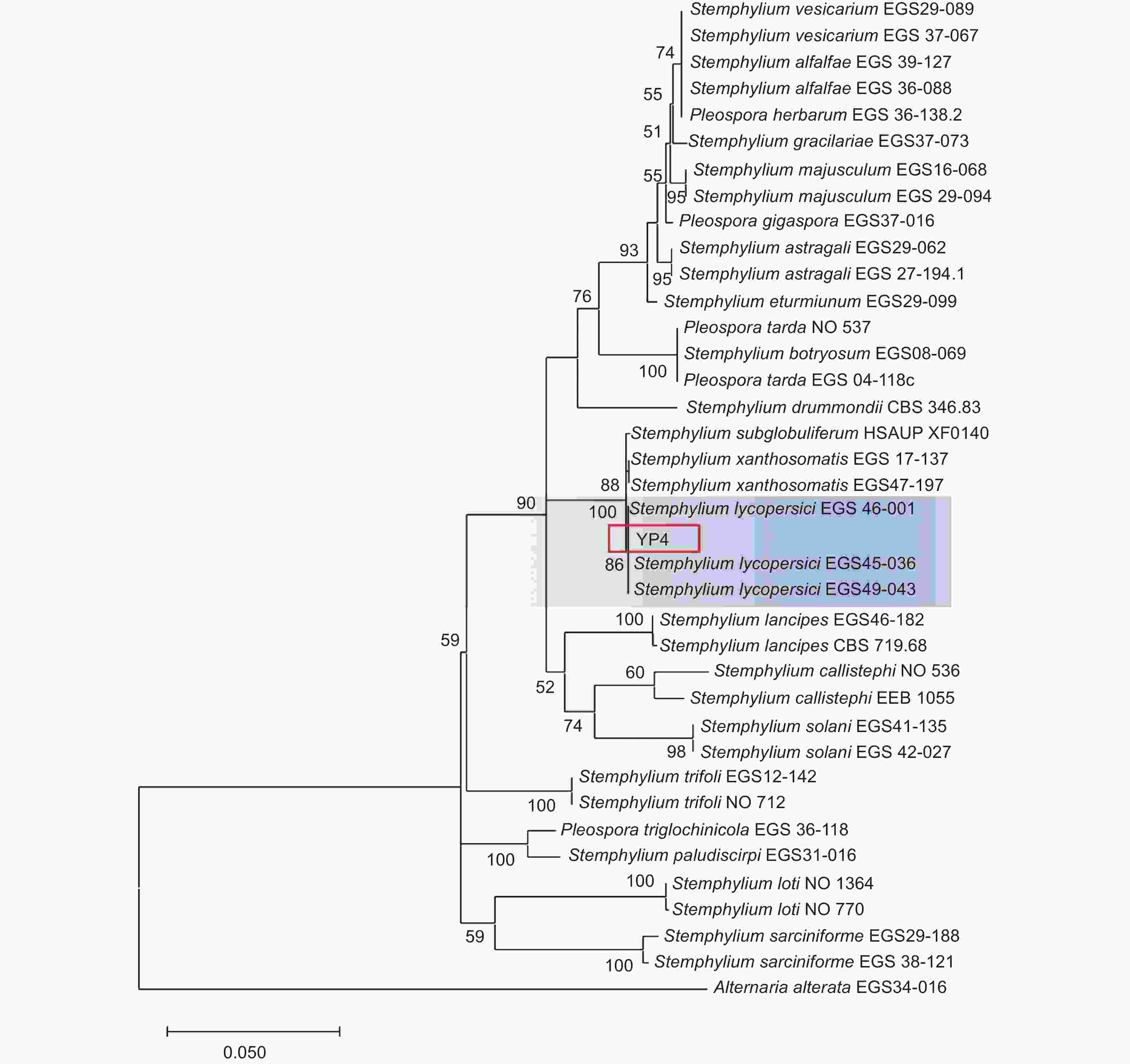
图 3 基于最大似然法(ML) ITS、gpd和EF-1α联合基因系统发育树构建
Figure 3. Maximum likelihood phylogenetic tree based on combined datasets of ITS,gpd and EF-1α
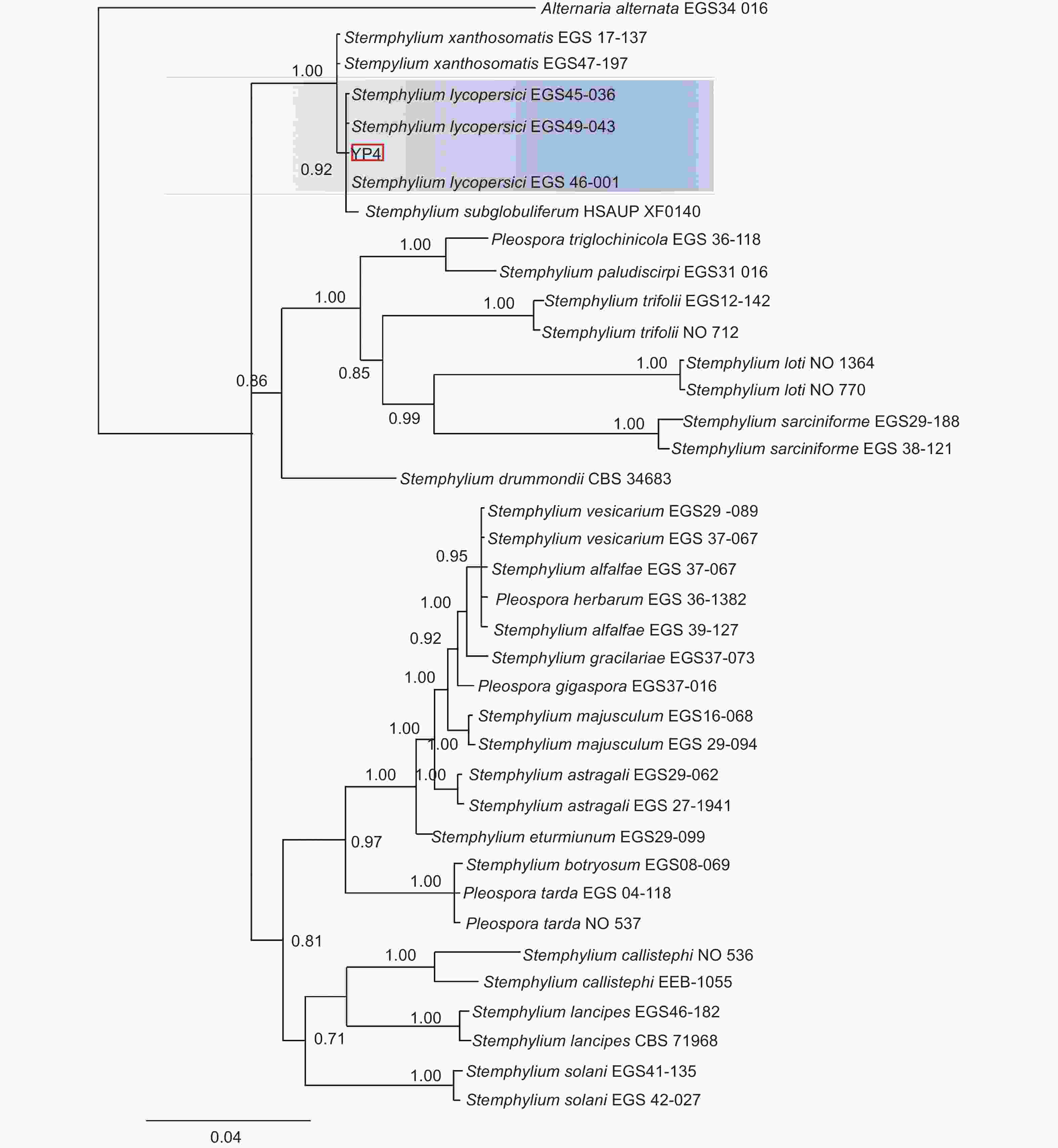
图 4 基于贝叶斯法(BI)ITS、gpd和EF-1α联合基因系统发育树构建
Figure 4. Bayesian phylogenetic tree based on combined datasets of ITS,gpd and EF-1α
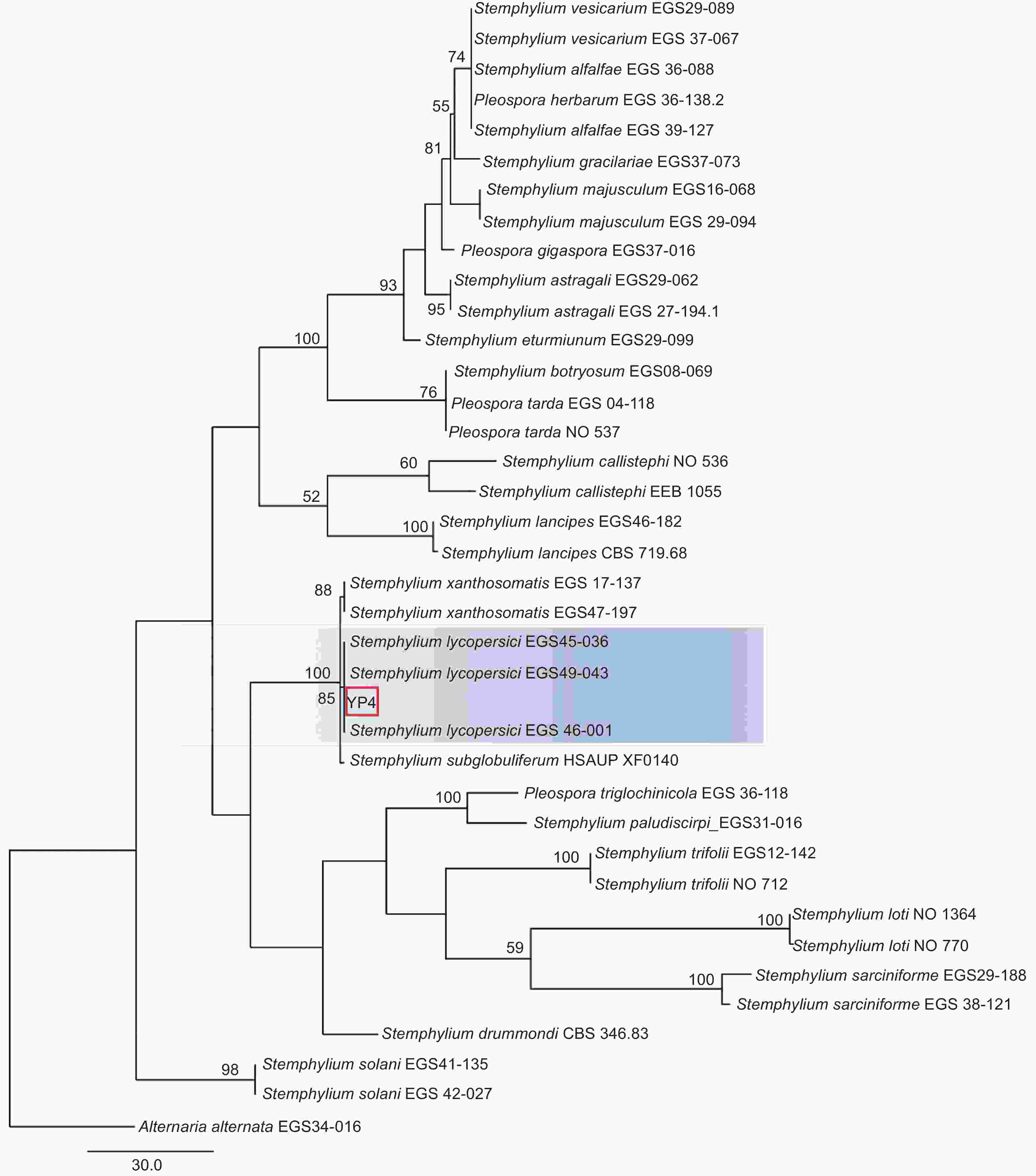
图 5 基于最大简约法(MP)ITS、gpd和EF-1α联合基因系统发育树构建
Figure 5. Phylogenetic tree (MP) based on combined datasets of ITS,gpd and EF-1α
-
在对金银花活体叶片接种9 d后,叶片上产生病斑,呈不规则状,边界不清晰,横跨叶脉,并产生灰白色霉层,而对照组叶片无明显症状(图6)。根据柯赫氏法则,将接种叶片进行再分离,分离得到的菌株的形态特征与接种所用菌株相同。

