


-
在全球气候变化背景下,中国政府在第七十五届联合国大会上提出二氧化碳排放力争于2030年前达到峰值、2060年前实现碳中和的重大战略目标,人工林碳汇是其中的重要组成部分[1]。影响林分年内生长的因素复杂,包括温度、降水等气候因子以及林分密度、竞争等结构特征。林分密度是最重要的林分结构特征之一[2],与径向生长有直接关系,如Jordan Bello等[3]发现,低密度林中无柄橡树生长高于中密度林。在其他许多关于温带针叶林径向生长影响因子的研究中,太阳辐射、降水、饱和水汽压差和日最低气温等因子则表现出不同规律[4-9]。
华北落叶松(Larix principis-rupprechtii Mayr.)作为宁夏六盘山地区的主要造林树种,发挥着水土保持、水源涵养、固碳释氧等多种生态功能。目前关于华北落叶松林分的径向生长的年内变化特征,特别是对不同密度人工林如何响应环境因子变化,尽管已有较为丰富的研究成果[10-12],但受限于不同观测方法和分析手段及研究时段,当前研究结果中对同一因子影响的正负性和主导因子判定存在不同结论,如熊伟[10]等发现日最低温度与径向生长量有显著正相关关系,王亚蕊等[13]的研究则表明,该变量与生长量呈不显著负相关;管伟等[11]研究结果表明,空气相对湿度与生长存在正相关,Liu[12]的研究结果则表现为不显著的负相关关系。本研究通过线性混合效应模型分析了各环境因子对华北落叶松径向生长的影响,以期从另一角度探讨环境因子的驱动作用,并与相关分析结果对比验证。本研究拟解决的主要问题为:(1)分析不同密度下华北落叶松人工林年内径生长的动态变化;(2)确定影响其生长的主要环境因子,并量化各因素影响华北落叶松人工林径向生长的相对贡献。为此,本研究利用树木生长尺监测了六盘山华北落叶松树木个体的径向生长动态,并分析其树干径向生长的季节变化格局,以明确不同密度下树木年内径向生长的主要影响因素,为准确评估和预测未来华北落叶松人工林树木径向生长和森林动态、实现可持续管理与多功能维持提供科学依据。
-
研究区位于宁夏六盘山南坡的香水河小流域(106°12′~106°16′ E,35°27′~35°33′ N),海拔2 070~2 931 m,小流域总面积43.7 km²。研究区属于温带半湿润气候,雨热不同期,年均气温6.8 ℃,年均降水量710.3 mm,降水主要集中在6—9月份,全年无霜期90~130 d。土壤类型为森林灰褐土,土壤质地为壤土,土壤厚度为80 cm。石砾含量为8%~18%。研究区内乔木以华北落叶松人工林为主,天然次生林有白桦(Betula platyphlla Suk.)、华山松(Pinus armandii Franch.)等。灌木林主要有虎榛子(Ostyopsis davidiana Decne.)、灰栒子(Cotongaster acutifolius Turcz.)等。林下草本主要有苔草(Cyperaceae carex L.)、东方草莓(Fragaria orientali Lozinsk.)等。
-
在香水河小流域内选择4块不同林分密度的华北落叶松人工纯林样地,样地面积40 m × 40 m。为消除小地形影响,样地坡向相同(东南坡向)、坡度接近、坡位相同(均为下坡位)且海拔高度基本一致的相邻样地(表1)。对于样地间坡度差异,取林分密度接近、坡度有差异的2个高密度样地(样地3、4)进行了方差分析,结果表明其平均累积径向生长增量差异不显著,本研究样地间坡度差异不影响生长量分析。
表 1 样地基本信息
Table 1. General characteristics of the sample plot
样地编号
Sample NO.海拔
Elevation/m坡度
Slope/(°)林龄
Age/a平均胸径
DBH/cm平均树高
Height/m优势木平均高
Average height of
dominant trees/m样本量
Sample
size林分密度
Stand density/
(株·hm−2)1 2 336 18.0 39 21.4 ± 4.6 17.6 ± 2.3 20.9 ± 1.8 76 544 2 2 351 28.8 39 20.8 ± 3.0 19.3 ± 1.5 21.2 ± 1.1 91 742 3 2 300 29.0 39 19.0 ± 3.9 17.7 ± 2.4 20.5 ± 1.5 151 999 4 2 346 16.2 39 19.7 ± 4.0 19.5 ± 2.9 22.4 ± 1.5 144 1 020 -
将4个样地内的462株华北落叶松均选为样树,在4个样地内每棵树干距地1.3 m处刮去干枯树皮,安装QT-W304树木生长尺。观测时间从2022年5月开始,记录开始生长时的初始刻度值,每隔5~7 d观测生长尺刻度变化,观测至10月底生长季结束。最终获取到438株样木的生长尺有效数据。
-
在距离研究样地100 m处的空旷草地内设有标准气象观测场,使用 Weatherhawk232自动气象站监测气象因子,监测指标包括空气温度、降水量、空气相对湿度(以下简称空气湿度)、太阳辐射、风速(m·s−1),每隔1 min采集1次数据,每隔5 min记录1次平均值。利用空气温度和相对湿度计算饱和水汽压差[14]。
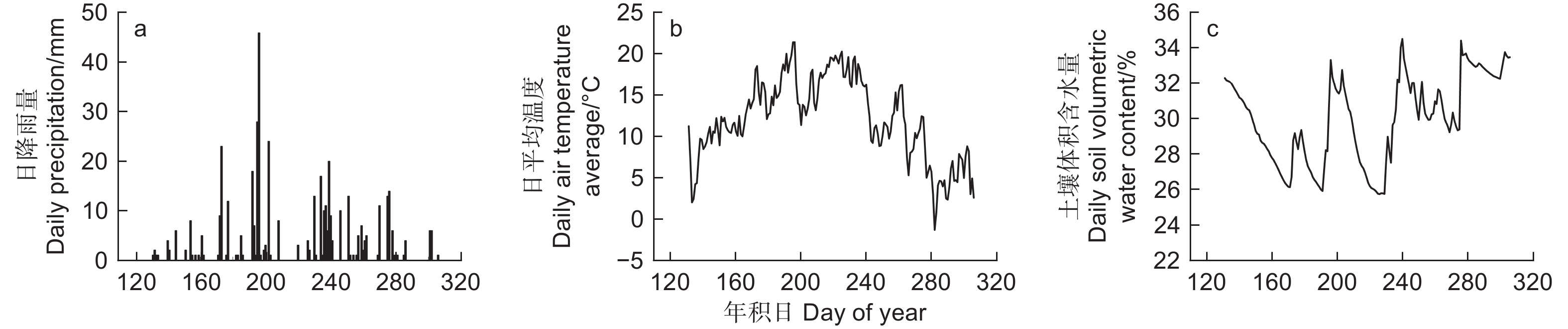
在样地内按 0~10、10~20、20~40 、40~60 和60~80 cm土层深度布设 EC-5 土壤水分传感器(Decagon,美国),并利用 EM 50 数据采集器(Decagon,美国)每 5 min 记录 1 次数据。本研究环境因子分析提到的土壤含水量均为土壤体积含水量,为0~80 cm土层实测土壤体积含水量的平均值(图1)。

图 1 研究期间主要环境因子变化
Figure 1. Variations of environmental major factors during the study period
-
Gompertz 函数在树木径向生长拟合中有广泛的应用[15],可以用来确定树木的生长季。使用Gompertz函数对不同密度华北落叶松生长季内平均累积径向生长量随时间变化进行拟合,然后对拟合函数求导得到华北落叶松生长速率。Gompertz函数模拟公式如下式[16]:
$ Y=A \times \exp \left[\mathrm{e}^{(\beta-k t)}\right] $
(1) 式中,Y为树干累积径向生长量;A为上渐近线;β表示与Y初始值有关的参数;k表示决定生长速率的参数;t表示时间;将拟合方程求导,得到华北落叶松的径向生长速率。
采用树径变化速率法判定快速增大阶段[13,17-18],即认为树干直径变化速率开始大于4 µm·d−1时为快速生长期的开始时间,开始小于4 µm·d−1时为结束时间[17]。
