

-
林下植被一般由林下灌木层、草本层和地被苔藓层等组成,是森林的重要组成部分,在森林生态系统演替[1]、土壤养分循环[2]、水土保持[3]、生物多样性维持[4]等方面具有独特的功能和作用。森林经营管理大都在林下作业,会对森林下层植被产生直接或间接影响[5]。研究表明,间伐、除草、割灌等抚育等措施通过增加可利用空间和资源,能促进林下植被更新与生长,改变林下植被物种组成,进而影响森林的生态功能[6-7]。森林间伐也可能会使部分林下植物优势地位增加,从而减少林下植物多样性[8]。此外,林下植被对森林管理的响应会因森林类型、林分演替阶段和扰动发生时间的不同存在一定的差异,具体机制有待进一步探讨。
千岛湖是浙江省最为重要的饮用水源保护地,也是华东地区重要的战略储备水源地[9],千岛湖地区森林生态系统的提质增效是保障区域水资源安全的重要途径。该地区森林类型以马尾松次生林为主,但由于长期较高密度的纯林经营,造成了林地退化、生产力下降、水源涵养能力低、生物多样性较差等生态问题,并伴有不同程度的松材线虫病侵害,严重威胁着林地的可持续经营[10]。为此,区域大力开展马尾松林的阔叶化改造,促进马尾松低效纯林向高质、稳定的针阔混交和阔叶混交林进行演替。
当前,关于马尾松林下植被相关研究多集中在间伐强度、林分密度等对林下植物多样性和群落结构的影响[11-12],尚缺乏关于马尾松林地不同改造措施下的林下植被稳定性与多样性关系的研究。而林下植物多样性和群落稳定性对于理解整个森林生态系统的稳定性及功能多样性至关重要,也是林地可持续经营的重要依据。因此,本研究以淳安县千岛湖林区马尾松林阔叶化改造后形成的不同林分类型为对象,分析其林下植被物种组成、多样性及其与林地特征因子的关系,比较不同林分类型的林下植物群落稳定性差异及其与物种多样性之间的关系,以期为千岛湖区马尾松林的可持续经营提供数据支撑和理论依据。
-
研究区位于浙江省杭州市淳安县浪川乡(29°28′~29°30′ N, 118°36′~118°38′ E),属中亚热带季风气候,温暖湿润,雨热同期,四季分明,年均降水量1 515 mm,年均气温18℃,常年盛行东北风,年均风速2.1 m·s−1。区域主要为山地丘陵,土壤以红壤、黄壤、岩性土为主。植被以马尾松次生林为主,优势乔木种为马尾松(Pinus massoniana Lamb.)、杉木(Cunninghamia lanceolata (Lamb.) Hook.)、青冈(Quercus glauca Thunb.)、苦槠(Castanopsis sclerophylla (Lindl.) Schottky)、枫香(Liquidambar formosana Hance)等,优势灌木种为檵木(Loropetalum chinense (R. Br.) Oliv.)、油茶(Camellia oleifera Abel)、江南越橘(Vaccinium mandarinorum Diels)等,草本有芒萁(Dicranopteris pedate (Houtt.) Nakaike)、蕨(Pteridium aquilinum var. latiusculum (Desv.)Underw.e)等,藤本有菝葜(Smilax china L.)、刺花悬钩子(Rubus aculeatiflorus Hayata)等。
-
研究区马尾松林阔叶化改造项目始于2018年,改造前均为马尾松次生纯林,改造后逐渐向针阔混交和阔叶混交林发展,改造措施包括疏伐促新、疏伐补阔、皆伐促新和皆伐补阔4种类型。其中:(I)疏伐促新:伐除约70%的马尾松,无割灌、除草和补植措施;(II)疏伐补阔:伐除约40%的马尾松,无割灌、除草,补植枫香、油茶;(III)皆伐促新:伐除所有马尾松,对林下幼阔和灌木进行抚育,保留建群种;(IV)皆伐补阔:伐除所有马尾松,对林下幼阔和灌木进行抚育,保留建群种,补植枫香、木荷(Schima superba Gardner & Champ.)。分别在4类改造后的林分中设置标准样地3个,并在周边无采伐抚育措施的马尾松次生纯林(V)设置对照样地3个,各样地立地条件类似,共设置15个样地,样地大小为20 m × 20 m,各样地间隔大于30 m。
-
植被调查时间在2022年7月和12月,对每个标准样地内的乔木进行每木检尺,记录胸径、树高、冠幅,并估测郁闭度。各类型样地林分特征如表1所示。将标准样地划分为16个5 m × 5 m小样地,随机选择3个小样地进行灌木层调查(将胸径小于5 cm、高度低于3 m的乔木幼苗或幼树也归为灌木层,同时将木质藤本植物也归入灌木层),测定植物的地径、树高、冠幅。并在每个小样地内随机设置5个1 m × 1 m的草本层样方,测定草本植物种类、多度、盖度等指标。
表 1 研究区不同类型林分特征
Table 1. Stand characteristics of different types in the study area
林型(编号)
Type (Number)郁闭度
Canopy density主要树种