图 6 致病性接种实验金银花叶片症状
Figure 6. Symptoms of leaves of Lonicera japonica after inoculation of the strain YP4
在对番茄离体叶片接种48 h后,接种叶片上产生黑色病斑,呈不规则状,边界不清晰,横跨叶脉。而对照组叶片无明显症状(图7)。从表现症状的番茄离体叶片病斑上均能再次分离得到所接种的真菌菌株。

图 7 致病性接种实验番茄叶片症状
Figure 7. Symptoms of leaves of Solanum lycopersicum after inoculation of the strain YP4
-
本研究中所获得的菌株在系统发育学上与番茄匍柄霉聚为一支,在形态学上虽未能观察到该菌株的分生孢子和分生孢子梗,但其菌落形态与番茄匍柄霉相同,并且将该菌株接种到金银花叶片和番茄叶片上均可引起明显症状。因此,本研究结合形态学、系统发育学以及致病性分析,将引起河北省金银花黑斑病的病原菌鉴定为番茄匍柄霉(Stemphylium lycopersici (Enjoji) Yamamoto)。
由番茄匍柄霉引起的金银花黑斑病与金银花褐斑病相比有很大的不同:褐斑病的症状为发病初期在叶上形成褐色小点,后扩大成褐色圆斑或不规则病斑,病斑边缘受叶脉限制呈多棱状[17]。而这种黑斑病初期为芝麻状小黑斑,后期扩大为不规则状黑斑,并且能够横跨叶脉。
番茄匍柄霉隶属匍柄霉属(Stemphylium Wallr.),该属是1883年由Wallroth以簇孢匍柄霉(S. botryosum)为模式种创立的。本属内大部分种类通过产生分生孢子进行无性繁殖,有一些种有性型为格孢腔菌属 (Pleospora Rabenh. ex Ces. & De Not.),隶属于格孢腔菌目 (Pleosporales),格孢腔菌科 (Pleosporaceae)。匍柄霉属包括多种腐生真菌及病原真菌,其中病原真菌可危害包括多种农作物在内的寄主植物。匍柄霉属在形态上与其相近属链格孢属(Alternaria Nees)相似,它们的分生孢子都是多隔且着色的,形成于由菌丝产生的分生孢子团上,而两属的区别在于匍柄霉属可在菌丝顶端的分生孢子团上产生连续的分生孢子[16, 18-19]。匍柄霉属中的番茄匍柄霉也是一种世界性分布的重要植物病原菌,能够引起番茄、辣椒、茄子、莴苣和人参果等多种寄主植物产生叶斑病,于1931年被Enjoji从日本的番茄叶片上被首次分离出来[20-23]。