利用线性混合效应模型分析2022年华北落叶松日径向生长量与环境因子之间的关系,将胸径(表征树木个体间的竞争能力)、土壤水分、平均气温、饱和蒸汽压差、降水量、太阳辐射和风速作为固定效应,将林分密度(表征林分尺度的树木竞争)、生长时期(以月份表征,作为生长阶段的次一级分期)作为随机效应。用R语言中“lmerTest”包的“lmer”函数进行线性混合效应模型的分析。利用glmm.hp包[19]分析影响树木径向生长各因素的相对重要性,使用Origin 2018进行绘图。
-
图2为生长季节内华北落叶松的径向变化图,华北落叶松径向生长的季节变化呈现“S”型。2022年DOY 152—163(6月1日—6月13日)起,华北落叶松径向生长量开始大幅度增加;参考王亚蕊等[13]对六盘山华北落叶松研究结果,可将本研究生长季内树木直径变化过程分为3个阶段:1)相对稳定阶段(5月及9月上旬以后,DOY138—150及DOY245以后),此时树木直径净增量基本为零;2)快速增大阶段(6月初—7月下旬,DOY152—210),此时树木直径快速增大,但因树木蒸腾失水和土壤体积含水量变化等影响而呈波动增长趋势;3)涨缩波动阶段(8月上旬—9月上旬,DOY212—245),此时树木直径虽波动增长,但生长较慢(图2)。
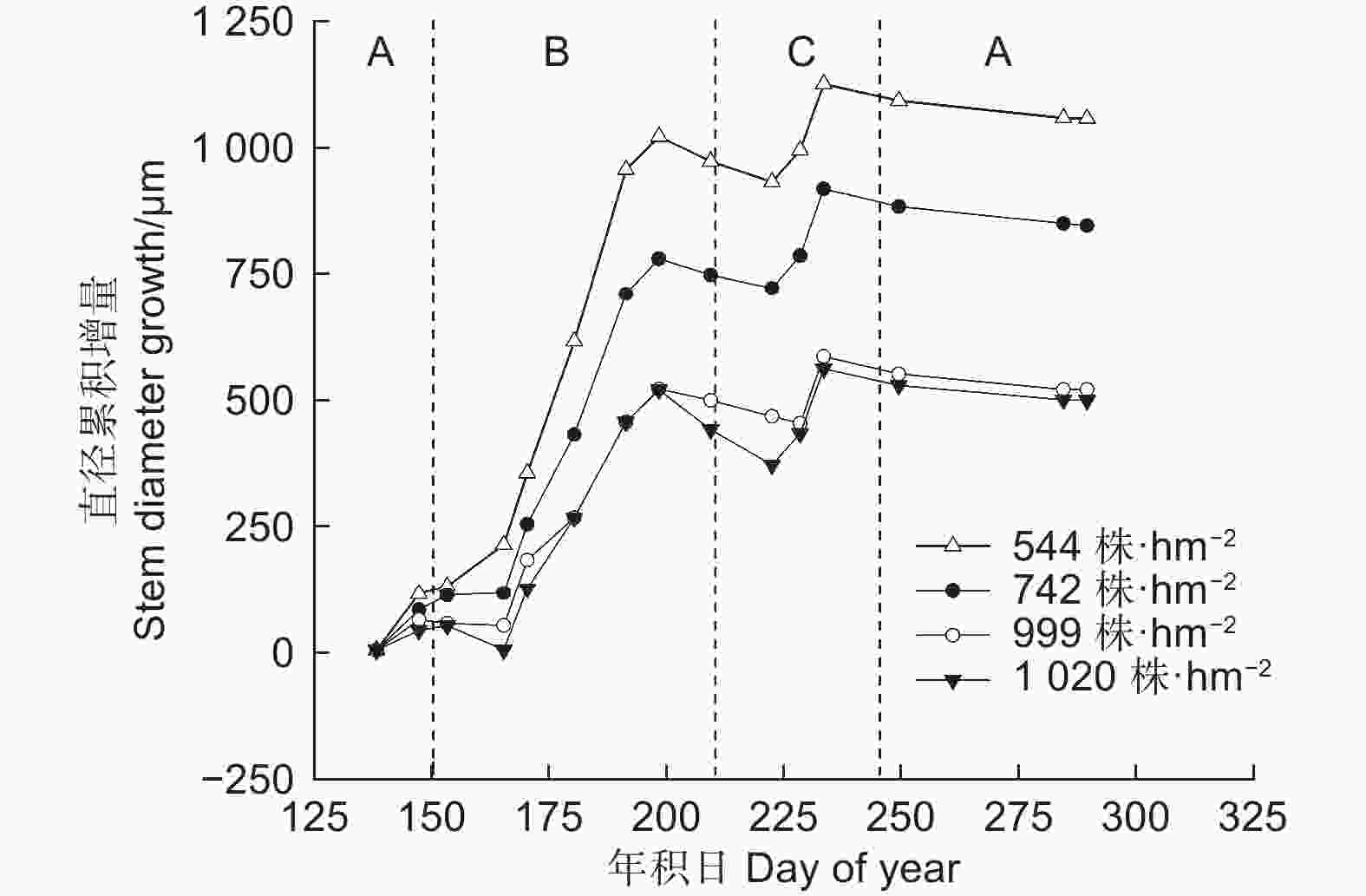
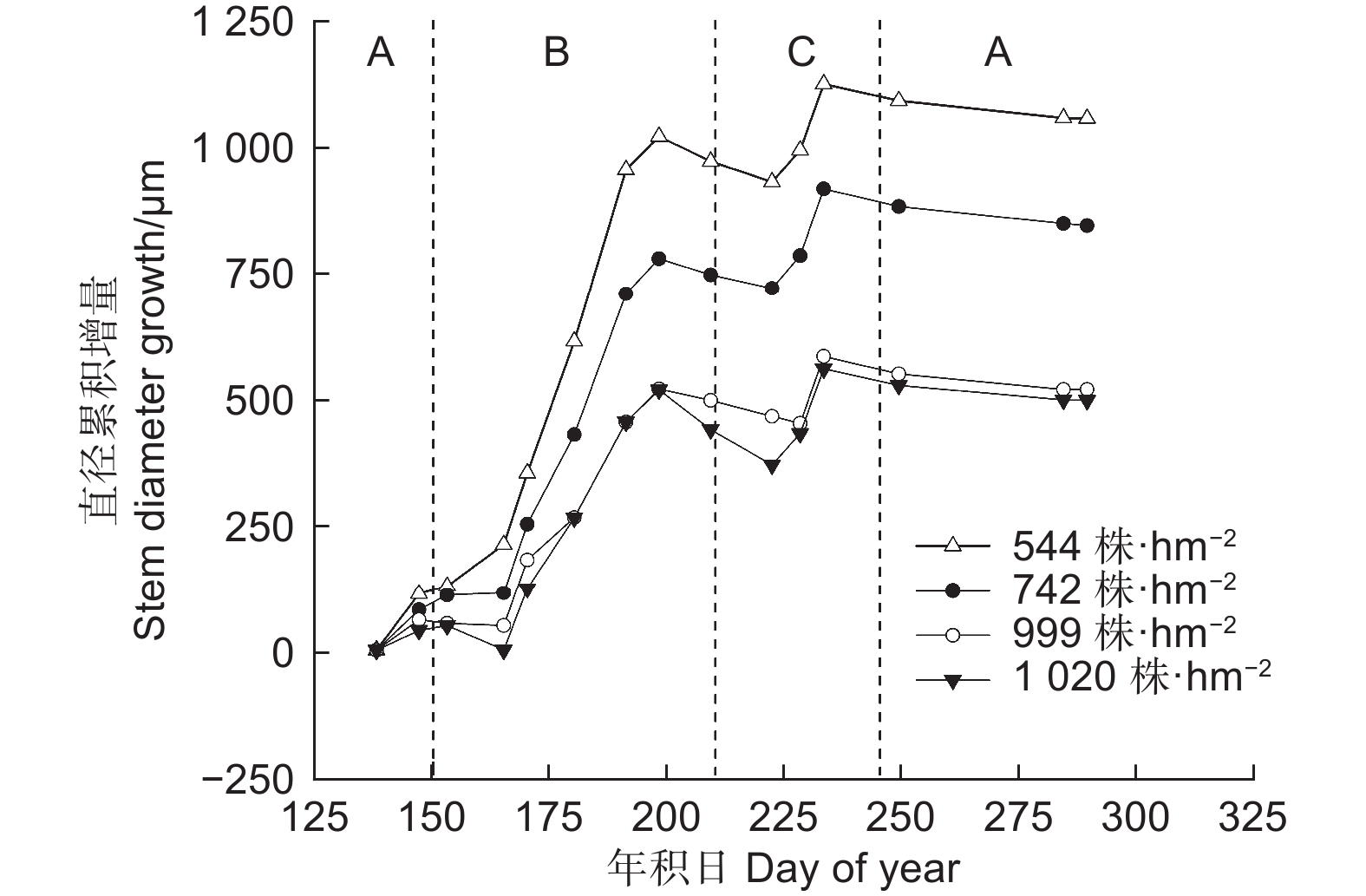
图 2 不同林分密度华北落叶松径向变化的生长季内格局
Figure 2. Seasonal pattern of stem diameter variation of Larix principis-rupprechtii at different stand densities
图3为2022年测量和模拟的不同林分密度下华北落叶松平均累积径向生长量和日平均径向生长速率变化图,由图3a可知,Gompertz函数模拟结果显示,华北落叶松平均累积径向生长量拟合曲线的决定系数(R2)均≥0.94,模型模拟与实测数据趋势基本一致。华北落叶松年内径向生长呈“S”型。不同林分密度下,华北落叶松的生长开始于5月—6月初(DOY121—163),7月中下旬(DOY198—209)达到高峰,9月上旬左右(DOY245)停止生长。

图 3 不同林分密度Gompertz函数拟合图(a)及生长速率变化(b)
Figure 3. Gompertz function-modeled curves of the cumulative stem growth(a) and daily growth rate(b) at different stand densities
图3b为模拟得到的华北落叶松日平均径向生长速率,本研究中分析华北落叶松树干径向生长的快速增大阶段为DOY 152—212(6月至7月),即生长速率>4 µm·d−1的阶段。4个林分密度的华北落叶松快速生长期分别为58,57,44和36 d,表现出林分密度越小快速增大阶段越长的趋势,随着林分密度的增大,快速增大阶段持续时间缩短。从生长速率来看,2022年华北落叶松径向生长速率曲线表现为单峰型;4个林分密度下最大生长速率依次为32.6、23.6、17.6 和22.5 µm·d−1,林分密度越低,最大生长速率越高。最大生长速率出现在DOY 171—174(6月20日—6月23日)左右。
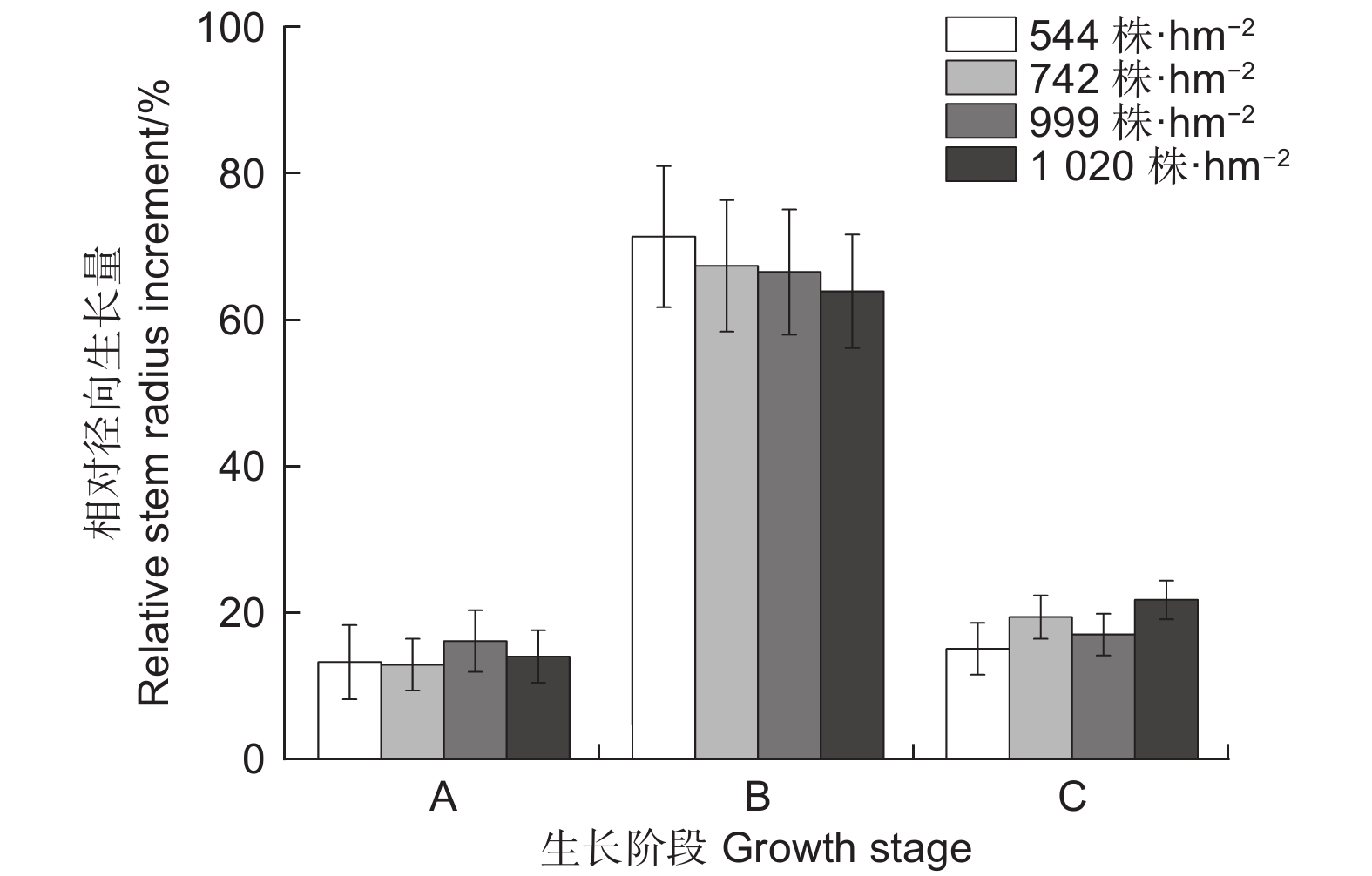
不同林分密度下生长的季节变化趋势不尽相同(图3b)。随着林分密度增大,华北落叶松快速增大阶段开始的时间较晚,结束时间较早,导致径向生长快速增大阶段时间少于低林分密度树木。但快速生长期开始时间均为6月初,快速生长期结束时间为7月末。就整个生长季而言,华北落叶松5月中旬开始生长,但生长速率较低,8月末生长结束,9月停止生长。由图4可以看出,2022年各密度下华北落叶松径向生长主要发生在阶段B快速增大阶段(6月初—7月下旬,DOY152—210),在544、742、999和1 020 株·hm−2的密度下,华北落叶松快速增大阶段相对径向生长量分别为71.5%、67.5%、66.7%和64.1%,除1 020 株·hm−2密度外,不同密度华北落叶松径向生长在快速增大阶段(阶段B)均超过整个生长季的2/3;相对稳定阶段和涨缩波动阶段华北落叶松径向生长均较低。
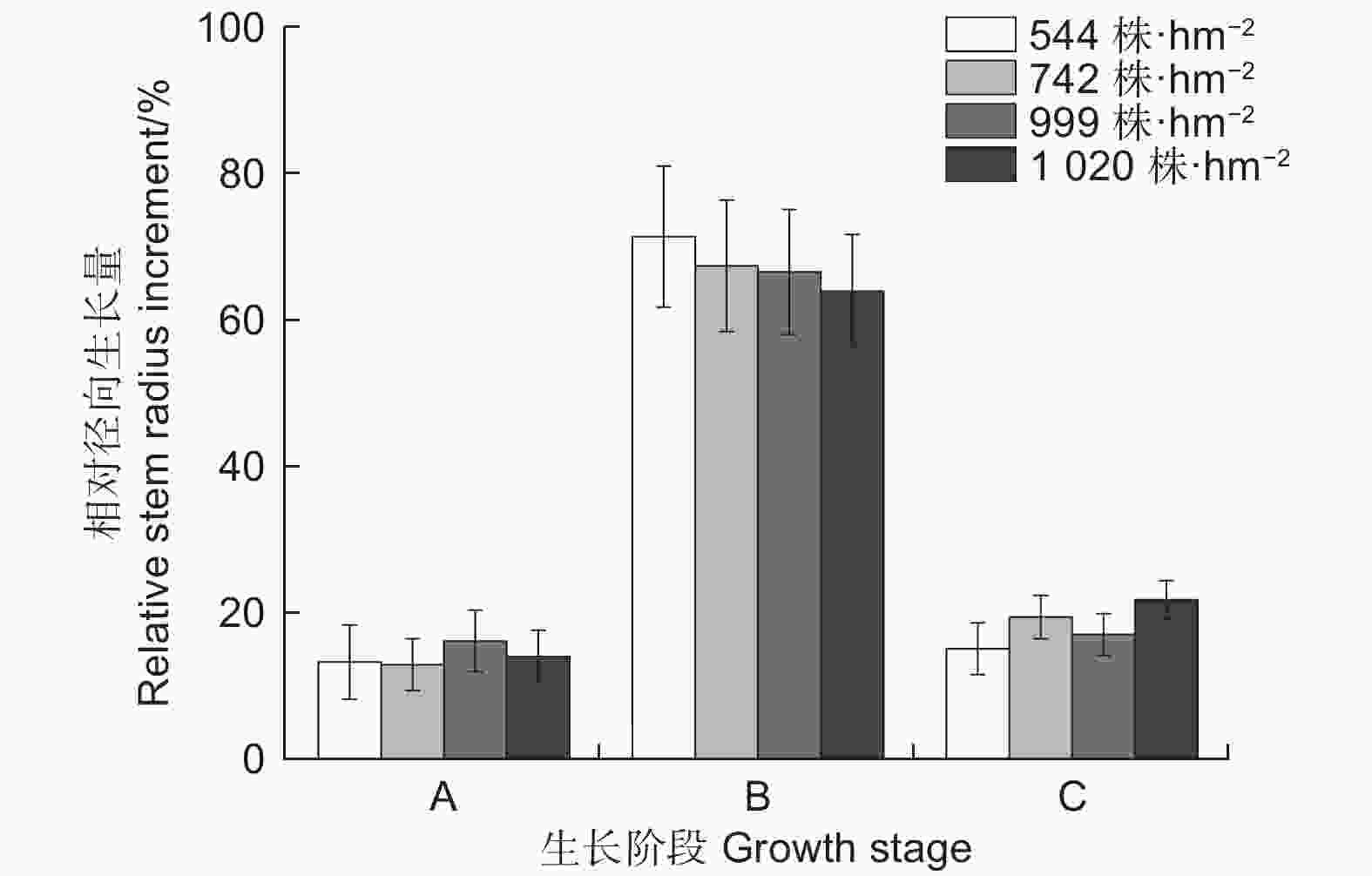
图 4 2022年生长季华北落叶松不同生长阶段相对径向生长量
Figure 4. Monthly relative stem radius increment of Larix principis-rupprechtii during growing season in 2022
-
表2为混合效应模型分析的气象、树木大小、林分密度和生长时期对华北落叶松日径向生长的影响结果。由表2可见,树木胸径、日平均气温、土壤含水量、降水量、饱和水汽压差、太阳辐射和风速均对华北落叶松日径向生长量有显著影响。作为表征树木之间竞争强弱的因子,树木胸径与华北落叶松的径向生长呈正相关性,即胸径越大,树木生长越快。对气候因素来说,树木日径向生长随平均气温、降水量和风速增加而显著增加,但随土壤体积含水量、饱和水汽压差和太阳辐射增加而显著下降,这可能与影响华北落叶松树木生长直接相关的冠层蒸腾作用有关。对比不同因素在决定华北落叶松日径向生长中的作用发现,土壤水分和平均气温是影响日径向生长的最主要因子,其相对贡献率分别达到30.30%和27.24%。太阳辐射、风速和饱和蒸气压差作为影响树木蒸腾作用的主要因素,它们的相对贡献率合计为38.03%,个体竞争因素(胸径大小)对树木日径向生长的影响作用非常小。然而,林分密度(表征林分尺度的竞争)和生长时期(即表征其生长时期)也显著影响华北落叶松的日径向生长。模型模拟结果表明,同时考虑林分密度、生长季月份、树木大小以及气象因子的模型可解释72.0%的方差,但排除林分密度和生长时期因素后,树木大小和气象因子共同仅能解释46.4%的方差。
表 2 气象因素、树木大小、林分密度和生长时期对华北落叶松日径向生长量的影响
Table 2. Effects of daily meteorological variables, tree size, stand density and month in growth season on daily radial increment of Larix principis-rupprechtii plantation during growing season
模型变量
Predictors估计值
Estimate95%置信区间95%
Confidence intervals标准误
