

-
生态化学计量学是从元素计量角度探讨生态系统中多重化学元素平衡和耦合关系的学科,为揭示元素之间相互作用和养分平衡机制提供了有效途径[1]。碳(C)、氮(N)和磷(P)是植物生长发育的主要元素,其化学计量比能够反映植物、凋落物和土壤的养分平衡状况[2]。凋落物是森林生态系统重要的养分储藏库,植物生长发育所需养分约70%来源于凋落物的分解和释放[3]。凋落物分解过程中C、N、P含量的变化及其化学计量特征是影响凋落物分解速率及养分归还的重要因素,与土壤C、N、P循环过程密切相关[4]。研究表明,凋落物中N含量升高或者C∶N的降低会加快凋落物分解,加速养分的释放,C∶N和C∶P也可以反映土壤中N和P的供应情况[5]。因此,开展植物凋落物分解过程中生态化学计量特征的研究,是探究森林生态系统养分循环和调控机制的重要途径之一。
温度是影响凋落物产量及分解的重要气候因子,温度升高加速了植物体的代谢速率,提高了凋落物产量与基质质量,从而影响森林生态系统物质循环的速率[6]。研究发现,凋落物产量与年平均气温呈线性正相关关系;植物叶片可通过降低N、P含量、提高C∶N比来适应长期的高温环境[7];增温处理明显缩短了N、P释放的周期[8]。凋落叶作为森林凋落物的主体,其养分归还的速率直接影响森林生态系统的养分供应水平。不同发育阶段的森林凋落叶在物理结构与化学组成上存在明显差异,直接影响着凋落叶的分解速率与养分释放[9]。然而,在全球变暖的大背景下,温度升高是否会加速森林凋落叶分解过程中养分的释放?温度升高对森林凋落叶分解的影响是否随林分发育阶段的不同而变化?目前还尚不清楚,而这些问题的阐释,对于全面认识凋落物分解过程中养分的释放规律具有重要的科学意义。
杉木(Cunninghamia lanceolata (Lamb.) Hook.)是我国栽植面积最大的速生树种之一,主要分布于我国南亚热带地区,在我国林业生产中具有举足轻重的地位[10]。杉木人工林具有明显的枯枝叶宿存特性,其枯死枝叶并不马上脱落而会宿存在树干多年,导致林地凋落物产量少,加之杉木凋落物基质质量差,凋落物分解缓慢,形成养分循环障碍[11]。已有研究证实,亚热带地区受全球气候变暖带来的气温增幅明显高于全球平均增温[12],温度升高已成为影响杉木人工林凋落物分解及养分释放的重要因子,对改变杉木人工林生态系统养分平衡具有重要影响。李澳归等[13]发现土壤增温显著改变了杉木凋落物的理化性质,凋落物N、P含量的提高以及抗拉强度的减小对凋落物初期的分解具有明显的促进作用。杨成邦等[14]认为温度升高显著降低了杉木人工林土壤中的N含量,不利于森林生态系统生产力的提高。王书丽等[15]发现温度升高使杉木一年生与两年生凋落叶分解过程中C∶N、C∶P明显增大。然而,以往研究更多关注于温度升高对某一发育阶段杉木凋落物分解过程的影响,而关于不同发育阶段杉木凋落物分解过程中养分释放及化学计量比对温度升高的响应研究还鲜见报道。为实现杉木人工林大径材的培育目标,杉木成熟林、过熟林林分占比不断增加,探究温度升高对这些发育阶段杉木林分凋落物分解过程的影响,对于培育杉木大径材、改善杉木人工林土壤肥力具有重要的指导意义。
有鉴于此,本研究以中龄林(18年生)、成熟林(30年生)和过熟林(42年生)杉木人工林的凋落叶为研究对象,通过设计不同温度处理(25、30、35 ℃)在室内模拟凋落叶分解试验,测定不同温度处理下不同发育阶段杉木凋落叶在第15、30、72、118、172、264 d的C、N、P含量,比较不同分解时间C、N、P残留率的变化规律及其化学计量特征,以期揭示温度升高对不同发育阶段杉木凋落叶分解过程中养分释放及化学计量比的影响规律,为杉木人工林科学培育及合理经营提供理论支撑。
-
研究区位于福建省三明市三元区福建农林大学莘口教学林场(26º10′ N,117º27′ E),该地处于中亚热带南缘,位于武夷山脉与戴云山脉之间,属于中亚热带季风气候,温暖湿润,降雨量充足,年平均气温19.5 ℃,年平均降雨量1 700 mm,年平均蒸发量1 585 mm,年平均日照时间1 892 h,年平均相对湿度78.0%,无霜期300 d左右,研究区域土壤是由粉沙页岩发育而来的山地红壤,pH为4.40~4.60,有机质含量为35.3~42.4 g·kg−1 [11]。
-
2021年7月,在杉木人工纯林中选择中龄林(18年生)、成熟林(30年生)和过熟林(42年生)为研究对象,三种林分分别于2003年、1991年和1979年营造。采用课题组自制的凋落物收集器对凋落物进行收集,每个发育阶段杉木人工林分别设置3个20 m × 20 m的标准样地,每个样地中随机设置10个面积为1 m × 1 m的凋落物收集框,设置在距离地面约50 cm处[11]。2021年10月,把收集到的凋落物带回实验室,将针叶从小枝分离,去除新鲜叶片和开始分解的叶片,选取部分样品用于初始(即分解第0 d)指标测定。采用随机取样法在各发育阶段杉木人工林内取表层土(0~20 cm),挑出杂物和石子,做好标记后带回实验室备用。
选择定制的长方型塑料盒进行分解试验,其规格为长17 cm、宽11 cm、高5 cm,在分解盒中平铺装入原生境地表土壤200 g。根据课题组长期观测发现,杉木凋落叶产量约为119~222 g·m−2·a−1[11],因此在分解盒中平铺5 g凋落叶,为方便后期收集不同分解时间的凋落叶,土壤与凋落叶之间用1 mm孔径尼龙网相隔。将不同发育阶段杉木凋落叶分解盒随机放置在为25、30、35 ℃的植物培养箱(BPC500-2H, 福建九圃生物科技有限公司)进行分解,培养箱湿度始终保持在80%左右,以消除水分因子对凋落物分解的影响。在分解第15、30、72、118、172、264 d时进行凋落叶取样,每次随机选取各温度处理的3个分解盒,总共162个分解盒(3个温度处理 × 3个发育阶段 × 6个取样时间 × 3个重复)。不同发育阶段杉木凋落叶的初始化学性质见表1。
表 1 不同发育阶段杉木凋落叶初始化学性质
Table 1. Initial chemical properties of Chinese fir leaf litter among different developmental stages
发育阶段
Developmental stageC含量
C content/(g·kg−1)N含量
N content/(g·kg−1)P含量
P content/(g·kg−1)中龄林(18 a) 489 ± 1.370 A 19.9 ± 0.372 A 0.487 ± 0.013 3 A 成熟林(30 a) 493 ± 0.142 A 19.9 ± 0.698 A 0.432 ± 0.014 6 B 过熟林(42 a) 490 ± 0.211 A 19.3 ± 1.030 A 0.503 ± 0.013 5 A 注:同列不同大写字母表示不同发育阶段凋落叶间差异显著(P < 0.05)
Note: Uppercase letter in the same column showed significant difference between different developmental stages (P < 0.05) -