有研究表明,该种匍柄霉通过合成和释放植物毒性次生代谢物破坏植物细胞,从而导致植物发病[24]。在中国,番茄匍柄霉于2001年首次在山东省泰安市被发现,其寄主植物为番茄[18];随后,在我国多次发现番茄匍柄霉引起番茄产生灰叶斑病[25-27];此外,在国内番茄匍柄霉也可危害茄子、辣椒、柑橘、向日葵等经济作物的叶部[28-30],还有研究发现甜瓜根部病害也与番茄匍柄霉有关[31]。而本研究是番茄匍柄霉在我国危害金银花的首次报道,进一步拓宽了该病菌的寄主范围。
-
本研究对采自河北省金银花样品进行了分离与纯化,结合形态学和多基因片段分析对其病原菌进行了分类鉴定,并依照科赫氏法则进行了致病性测定,发现了一种由番茄匍柄霉(Stemphylium lycopersici )引起的金银花新型叶斑病,并将该病害命名为金银花黑斑病,为金银花病害的科学防控提供了一定的理论依据。
金银花黑斑病病原菌番茄匍柄霉的分离与鉴定
Isolation and Identification of Pathogenic Fungus Stemphylium lycopersici Causing the Black Spot Disease on Honeysuckle (Lonicera japonica Thunb.)
