



-
红树林分布于热带和亚热带海湾、河口滩涂上以红树植物为主的常绿乔木、灌木组成的木本植物群落,具有促淤防浪护岸,护堤固滩、促淤固滩、净化水体、美化景观、维持湿地生物多样性以及固碳储碳等重要生态功能[1-2]。温度是限制红树植物地理分布的重要因素[3]。中国红树林植物种类随纬度的升高和平均温度的下降而减少,其中,最南的海南省红树植物种类有24种,而人工引种最北界浙江仅有1种[4]。近些年来,全球变暖已经成为不争的事实,这势必对红树林生长产生影响。红树林是嗜热植物,主要分布在南北半球25 ℃等温线内,而以全球变暖为主要特征的全球气候变化可促进红树林向更高纬度引种,目前红树林的自然分布北界可由现在的福建福鼎北扩至浙江,甚至已经开始在杭州尝试种植[5]。然而,另一方面,因气候变化带来的频繁极端低温灾害又对红树林生态系统造成了极大的伤害和严重威胁,尤其是生长最北缘的红树植物[6-7]。多年引种经验表明,限制我国高纬度地区红树分布的主要因素是温度,其中极端最低温度是主要限制因素之一[4],这主要表现在极端低温会引起红树植物光合器官受损,抗氧化系统失衡,光合电子传递受阻,生长发育停滞,叶片发黄脱落,植株营养物质损失,成熟植株提前落花落果,甚至死亡等[8-10],详见图1。因此,解析红树林对低温胁迫响应与适应机制已成为提高红树林抗寒性的重要课题。

图 1 2015年不同树龄红树林极端低温伤害症状
Figure 1. The symptoms of extremely cold events damage on mangroves in 2015
秋茄(Kandelia obovate Sheue et al.)为红树科秋茄树属常绿灌木或小乔木,是我国境内分布最广、纬度分布最高、最耐寒的红树植物,在我国引种成林的最北缘为浙江西门岛[11]。红榄李(Lumnitzera littore (Jack.)Voigt)是使君子科榄李属的红树植物,主要分布于东非到西太平洋、澳大利亚亚热带地区和中南半岛,目前我国野生种数量仅有9株,唯一分布地在三亚铁炉港[12]。红榄李生长于易受人为干扰的高潮带,是嗜热窄分布的濒危红树植物 [13-14]。研究认为,红榄李濒危的主要原因主要包括:种群遗传性多样性较低,野生个体数量较少,有性生殖存在障碍等[15]。一般来说,红榄李对温度的要求也较为苛刻。据调查,红榄李仅能生长在年平均温度为21~25 ℃,且全年无霜的滩涂上,这导致红榄李对低温的影响较为敏感。目前,对红树植物低温胁迫的研究主要集中在秋茄、白骨壤、桐花树等[16-17]上,而将我国分布最南(低温敏感)的红榄李与最北(耐寒)秋茄置于相同低温下比较其对温度耐性的生理机制研究还报道甚少。本研究以低温敏感型红榄李和耐寒型秋茄幼苗为试验材料,通过比较分析两种红树植物在不同低温时间处理下的光合生理参数、荧光参数、光合色素含量、活性氧含量、抗氧化系统中关键酶活性以及叶绿体超微结构、气孔开放特征等变化,探讨两种红树植物响应低温的光合生理和抗氧化特性差异,以期为我国红树林资源保护与抗寒机制研究提供理论依据。
-
2021年10月初,分别将来自福建漳江口红树林国家级自然保护区(117°24′~117°30′ E,23°53′~23°56′ N)和三亚铁炉港红树林自然保护区(109°42′~109°44′ E,18°15′~18°17′ N)已培育1 a的健壮秋茄、红榄李幼苗(生长在规格为13 cm × 13 cm营养杯中)装入高30 cm、直径为20 cm的聚乙烯塑料桶内。小心剪开营养杯的底部,使幼苗根系能够直接接触塑料桶底部,加入一定量的Hoaglandʼs营养液,使塑料桶底部营养液3 cm左右深。将其摆放在人工气候室(昼夜温度28 ℃/25 ℃)内培养,适应恢复15 d。恢复期间每2 d更换一次Hoaglandʼs营养液。
-
选择大小一致的秋茄和红榄李幼苗,移至昼夜温度为25 ℃ /20 ℃,光照周期为12 h /12 h,光强为 400 μmol·m−2·s−1,相对湿度为65%~70%的智能培养箱内,适应10 d。随后将两种红树植物幼苗均分成2组,一组仍保留在温度25 ℃ (昼) /20 ℃ (夜)的光照培养箱内,作为对照;另一组移入温度为7 ℃(昼)/4 ℃(夜)的冷光源植物生长箱(DGX-260E)进行低温胁迫处理(简称为LS),在低温处理24、48 h时,分别取对照和两种红树植物幼苗顶端完全展开的倒三对叶片进行光合、荧光以及生理指标测定;每个处理3盆,3次重复。
-
使用Li-6400型便携式光合仪于9:30—11:30 测定顶端完全展开的倒三对叶片的净光合速率(Pn)、细胞间隙CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率参数。测定时设定内置红蓝光源光强为400 μmol·m−2s−1,CO2浓度均为(410 ± 3)μmol·mol−1,气体流速设为500 μmol·s−1。气孔限制值(Ls)计算公式为:Ls=1−Ci/Ca
-
使用FMS2调制式荧光测定仪(英国Hansatech公司)进行叶片叶绿素荧光参数测定。测定时先测定光适应下的稳态荧光(Fs)、最大荧光(Fm’)等参数,暗环境下适应30 min后,测定初始荧光(Fo)、最大荧光(Fm)和PSII最大光化学效率(Fv/Fm)。根据Yuan等[18]的方法计算PSII实际光化学效率(ФPSII)、光化学猝灭系数(qP)以及非光化学猝灭系数(NPQ)。
-
称取0.1 g秋茄、红榄李幼苗叶片剪碎放入25 mL的1:1无水乙醇和丙酮提取液中。置于黑暗环境下提取至叶片发白,测定提取液在665、649和470 nm处的吸光值,参照李合生[19]方法计算叶绿素a、叶绿素b、总叶绿素以及类胡萝卜素含量。
-
分别取1.0 g秋茄、红榄李幼苗叶片,加3 mL 50 mmol·L−1 pH7.5 Tris-HCL缓冲液后进行冰浴研磨,离心30 min,取上清液待测。用氯化硝基四氮唑蓝(NBT)法测定SOD活性[20],采用愈创木酚法测定POD活性[20];按Du和Bramlage[21]方法测定MDA含量。
-
分别取各实验小组叶片进行裁剪为长条形1 × 2 mm(避开主脉),将取样叶片正反面标记进行真空处理,确保样品沉入固定液中,其中叶绿体透射电镜标本采用5%戊二醛固定液,气孔扫描电镜标本选用2.5%的戊二醛固定液。使用JEOL JEM-1230透射电镜和Zeiss Gemini 300扫描电镜进行样本观察。
测量之前先用图片中的10 µm微尺定标,利用ImageJ软件测量单个气孔的气孔长、气孔宽,并按照张翠等[24]方法计算气孔开度。同时,利用软件中的polygon工具测量气孔面积。每个处理测量3个样本,每个样本测量10个视野。
-
通过SPSS 10.0 统计软件进行单因素方差分析(One-way ANOVA)进行数据比较,采用Duncanʼs法检验处理间差异的显著水平(P<0.05)。图中数据均为平均值 ± 标准偏差,通过SigmaPlot 10.0绘图软件作图。
-
由图2可知,在低温胁迫48 h时,秋茄幼苗叶片并未产生明显的变化,而同样低温处理下红榄李幼苗叶片出现萎蔫现象,即叶片明显下垂。

图 2 低温胁迫对两种红树植物幼苗生长情况的影响
Figure 2. Effects of low temperature stress on growth of two mangrove seedlings
由表1可知,秋茄和红榄李幼苗叶片净光合速率(Pn)均随着低温胁迫时间的增长而显著下降,甚至低温胁迫48 h处理下红榄李幼苗叶片Pn甚至降至−0.33 µmol·m−2 ·s−1。与对照相比,低温胁迫下两种红树植物叶片Pn下降幅度存在着较大的差异,且降低幅度随胁迫时间延长而增加。例如,与对照相比,低温胁迫48 h处理下,秋茄幼苗叶片Pn下降幅度约为82.1%,而红榄李幼苗叶片Pn下降幅度约为105.2%。此外,低温胁迫48 h处理下,秋茄幼苗叶片Pn下降幅度为红榄李约为79%,表明秋茄耐寒性较高于红榄李。