SEp值
p value偏相关系数
Partial R2相对贡献率
Relative importance/%固定效应 Fixed effects 截距 Intercept 0.06 0.05~0.06 0.003 7 <0.001 胸径 DBH 0.00 0.00~0.00 0.000 0 <0.001 0.004 5 0.97 土壤含水量 Soil content −0.20 −0.22~−0.19 0.006 6 <0.001 0.140 7 30.30 平均气温 Mean temperature 0.00 0.00~0.00 0.000 0 <0.001 0.126 5 27.24 饱和水汽压差 VPD −0.01 −0.01~−0.01 0.000 9 <0.001 0.027 9 6.01 降水量 Rainfall 0.00 0.00~0.01 0.000 6 <0.001 0.016 1 3.47 太阳辐射 Solar radiation −0.00 −0.00~−0.00 0.000 0 <0.001 0.063 3 13.63 风速 Wind speed 0.02 0.01~0.02 0.000 6 <0.001 0.085 4 18.39 随机效应 Random effects σ2 0.00 τ00 月 0.00 τ00 林分密度 0.00 ICC 0.48 N林分密度 4 N月 6 观测数量 Observations 6 568 边际R2/条件R2
Marginal R2/Conditional R20.464/0.720 注:σ2为残差的方差,τ为随机效应引起的方差,ICC为类内相关系数,N是月或林分密度分类的数量,边际R2是固定效应的变异系数,条件R2是固定效应和随机效应影响的变异系数。
Note: σ2 is the variance of residuals, τ is the variance caused by random effects, ICC is intra-class correlation coefficient, N is the number of plots, Marginal R2 is the coefficient of variation for fixed effects, and Conditional R2 is the coefficient of variation for fixed and random effects. -
研究结果表明,六盘山南坡华北落叶松树木年内径向的生长季变化时间阶段性明显,大致可分为相对稳定、快速增大、涨缩波动3个阶段,与王亚蕊等[13]和马菁[20]对六盘山半湿润区华北落叶松的研究结果总体相似,但存在一些差异。本研究中树径在快速增大阶段(径向生长速率>4 µm·d−1),其生长速率较大;另外在第3阶段(涨缩波动)中,王亚蕊等[13]的研究结果表明树径在这一阶段呈波动减小趋势,而本研究中尽管这一阶段的生长速率较低,但树径仍在增长,即波动增长。熊伟等[10]在2007年对六盘山南坡华北落叶松研究结果表明,6—7月日均生长量在27. 0~44. 2 µm,本研究结果为在快速增大阶段(6月初—7月下旬,DOY152—210),华北落叶松日平均生长速率为17.6~32.5 µm·d−1,结果对比表明,近几年来,六盘山南坡华北落叶松生长较缓慢。这些差异一方面可能是以上研究主要是基于单株尺度,或选择代表性单木(通常是优势木)进行与环境因子等的相关分析,这一过程可能通过系统误差进而影响相关分析结果,本研究的径向生长数据来自全样地生长尺监测,有助于更准确地把握不同林分密度下,华北落叶松对环境因子的响应。另一方面则可能与树龄对年际生长的影响有关,有待开展较长时间尺度的生长特征对比研究。
在生长季时间尺度,华北落叶松径向变化曲线呈现“S”型;这与管伟等[11]的研究结果相似。本研究发现,6—7月是研究区华北落叶松径向生长的关键时期,超过2/3的径向生长发生在这一时段。在研究时段内,华北落叶松在其生长初期和快速生长期遭受了两次土壤体积含水量减少的“弱干旱”事件(DOY130—170和DOY210—230,见图1c),在生长初期,干旱可能会导致叶生长减缓或停止,进而导致气孔导度和碳同化速率降低,从而影响树木径向生长。即在土壤体积含水量减少阶段,树木径向生长首先会受到影响。土壤体积含水量减少导致的生理干旱限制了形成层细胞活动和细胞增大过程,导致华北落叶松生长速率减慢,从而使得2022年6月初期径向生长量减少。然而,由于土壤体积含水量含量相对较高,并未对树木造成不可逆的损伤,随着6、7月降水的补充和温度的升高,加之这一时期正处于生长生理期,华北落叶松的径向生长得到快速恢复。然而,由于8月上中旬持续无有效降水(伏旱),导致土壤体积含水量再次持续下降,继而中断了树木的径向生长,此后8月下旬的大量降水补偿了伏旱导致的径向生长量的减少,表现出华北落叶松对干旱的适应性。
径向生长观测手段对在不同尺度生长特征的刻画可能也有一定影响。树木径向生长对环境变化十分敏感,目前,对树木径向生长的研究多基于树木年代学和径向生长记录仪展开。其中树木年代学方法因定年准确、分辨率较高、方法成熟等优点,被广泛应用于树木生长与气候的关系研究中[21-28],可在年际及以上时间尺度上揭示树木径向生长的年际变化及其环境响应[28],但却无法连续监测年内尺度上树木的生长动态。另一方面,树木径向生长记录仪能够连续监测树木在日、月或季节尺度上的径向生长变化,揭示较短的时间尺度上影响树木生长的机理机制[29-39],但由于其观测仪器成本较高,观测样本较少,常导致研究结果存在较大不确定性。总体而言,这两种方法各有所长,但都难以保证在较大样本量观测的同时获取年内尺度上树木的径向生长变化特征,从而制约了更为精细时间尺度上对不同树木生长影响因素的确定,有必要使用多种手段进行观测,以补充树木年代学方法和径向生长记录仪以外、在数据尺度方面的不同需求。
-
树木径向生长同时受水分和温度等气象因子、林分密度等生物环境、树木个体竞争及生长时期等多种因素的综合影响。很多研究发现,不同地区或物种的树木径向生长受不同气候因子的影响,例如金敏艳等[7]发现,生长季干旱状况是决定祁连圆柏径向生长量最主要的气候因子;管伟等[11]则指出,降水和气温是影响落叶松直径变化的主要因子;张慧等[40]研究发现,滇西北长苞冷杉和大果红杉的日增长量都与降水呈显著正相关,但两树种的负相关因子则有所不同;高佳妮等[41]研究认为,除降水和气温以外,饱和水汽压差也对油松日径向生长量有显著影响。而本研究发现,华北落叶松日径向生长主要受日均气温、土壤体积含水量、降水量、饱和水汽压差、太阳辐射和风速的显著影响,这表明华北落叶松树木径向生长同时受多种气候因素的综合作用,特别是水分和热量因素的综合影响。