将每次获得的凋落叶样本置于烘箱内75 ℃烘干至质量恒定,称干质量,采用粉碎机粉碎后过筛(φ=0.15 mm),使用碳氮元素分析仪(Vario Max, Elementar, 德国)测定全C、全N含量。使用电感耦合等离子体发射光谱仪(ICP-OES,OPTIMA 8000, Perkin Elmer, 美国)测定全P含量。测定前,采用微波消解仪进行样本消解(ETHOS UP, Milestone, 美国),消解液为5 mL硝酸 + 1 mL过氧化氢(30%)。通过各时期测定的全C、全N、全P含量计算出各元素的残留率E,公示如下[16]:
$ E={W}_{t}\times {C}_{t}/({W}_{0}\times {C}_{0}) \times 100{\text{%}}$
(1) 式(1)中:W0为初始凋落物的质量/g,Wt为时间t取样时凋落叶质量/g,C0为初始凋落叶元素含量/(g·kg−1),Ct为时间t取样时凋落叶元素含量/(g·kg−1)。
应用Olson负指数衰减模型对各元素残留率(E)与时间(t)进行拟合,拟合方程如下[17]:
$ E=\mathrm{a} \times \mathrm{e}^{-k t} $
(2) 式(2)中,E为元素残留率/%,a为拟合参数,k为年分解系数,t为分解时间。
元素分解95%(T95%)所需时间,公式如下:
$ {T}_{95\%}=-\mathrm{l}\mathrm{n}(1-0.95)/k $
(3) 式(3)中,k为年分解系数。
-
不同温度处理下不同发育阶段杉木凋落叶分解过程中C、N、P残留率与化学计量比的差异采用单因素方差分析(One-way ANOVA)和Duncan多重比较。采用重复测量方差分析检验温度、发育阶段、分解时间及其相互作用对凋落叶分解过程中C、N、P残留率及其化学计量比的影响。C、N、P残留率及其化学计量比之间的相关性采用Pearson法进行分析。利用SPSS 25.0软件和Origin 2021进行数据统计及做图。图表中数据均为平均值 ± 标准差(n = 3)。
-
通过重复测量方差分析(表2)可知,温度对凋落叶分解过程中N、P残留率变化有极显著影响(P < 0.01),发育阶段对N残留率有极显著影响,分解时间对C、N、P残留率有极显著影响;温度 × 发育阶段、发育阶段 × 分解时间、温度 × 分解时间、温度 × 发育阶段 × 分解时间的交互效应对分解过程中C、N残留率均有极显著影响,发育阶段 × 分解时间、温度 × 发育阶段 × 分解时间的交互效应对P残留率的影响均达显著水平(P < 0.05)。
表 2 温度、发育阶段、分解时间及其交互效应对杉木凋落叶C、N、P残留率及其化学计量特征的影响(F值)
Table 2. Effects of temperature, developmental stage, decomposition time and their interaction effects on C, N and P residual rates and stoichiometric characteristics of Chinese fir litters (F value)
指标 Indicator C N P C∶N C∶P N∶P TE 1.78 82.00** 7.49** 45.00** 9.09** 39.40** D 0.54 11.30** 1.66 6.12** 1.00 4.53* T 58.90** 37.20** 53.10** 56.10** 54.30** 17.90** TE × D 5.44** 8.50** 0.78 14.50** 0.62 21.50** D × T 6.36** 30.00** 1.90* 54.10** 1.05 1.65 TE × T 10.20** 65.70** 1.27 39. 80** 3.85** 26.00** TE × D × T 4.89** 10.50** 2.82** 7.21** 2.92** 9.26** 注:* P < 0.05;**P < 0.01;TE-温度;D-发育阶段;T-分解时间
Notes: * P < 0.05; **P < 0.01; TE- temperature; D- developmental stage; T- decomposition time由图1可知,不同发育阶段杉木凋落叶分解过程中C、N、P残留率随分解时间的进行总体呈逐渐减小趋势。C在整个分解过程中表现为释放—富集—释放的变化趋势,不同发育阶段杉木凋落叶C在分解前15 d均呈明显释放状态;中龄林、成熟林在30~72 d发生一定程度的富集;分解72~172 d时,各发育阶段杉木凋落叶C均表现释放过程。分解264 d时,35 ℃处理下中龄林、成熟林与过熟林凋落叶C残留率比25 ℃处理分别减少了6.10%、5.44%、5.25%,且25与35 ℃处理间的差异达到极显著水平(P < 0.01)。
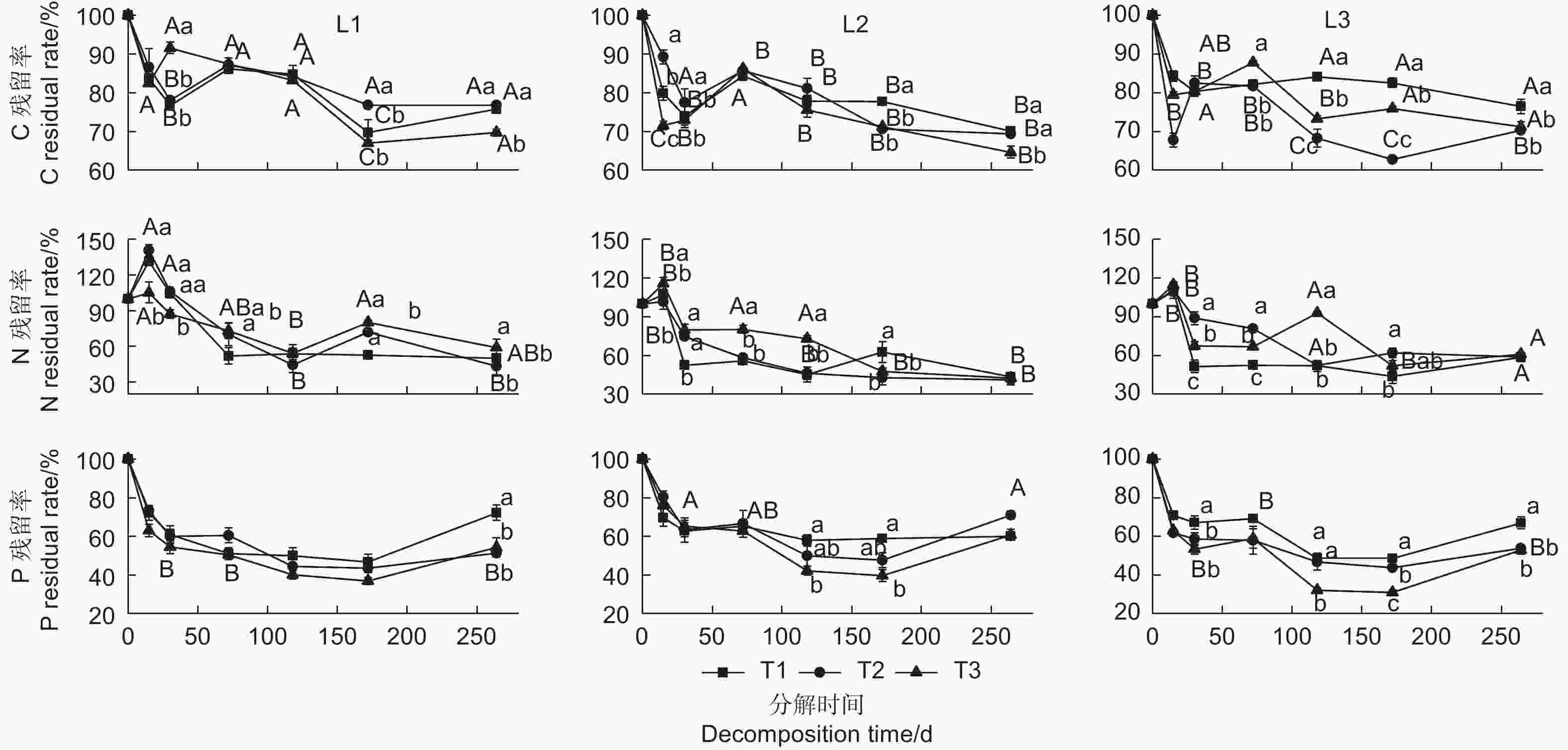
图 1 不同温度下不同发育阶段杉木凋落叶分解过程中C、N、P残留率的变化
Figure 1. Changes of the residual rates of C,N and P of leaf litter among different developmental Chinese fir plantation under different temperature treatments
凋落叶分解过程中N残留率变化呈富集-释放的变化趋势。分解15 d时,各温度处理下不同发育阶段凋落叶N残留率均发生不同程度的增加,表现出富集过程,升温处理(35 ℃)显著增加了成熟林凋落叶N的残留率,但升温处理(35 ℃)显著降低了中龄林凋落叶N的残留率;分解15~30 d时,各发育阶段杉木凋落叶N均表现为快速释放过程;分解30~172 d时,各阶段杉木凋落叶N残留率呈波动下降,N释放过程变缓。试验结束时(264 d),中龄林凋落叶N残留率表现为25 ℃处理显著低于35 ℃处理(P < 0.05),平均减少了9.03%;而成熟林、过熟林凋落叶在不同温度处理下N残留率没有显著差异(P > 0.05)。
凋落叶分解过程中P残留率表现为释放—富集模式,各发育阶段杉木凋落叶P在分解前30 d表现为快速释放阶段,30~172 d为缓慢释放,而在172~264 d时则发生一定程度富集。成熟林、过熟林凋落叶分解到118、172 d时均表现为35 ℃处理下P残留率显著低于25 ℃处理(P < 0.05);分解第264 d,中龄林与过熟林凋落叶P残留率在35 ℃处理下显著低于25 ℃处理,分别减少了18.0%、14.1%。在30、35 ℃处理下,凋落叶分解过程中成熟林P残留率明显大于中龄林、过熟林。
-