表 1 低温胁迫对两种红树植物幼苗叶片气体交换参数的影响
Table 1. Effects of low temperature stress on gas exchange parameters in leaves of two mangrove seedlings
红树植物
Mangrove plants处理
Treatment净光合速率
Pn/(μmol·m−2·s−1)气孔导度
Gs/(mmol·m−2·s−1)胞间CO2浓度
Ci/(μmol·mol−1)气孔限制值
Ls秋茄
K. obovata对照 6.38 ± 0.54 a 147.87 ± 0.93 a 317.03 ± 12.25 b 0.25 ± 0.03 c 24 h 3.41 ± 1.09 b 111.43 ± 0.30 b 376.99 ± 11.31 a 0.14 ± 0.05 d 48 h 1.14 ± 0.13 c 43.69 ± 0.72 d 249.14 ± 15.83 c 0.55 ± 0.10 a 红榄李
L. littore对照 6.30 ± 0.27 a 151.87 ± 6.84 a 252.80 ± 6.95 c 0.38 ± 0.01 b 24 h 0.19 ± 0.01 d 68.54 ± 8.79 c 322.45 ± 7.06 b 0.21 ± 0.02 c 48 h -0.33 ± 0.06 d 14.84 ± 3.20 d 391.10 ± 5.91 a 0.05 ± 0.01 e 注:同列不同小写字母表示不同处理间存在差异显著(P<0.05)。下同
Notes: Different lowercase letters of the same column indicate significant difference at 0.05 level. The same below不同低温胁迫处理下两种红树植物叶片气孔导度(Gs)变化趋势与Pn相似(表1)。与对照相比,低温胁迫24 h处理下,秋茄和红榄李幼苗叶片Gs分别下降25%和55%,而低温胁迫48 h处理下两种红树植物叶片Gs分别下降70%和90%,这表明红榄李幼苗叶片Gs对低温响应比秋茄更为敏感。
由表1可知,随低温处理时间延长秋茄幼苗叶片胞间CO2浓度(Ci)呈先增后降的趋势,而红榄李幼苗叶片Ci却逐渐增加。在低温胁迫24 h处理下,秋茄、红榄李幼苗叶片Ci为对照的1.19倍和1.28倍。然而,与对照相比,低温胁迫48 h处理下,秋茄幼苗叶片Ci降低了21%,而红榄李幼苗叶片Ci则增加了55%。
随低温胁迫时间延长,红榄李幼苗叶片气孔限制值(Ls)持续降低,而秋茄幼苗叶片气孔限制值呈先降后增的趋势(表1)。与对照相比,低温胁迫24 h处理下,秋茄、红榄李幼苗叶片Ls分别降低44%和45%;低温胁迫48 h处理下秋茄植物叶片Ls上升了120%,而红榄李植物叶片气孔限制值降低了87%。
-
由表2可知,随着低温胁迫的时间延长,秋茄和红榄李幼苗叶片Fv/Fm、ΦPSⅡ和qP均逐渐减少。低温胁迫下,红榄李幼苗叶片Fv/Fm、ΦPSⅡ和qP下降幅度均大于秋茄,且随着低温胁迫时间延长,各值降低幅度更大。与对照相比,低温胁迫24 h下,秋茄幼苗叶片Fv/Fm、ΦPSII、qP分别降低18%、21%以及25%,而红榄李幼苗叶片Fv/Fm、ΦPSⅡ和qP则分别降低32%、41%以及27%;低温胁迫48 h处理下,秋茄幼苗叶片Fv/Fm、ΦPSⅡ、qP分别降低39%、38%以及44%,而红榄李幼苗叶片Fv/Fm、ΦPSⅡ、qP分别降低达70%、77%以及51%。随着低温胁迫时间延长,秋茄幼苗叶片NPQ呈逐渐增加的趋势,而红榄李幼苗叶片NPQ却先增后降(表2)。与对照相比,低温胁迫24 h和48 h下,秋茄幼苗叶片NPQ分别增加26%和49%,低温胁迫24 h下红榄李幼苗叶片NPQ为对照1.28倍,而低温胁迫48 h处理下叶片NPQ为对照的36%。
表 2 低温胁迫对两种红树植物幼苗叶片叶绿素荧光参数的影响
Table 2. Effects of low temperature stress on chlorophyll fluorescence parameters of leaves of two mangrove seedlings
红树植物
Mangrove plants处理
TreatmentFv/Fm ΦPSⅡ qP NPQ 秋茄
K. obovata对照 0.85 ± 0.01 a 0.58 ± 0.04 a 0.69 ± 0.06 a 0.53 ± 0.29 cd 24 h 0.70 ± 0.02 b 0.46 ± 0.07 b 0.52 ± 0.09 b 0.67 ± 0.28 b 48 h 0.52 ± 0.02 c 0.36 ± 0.04 c 0.39 ± 0.06 c 0.79 ± 0.20 a 红榄李
L. littore对照 0.69 ± 0.01 b 0.56 ± 0.04 a 0.49 ± 0.06 b 0.47 ± 0.05 d 24 h 0.47 ± 0.04 c 0.33 ± 0.01 c 0.36 ± 0.04 c 0.60 ± 0.04 bc 48 h 0.21 ± 0.04 d 0.13 ± 0.03 d 0.24 ± 0.03 d 0.17 ± 0.04 e -
由表3可知,随低温胁迫延长秋茄和红榄李幼苗叶片叶绿素(Chl)含量、类胡萝卜素(Car)含量、Chl a/Chl b值均逐渐减少(表3)。低温处理下两种红树植物相比,秋茄幼苗叶片Chl含量、Car含量、Chl a/Chl b值以及Car/Chl值均比红榄李的值高。
表 3 低温胁迫对两种红树植物幼苗叶片光合色素的影响
Table 3. Effects of low temperature stress on photosynthetic pigments in leaves of two mangrove seedlings
红树植物
Mangrove plants处理
Treatment叶绿素含量
Chl content/(mg·g−1)类胡萝卜素含量
Car content/(mg·g−1)叶绿素a/叶绿素b比
Chl a/Chl b类胡萝卜素/叶绿素比
Car/Chl秋茄
K. obovata对照 1.14 ± 0.15 a 0.20 ± 0.01 a 3.05 ± 0.04 a 0.17 ± 0.01 b 24 h 0.96 ± 0.19 b 0.19 ± 0.01 a 2.85 ± 0.03 ab 0.20 ± 0.02 a 48 h 0.93 ± 0.05 b 0.16 ± 0.01 b 2.56 ± 0.08 b 0.17 ± 0.01 b 红榄李
L. littore对照 0.90 ± 0.12 b 0.09 ± 0.03 c 2.11 ± 0.14 c 0.10 ± 0.03 d 24 h 0.69 ± 0.04 c 0.08 ± 0.02 cd 1.81 ± 0.12 cd 0.11 ± 0.01 d 48 h 0.53 ± 0.04 d 0.07 ± 0.02 d 1.71 ± 0.04 d 0.13 ± 0.01 c 与对照相比,低温胁迫24 h和48 h处理下,秋茄幼苗叶片Chl含量分别下降16%和18%;低温胁迫24 h处理下,秋茄幼苗叶片叶片Car/Chl值上升18%,而低温胁迫48 h处理下秋茄幼苗叶片Car/Chl值无显著性差异;两个低温胁迫处理相比,秋茄幼苗叶片Chl含量无显著差异变化,而Car/Chl值显著下降(P<0.05)。与对照相比,低温胁迫24 h处理下,秋茄幼苗叶片Car含量和Chl a/Chl b值均无显著差异变化,而低温胁迫48 h处理下秋茄幼苗叶片Car含量和Chl a/Chl b值均显著下降(P<0.05)。