本研究发现,树木径向生长速率与树木胸径呈显著正相关性,表明大树的生长速率更高,这与一些已有研究结论相似,如SeMyung Kwon等[42]研究认为竞争显著影响了沙地樟子松树木的生长,即胸径越大的树木径向生长量越大;康剑等[43]对阿尔泰山西伯利亚五针松树木径向生长影响因素的研究时发现,树木径向生长主要受到竞争树的胸径和生长季前期气温的影响。
林分密度作为衡量林分尺度上树木竞争强度的重要指标,它可能会通过影响树木对土壤体积含水量、养分和光照的利用而影响树木的径向生长[2-3]。本研究发现,不同林分密度下的径向生长速率差异较大,低密度林分的径向生长速率较大,而高密度林分的生长速率较低,这在很多研究中也有类似结果[3,44-48]。例如Jordan Bello等[3]发现,无柄橡树在低密度下径向生长量高于中等林分密度的生长。此外,在本研究中,2个密度相似的高密度林分,其生长速率无显著差异。同时,随着林分密度的增大,华北落叶松快速增大阶段(生长速率大于4 µm·d−1)开始时间推迟,结束时间提前,导致快速增大阶段时间缩短。例如在土壤体积含水量下降阶段,低密度林分生长速率下降较小,而高密度林分的生长速率下降相对更大,这或许与林分密度差异导致的树木生长对水热等资源的限制性因子有关。
-
针对宁夏六盘山不同密度华北落叶松人工林,基于树木年内径向生长观测,主要结论如下:华北落叶松人工林立木年内生长季径向累积生长呈“S”型变化,可分为相对稳定、快速增大和涨缩波动3个生长阶段;快速增大阶段为华北落叶松主要生长阶段,快速增大时间为6月初—7月下旬(DOY152—210),其累积生长量约占整个生长季的2/3;随林分密度增加,华北落叶松年内累积径向生长量呈下降趋势,其快速增大阶段有所缩短。在本研究中,低密度华北落叶松平均累积径向生长量更大。本研究应用线性混合效应模型分析发现,树木胸径、日均温、土壤体积含水量、降水量、饱和水汽压差、太阳辐射和风速均对华北落叶松日径向生长有显著影响,其中,土壤水分和平均气温是影响日径向生长的最主要气候因子,而个体间竞争的影响非常小。在林分尺度上,林分密度和生长时期也是影响径向生长的重要因素。未来在六盘山华北落叶松径向生长与气候研究中,可以考虑环境因子和林分密度交互作用对华北落叶松径向生长的影响。
六盘山南坡不同密度华北落叶松人工林年内径向生长动态及其影响因素
Intra-annual Radial Growth Dynamics of Larix principis-rupprechtii Plantations with Different Stand Densities and Its Response to Environmental Factors
-
摘要:
目的 探究不同密度下华北落叶松人工林林分径向生长的季节变化特征及其对环境因子的响应,为准确评估和预测未来区域尺度上华北落叶松人工林树木径向生长和森林动态、实现可持续管理与多功能维持提供科学依据。 方法 在宁夏六盘山南部香水河小流域,设置4种不同密度华北落叶松样地,使用树木生长尺对华北落叶松年内径向生长过程进行监测,并同步监测气象因子、土壤温度和湿度。 结果 表明:(1)在生长季时间尺度,华北落叶松径向变化呈“S”型,径向变化呈相对稳定、快速增大、涨缩波动3个阶段的时间格局,快速增大阶段主要发生在6月初—7月下旬,DOY152—210。(2)在544、742、999和1 020 株·hm−2的密度下,生长季节内华北落叶松林平均累积径向生长量分别为1 070 ± 230 µm、775 ± 181 µm、518 ± 214 µm和494 ± 124 µm;随林分密度的增加,华北落叶松年内累积径向生长量呈下降趋势,其快速增大阶段缩短;(3)混合效应模型分析结果表明,华北落叶松径向生长主要受土壤水分和平均气温控制,但林分密度和生长时期对年内径向生长也有显著影响;同时考虑林分密度、生长时期、树木大小以及气象因子等变量的模型可解释72.0%的方差,但排除林分密度和生长时期因素后,树木大小和气象因子共同仅能解释46.4%的方差。 结论 超过2/3的华北落叶松径向生长发生在快速生长期;华北落叶松平均累积径向生长量随密度增大而减小;林分密度、生长时期、树木大小和气象因子共同调控树木的年内径向生长。 Abstract:Objective To investigate the seasonal variation characteristics of radial growth of Larix principis-rupprechtii plantations with different densities and its responses to environmental factors for providing a scientific basis for accurate assessment and prediction of the radial growth of L. principis-rupprechtii at regional scales, and achieving sustainable management and multi-functional maintenance. Method In the small watershed of Xiangshui River in the south of Liupanshan, four sample plots of L. principis-rupprechtii with different densities were set up, and the intra-annual radial growth process of L. principis-rupprechtii was monitored using a manual band dendrometer, and meteorological factors, soil temperature and humidity were monitored simultaneously. Result The results indicated that: (1) in the growing season, the radial growth of L. principis-rupprechtii showed a "S" curve. The rapid increasing stage mainly occurred from early June to late June, DOY152‒210. (2) The average cumulative radial growth of L. principis-rupprechtii decreased with the increase of stand