应用Olson指数衰减模型对不同温度处理各发育阶段杉木凋落叶C、N、P元素残留率(E)与时间(t)进行拟合(表3),结果表明,C残留率模型拟合效果未达显著水平,N、P残留率拟合模型均达到显著水平(P < 0.05)。成熟林、过熟林杉木凋落叶分解过程中30 ℃处理下C周转期比25 ℃处理缩短了22.7%和18.8%,成熟林、过熟林凋落叶分解中35 ℃处理下N周转期比25 ℃分别缩短了34.4%和16.9%,P周转期分别缩短了38.4%和43.8%。
表 3 不同温度处理下各发育阶段杉木凋落叶C、N、P平均分解速率
Table 3. Average decomposition rates of C, N and P of leaf litter among different developmental stages under different temperature treatments
元素
Element温度处理
Temperature
treatment发育阶段
Developmental
stage分解方程
Decomposition
equation分解系数
Decomposition
coefficient95%分解所需时间
95% decomposition
time/a相关系数
Correlation
coefficientP值
P valueC 25 ℃ 中龄林 y=56.3e−0.412t 0.412 7.27 0.347 P > 0.05 成熟林 y=56.7e−0.391t 0.391 7.66 0.334 P > 0.05 过熟林 y=57.4e−0.378t 0.378 7.93 0.337 P > 0.05 30 ℃ 中龄林 y=57.0e−0.383t 0.383 7.82 0.338 P > 0.05 成熟林 y=59.8e−0.506t 0.506 5.92 0.464 P < 0.05 过熟林 y=56.7e−0.465t 0.465 6.44 0.390 P > 0.05 35 ℃ 中龄林 y=58.3e−0.456t 0.456 6.57 0.401 P > 0.05 成熟林 y=56.4e−0.379t 0.379 7.91 0.327 P > 0.05 过熟林 y=57.2e−0.340t 0.340 8.81 0.303 P > 0.05 N 25 ℃ 中龄林 y=81.3e−0.905t 0.905 3.31 0.569 P < 0.01 成熟林 y=80.1e−0.945t 0.945 3.17 0.666 P < 0.01 过熟林 y=76.8e−0.778t 0.778 3.85 0.537 P < 0.05 30 ℃ 中龄林 y=108 e−1.39t 1.390 2.16 0.775 P < 0.01 成熟林 y=86.9e−1.32t 1.320 2.27 0.829 P < 0.01 过熟林 y=94.0e−0.737t 0.737 4.07 0.957 P < 0.01 35 ℃ 中龄林 y=93.5e−0.710t 0.710 4.22 0.708 P < 0.01 成熟林 y=114. e−1.44t 1.440 2.08 0.838 P < 0.01 过熟林 y=93.2e−0.936t 0.936 3.20 0.601 P < 0.01 P 25 ℃ 中龄林 y=.79.7e−0.695t 0.695 4.31 0.614 P < 0.05 成熟林 y=75.0e−0.462t 0.462 6.48 0.589 P < 0.01 过熟林 y=74.6e−0.503t 0.503 5.96 0.513 P < 0.05 30 ℃ 中龄林 y=73.5e−0.809t 0.809 3.70 0.711 P < 0.01 成熟林 y=75.2e−0.479t 0.479 6.25 0.464 P < 0.01 过熟林 y=72.1e−0.738t 0.738 4.06 0.654 P < 0.05 35 ℃ 中龄林 y=65.2e−0.701t 0.701 4.27 0.555 P < 0.05 成熟林 y=73.7e−0.751t 0.751 3.99 0.553 P < 0.05 过熟林 y=65.53e−0.895t 0.895 3.35 0.561 P < 0.05 -
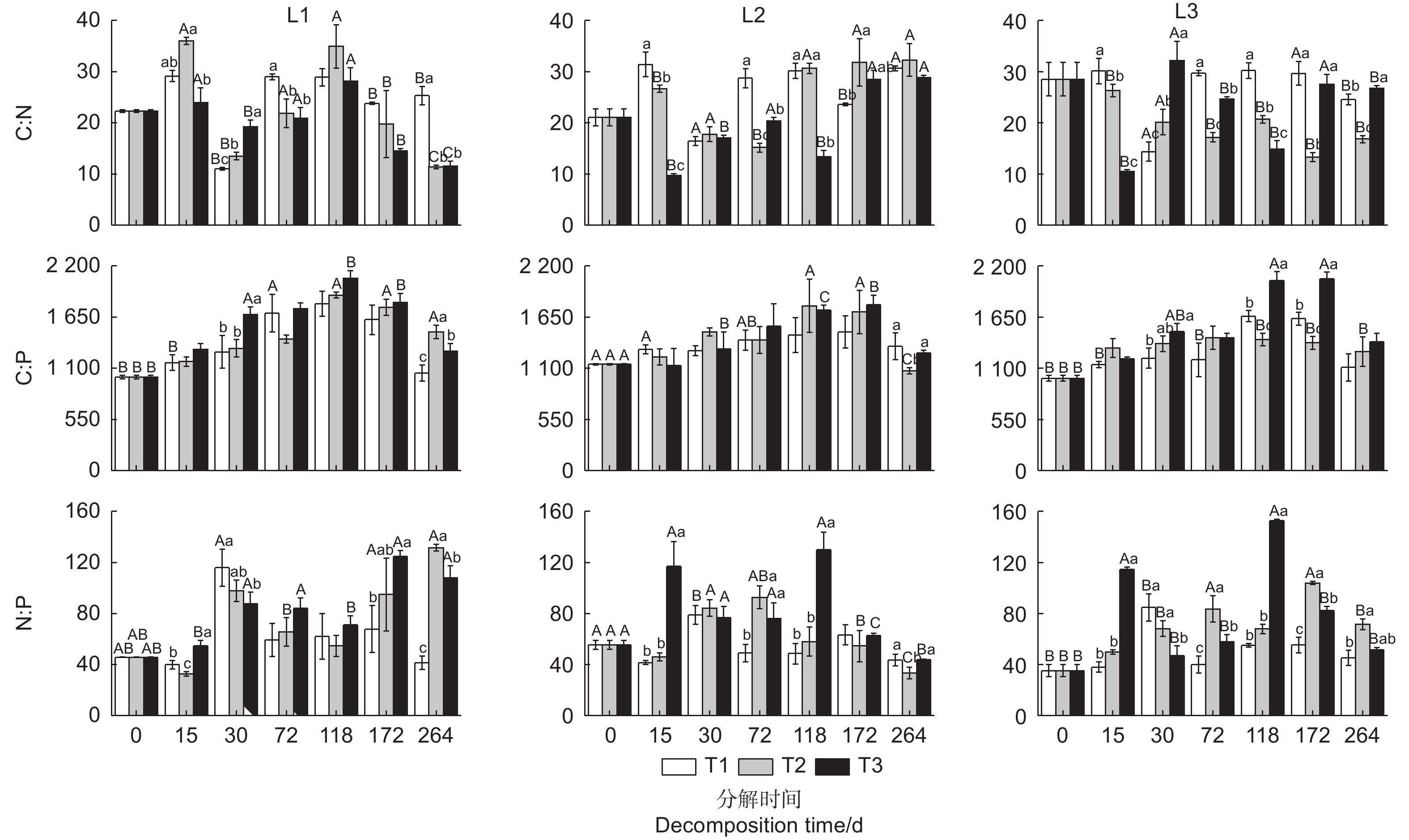
通过重复测量方差分析(表2)可知,温度、发育阶段及分解时间对凋落叶分解过程中C∶N、C∶P、 N∶P变化均有显著影响(除发育阶段对C∶P的影响外),温度 × 发育阶段交互效应对C∶N和 N∶P影响达极显著水平,发育阶段 × 分解时间交互效应对C∶N影响极显著,温度 × 分解时间、温度 × 发育阶段 × 分解时间交互效应对C∶N、C∶P和 N∶P影响极显著。
不同发育阶段杉木凋落叶分解过程中C∶N呈波动变化,C∶N变幅为9.32~39.0(图2)。各发育阶段凋落叶分解过程中均表现为25 ℃处理下C∶N明显大于35 ℃处理(除中龄林凋落叶分解第30 d,成熟林凋落叶分解30、172 d,过熟林凋落叶分解30、264 d外)。其中,中龄林凋落叶分解第72、264 d,成熟林分解第15、72 d、118 d,过熟林分解第15、72、118 d时均表现为25 ℃处理C∶N显著高于35 ℃处理(P < 0.05)。

图 2 不同温度下不同发育阶段杉木凋落叶分解过程中化学计量比的变化
Figure 2. Changes of stoichiometric ratios of leaf litter among different developmental Chinese fir plantations under different temperature treatments
各发育阶段杉木凋落叶分解在0~172 d时C∶P逐渐增大,172~264 d时C∶P逐渐减小,且凋落叶分解过程中C∶P变幅在949~2 194之间(图2)。不同发育阶段凋落叶分解过程中C∶P均表现35 ℃处理大于25 ℃处理(除成熟林凋落叶分解15、30、264 d外),其中,中龄林凋落叶分解30、264 d,过熟林分解30 、118 、172 d时两个温度处理间差异达显著水平(P < 0.05)。
不同发育阶段杉木凋落叶分解过程中N∶P呈波动变化规律(图2),N∶P的变化范围在32.7~153。除中龄林分解30 d,成熟林分解30、172、264 d,过熟林分解30 d外,各发育阶段分解过程中表现为35 ℃处理下N∶P高于25 ℃处理,其中,中龄林凋落叶分解15、172、264 d,成熟林分解15、72、118 d,过熟林分解15、72、118、172 d时两个温度处理间差异达显著水平(P < 0.05)。
-
杉木凋落叶分解过程中C、N、P残留率及其化学计量特征之间存在明显的相关性(表4)。其中,杉木凋落叶分解过程中C与C∶N、C∶P呈极显著正相关(P < 0.01),与N呈显著负相关(P < 0.05);N与C∶N呈极显著负相关,与 N∶P呈极显著正相关;P与C∶P、 N∶P呈极显著负相关; N∶P与C∶N呈极显著负相关,与C∶P呈极显著正相关。