与对照相比,低温胁迫24 h和48 h处理下,红榄李幼苗叶片Chl含量分别降低23%和41%,Car/Chl值分别上升10%和30%;两个低温胁迫处理相比,低温胁迫24 h处理下红榄李幼苗叶片Chl含量显著高于低温胁迫48 h处理(P<0.05),两个低温胁迫处理下红榄李幼苗叶片Car/Chl值均上升。与对照相比,低温胁迫24 h处理下红榄李幼苗叶片Car含量和Chl a/Chl b值均无显著差异变化,而低温胁迫48 h处理下红榄李幼苗叶片Car含量和Chl a/Chl b值均显著下降(P<0.05)。
-
秋茄和红榄李幼苗叶片SOD活性均随低温胁迫延长逐渐降低,且红榄李幼苗下降的幅度较大(表4)。比如,与对照相比,低温胁迫24 h和48 h处理下,秋茄幼苗叶片SOD活性分别降低2%和61%,而红榄李幼苗叶片SOD活性分别降低51%和85%。不同低温处理间相比,低温胁迫24 h处理下,秋茄叶片SOD活性显著高于低温胁迫48 h处理,而与对照之间无显著差异;低温胁迫24 h处理下,红榄李幼苗叶片SOD活性与对照和低温胁迫48 h处理均存在显著差异(P<0.05)。
表 4 低温胁迫对两种红树植物叶片超氧岐化酶(SOD)、过氧化物酶(POD)活性以及丙二醛(MDA)含量的影响
Table 4. Effects of low temperature stress on the activities of superoxide dismutase (SOD), peroxidase (POD) and content of malondialdehyde (MDA) in leaves of two mangrove plants
红树植物
Mangrove plants处理
TreatmentSOD活性
SOD activity/(U·g−1 )POD活性
POD activity/(U·g−1)丙二醛含量
MDA content/(mmol·g−1)秋茄
K. obovata对照 59.99 ± 0.64 a 1.98 ± 0.57 c 1.01 ± 0.01 e 24 h 58.80 ± 1.89 a 4.21 ± 0.63 a 1.36 ± 0.11 d 48 h 23.61 ± 1.57 b 2.82 ± 0.37 b 1.65 ± 0.04 c 红榄李
L. littore对照 59.93 ± 2.84 a 2.53 ± 0.17 b 1.29 ± 0.02 d 24 h 29.21 ± 1.65 b 1.20 ± 0.01 d 1.98 ± 0.04 b 48 h 9.02 ± 1.88 c 0.16 ± 0.15 e 2.42 ± 0.01 a 随低温胁迫时间延长,秋茄幼苗叶片POD活性先增后降,而红榄李幼苗叶片POD活性一直持续下降(表4)。与对照相比,低温胁迫24 h和48 h处理下,秋茄幼苗叶片POD活性增加了113%和42%,而红榄李幼苗叶片POD活性分别降低53%和94%。
随着低温胁迫延长两种红树植物叶片MDA含量均逐渐增加,且红榄李幼苗叶片MDA含量高于秋茄(表4)。低温胁迫24 h、48 h处理下,秋茄幼苗叶片MDA含量分别为对照的1.35倍和1.63倍,而红榄李幼苗叶片MDA含量分别为对照1.53倍和1.88倍。
-
由表5可知,随着低温胁迫时间延长,秋茄和榄李幼苗叶片H2O2和O2·−含量均逐渐增加。与对照相比,低温胁迫24 h和48 h处理下秋茄幼苗叶片H2O2、O2·−含量分别增加了20%、27%和44%、32%;低温胁迫24 h、48 h处理下红榄李幼苗叶片H2O2和O2·−含量分别增加了31%、37%与60%、43%。
表 5 低温胁迫对两种红树植物叶片过氧化氢(H2O2)含量和超氧阴离子(O2·−)的影响
Table 5. Effects of low temperature stress on the contents of hydrogen peroxide (H2O2) and superoxide anion (O2·−) in leaves of two mangrove plants
红树植物
Mangrove plants处理
TreatmentH2O2含量
H2O2 content/
(μmol·g−1)O2·−含量
O2·− content/
(nmol·g−1)秋茄
K. obovata对照 63.74 ± 4.76 c 410.01 ± 0.41 b 24 h 76.47 ± 10.52 b 520.21 ± 0.42 a 48 h 91.68 ± 2.22 a 541.65 ± 0.53 a 红榄李
L. littore对照 3.49 ± 0.33 f 30.11 ± 0.12 d 24 h 4.56 ± 1.10 e 41.12 ± 0.11 c 48 h 5.57 ± 0.95 d 43.12 ± 0.13 c -
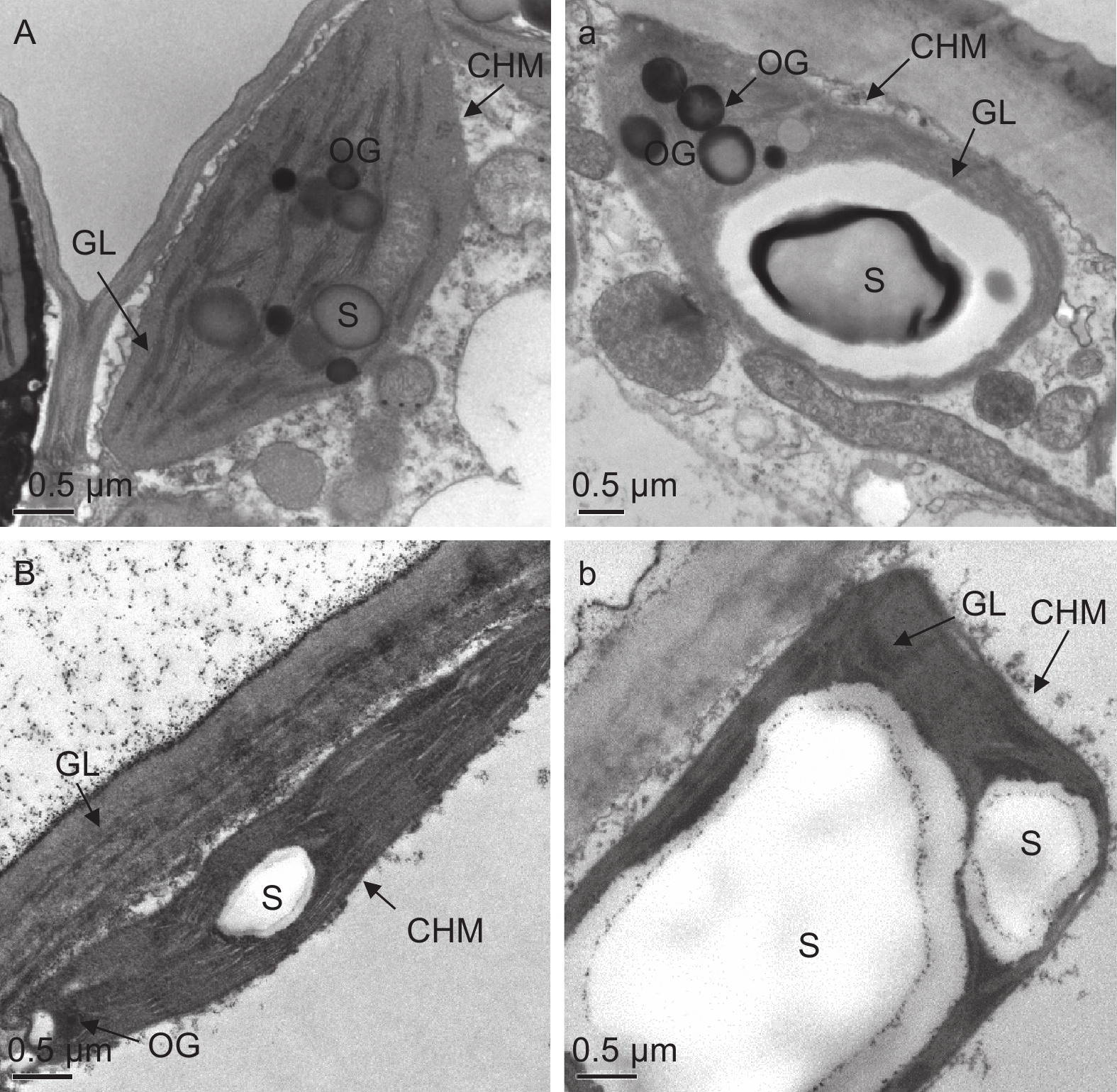
由图3可知,对照条件下,秋茄(图3A)和红榄李(图3B)幼苗叶片叶绿体紧密分布在细胞壁周围且结构完整,呈现为饼状或者凸透镜状,基粒片层排列较为紧密有序,膜清晰可辨,可以观察到叶绿体内有淀粉粒(S)和嗜锇体(OG)。与对照相比,低温胁迫48 h处理下秋茄(图3 a)和红榄李(图3 b)的叶绿体与对照相比,其叶绿体明显膨胀,淀粉粒肿胀变大且基粒片层间距加大,叶绿体膜较对照状态下出现不清晰或破裂等现象,其中红榄李幼苗叶片叶绿体结构受低温胁迫伤害更大,如叶绿体数目、单位基粒数和基粒片层数均减少,淀粉粒异常膨大。
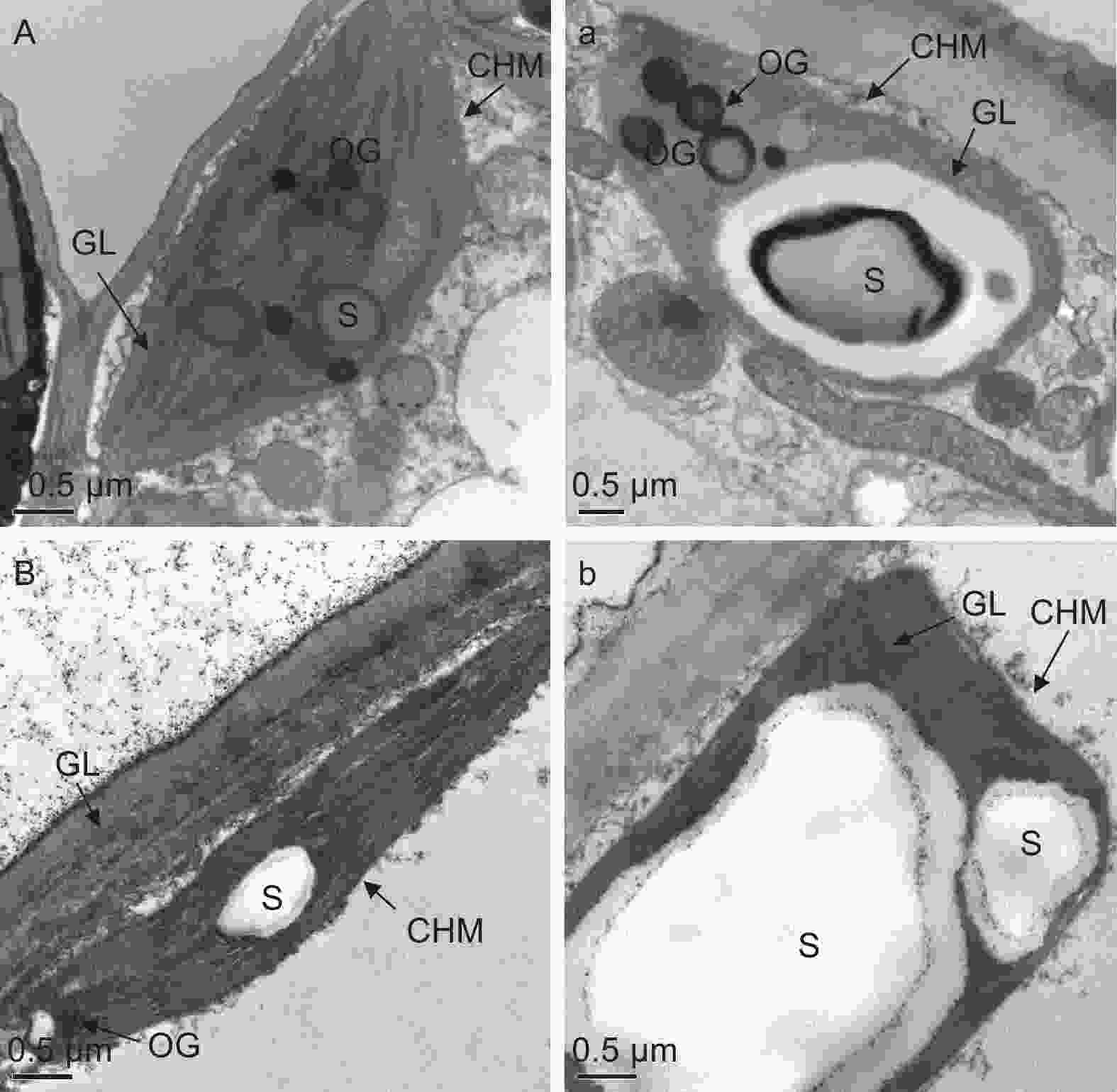
图 3 低温胁迫下两种红树植物叶绿体超微结构
Figure 3. Effects of low temperature stress on the ultrastructure of chloroplasts in two mangrove plants
-
由图4可知,低温胁迫下秋茄和红榄李幼苗叶片气孔开张度均明显小于对照。通过计算,低温胁迫下秋茄幼苗叶片气孔长、宽以及开张度均显著高于红榄李(P<0.05)。与对照相比,低温胁迫48 h处理下秋茄、红榄李幼苗叶片气孔长、宽以及开张度长并未显著变化。同样,秋茄幼苗叶片气孔开张度宽均显著高于红榄李(P<0.05)。与对照相比,低温胁迫48 h处理下秋茄和红榄李幼苗气孔开张度宽分别降低37%和80%,气孔面积分别上升了25%和24%。

图 4 低温胁迫对两种红树植物叶片气孔形态的影响
Figure 4. Effects of low temperature stress on the leaf stomata of two mangrove plants
-