density. In the plantations with densities of 544, 742, 999 and 1 020 N·hm−2, the average cumulative radial growth of L. principis-rupprechtii was 1 070 ± 230, 775 ± 181, 518 ± 214 and 494 ± 124 μm, respectively. With the increase of stand density, the cumulative radial growth of L. principis-rupprechtii showed a downward trend during the year, but its rapid increasing stage was shortened. (3) The results of mixed-effect model showed that the radial growth of L. principis-rupprechtii was mainly affected by soil moisture and average temperature, but stand density and growth period also had significant effects on radial growth during the year. The model including the variables of stand density, growth period, tree size and meteorological factors could explain 72.0% of the variance, but after excluding the factors of stand density and growth period, model with tree size and meteorological factors could only explain 46.4% of the variance. Conclusion More than two-thirds of the radial growth of L. principis-rupprechtii occurrs the rapid growth period. The average cumulative radial growth of L. principis-rupprechtii decreases with increasing density. Stand density, growth stage, tree size and meteorological factors jointly regulate the intra-annual radial growth of trees. -
图 1 研究期间主要环境因子变化
Figure 1. Variations of environmental major factors during the study period
图 2 不同林分密度华北落叶松径向变化的生长季内格局
Figure 2. Seasonal pattern of stem diameter variation of Larix principis-rupprechtii at different stand densities
图 3 不同林分密度Gompertz函数拟合图(a)及生长速率变化(b)
Figure 3. Gompertz function-modeled curves of the cumulative stem growth(a) and daily growth rate(b) at different stand densities
图 4 2022年生长季华北落叶松不同生长阶段相对径向生长量
Figure 4. Monthly relative stem radius increment of Larix principis-rupprechtii during growing season in 2022
表 1 样地基本信息
Table 1. General characteristics of the sample plot
样地编号
Sample NO.海拔
Elevation/m坡度
Slope/(°)林龄
Age/a平均胸径
DBH/cm平均树高
Height/m优势木平均高
Average height of
dominant trees/m样本量
Sample
size林分密度
Stand density/
(株·hm−2)1 2 336 18.0 39 21.4 ± 4.6 17.6 ± 2.3 20.9 ± 1.8 76 544 2 2 351 28.8 39 20.8 ± 3.0 19.3 ± 1.5 21.2 ± 1.1 91 742 3 2 300 29.0 39 19.0 ± 3.9 17.7 ± 2.4 20.5 ± 1.5 151 999 4 2 346 16.2 39 19.7 ± 4.0 19.5 ± 2.9 22.4 ± 1.5 144 1 020  下载: 导出CSV
下载: 导出CSV
表 2 气象因素、树木大小、林分密度和生长时期对华北落叶松日径向生长量的影响
Table 2. Effects of daily meteorological variables, tree size, stand density and month in growth season on daily radial increment of Larix principis-rupprechtii plantation during growing season
模型变量
Predictors估计值
Estimate95%置信区间95%
Confidence intervals标准误
SEp值
p value偏相关系数
Partial R2相对贡献率