表 4 杉木凋落叶分解过程中C、N、P残留率及其化学计量特征之间的相关性
Table 4. Correlation analysis between C, N and P residual rates and their stoichiometric ratios in leaf litter of Chinese fir plantations during decomposition process
指标 Indicator C N P C∶N C∶P N∶P C 1.000 N −0.163* 1.000 P −0.108 −0.035 1.000 C∶N 0.266** −0.944** 0.039 1.000 C∶P 0.404** −0.038 −0.918** 0.066 1.000 N∶P −0.084 0.859** −0.486** −0.821** 0.427** 1.000 注/Notes:* P < 0.05;**P < 0.01 -
凋落物分解是森林生态系统养分归还的主要途径,包括粉碎、淋溶和有机物的分解代谢3个过程。本研究发现,杉木凋落叶C素在研究期表现为释放—富集—释放的过程,对于中龄林和成熟林而言,升温处理对分解前172 d杉木凋落叶C残留率影响普遍不显著,但升温处理(35 ℃)在分解172、264 d时凋落叶C残留率显著低于25 ℃处理(图1),说明升温加快了这两个发育阶段杉木凋落叶C素的释放。在凋落物分解0~172 d内,主要是可溶性成分淋溶损失与易分解化合物的降解过程,受降雨淋溶冲刷影响较大,因此温度变化对此分解过程中C释放的影响并不明显,而在分解172 d后,温度升高提高了土壤微生物活性,显著加快了凋落物分解和C释放。这与研究中发现气候变暖导致大陆性气候的草地和针叶林在分解后期C释放更快的结果相似[18]。
本研究中N素在分解过程中表现为先富集后释放的变化规律(图1),这可能是因为凋落物分解过程中微生物对N需求量较高,分解前15 d内凋落叶中N不能满足微生物生长需求,需要从外界环境中吸收N,表现为N富集;但随着时间推移微生物持续分解含N有机物,逐渐能够满足自身对N的需要,进而表现为N释放。但不同发育阶段杉木凋落叶N释放的快慢不同,中龄林凋落叶分解第72、172、264 d,成熟林与过熟林分解第15、30、72 d时,N残留率表现为25 ℃处理显著低于35 ℃处理(P < 0.05),这与姜沛沛等[19]研究发现凋落叶分解过程中N素含量随温度增高而增大结果相似。
本研究发现分解过程中P表现为先释放后富集的规律,在分解30 d前呈明显快速释放状态,30~172 d时P呈缓慢释放的状态(图1)。这可能是由于热带与亚热带森林土壤P匮乏,且易被酸性土壤的铁铝氧化物固定,土壤中有效磷偏低导致微生物可利用P不足[20],因此凋落物分解过程中微生物很难从环境获取P,需要从凋落叶中获取P,从而使P残留率快速下降。本研究中,中龄林凋落叶分解第264 d,成熟林分解第118、172 d,过熟林分解第30、118、172、264 d时均表现为35 ℃处理下P残留率显著小于25 ℃处理,说明升温促进了这些林分凋落叶中P的释放,这与温度升高导致凋落叶分解过程中微生物或磷酸酶活性的提高有关[21]。
-
凋落叶C、N、P化学计量比的动态变化反映了凋落物的分解速率、土壤中N、P供应情况以及植物对养分的重吸收水平[22]。一般认为,凋落叶C∶N越小,凋落叶的N含量越高,难分解化合物较少,凋落物分解速率更快[23]。本研究发现,不同发育阶段杉木凋落叶分解过程中C∶N变幅为9.32~39.0(图2),低于凋落物分解研究中报道的凋落物C∶N临界阈值(40)[24],间接说明在整个凋落叶分解过程中基本可以满足微生物对N的需求,从而表现出N释放。本研究中,温度升高处理(35 ℃)的C∶N大多小于25 ℃处理,其中中龄林凋落叶分解第72、264 d、成熟林分解15、72、118 d、过熟林分解15、72、118 d时C∶N在这两个温度处理下差异显著(P < 0.05)。温度升高引起凋落叶分解的相关微生物及酶活性升高,但不同微生物类群对温度的响应存在差异,升温可能导致分解C有机物的微生物活性明显高于分解N的微生物活性,致使凋落物分解过程中C释放量大于N释放量,从而使升温处理下凋落叶C∶N明显降低。
植物对P的利用效率可以通过C∶P来衡量[25],本研究中杉木凋落叶分解过程中C∶P变幅在949~2 194之间(图2),明显高于C∶P释放临界阈值(600)[24],说明凋落叶分解过程中P的含量明显偏低,这与Chen等[26]对亚热带地区凋落物化学计量特征的研究结果一致。除了成熟林分解15、30、264 d外,各发育阶段凋落叶分解过程中C∶P均表现35 ℃处理大于25 ℃处理,这与Zhang等[25]报道的不同生态系统成分和不同生态系统结构中C∶P随温度升高而增大的研究结果一致,即增温提高了凋落叶磷酸酶活性,加快了凋落叶中P的释放,导致凋落叶C∶P增大。
N∶P是判断生态系统受N或P限制的敏感性指数,研究发现当凋落物 N∶P的大于25或其P含量低于0.22 g·kg−1时,凋落物 N∶P值越大,说明凋落物分解受到P的限制越强[27-28]。本研究中杉木凋落叶分解过程中 N∶P变幅为32.7~153(图2),均高于临界值25,说明各发育阶段杉木凋落叶分解过程中均受到P的限制。除中龄林和过熟林凋落叶分解第30 d,成熟林分解第30、172、264 d外,各发育阶段凋落叶分解过程中 N∶P随温度升高(35 ℃)而增大,这与王书丽等[15]研究发现不同年龄杉木针叶分解过程中 N∶P随温度升高而增大的结果一致。这可能与温度升高提高了分解过程中微生物对凋落叶中P的利用导致P大量释放,从而使 N∶P增大,这与分解过程前期P呈释放状态的结果一致。
-
不同发育阶段杉木凋落叶分解过程中C、N、P释放率及其化学计量特征差异明显,与自身凋落叶初始基质质量密切相关。本研究中成熟林杉木凋落叶初始C、N含量最高、P含量最低(表1)。成熟林杉木比中龄林阶段生长发育要相对缓慢,新陈代谢强度减弱,养分循环速率下降[29],则凋落叶C、N富集且含量偏高。成熟林凋落叶初始P含量显著低于中龄林和过熟林阶段(P < 0.05),这与邱勇斌等[30]研究结果相似,即P含量没有按照林龄增加而呈下降趋势。成熟林阶段土壤中P被前期生长所消耗,为保持旺盛生长的需求,P可通过叶片的养分重吸收而被再利用,从而导致凋落叶中P含量明显偏低;中龄林杉木处于快速生长阶段,所需P主要来源于土壤,叶片的养分重吸收作用不强,凋落叶P含量较高;而过熟林杉木处于衰老阶段,缓慢的新陈代谢速率导致养分需求量低,而林地养分的主要来源仍是凋落物的分解,因此过熟林杉木更倾向于将养分分配给凋落物,通过分解过程释放给土壤,从而保证了凋落叶中相对较高的P含量。