-
摘要:
目的 分离并鉴定引起金银花黑斑病的病原菌,为该病害的有效防控提供理论依据。 方法 采用常规组织分离法对发病金银花叶片进行分离,结合形态学、多基因序列分析(ITS、gpd和EF-1α序列)和致病性测定进行病原物种鉴定。 结果 在形态学上,分离获得的菌株YP4在PDA培养基上的菌落呈灰白或黄白色棉毛状,与番茄匍柄霉的菌落特征相符;在基于ITS、 gpd和EF-1α序列联合构建的3种系统发育树中,供试菌株YP4均以较高支持率与番茄匍柄霉(Stemphylium lycopersici)聚在同一分支上;致病性接种实验结果表明,菌株YP4接种至金银花叶片上和番茄叶片上均可产生病斑,并可再分离获得其纯菌株;因此,结合形态学、分子系统发育学以及致病性测定,该病原真菌被鉴定为番茄匍柄霉。 结论 本研究明确了引起河北省金银花黑斑病的病原菌为番茄匍柄霉,这是该病菌在我国危害金银花的首次报道。 Abstract:Objective To clarify the pathogen causing black spot disease on Lonicera japonica, and to provide a theoretical basis for effective prevention and control of this disease. Method The strains of the pathogen were isolated by conventional tissue plate method, and were identified by the morphological characters and phylogenetic analysis of ITS, gpd and EF-1α sequences. Its pathogenicity was tested through in vitro inoculating healthy leaves of L. japonica and Solanum lycopersicum. Result The isolated fungal strain YP4 morphologically formed greyish to yellowish white, cottony colonies on Potato Dextrose Agar, which were consistent with published descriptions of Stemphylium lycopersici. In the three phylogenetic trees based on ITS, gpd and EF-1α sequences, the isolate YP4 fell into the S. lycopersici clade with strong support. Based on the pathogenicity measurement of Koch's rule, the strain YP4 was pathogenic to leaves of L. japonica and S. lycopersicum. Conclusion Combined with the morphological characteristics, the pathogenicity determination and phylogenetic analysis of ITS, gpd and EF-1α sequences, the pathogen causing black spot disease on honeysuckle is identified as S. lycopersici. To our knowledge, this is the first report of S. lycopersici causing black spot of L. japonica in China. -
Key words:
- Lonicera japonica Thunb.
- / black spot
- / Stemphylium lycopersici
-
图 1 金银花黑斑病不同时期的危害症状
Figure 1. Symptoms of the black spot disease in different periods on Lonicera japonica
图 3 基于最大似然法(ML) ITS、gpd和EF-1α联合基因系统发育树构建
Figure 3. Maximum likelihood phylogenetic tree based on combined datasets of ITS,gpd and EF-1α
图 4 基于贝叶斯法(BI)ITS、gpd和EF-1α联合基因系统发育树构建
Figure 4. Bayesian phylogenetic tree based on combined datasets of ITS,gpd and EF-1α
图 5 基于最大简约法(MP)ITS、gpd和EF-1α联合基因系统发育树构建
Figure 5. Phylogenetic tree (MP) based on combined datasets of ITS,gpd and EF-1α
图 6 致病性接种实验金银花叶片症状
Figure 6. Symptoms of leaves of Lonicera japonica after inoculation of the strain YP4
图 7 致病性接种实验番茄叶片症状
Figure 7. Symptoms of leaves of Solanum lycopersicum after inoculation of the strain YP4
表 1 用于分子系统发育分析的参考菌株信息
Table 1. The information of strains used for phylogenetic analysis
种名
Species菌株