Relative importance/%固定效应 Fixed effects 截距 Intercept 0.06 0.05~0.06 0.003 7 <0.001 胸径 DBH 0.00 0.00~0.00 0.000 0 <0.001 0.004 5 0.97 土壤含水量 Soil content −0.20 −0.22~−0.19 0.006 6 <0.001 0.140 7 30.30 平均气温 Mean temperature 0.00 0.00~0.00 0.000 0 <0.001 0.126 5 27.24 饱和水汽压差 VPD −0.01 −0.01~−0.01 0.000 9 <0.001 0.027 9 6.01 降水量 Rainfall 0.00 0.00~0.01 0.000 6 <0.001 0.016 1 3.47 太阳辐射 Solar radiation −0.00 −0.00~−0.00 0.000 0 <0.001 0.063 3 13.63 风速 Wind speed 0.02 0.01~0.02 0.000 6 <0.001 0.085 4 18.39 随机效应 Random effects σ2 0.00 τ00 月 0.00 τ00 林分密度 0.00 ICC 0.48 N林分密度 4 N月 6 观测数量 Observations 6 568 边际R2/条件R2
Marginal R2/Conditional R20.464/0.720 注:σ2为残差的方差,τ为随机效应引起的方差,ICC为类内相关系数,N是月或林分密度分类的数量,边际R2是固定效应的变异系数,条件R2是固定效应和随机效应影响的变异系数。
Note: σ2 is the variance of residuals, τ is the variance caused by random effects, ICC is intra-class correlation coefficient, N is the number of plots, Marginal R2 is the coefficient of variation for fixed effects, and Conditional R2 is the coefficient of variation for fixed and random effects.
下载: 导出CSV
-
[1] Huang J G, Ma Q Q, Rossi S, et al. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117: 20645-20652. [2] David I F. Linking forest growth with stand structure: Tree size inequality, tree growth or resource partitioning and the asymmetry of competition[J]. Forest Ecology and Management, 2019, 447: 139-157. doi: 10.1016/j.foreco.2019.05.053 [3] Jordan B, Patrick V, Thomas P, et al. How do mixing tree species and stand density affect seasonal radial growth during drought events[J]. Forest Ecology and Management, 2019, 432: 436-445. doi: 10.1016/j.foreco.2018.09.044 [4] Delpierre N, Lireux S, Hartig F, et al. Chilling and forcing temperatures interact to predict the onset of wood formation in Northern Hemisphere conifers[J]. Global Change Biology, 2019, 25: 1089-1105. doi: 10.1111/gcb.14539 [5] Rossi S, Deslauriers A, Gricar J, et al. Critical temperatures for xylogenesis in conifers of cold climates[J]. Global Ecology and Biogeography, 2008, 17: 696-707. doi: 10.1111/j.1466-8238.2008.00417.x [6] Rossi S, Deslauriers A, Anfodillo T, et al. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes[J]. Oecologia, 2007, 152(1): 1-12. doi: 10.1007/s00442-006-0625-7 [7] 金敏艳, 李进军, 车宗玺, 等. 祁连山中部祁连圆柏年内径向生长对气候因子的响应[J]. 生态学报, 2020, 40(21):7699-7708. [8] Jiang Y, Wang B Q, Dong M Y, et al. Response of daily stem radial growth of Platycladus orientalis to environmental factors in a semi-arid area of North China[J]. Trees Structure and Function, 2015, 29(1): 87-96. doi: 10.1007/s00468-014-1089-8 [9] Gao J N, Yang B, He M H, et al. Intra-annual stem radial increment patterns of Chinese pine, Helan Mountains, Northern Central China[J]. Trees-Structure and Function, 2019, 33(3): 751-763. doi: 10.1007/s00468-019-01813-w [10] 熊 伟, 王彦辉, 于澎涛, 等. 六盘山南坡华北落叶松(Larix principis-rupprechtii)树干直径生长及其对气象因子的响应[J]. 生态学报, 2007, 27(2):432-441. [11] 管 伟, 熊 伟, 王彦辉, 等. 六盘山北侧华北落叶松树干直径生长变化及其对环境因子的响应[J]. 林业科学, 2007, 43(9):1-6. [12] Liu Z B, Wang Y H, Tian A, et al. Intea-annual variation of stem radius of Larix principis-rupprechtii and its response to environmental factors in Liupan Mountains of Northwest China[J]. Forests, 2017, 8(10): 382. doi: 10.3390/f8100382 [13] 王亚蕊, 刘泽彬, 王彦辉, 等. 