除不同发育阶段凋落叶自身属性的差异外,外界温度的升高对不同发育阶段凋落叶分解过程中C、N、P的释放及化学计量比也产生重要影响。本研究发现升温明显缩短了成熟林与过熟林凋落叶N和P的周转期,35 ℃处理下N周转期比25 ℃处理分别缩短了34.4%和16.9%,P周转期分别缩短了38.4%和43.8%(表3)。通过对温带森林长期增温实验表明,升温能显著提高土壤有机氮矿化速率和硝化速率,从而促进凋落叶中N释放[31]。也有研究发现增温能够显著提高土壤酸性磷酸酶的活性,使有机磷转化为无机磷,能更好地满足微生物对P的需求,加快凋落物分解,从而缩短了凋落叶—土壤—植物间P的周转期[32]。另外,本研究发现,中龄林、成龄林和过熟林凋落叶分解过程中 N∶P变幅分别为32.7~131、33.4~130、37.9~153(图2),说明不同发育阶段杉木凋落叶分解过程中受P限制的程度不同,表现为过熟林阶段明显高于中龄林和成熟林阶段。这可能是由于杉木人工林生长到过熟林阶段,土壤中有效P已经过度消耗[33],土壤微生物活性受到强烈的P限制,导致凋落物分解中P释放加快,从而使 N∶P相对增大。可见,P不足是影响不同发育阶段杉木生长发育的重要限制因子,为了实现杉木大径材的培育目标,建议可根据杉木各发育阶段的生长需求,适当增加P肥的施用,对提高杉木人工林长期生产力水平具有重要的指导意义。
-
通过室内模拟升温试验,研究了温度升高对不同发育阶段杉木凋落叶分解过程中C、N、P残留率及其化学计量特征的影响,发现:(1)升温能够加速成熟林、过熟林杉木凋落叶N、P的周转速率;(2)升温总体增加了各发育阶段杉木凋落叶分解过程中的C∶P、 N∶P,降低了C∶N;杉木凋落叶分解过程中C∶P, N∶P均明显大于临界阈值(600和25),说明杉木凋落叶分解过程明显受P限制;(3)杉木凋落叶分解过程中C、N、P的释放及其化学计量特征受温度影响明显,而凋落叶的初始基质质量也是其重要的影响因素。开展凋落物分解对升温的响应研究,有利于阐明未来全球气候变化背景下,杉木人工林生态系统中凋落叶—土壤—植物的养分周转平衡,为杉木人工林生产力维持提供科学指导。然而,凋落物分解往往与微生物、土壤性质密切相关,建议未来结合植物、土壤的化学计量特征以及野外试验,多尺度、多维度、全方位揭示杉木人工林生态系统对全球气候变暖的响应规律。
不同温度下杉木凋落叶分解过程中碳氮磷释放及其化学计量比变化
Carbon, Nitrogen and Phosphorus Release and Their Stoichiometric Ratio Change During Decomposition of Cunninghamia lanceolata Leaf Litter at Different Temperatures
-
摘要:
目的 研究升温对不同发育阶段杉木凋落叶分解过程中碳(C)、氮(N)、磷(P)释放规律及其生态化学计量特征的影响,揭示气候变暖背景下杉木人工林凋落叶分解过程中养分释放特征。 方法 收集中龄林(18年生)、成熟林(30年生)和过熟林(42年生)3个发育阶段的杉木凋落叶,设置25、30和35 ℃3个温度梯度进行室内模拟分解试验。 结果 (1)在264 d的分解周期内,各发育阶段杉木凋落叶C、N、P残留率总体表现为随分解时间的增加而减小,但不同元素残留率变化模式不同,C残留率表现为释放—富集—释放模式,N残留率表现为富集—释放模式,P残留率表现为释放—富集模式。(2)拟合模型结果表明,成熟林与过熟林凋落叶分解过程中N周转期在35 ℃处理下比25 ℃处理分别缩短了34.4%和16.9%,P周转期分别缩短了38.4%和43.8%。(3)不同发育阶段杉木凋落叶分解过程中C∶N、 N∶P比总体呈波动变化,C∶P比呈先增大后减小的变化趋势,杉木凋落叶C∶N、C∶P、 N∶P比变幅分别为9.32~39.0、949~2 194、32.7~153,升温处理在凋落叶分解过程总体上增大了C∶P、 N∶P,降低了C∶N比。 结论 温度升高能够缩短成熟林、过熟林凋落叶分解过程中N、P的周转期,提高各发育阶段杉木凋落叶分解过程中的C∶P、 N∶P,说明杉木凋落叶分解明显受P限制,建议根据不同发育阶段杉木的生长需求,适当增加磷肥的施用。 Abstract:Objective To explore the effects of rising temperature on the release of carbon (C), nitrogen (N) and phosphorus (P) during leaf litter decomposition of Chinese fir at different developmental stages and their stoichiometric variations under the scenarios of climate warming. Methods The leaf litter of Chinese fir at three developmental stages, i.e. middle-aged forest (18-years old), mature forest (30-years old) and overmature forest (42-years old), was collected for decomposition experiments at three temperature treatments (i.e., 25, 30 and 35 ℃). Results (1) During the 264-day decomposition period, the residual rates of C, N and P in leaf litter at each development stage generally decreased over time. The residual rates of C, N and P showed patterns with release-enrichment-release, release-release, and release-enrichment, respectively. (2) The results of Olson fitting model revealed that the N turnover time of mature forest and over-mature forests during litter decomposition was shortened by 34.4 and 16.9%, respectively under 35 ℃ compared with 25 ℃. In comparison, the P turnover time was shortened by 38.4% and 43.8%, respectively. (3) During litter decomposition, the ratios of C∶N and N∶P generally fluctuated, and C∶P increased first and then decreased. The variation of C∶N, C∶P and N∶P ratios were 9.32-39.0, 949-2 194 