光合作用是植物生长发育的物质基础,而低温胁迫下植物则通过光合作用强弱可以反映其抗寒能力[25-26]。研究认为,逆境胁迫引起的植物光合作用限制因子可分为气孔限制和非气孔限制,其中气孔限制导致的叶片光合速率下降主要表现气孔关闭引起Ci、Gs均降低,Ls升高;而非气孔限制则主要表现为Ci升高,Ls下降,这归因于逆境会破坏叶绿体结构,使光合色素合成受到抑制,电子传递受阻等[27-30]。前期研究还发现,低温胁迫会使红树植物秋茄的叶绿体超微结构发生改变,基粒溶解、基层片层断裂,膜破裂,甚至解体,最终引起叶片净光合速率降低,这也是非气孔限制的主要表现之一[25]。然而,低温如何影响红榄李幼苗叶片光合能力的研究仍未有相关报道。在本研究中,48 h 7 ℃(昼)/4 ℃(夜)处理对秋茄幼苗影响较小,其植株未有低温伤害症状,而在同样温度下红榄李幼苗却出现萎蔫现象,且叶片净光合速率值为负值,表明秋茄耐寒性远高于红榄李。本研究还发现,低温胁迫24 h处理下秋茄幼苗叶片Pn、Gs以及Ls均降低,而Ci升高;低温胁迫48 h处理下,秋茄幼苗叶片Pn、Gs以及Ci均降低,而Ls升高,表明低温胁迫24 h处理下秋茄幼苗叶片Pn的下降主要受到非气孔限制的影响,这可能是秋茄对低温的应急反应。当胁迫时间延迟至48 h时,其Pn的降低由气孔限制影响,主要表现在气孔开张度减少(图4B和表6),与此同时,非气孔限制仍存在,如叶绿体超微结构(图3a)受到轻微伤害。与秋茄不同,低温胁迫24 h和48 h处理下红榄李幼苗叶片Pn、Gs以及Ls均降低,而Ci均升高,表明低温胁迫下,红榄李幼苗叶片Pn的降低主要由非气孔限制起着主要作用,表现在叶绿体超微结构受到严重破坏,如叶绿体数目、单位基粒数和基粒片层数均减少,淀粉粒异常膨大等(图3b),叶片气孔基本关闭(图4D)。综上所述,非气孔限制是影响红榄李幼苗不耐寒的一个重要因素。
表 6 低温胁迫对两种红树植物叶片气孔指标的影响
Table 6. Effects of low temperature stress on leaf stomatal indexes of two mangrove plants
红树植物
Mangrove plants处理
Treatment气孔长
Stomata length/μm气孔宽
Stomata width/μm气孔开张度
Stomatal aperture气孔面积 长
Length/μm宽
Width/μmStomatal Area/
μm2秋茄
K. obovata对照 36.49 ± 0.41 a 23.36 ± 0.21 a 31.81 ± 0.42 a 10.07 ± 0.18 a 367.59 ± 2.32 b 低温 39.29 ± 0.36 a 21.71 ± 0.14 a 28.73 ± 0.23 a 6.37 ± 0.24 b 458.31 ± 2.68 a 红榄李
L. littore对照 24.75 ± 0.07 b 10.29 ± 0.10 b 18.42 ± 0.16 b 2.79 ± 0.31 c 154.54 ± 2.31 d 低温 24.21 ± 0.21 b 10.27 ± 0.22 b 18.16 ± 0.37 b 0.55 ± 0.11 d 192.02 ± 8.46 c 植物叶片的叶绿素荧光参数(Fv/Fm、ΦPSII、qP等)可作为判定植物耐寒能力的重要指标,其中电子传递有效性和光化学效率的差异是影响植物耐寒性的主要原因[31]。研究认为,低温胁迫直接降低秋茄叶片Fv/Fm、ΦPSII和qP等,引起PSII反应中心出现过剩的激发能,致使叶片发生PSII光抑制,最终导致叶片Pn下降[25]。本研究中,随着低温胁迫时间的延长,秋茄、红榄李幼苗叶片Fv/Fm、ΦPSII和qP均下降,且低温胁迫48 h处理下红榄李幼苗叶片各荧光参数下降幅度均高于秋茄,表明低温胁迫会使两种红树植物的PSII反应中心发生部分失活或伤害,降低反应中心对激发能捕获能力,影响叶片净光合速率。然而,低温胁迫对红榄李的伤害更为严重,这也验证了红榄李叶片Pn下降是由于非气孔限制引起。前期研究还发现,低温胁迫提高秋茄幼苗叶片NPQ值,表明植株通过以热耗散方式散失过多的光能,从而减轻光系统伤害[32],这与本研究结果一致。然而,红榄李幼苗仅在低温胁迫24 h处理下,叶片NPQ显著高于对照,而低温胁迫48 h处理下,其值却显著下降,这表明低温胁迫时间延长后红榄李已不能通过增加热耗散来避免伤害,从而丧失了防御能力。
光合色素能够在植物光合作用中参与吸收、传递光能或引起原初光化学反应,其含量的变化可反映植物光合作用的强弱。光合色素主要包括Chl(Chl a和Chl b)和Car,其中Chl在光合作用的光吸收中其核心作用,而Car不仅具有天线色素作用,而且还可以减少ROS[33]。前期研究发现,低温胁迫下Chl a/Chl b值能反映光合能力强弱,且Chl a/Chl b值与秋茄抗寒性有正相关[34]。研究还认为,叶绿素含量的降低主要是低温胁迫通过增加叶绿素酶活性,促进叶绿素的降解[35]。本研究中,随着低温胁迫时间的延长,秋茄和红榄李幼苗叶片叶绿素含量、Chl a/Chl b值均呈下降的趋势,且低温胁迫24 h 和48 h 处理下,秋茄幼苗叶片叶绿素含量和Chl a/Chl b值均无显著性差异,而红榄李幼苗叶片叶绿素含量却存在显著差异,这表明低温胁迫时间延长可能也会加快红榄李幼苗叶片叶绿素酶活性,促进叶绿素的降解,而相同低温胁迫下秋茄幼苗却未有相似现象,这可能归因于较高的Chl a/Chl b值,即秋茄的抗寒能力高于红榄李。对两种红树叶片Car含量变化分析,秋茄幼苗叶片Car合成对低温胁迫时间较红榄李敏感,即在低温胁迫下秋茄幼苗叶片Car参与清除ROS的作用相比于红榄李较强,如低温胁迫24 h和48 h处理下秋茄幼苗叶片Car含量存在显著差异,而红榄李幼苗而无显著差异。Car/Chl值与植物清除活性氧的能力有关,逆境下植物Car/Chl值变化可与清除ROS的抗氧化酶活性一致[36]。低温胁迫24h处理下,秋茄幼苗叶片Car/Chl值显著增加,而随着低温胁迫时间延长,其值却下降。这主要是因为秋茄幼苗受到轻微低温胁迫时,植株出现应急反应,迅速提升Car/Chl值,进而减轻ROS对细胞的伤害;之后随着低温胁迫时间延长,这种自我保护作用减弱。与秋茄不同,随着低温时间延长红榄李幼苗叶片Car/Chl值一直持续增加,这可能与低温胁迫下红榄李幼苗叶片ROS大量积累有关。
低温胁迫会使不耐寒红树植物体内大量ROS累积,引起细胞膜脂过氧化,破坏膜结构,而耐寒红树植物却在低温胁迫下植株呈现较轻的伤害程度,这与清除ROS的抗氧化酶活性和Car含量有关[12]。研究认为,当植物抵抗低温胁迫时,其首先启动SOD,将O2·−转化为H2O2和O2,POD和CAT则进一步将H2O2分解为H2O和O2,从而减轻活性氧对膜系统的伤害,增强植株的抗寒性[37]。本研究中,一方面,随着低温胁迫时间延长秋茄、红榄李幼苗叶片SOD活性均降低,致使清除O2·−能力降低,使其含量增加。另一方面,在两种红树植物体内SOD催化O2·−歧化为H2O2和O2后,POD对清除H2O2将起着重要作用;相同低温胁迫下秋茄幼苗叶片POD活性显著增加,而红榄李幼苗叶片POD活性显著降低。此外,秋茄幼苗叶片MDA含量增加幅度小于红榄李。说明POD在低温胁迫下两种红树植物抗氧化清除酶系统中起着主要作用[16],其中低温胁迫会抑制红榄李幼苗叶片抗氧化酶活性,提高了H2O2含量,增加了膜脂过氧化产物MDA积累,从而加剧膜的损伤;相反,秋茄却有较强的防御机制,这也是其耐寒性高于红榄李的原因之一。
-
随着低温胁迫时间延长秋茄和红榄李幼苗叶光合能力均逐渐降低,但红榄李光合作用降低的幅度更大,尤其是在低温胁迫48 h处理下,表明秋茄耐寒性较红榄李高。这可能是因为:低温胁迫48 h处理下,红榄李幼苗叶片光合色素下降,叶绿体超微结构受到破坏,气孔开张度减少,光合电子传递效率降低,PSII反应中心受到伤害,ROS增加,膜系统受损,致使光合能力下降,而秋茄受到的低温伤害较红榄李轻。研究结果不仅丰富了我国红树林抗寒研究内容,而且也为高纬度红树林引种提供了重要参考依据。
低温胁迫对不同耐寒红树植物幼苗光合生理和抗氧化特性的影响
Effects of Low Temperature Stress on Leaf Photosynthetic Physiology and Antioxidant Characteristics in Mangrove Plants Seedlings with Different Cold Tolerance