Strains登录号 GenBank Accession No. ITS gpd EF-1α Pleospora herbarum EGS 36-138.2 AF442785 AF443884 P. gigaspora EGS37-016 AY329174 AY316975 AY324678 P. tarda EGS 04-118c AF442782 AF443881 P. tarda NO 537 AF443879 AF442780 P. triglochinicola EGS 36-118 AF443901 AF442802 AY324753 Stemphylium vesicarium EGS29-089 AY329229 AY317033 AY324711 S. vesicarium EGS 37-067 AF442803 AF443902 AY324708 S. majusculum EGS 29-094 AF442792 AF443891 S. alfalfae EGS 36-088 AF442775 AF443874 S. majusculum EGS16-068 AY329228 AY317032 AY324710 S. alfalfae EGS 39-127 AF442774 AF443873 S. astragali EGS29-062 AY329181 AY316984 AY324684 S. gracilariae EGS37-073 AY329217 AY317021 AY324709 S. astragali EGS 27-194.1 AF442777 AF443876 S. eturmiunum EGS29-099 AY329230 AY317034 AY324712 S. subglobuliferum HSAUP XF0140 AY751454 AY751459 S. xanthosomatis EGS 17-137 AF442804 AF443903 AY324758 S. xanthosomatis EGS47-197 AY329223 AY317027 AY324764 S. lycopersici EGS 46-001 AF442790 AF443889 S. lycopersici EGS45-036 AY329225 AY317029 AY324766 S. lycopersici EGS49-043 AY329256 AY317060 AY324770 S. callistephi EEB 1055 AF229482 AY278822 JQ672393 S. callistephi NO 536 AF442783 AF443882 AY324750 S. solani EGS 42-027 AF442797 AF443896 S. solani EGS41-135 AY329214 AY317018 AY324759 S. lancipes CBS 719.68 GQ395367 GQ395373 S. lancipes EGS46-182 AY329203 AY317007 AY324742 S. paludiscirpi EGS31-016 AY329231 AY317035 AY324769 S. trifolii EGS12-142 AY329218 AY317022 S. trifolii NO 712 AF442800 AF443899 S. drummondii CBS 346.83 GQ395365 KU850687 S. loti NO 1364 AF442788 AF443887 AY324776 S. loti NO 770 AF442789 AF443888 AY324775 S. sarciniforme EGS29-188 AY329221 AY317025 AY324746 S. sarciniforme EGS 38-121 AF442793 AF443892 AY324743 S. botryosum EGS08-069 AY329168 AY316968 AY324671 Alternaria alternata EGS34-016 AY751456 AY278808 AH013339  下载: 导出CSV
下载: 导出CSV
-
[1] LI R J, KUANG X P, WANG W J, et al. Comparison of chemical constitution and bioactivity among different parts of Lonicera japonica Thunb[J]. Journal of the Science of Food and Agriculture, 2019, 100: 614-622. [2] SHANG X F, PAN H, LI M X, et al. Lonicera japonica Thunb. : Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine[J]. Journal of Ethnopharmacology, 2011, 138: 1-21. doi: 10.1016/j.jep.2011.08.016 [3] 胡凯基, 王浩楠, 袁卉馥. 我国金银花研究进展浅析[J]. 河北北方学院学报(自然科学版), 2022, 38(7):38-42. [4] 秦祎婷, 朱艳霞, 郭玉海, 等. 河北巨鹿金银花生产现状与发展建议[J]. 中国现代中药, 2012, 14(2):34-36. doi: 10.3969/j.issn.1673-4890.2012.02.008 [5] 曾令祥. 金银花主要病虫害及防治技术[J]. 贵州农业科学, 2004(4):68-70. [6] 简美玲, 郑基焕, 毛润乾. 广东省金银花主要病虫害调查初报[J]. 广东农业科学, 2011, 38(14):74-76. doi: 10.3969/j.issn.1004-874X.2011.14.026 [7] HUANG X, ZHENG X, XU J, et al. First report of brown leaf spot on honeysuckle caused by Corynespora cassiicola in China[J]. Plant Disease, 2016, 100(11): 2326-2326. [8] CHANG C R, SHI H H, QU J H, et al. First report of brown leaf spot on Lonicera japonica caused by Corynespora cassiicola in Taiwan[J]. Plant Disease, 2020, 104(3): 989. [9] DE MIRANDA B E C, FERREIRA B W, ALVES J L, et al. Pseudocercospora lonicerigena a leaf spot fungus on the invasive weed Lonicera japonica in Brazil[J]. Australasian Plant Pathology, 2014, 43(3): 339-345. [10] 张永信, 刘晓杰, 冯争光, 等. 金银花褐斑病病原检测和ITS鉴定[J]. 河北农业大学学报, 2016, 39(4):73-77. doi: 10.13320/j.cnki.jauh.2016.0087 [11] 谢学文, 李宝聚, 石延霞, 等. 一种快速诱导匍柄霉菌产孢的方法: 中国, 201610335021.8[P]. 2016-05-19. [12] 熊晨琳, 王 群, 王 石, 等. 双玻片插片诱导新西兰匍柄霉产生分生孢子的方法[J]. 