and 32.7-153, respectively. In the decomposition process of leaf litter, rising temperature increased C∶P, N∶P, and decreased C∶N. Conclusion Rising temperature can shorten the turnover rate of N and P during litter decomposition in mature and overmature forests by increasing C∶P and N∶P ratios during litter decomposition, indicating that litter decomposition of Chinese fir is obviously limited by P. It is suggested to appropriately increase the application of P fertilizer according to the growth needs of Chinese fir at different developmental stages. -
图 1 不同温度下不同发育阶段杉木凋落叶分解过程中C、N、P残留率的变化
Figure 1. Changes of the residual rates of C,N and P of leaf litter among different developmental Chinese fir plantation under different temperature treatments
图 2 不同温度下不同发育阶段杉木凋落叶分解过程中化学计量比的变化
Figure 2. Changes of stoichiometric ratios of leaf litter among different developmental Chinese fir plantations under different temperature treatments
表 1 不同发育阶段杉木凋落叶初始化学性质
Table 1. Initial chemical properties of Chinese fir leaf litter among different developmental stages
发育阶段
Developmental stageC含量
C content/(g·kg−1)N含量
N content/(g·kg−1)P含量
P content/(g·kg−1)中龄林(18 a) 489 ± 1.370 A 19.9 ± 0.372 A 0.487 ± 0.013 3 A 成熟林(30 a) 493 ± 0.142 A 19.9 ± 0.698 A 0.432 ± 0.014 6 B 过熟林(42 a) 490 ± 0.211 A 19.3 ± 1.030 A 0.503 ± 0.013 5 A 注:同列不同大写字母表示不同发育阶段凋落叶间差异显著(P < 0.05)
Note: Uppercase letter in the same column showed significant difference between different developmental stages (P < 0.05) 下载: 导出CSV
下载: 导出CSV
表 2 温度、发育阶段、分解时间及其交互效应对杉木凋落叶C、N、P残留率及其化学计量特征的影响(F值)
Table 2. Effects of temperature, developmental stage, decomposition time and their interaction effects on C, N and P residual rates and stoichiometric characteristics of Chinese fir litters (F value)
指标 Indicator C N P C∶N C∶P N∶P TE 1.78 82.00** 7.49** 45.00** 9.09** 39.40** D 0.54 11.30** 1.66 6.12** 1.00 4.53* T 58.90** 37.20** 53.10** 56.10** 54.30** 17.90** TE × D 5.44** 8.50** 0.78 14.50** 0.62 21.50** D × T 6.36** 30.00** 1.90* 54.10** 1.05 1.65 TE × T 10.20** 65.70** 1.27 39. 80** 3.85** 26.00** TE × D × T 4.89** 10.50** 2.82** 7.21** 2.92** 9.26** 注:* P < 0.05;**P < 0.01;TE-温度;D-发育阶段;T-分解时间
Notes: * P < 0.05; **P < 0.01; TE- temperature; D- developmental stage; T- decomposition time
下载: 导出CSV
表 3 不同温度处理下各发育阶段杉木凋落叶C、N、P平均分解速率
Table 3. Average decomposition rates of C, N and P of leaf litter among different developmental stages under different temperature treatments
元素
Element温度处理
Temperature
treatment发育阶段
Developmental
stage分解方程
Decomposition
equation分解系数
Decomposition
coefficient95%分解所需时间
95% decomposition
time/a相关系数
Correlation
coefficientP值