-
摘要:
目的 探索两种不同耐寒红树植物对低温胁迫的生理响应机制,为红树林抗寒性研究提供重要理论依据。 方法 以秋茄、红榄李幼苗为试验材料,设置25 ℃ (昼) /20 ℃ (夜)、24 h 7 ℃(昼)/4 ℃(夜)、48 h 7 ℃(昼)/4 ℃(夜)3个处理,分别比较分析了两种红树植物幼苗叶片的光合参数、荧光特征、抗氧化能力、叶绿体超微结构等生理过程对低温响应的差异。 结果 低温胁迫抑制红榄李幼苗生长和光合作用,而对秋茄的影响却较小。在低温胁迫处理48 h后,红榄李幼苗叶片超氧歧化酶(SOD)、过氧化物酶(POD)活性显著降低,而超氧阴离子(O2 ·−)、过氧化氢(H2O2)含量、丙二醛(MDA)含量增加,同时气孔关闭,光合色素合成受阻,叶绿体超微结构受损,最终导致叶片净光合速率(Pn)、气孔导度(Gs)、最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSⅡ)、光化学猝灭系数(qP)以及非光化学猝灭系数(NPQ)等降低。相同低温胁迫下秋茄幼苗叶片Pn也受到抑制,但其Pn下降幅度约为红榄李的79%。 结论 秋茄耐寒能力较强于红榄李,这可能是因为秋茄在低温胁迫下不仅能够保持较高的气孔开张度、光合色素含量以及POD活性,维持较好的叶绿体超微结构,而且还会通过调控能量耗散减轻PSII光抑制,降低低温胁迫带来的膜脂过氧化伤害,最终维持自身较强的光合能力。 Abstract:Objective This study investigates the physiological response of two different cold-tolerant mangrove plant species to low temperature stress, which provides a theoretical basis for studying on cold resistance of mangrove. Method Based on the cold-tolerant variety Kandelia obovata and the non-cold-tolerant variety Lumnitzera littore , three temperature treatments were set for treatment, including 25 ℃/20 ℃ (day/night), 24 hours at 7 ℃/4 ℃ (day/night), and 48 hours at 7 ℃/4 ℃ (day/night). The differences in the of photosynthetic parameters and fluorescence characteristic, antioxidant capacity, chloroplast ultrastructure of two mangrove seedlings were compared and analyzed. Result Low-temperature stress inhibited the growth and photosynthesis of L. littore seedlings but had less effect on K. obovate. After 48 hours of low-temperature stress treatment, the activities of superoxide dismutase (SOD) and peroxidase (POD) in the leaves of L. littore seedlings were reduced, but the content of superoxide anion (O2·−), malondialdehyde (MDA) and hydrogen peroxide (H2O2) in L. littore seedlings were increased. It was also observed that the leaf stomata were closed, and the contents of photosynthetic pigments were decreased. Additionally, the ultrastructure of the chloroplast was damaged, which eventually led to a decrease in the net photosynthetic rate (Pn), stomatal conductance (Gs), maximum photochemical efficiency (Fv/Fm), actual photochemical efficiency (ΦPSII), photochemical quenching coefficient (qP), and non-photochemical quenching coefficient (NPQ) of the leaves. Under the same low-temperature stress, the photosynthesis of K. obovata seedlings’ leaves was also inhibited, but the decrease in leaf Pn was approximately 79% of that observed in L. littore. Conclusion K. obovata exhibits stronger cold tolerance than L. littore, possibly because K. obovata can not only maintain higher stomatal opening, photosynthetic pigment content, and POD activity, reducing the damage to chloroplast ultrastructure, but also regulate energy dissipation to alleviate PSII photoinhibition, thereby reducing the damage caused by low-temperature stress to membrane lipid peroxidation. Consequently, K. obovata maintains its robust photosynthetic capacity. -
Key words:
- mangrove
- / low temperature
- / photosynthetic fluorescence
- / antioxidant system
- / chloroplast structure
-
图 1 2015年不同树龄红树林极端低温伤害症状
Figure 1. The symptoms of extremely cold events damage on mangroves in 2015
图 2 低温胁迫对两种红树植物幼苗生长情况的影响
Figure 2. Effects of low temperature stress on growth of two mangrove seedlings
图 3 低温胁迫下两种红树植物叶绿体超微结构
Figure 3. Effects of low temperature stress on the ultrastructure of chloroplasts in two mangrove plants