六盘山半湿润区华北落叶松树干半径变化特征及其影响因素[J]. 应用生态学报, 2020, 31(10):3313-3321. [14] Campbell G S, Norman J M. An Introduction to Environmental Biophysics [M]. 2nd Ed. New York, USA: SpringerVerlag, 1998. [15] 牛思圆, 刘鹏举, 雷相东, 等. 基于连清样地数据的全国杉木人工林平均木树高-胸径模型[J]. 林业科学研究, 2023, 36(1):117-123. [16] Mäkinen H, Seo J W, Nöjd P, et al. Seasonal dynamics of wood formation: A comparison between pinning, microcoring and dendrometer measurements[J]. European Journal of Forest Research, 2008, 127: 235-245. doi: 10.1007/s10342-007-0199-x [17] Tian Q Y, He Z B, Xiao S C, et al. Response of stem radial growth of Qinghai spruce (Picea crassifolia) to environmental factors in the Qilian Mountains of China[J]. Dendrochronologia, 2017, 44: 76-83. doi: 10.1016/j.dendro.2017.04.001 [18] Duchesne L, Houle D, D’Orangeville L, et al. Influence of climate on seasonal patterns of stem increment of balsam fir in a boreal forest of Québec, Canada[J]. Agricultural and Forest Meteorology, 2012, 162: 108-114. [19] Lai J S, Yi Zou Y, Zhang S, et al. glmm. hp: an R package for computing individual effect of predictors in generalized linear mixed models[J]. Journal of Plant Ecology, 2022, 15(6): 1302-1307. doi: 10.1093/jpe/rtac096 [20] 马 菁. 六盘山华北落叶松多时间尺度树干径向生长的环境响应[D]. 北京: 北京林业大学, 2020: 52-54. [21] K. Svobodová , Langbehn T, Bjrklund J, et al. Increased sensitivity to drought across successional stages in natural Norway spruce (Picea abies (L. ) Karst. ) forests of the Calimani Mountains, Romania[J]. Trees, 2019, 33: 1345-1359. doi: 10.1007/s00468-019-01862-1 [22] Antoine C, Florent M, Morine L, et al. Thinning increases tree growth by delaying drought-induced growth cessation in a Mediterranean evergreen oak coppice[J]. Forest Ecology and Management, 2018, 409: 333-342. doi: 10.1016/j.foreco.2017.11.030 [23] 胡正生, 郑永宏, 陆呈瑜, 等. 窗口宽度对树木径向生长与气候要素之间关系的影响[J]. 东北林业大学学报, 2023, 51(2):40-42 + 69. doi: 10.13759/j.cnki.dlxb.2023.02.015 [24] 朱明阳, 林 琳, 佘雨龙, 等. 云南轿子山不同海拔急尖长苞冷杉径向生长动态及其低温阈值[J]. 植物生态学报, 2022, 46(9):1038-1049. [25] 解萍萍, 张博奕, 董一博, 等. 华北落叶松和白杄径向生长对干旱的生态弹性差异[J/OL]. 应用生态学报, 1-9[2023-05-31]. [26] 赵凡凡, 叶 茂, 康利飞, 等. 基于微树芯的塔里木河下游胡杨年内径向生长动态研究[J]. 干旱区资源与环境, 2021, 35(12):156-162. [27] 韦小练, 范泽鑫, Arisa Kaewmano, 等. 热带季节雨林多花白头树年内径向生长动态及其对环境因子的响应[J]. 应用生态学报, 2021, 32(10):3567-3575. [28] 于德水, 卢 杰, 张 萌, 等. 西藏色季拉山西藏红杉径向生长对气温和降水波动的响应[J]. 林业科学研究, 2023, 36(1):91-99. doi: 10.12403/j.1001-1498.20220144 [29] 牛豪阁, 张 芬, 于爱灵, 等. 祁连山东部青杄年内径向生长动态对气候的响应[J]. 