P valueC 25 ℃ 中龄林 y=56.3e−0.412t 0.412 7.27 0.347 P > 0.05 成熟林 y=56.7e−0.391t 0.391 7.66 0.334 P > 0.05 过熟林 y=57.4e−0.378t 0.378 7.93 0.337 P > 0.05 30 ℃ 中龄林 y=57.0e−0.383t 0.383 7.82 0.338 P > 0.05 成熟林 y=59.8e−0.506t 0.506 5.92 0.464 P < 0.05 过熟林 y=56.7e−0.465t 0.465 6.44 0.390 P > 0.05 35 ℃ 中龄林 y=58.3e−0.456t 0.456 6.57 0.401 P > 0.05 成熟林 y=56.4e−0.379t 0.379 7.91 0.327 P > 0.05 过熟林 y=57.2e−0.340t 0.340 8.81 0.303 P > 0.05 N 25 ℃ 中龄林 y=81.3e−0.905t 0.905 3.31 0.569 P < 0.01 成熟林 y=80.1e−0.945t 0.945 3.17 0.666 P < 0.01 过熟林 y=76.8e−0.778t 0.778 3.85 0.537 P < 0.05 30 ℃ 中龄林 y=108 e−1.39t 1.390 2.16 0.775 P < 0.01 成熟林 y=86.9e−1.32t 1.320 2.27 0.829 P < 0.01 过熟林 y=94.0e−0.737t 0.737 4.07 0.957 P < 0.01 35 ℃ 中龄林 y=93.5e−0.710t 0.710 4.22 0.708 P < 0.01 成熟林 y=114. e−1.44t 1.440 2.08 0.838 P < 0.01 过熟林 y=93.2e−0.936t 0.936 3.20 0.601 P < 0.01 P 25 ℃ 中龄林 y=.79.7e−0.695t 0.695 4.31 0.614 P < 0.05 成熟林 y=75.0e−0.462t 0.462 6.48 0.589 P < 0.01 过熟林 y=74.6e−0.503t 0.503 5.96 0.513 P < 0.05 30 ℃ 中龄林 y=73.5e−0.809t 0.809 3.70 0.711 P < 0.01 成熟林 y=75.2e−0.479t 0.479 6.25 0.464 P < 0.01 过熟林 y=72.1e−0.738t 0.738 4.06 0.654 P < 0.05 35 ℃ 中龄林 y=65.2e−0.701t 0.701 4.27 0.555 P < 0.05 成熟林 y=73.7e−0.751t 0.751 3.99 0.553 P < 0.05 过熟林 y=65.53e−0.895t 0.895 3.35 0.561 P < 0.05
下载: 导出CSV
表 4 杉木凋落叶分解过程中C、N、P残留率及其化学计量特征之间的相关性
Table 4. Correlation analysis between C, N and P residual rates and their stoichiometric ratios in leaf litter of Chinese fir plantations during decomposition process
指标 Indicator C N P C∶N C∶P N∶P C 1.000 N −0.163* 1.000 P −0.108 −0.035 1.000 C∶N 0.266** −0.944** 0.039 1.000 C∶P 0.404** −0.038 −0.918** 0.066 1.000 N∶P −0.084 0.859** −0.486** −0.821** 0.427** 1.000 注/Notes:* P < 0.05;**P < 0.01
下载: 导出CSV
-
[1] 贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论[J]. 植物生态学报, 2010, 34(1):2-6. doi: 10.3773/j.issn.1005-264x.2010.01.002 [2] ZHANG J H, HE N P, LIU C C, et al. Allocation strategies for nitrogen and phosphorus in forest plants[J]. Oikos, 2018, 127(10): 1506-1514. doi: 10.1111/oik.05517 [3] KRISHNA M P, MOHAN M. Litter decomposition in forest ecosystems: a review[J]. Energy, Ecology and Environment, 2017, 2(4): 236-249. doi: 10.1007/s40974-017-0064-9 [4] 崔 雪, 王海燕, 邹佳何, 等. 长白山针阔混交林凋落物-土壤生态化学计量特征[J]. 林业科学研究, 2023, 36(3):91-99. doi: 10.12403/j.1001-1498.20220226 [5] TONG R, ZHOU B Z, JIANG L N, et al. Leaf litter carbon, nitrogen and phosphorus stoichiometry of Chinese fir (Cunninghamia lanceolata) across China[J]. Global Ecology and Conservation, 2021, 27: e1542. [6] 宋 飘, 张乃莉, 马克平, 等. 全球气候变暖对凋落物分解的影响[J]. 生态学报, 2014, 34(6):1327-1339. [7] 田 地, 严正兵, 方精云. 植物生态化学计量特征及其主要假说[J]. 植物生态学报, 2021, 45(7):682-713. [8] REN H Y, QIN J, YAN B L, et al. Mass loss and nutrient dynamics during litter decomposition in response to warming and nitrogen addition in a desert steppe[J]. Frontiers of Agricultural Science and Engineering, 2018, 5(1): 64-70. doi: 10.15302/J-FASE-2017194 [9] YANG Q P, LI R, S ZHANG W D, et al. Decomposition of harvest residue needles of different needle ages in a Chinese fir (Cunninghamia lanceolata) plantation[J]. Plant and soil, 2018, 423(1-2): 273-284. doi: 10.1007/s11104-017-3515-3 [10] 国家林业和草原局. 中国森林资源报告(2014-2018)[M]. 北京: 中国林业出版社. 2019, 28-29. [11] ZHOU L L, SHALOM A D D, WU P F, et al. Litterfall production and nutrient return in different-aged Chinese fir (Cunninghamia lanceolata) plantations in South China[J]. Journal of Forestry Research, 2015, 26(1): 79-89. doi: 10.1007/s11676-014-0011-y [12] 吴小健, 石 焱, 朱嘉宁, 等. 