图 4 低温胁迫对两种红树植物叶片气孔形态的影响
Figure 4. Effects of low temperature stress on the leaf stomata of two mangrove plants
表 1 低温胁迫对两种红树植物幼苗叶片气体交换参数的影响
Table 1. Effects of low temperature stress on gas exchange parameters in leaves of two mangrove seedlings
红树植物
Mangrove plants处理
Treatment净光合速率
Pn/(μmol·m−2·s−1)气孔导度
Gs/(mmol·m−2·s−1)胞间CO2浓度
Ci/(μmol·mol−1)气孔限制值
Ls秋茄
K. obovata对照 6.38 ± 0.54 a 147.87 ± 0.93 a 317.03 ± 12.25 b 0.25 ± 0.03 c 24 h 3.41 ± 1.09 b 111.43 ± 0.30 b 376.99 ± 11.31 a 0.14 ± 0.05 d 48 h 1.14 ± 0.13 c 43.69 ± 0.72 d 249.14 ± 15.83 c 0.55 ± 0.10 a 红榄李
L. littore对照 6.30 ± 0.27 a 151.87 ± 6.84 a 252.80 ± 6.95 c 0.38 ± 0.01 b 24 h 0.19 ± 0.01 d 68.54 ± 8.79 c 322.45 ± 7.06 b 0.21 ± 0.02 c 48 h -0.33 ± 0.06 d 14.84 ± 3.20 d 391.10 ± 5.91 a 0.05 ± 0.01 e 注:同列不同小写字母表示不同处理间存在差异显著(P<0.05)。下同
Notes: Different lowercase letters of the same column indicate significant difference at 0.05 level. The same below 下载: 导出CSV
下载: 导出CSV
表 2 低温胁迫对两种红树植物幼苗叶片叶绿素荧光参数的影响
Table 2. Effects of low temperature stress on chlorophyll fluorescence parameters of leaves of two mangrove seedlings
红树植物
Mangrove plants处理
TreatmentFv/Fm ΦPSⅡ qP NPQ 秋茄
K. obovata对照 0.85 ± 0.01 a 0.58 ± 0.04 a 0.69 ± 0.06 a 0.53 ± 0.29 cd 24 h 0.70 ± 0.02 b 0.46 ± 0.07 b 0.52 ± 0.09 b 0.67 ± 0.28 b 48 h 0.52 ± 0.02 c 0.36 ± 0.04 c 0.39 ± 0.06 c 0.79 ± 0.20 a 红榄李
L. littore对照 0.69 ± 0.01 b 0.56 ± 0.04 a 0.49 ± 0.06 b 0.47 ± 0.05 d 24 h 0.47 ± 0.04 c 0.33 ± 0.01 c 0.36 ± 0.04 c 0.60 ± 0.04 bc 48 h 0.21 ± 0.04 d 0.13 ± 0.03 d 0.24 ± 0.03 d 0.17 ± 0.04 e
下载: 导出CSV
表 3 低温胁迫对两种红树植物幼苗叶片光合色素的影响
Table 3. Effects of low temperature stress on photosynthetic pigments in leaves of two mangrove seedlings
红树植物
Mangrove plants处理
Treatment叶绿素含量
Chl content/(mg·g−1)类胡萝卜素含量
Car content/(mg·g−1)叶绿素a/叶绿素b比
Chl a/Chl b类胡萝卜素/叶绿素比
Car/Chl秋茄
K. obovata对照 1.14 ± 0.15 a 0.20 ± 0.01 a 3.05 ± 0.04 a 0.17 ± 0.01 b 24 h 0.96 ± 0.19 b 0.19 ± 0.01 a 2.85 ± 0.03 ab 0.20 ± 0.02 a 48 h 0.93 ± 0.05 b 0.16 ± 0.01 b 2.56 ± 0.08 b 0.17 ± 0.01 b 红榄李
L. littore对照 0.90 ± 0.12 b 0.09 ± 0.03 c 2.11 ± 0.14 c 0.10 ± 0.03 d 24 h 0.69 ± 0.04 c 0.08 ± 0.02 cd 1.81 ± 0.12 cd 0.11 ± 0.01 d 48 h 0.53 ± 0.04 d 0.07 ± 0.02 d 1.71 ± 0.04 d 0.13 ± 0.01 c
下载: 导出CSV
表 4 低温胁迫对两种红树植物叶片超氧岐化酶(SOD)、过氧化物酶(POD)活性以及丙二醛(MDA)含量的影响
Table 4. Effects of low temperature stress on the activities of superoxide dismutase (SOD), peroxidase (POD) and content of malondialdehyde (MDA) in leaves of two mangrove plants
红树植物
Mangrove plants处理
TreatmentSOD活性
SOD activity/(U·g−1 )POD活性
POD activity/(U·g−1)丙二醛含量
MDA content/(mmol·g−1)秋茄
K. obovata对照 59.99 ± 0.64 a 1.98 ± 0.57 c 1.01 ± 0.01 e 24 h 58.80 ± 1.89 a 4.21 ± 0.63 a 1.36 ± 0.11 d 48 h 23.61 ± 1.57 b 2.82 ± 0.37 b 1.65 ± 0.04 c 红榄李
L. littore对照 59.93 ± 2.84 a 2.53 ± 0.17 b 1.29 ± 0.02 d 24 h 29.21 ± 1.65 b 1.20 ± 0.01 d 1.98 ± 0.04 b 48 h 9.02 ± 1.88 c 0.16 ± 0.15 e 2.42 ± 0.01 a
下载: 导出CSV
表 5 低温胁迫对两种红树植物叶片过氧化氢(H2O2)含量和超氧阴离子(O2·−)的影响
Table 5. Effects of low temperature stress on the contents of hydrogen peroxide (H2O2) and superoxide anion (O2·−) in leaves of two mangrove plants
红树植物
Mangrove plants处理
TreatmentH2O2含量
H2O2 content/
(μmol·g−1)O2·−含量
O2·− content/
(nmol·g−1)秋茄
K. obovata对照 63.74 ± 4.76 c 410.01 ± 0.41 b 24 h 76.47 ± 10.52 b 520.21 ± 0.42 a 48 h 91.68 ± 2.22 a 541.65 ± 0.53 a 红榄李