菌物研究, 2017, 15(2):129-132. [13] WHITE T J, BRUNS S, LEE S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[J]. PCR Protocols, A Guide to Methods and Application, 1990(1): 315-322. [14] BERBEE M L, PIRSEYEDI M, HUBBARD S, et al. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences[J]. Mycologia, 1999, 91(6): 964-977. doi: 10.1080/00275514.1999.12061106 [15] DENG J X, PAUL N C, LI M J, et al. Stemphylium platycodontis sp. nov., isolated from Platycodon grandiflorus in Korea[J]. Mycological Progress, 2013, 13(3): 477-482. [16] INDERBITZIN P, MEHTA Y R, BERBEE M L. Pleospora species with Stemphylium anamorphs: a four locus phylogeny resolves new lineages yet does not distinguish among species in the Pleospora herbarum clade[J]. Mycologia, 2009, 101(3): 329-339. doi: 10.3852/08-071 [17] SUDHIR K, GLEN, S, LI M, et al. MEGA X: Molecular evolutionary genetics analysis across computing platforms[J]. Molecular biology and evolution, 2018, 35: 1547-1549. doi: 10.1093/molbev/msy096 [18] 薛 峰, 张修国, 王 勇, 等. 中国匍柄霉研究II一新种及四个中国新记录[J]. 菌物学报, 2005(3):322-329. doi: 10.3969/j.issn.1672-6472.2005.03.003 [19] BESSADAT N, HAMON B, BATAILLÉ-SIMONEAU N, et al. Characterization of Stemphylium spp. associated with tomato foliar diseases in Algeria[J]. Phytopathologia Mediterranea, 2022, 61(1): 39-53. doi: 10.36253/phyto-13033 [20] ZHANG D Z, CHI W J, WANG C, et al. Pathogenic process-associated transcriptome analysis of Stemphylium lycopersici from Tomato[J]. International Journal of Genomics, 2022: 1-12. [21] NASEHI A, KADIR J, NASR-ESFAHANI M, et al. Identification of the new pathogen (Stemphylium lycopersici) causing leaf spot on Pepino (Solanum muricatum)[J]. Journal of Phytopathology, 2015, 164(6): 421-426. [22] NASEHI A, KADIR J B, NASR-ESFAHANI M, et al. Analysis of genetic and virulence variability of Stemphylium lycopersici associated with leaf spot of vegetable crops[J]. European journal of plant pathology, 2014, 140(2): 261-273. doi: 10.1007/s10658-014-0460-3 [23] ENJOJI S. Two diseases of tomato (2) (in Japanese)[J]. Journal of Plant Protection, 1931, 18: 48-53. [24] MEDINA R, FRANCO M E E, LUCENTINI C G, et al. Secondary metabolites synthesized by Stemphylium lycopersici and Fulvia fulva, necrotrophic and biotrophic fungi pathogen of tomato plants[J]. Current Plant Biology, 2019, 20: 100122. doi: 10.1016/j.cpb.2019.100122 [25] 周艳芳, 赵伟强, 王燕春. 赤峰地区番茄灰叶斑病病原菌的鉴定[J]. 北方园艺, 2017(22):55-58. [26] 周艳芳, 李戌清, 田忠玲, 等. 浙江番茄灰叶斑病病原菌[J]. 菌物学报, 2016, 35(9):1151-1156. [27] 李宝聚, 周艳芳, 李金萍, 等. 李宝聚博士诊病手记(三十)番茄匍柄霉叶斑病(灰叶斑病)的诊断与防治[J]. 中国蔬菜, 2010(23):24-26. [28] 黄文枫, 王 敏, 胡艳平, 等. 海南辣椒白斑病病菌鉴定及田间药效试验[J]. 热带生物学报, 2018, 9(2):203-206. doi: 10.15886/j.cnki.rdswxb.2018.02.012 [29] 刘 麟, 刘 欢, 王 娜, 等. 海南岛南繁基地向日葵新病害病原菌的分离和鉴定[J]. 中国植保导刊, 2021, 41(7):14-20. [30] 崔一平, 彭埃天, 宋晓兵, 等. 广东省江门市‘新会柑’叶斑病病原菌鉴定[J]. 植物保护, 2022, 48(4):236-239. doi: 10.16688/j.zwbh.2021311 [31] 郝芳敏, 臧全宇, 马二磊, 等. 甜瓜潜在病原菌番茄匍柄霉菌株的分离与鉴定[J]. 现代农业科技, 2020(20):89-90 + 95. doi: 10.3969/j.issn.1007-5739.2020.20.034 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 2141
- HTML全文浏览量: 1155
- PDF下载量: 57
- 被引次数: 0