全球变暖对森林凋落物的影响[J]. 世界林业研究, 2023, 36(01):26-32. doi: 10.13348/j.cnki.sjlyyj.2022.0079.y [13] 李澳归, 林成芳, 胡明艳, 等. 增温对亚热带杉木枝和叶凋落物理化性质的影响[J]. 应用生态学报, 2022, 33(10):2711-2717. [14] 杨成邦, 张 丽, 高艳丽, 等. 增温对湿润亚热带杉木幼林和成熟林土壤无机氮的影响[J]. 应用生态学报, 2020, 31(9):2849-2856. [15] 王书丽, 黄立君, 袁 希, 等. 氮添加和升温对杉木林凋落物分解及碳氮磷化学计量特征的影响[J]. 生态学杂志, 2020, 39(9):2842-2850. [16] 马志良, 高 顺, 杨万勤, 等. 不同季节亚热带常绿阔叶林6个树种凋落叶钙、镁、锰的释放特征[J]. 应用生态学报, 2015, 26(10):2913-2920. [17] OLSON S J. Energy Storage and the balance of producers and decomposers in ecological systems[J]. Ecology, 1963, 44(2): 322-331. doi: 10.2307/1932179 [18] ZHANG F, FENG Y H, SONG S S, et al. Temperature sensitivity of plant litter decomposition rate in China's forests[J]. Ecosphere, 2021, 12(6): e03541. doi: 10.1002/ecs2.3541 [19] 姜沛沛, 曹 扬, 陈云明, 等. 陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征[J]. 生态学报, 2017, 37(2):443-454. [20] 陈嘉琪, 赵光宇, 李仰龙, 等. 杉木人工林土壤磷素形态及含量的林龄变化[J]. 林业科学, 2022, 58(5):10-17. [21] 程 严, 列志旸, 刘旭军, 等. 增温对南亚热带针阔叶混交林凋落物分解酶活性的影响[J]. 应用与环境生物学报, 2021, 27(4):923-929. [22] 李明军, 喻理飞, 杜明凤, 等. 不同林龄杉木人工林植物-凋落叶-土壤C、N、P化学计量特征及互作关系[J]. 生态学报, 2018, 38(21):7772-7781. [23] 张乃木, 王克勤, 宋娅丽, 等. 滇中亚高山森林林下植被和凋落物生态化学计量特征[J]. 林业科学研究, 2020, 33(4):127-134. [24] PARTON W, SILVER W L, BURKE I C, et al. Global-scale similarities in nitrogen release patterns during long-term decomposition[J]. Science, 2007, 315(5810): 361-364. doi: 10.1126/science.1134853 [25] ZHANG J, ZHAO N, LIU C, et al. C∶ N∶ P stoichiometry in China's forests: from organs to ecosystems[J]. Functional Ecology, 2018, 32(1): 50-60. doi: 10.1111/1365-2435.12979 [26] CHEN B, CHEN L, JIANG L, et al. C∶ N∶ P stoichiometry of plant, litter and soil along an elevational gradient in subtropical forests of China[J]. Forests, 2022, 13(3): 372. doi: 10.3390/f13030372 [27] YUE K, FORNARA D A, YANG W, et al. Effects of three global change drivers on terrestrial C∶ N∶ P stoichiometry: a global synthesis[J]. Global Change Biology, 2017, 23(6): 2450-2463. doi: 10.1111/gcb.13569 [28] 许宇星, 王志超, 张丽丽, 等. 不同种植年限尾巨桉人工林叶片-凋落物-土壤碳氮磷化学计量特征[J]. 林业科学研究, 2018, 31(6):168-174. [29] 陈日升, 康文星, 周玉泉, 等. 杉木人工林养分循环随林龄变化的特征[J]. 植物生态学报, 2018, 42(2):173-184. [30] 邓博文, 许瑶瑶, 陈逸飞, 等. 中国针叶林优势树种叶片氮磷钾生态化学计量特征及内稳态分析[J]. 林业科学研究, 2020, 33(6):81-87. doi: 10.13275/j.cnki.lykxyj.2020.06.010 [31] MELILLO J M, BUTLER S, JOHNSON J, et al. Soil warming, carbon–nitrogen interactions, and forest carbon budgets[J]. Proceedings of the National Academy of Sciences, 2011, 108(23): 9508-9512. doi: 10.1073/pnas.1018189108 [32] GAO J, WANG E, REN W, et al. Effects of simulated climate change on soil microbial biomass and enzyme activities in young Chinese fir (Cunninghamia lanceolata) in subtropical China[J]. Acta Ecologica Sinica, 2017, 37(4): 272-278. doi: 10.1016/j.chnaes.2017.02.007 [33] 曹 娟, 闫文德, 项文化, 等. 湖南会同3个林龄杉木人工林土壤碳、氮、磷化学计量特征[J]. 林业科学, 2015, 51(7):1-8. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1816
- HTML全文浏览量: 909
- PDF下载量: 64
- 被引次数: 0