L. littore对照 3.49 ± 0.33 f 30.11 ± 0.12 d 24 h 4.56 ± 1.10 e 41.12 ± 0.11 c 48 h 5.57 ± 0.95 d 43.12 ± 0.13 c
下载: 导出CSV
表 6 低温胁迫对两种红树植物叶片气孔指标的影响
Table 6. Effects of low temperature stress on leaf stomatal indexes of two mangrove plants
红树植物
Mangrove plants处理
Treatment气孔长
Stomata length/μm气孔宽
Stomata width/μm气孔开张度
Stomatal aperture气孔面积 长
Length/μm宽
Width/μmStomatal Area/
μm2秋茄
K. obovata对照 36.49 ± 0.41 a 23.36 ± 0.21 a 31.81 ± 0.42 a 10.07 ± 0.18 a 367.59 ± 2.32 b 低温 39.29 ± 0.36 a 21.71 ± 0.14 a 28.73 ± 0.23 a 6.37 ± 0.24 b 458.31 ± 2.68 a 红榄李
L. littore对照 24.75 ± 0.07 b 10.29 ± 0.10 b 18.42 ± 0.16 b 2.79 ± 0.31 c 154.54 ± 2.31 d 低温 24.21 ± 0.21 b 10.27 ± 0.22 b 18.16 ± 0.37 b 0.55 ± 0.11 d 192.02 ± 8.46 c
下载: 导出CSV
-
[1] NAIDOO. The mangroves of africa: A review[J]. Marine Pollution Bulletin, 2023, 190: 114859. doi: 10.1016/j.marpolbul.2023.114859 [2] YU C, FENG J, YUE W, et al. The role of blue carbon stocks becomes more labile with mangrove development[J]. Ecological Indicators, 2023, 154: 110634. doi: 10.1016/j.ecolind.2023.110634 [3] QUISTHOUDT K, SCHMITZ N, RANDIN CF, et al. Temperature variation among mangrove latitudinal range limits worldwide[J]. Trees, 2012, 26: 1919-1931. doi: 10.1007/s00468-012-0760-1 [4] 王文卿, 王 瑁. 中国红树林[M]. 北京: 科学出版社, 2007. [5] 王友绍. 全球气候变化对红树林生态系统的影响、挑战与机遇[J]. 热带海洋学报, 2021, 40(3):1-14. [6] LIU K, LIU L, LIU H X, et al. Exploring the effects of biophysical parameters on the spatial pattern of rare cold damage to mangrove forests[J]. Remote Sensing of Environment, 2014, 150: 20-33. doi: 10.1016/j.rse.2014.04.019 [7] ELLIS W L, BOWLES J W, ERICKSON A, et al. Alteration of the chemical composition of mangrove (Laguncularia racemosa) leaf litter fall by freeze damage[J]. Estuarine, Coastal and Shelf Science, 2006, 68(1-2): 363-371. doi: 10.1016/j.ecss.2006.02.017 [8] WANG W Q, YOU S Y, WANG Y B, et al. Influence of frost on nutrient resorption during leaf senescence in a mangrove at its latitudinal limit of distribution[J]. Plant and Soil, 2011, 342(1-2): 105-115. doi: 10.1007/s11104-010-0672-z [9] 吴中华, 赵昊阳, 曾思洁, 等. 不同种源地木榄光合作用对低温寒害的响应[J]. 林业科学研究, 2023, 36(2):195-202. [10] FEI J, WANG Y S, CHENG H, et al. Comparative physiological and proteomic analyses of mangrove plant Kandelia obovate under cold stress[J]. Ecotoxicology, 2021, 30(9): 1826-1840. doi: 10.1007/s10646-021-02483-6 [11] ZHENG C F, LIU W C, QIU J B, et al. Comparison of physiological characteristics of Kandelia obovata at different ages in winter in the northernmost mangrove transplanted area of China[J]. Acta Ecologica Sinica, 2013, 33(3): 132-138. doi: 10.1016/j.chnaes.2013.03.002 [12] 张 颖, 钟才荣, 李诗川, 等. 濒危红树植物红榄李[J]. 林业资源管理, 2013, 5:103-107 + 151. [13] 张 颖, 钟才荣, 杨 勇, 等. 濒危红树植物红榄李种质资源挽救[J]. 分子植物育种, 2018, 16(12):4112-4118. [14] 张 颖, 李燕华, 张晓楠, 等. 濒危红树植物红榄李开花生物学特征及繁育系统[J]. 应用与环境生物学报, 2017, 23(1):77-81. [15] 张 颖, 陈光程, 钟才荣. 中国濒危红树植物研究与恢复现状[J]. 