生态学报, 2018, 38(20):7412-7420. [30] Yanfang Wan, Pengtao Yu, Xiaoqing Li, et al. Seasonal Pattern of Stem Diameter Growth of Qinghai Spruce in the Qilian Mountains, Northwestern China[J]. Forests, 2019, 11(5): 494. [31] Yanfang Wan, Pengtao Yu, Xiaoqing Li, et al. Divergent Seasonal Patterns of Qinghai Spruce Growth with Elevation in Northwestern China[J]. Forests, 2022, 13(3): 388. doi: 10.3390/f13030388 [32] Aldea J, Bravo F, Bravo-Oviedo A, et al. Thinning enhances the species-specific radial increment response to drought in Mediterranean pine-oak stands[J]. Agricultural and Forest Meteorology, 2017, 237: 371-383. [33] Jiménez M N, Navarro F B, Sánchez-Miranda A, et al. Using stem diameter variations to detect and quantify growth and relationships with climatic variables on a gradient of thinned Aleppo pines[J]. Forest Ecology and Management, 2019, 442: 53-62. doi: 10.1016/j.foreco.2019.03.061 [34] Feng Xue, Yuan Jiang, Manyu Dong, et al. Seasonal and daily variations in stem water relations between co-occurring Larix principis-rupprechtii and Picea meyeri at different elevations[J]. Forest Ecology and Management, 2022, 504: 119821. doi: 10.1016/j.foreco.2021.119821 [35] 董满宇, 江 源, 杨浩春, 等. 芦芽山林线白杄生长季径向生长动态[J]. 植物生态学报, 2012, 36(9):956-964. [36] Biondi F, Hartsough P C, Estrada I G. Daily weather and tree growth at the tropical treeline of North America[J]. Arctic, Antarctic, and Alpine Research, 2005, 37: 16-24. doi: 10.1657/1523-0430(2005)037[0016:DWATGA]2.0.CO;2 [37] Wang W B, Zhang F, Yuan L M, et al. Environmental factors effect on stem radial variations of Picea crassifolia in Qilian Mountains, Northwestern China[J]. Forests, 2016, 7(10): 210. [38] Deslauriers A, Rossi S, Anfodillo T. Dendrometer and intra-annual tree growth: What kind of information can be inferred[J]. Dendrochronologia, 2007, 25(2): 113-124. doi: 10.1016/j.dendro.2007.05.003 [39] Xiaodong Ma, Zhuo Pang, Juying Wu, et al. Seasonal pattern of stem radial growth of Salix matsudana and its response to climatic and soil factors in a semi-arid area of North China[J]. Global Ecology and Conservation, 2021, 28: e01701. doi: 10.1016/j.gecco.2021.e01701 [40] 张 慧, 付培立, 林友兴, 等. 滇西北白马雪山长苞冷杉和大果红杉年内径向生长动态及其对环境因子的响应[J]. 应用生态学报, 2022, 33(11):2881-2888. [41] 高佳妮, 杨 保, 秦 春. 树木年内径向生长对干旱事件的响应—以贺兰山油松为例[J]. 应用生态学报, 2021, 32(10):3505-3511. [42] SeMyung Kwon, 潘磊磊, 时忠杰, 等. 不同竞争强度下的沙地樟子松天然林树木径向生长及其气候响应[J]. 生态学杂志, 2019, 38(7):1962-1972. [43] 康 剑, 梁寒雪, 蒋少伟, 等. 竞争和气候对新疆阿尔泰山西伯利亚五针松树木径向生长的影响[J]. 植物生态学报, 2020, 44:1195-1202. [44] 郑 颖, 冯 健, 于世河, 等. 辽东山区不同密度落叶松人工幼龄林林木生长和土壤养分特性[J]. 中南林业科技大学学报, 2022, 42(1):94-103. [45] Trouvé R, Bontemps J D, Collet C, et al. Radial growth resilience of sessile oak after drought is affected by site water status, stand density, and social status[J]. Trees, 2017, 31: 517-529. doi: 10.1007/s00468-016-1479-1 [46] Sonia Condés, Miren Del Rio, Hubert Sterba. Mixing effect on volume growth of Fagus sylvatica and Pinus sylvestris is modulated by stand density[J]. Forest Ecology and Management, 2013, 292: 86-95. doi: 10.1016/j.foreco.2012.12.013 [47] 王翰琛, 张雄清, 张建国, 等. 杉木人工林不同密度间伐林分生长优势的变化规律分析[J]. 林业科学研究, 2021, 34(5):32-38. [48] 张 程, 欧阳林男, 陈少雄. 3种初植密度桉树林分生长、材种出材量及经济效益动态分析[J]. 林业科学研究, 2021, 34(4):58-65. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1532
- HTML全文浏览量: 1062
- PDF下载量: 41
- 被引次数: 0