应用海洋学学报, 2021, 40(1):142-153. [16] PENG Y L, WANG Y S, FEI J, et al. Ecophysiological differences between three mangrove seedlings (Kandelia obovata, Aegiceras corniculatum, and Avicennia marina) exposed to chilling stress[J]. Ecotoxicology, 2015, 24(7-8): 1722-1732. doi: 10.1007/s10646-015-1488-7 [17] PENG Y L, WANG Y S, FEI J, et al. Isolation and expression analysis of two novel C-repeat binding factor (CBF) genes involved in plant growth and abiotic stress response in mangrove Kandelia obovate[J]. Ecotoxicology, 2020, 29(6): 718-725. doi: 10.1007/s10646-020-02219-y [18] YUAN L Y, SHU S, GUO S, et al. Effects of 24-epibrassinolide on the photosynthetic characteristics, antioxidant system, and chloroplast ultrastructure in Cucumis sativus L. under Ca(NO3)2 stress[J]. Photosynthesis Research, 2012, 112(3): 205-214. doi: 10.1007/s11120-012-9774-1 [19] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000. [20] TAN W, LIU J, DAI T, et al. Alterations in photosynthesis and antioxidant enzyme activity in winter wheat subjected to post-anthesis water-logging[J]. Photosynthetica, 2008, 46(1): 21-27. doi: 10.1007/s11099-008-0005-0 [21] Du Z Y, Bramlage W J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts[J]. Journal of Agricultural and Food Chemistry, 1992, 40(9): 1566-1570. doi: 10.1021/jf00021a018 [22] 彭 建, 王丹英, 徐春梅, 等. 钼酸铵法测定水稻过氧化氢酶活性[J]. 中国农学通报, 2009, 25(16):61-64. [23] 李忠光, 龚 明. 植物中超氧阴离子自由基测定方法的改进[J]. 云南植物研究, 2005, 27(2):211-216. [24] 张 翠, 马 瑞, 谭立佳, 等. 兰州市10种常用园林绿化树种叶表面微结构对其滞尘量的影响[J]. 甘肃农业大学学报, 2023, 58(4):192-200 + 211. [25] LIU W C, ZHENG C F, CHEN J N, et al. Cold acclimation improves photosynthesis by regulating the ascorbate-glutathione cycle in chloroplasts of Kandelia Obovate[J]. Journal of Forest Research, 2019, 30(3): 755-765. doi: 10.1007/s11676-018-0791-6 [26] ZHAO S X, QU X L, ZHANG Y D, et al. Over-activation of cold tolerance in arabidopsis causes carbohydrate shortage compared with Chorispora bungeana[J]. Journal of Plant Physiology, 2023, 289: 154083. doi: 10.1016/j.jplph.2023.154083 [27] FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 1982, 33(1): 317-345. doi: 10.1146/annurev.pp.33.060182.001533 [28] 褚 润, 陈年来. UV-B 辐射增强对芦苇光合生理及叶绿体超微结构的影响[J]. 应用生态学报, 2017, 28(11):3515-3520. [29] JU S M, WANG L P, CHEN J Y. Effects of silicon on the growth, photosynthesis and chloroplast ultrastructure of Oryza sativa L. seedlings under acid rain stress[J]. Silicon, 2020, 12(3): 655-664. doi: 10.1007/s12633-019-00176-8 [30] 李庆会, 徐 辉, 周 琳, 等. 低温胁迫对2个茶树品种叶片叶绿素荧光特性的影响[J]. 植物资源与环境学报, 2015, 24(2):26-31. [31] 胡春梅, 候喜林, 王 旻. 低温胁迫对不结球白菜光合及叶绿素荧光特性的影响[J]. 西北植物学报, 2008, 28(12):2478-2484. [32] 郑春芳, 刘伟成, 魏 龙, 等. 外施褪黑素对低温胁迫下红树植物秋茄光合作用和抗坏血酸-谷胱甘肽循环的调控[J]. 植物生理学报, 2019, 55(8):1211-1221. [33] 铁得祥, 胡红玲, 喻秀艳, 等. 桢楠幼树光合特性对镉胁迫的响应[J]. 生态学报, 2020, 40(11):3738-3746. [34] ZHENG C F, YE Y, LIU W C, et al. Recovery of photosynthesis, sucrose metabolism, and proteolytic enzymes in Kandelia obovata from rare cold events in the northernmost mangrove, China[J]. Ecological Processes, 2016, 5: 9. doi: 10.1186/s13717-016-0047-3 [35] KUMAR N, GUPTA S, TRIPATHI A N. Gender-specific responses of Piper betle L. to low temperature stress: changes in chlorophyllase activity[J]. Biologia Plantarum, 2006, 50: 705-708. doi: 10.1007/s10535-006-0111-4 [36] 陈健妙, 郑青松, 刘兆普, 等. 麻疯树((Jatropha curcas L. )幼苗生长和光合作用对盐胁迫的响应[J]. 生态学报, 2009, 29(3):1356-1365. [37] PING D, ZHAI M L, WANG A G, et al. Exogenous methyl jasmonate enhanced the antioxidant capacity of Malus baccata by stimulating jasmonate signalling under suboptimal low root-zone temperature[J]. Scientia Horticulturae, 2023, 321: 112292. doi: 10.1016/j.scienta.2023.112292 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 767
- HTML全文浏览量: 441
- PDF下载量: 22
- 被引次数: 0