Main species树种占比/%
Proportion重要值
Important value胸径
DBH/cm树高
Height/m冠幅
Crown breadth/m疏伐促新(I)
Thinning and
promoting
regeneration0.70 杉木
Cunninghamia lanceolata32.4 0.225 11.83 ± 4.53 b 8.71 ± 1.92 b 3.17 ± 0.96 b 马尾松
Pinus massoniana4.5 0.151 36.40 ± 2.17 a 21.17 ± 2.04 a 6.27 ± 0.98 a 老鼠矢
Symplocos stellaris Brand15.5 0.068 4.00 ± 1.76 c 4.28 ± 1.18 c 1.93 ± 0.69 c 疏伐补阔(II)
Thinning and
supplementing
broadleaf0.65 马尾松
Pinus massoniana31.3 0.445 12.11 ± 5.30 a 8.83 ± 2.71 a 3.08 ± 1.00 a 油茶
Camellia oleifera35.6 0.247 4.64 ± 2.16 b 3.39 ± 0.29 b 1.90 ± 0.60 b 槲栎
Quercus aliena9.8 0.083 4.13 ± 0.90 b 3.37 ± 0.27 b 1.74 ± 0.33 b 皆伐促新(III)
Clear cutting and
promoting
regeneration0.55 苦槠
Castanopsis sclerophylla27.4 0.293 11.25 ± 4.25 b 7.20 ± 2.19 b 2.91 ± 1.25 b 杉木
Cunninghamia lanceolata11.6 0.188 16.78 ± 2.72 a 9.95 ± 0.96 a 3.56 ± 0.84 a 枫香
Liquidambar formosana25.3 0.166 2.68 ± 0.28 c 3.17 ± 0.27 c 1.20 ± 0.30 c 皆伐补阔(IV)
Clear cutting and
supplementing
broadleaf0.30 枫香
Liquidambar formosana25.0 0.451 9.03 ± 1.88 a 3.00 ± 0.25 b 1.40 ± 0.35 b 野柿
Diospyros kaki var.
sylvestris
Makino50.0 0.285 1.08 ± 0.04 c 4.70 ± 0.28 a 2.68 ± 0.33 a 紫叶李
Prunus cerasifera
‘Atropurpurea’25.0 0.264 4.72 ± 1.03 b 3.04 ± 0.30 b 1.64 ± 0.45 b 对照(V)
Control group0.75 马尾松
Pinus massoniana40.5 0.555 21.30 ± 4.59 a 21.90 ± 4.29 a 4.13 ± 1.40 a 檵木
Loropetalum chinense30.0 0.167 4.55 ± 1.11 b 4.97 ± 1.33 b 1.74 ± 0.72 b 油茶
Camellia oleifera7.9 0.080 5.30 ± 1.99 b 3.56 ± 1.31 b 2.24 ± 148 b -
在各标准样地的上、中、下坡位分别采集土壤样品各1份,去除地表枯落物,在0~10 cm和10~20 cm中间部位,分别用环刀获取原状土壤,利用烘干法测定土壤密度(SBD)和含水量(SM)。另外,挖取少许0~10 cm 和10~20 cm土壤样品带回实验室。样品经风干、粉碎、过筛(2 mm)后,测定土壤pH、全氮(TN)、全磷(TP)、全钾(TK)、水解性氮(HN)、速效磷(AP)、速效钾(AK)、有机质(SOM),上述指标测定方法参考《土壤调查实验室分析方法》[13]。
-
重要值:
$ IV=\frac{Dr + Pr + Fr}{3} $
(1) 上式中,Dr为相对多度,Pr为相对显著度,Fr为相对频度。
丰富度指数(Margalef index, H):
$ H=\frac{S-1}{InN} $
(2) 香农-威纳指数(Shannon-Wiener index, H'):
$ H{'}={\sum }_{i=1}^{s}{P}_{i}\mathit{In}{P}_{i} $
(3) 均匀度指数(Pielou index, J):
$ J=\frac{H'}{InS} $
(4) 生态优势度指数(Simpson index, D):
$ D=1-{\sum} _{i=1}^{s}{{p}_{i}}^{2}$
(5) 上式中,S为样地内植物种类总数,N为样地内植物个体总数,Pi为第i个种的个体数(Ni)占样地内所有种个体总数(N)的比值。
-
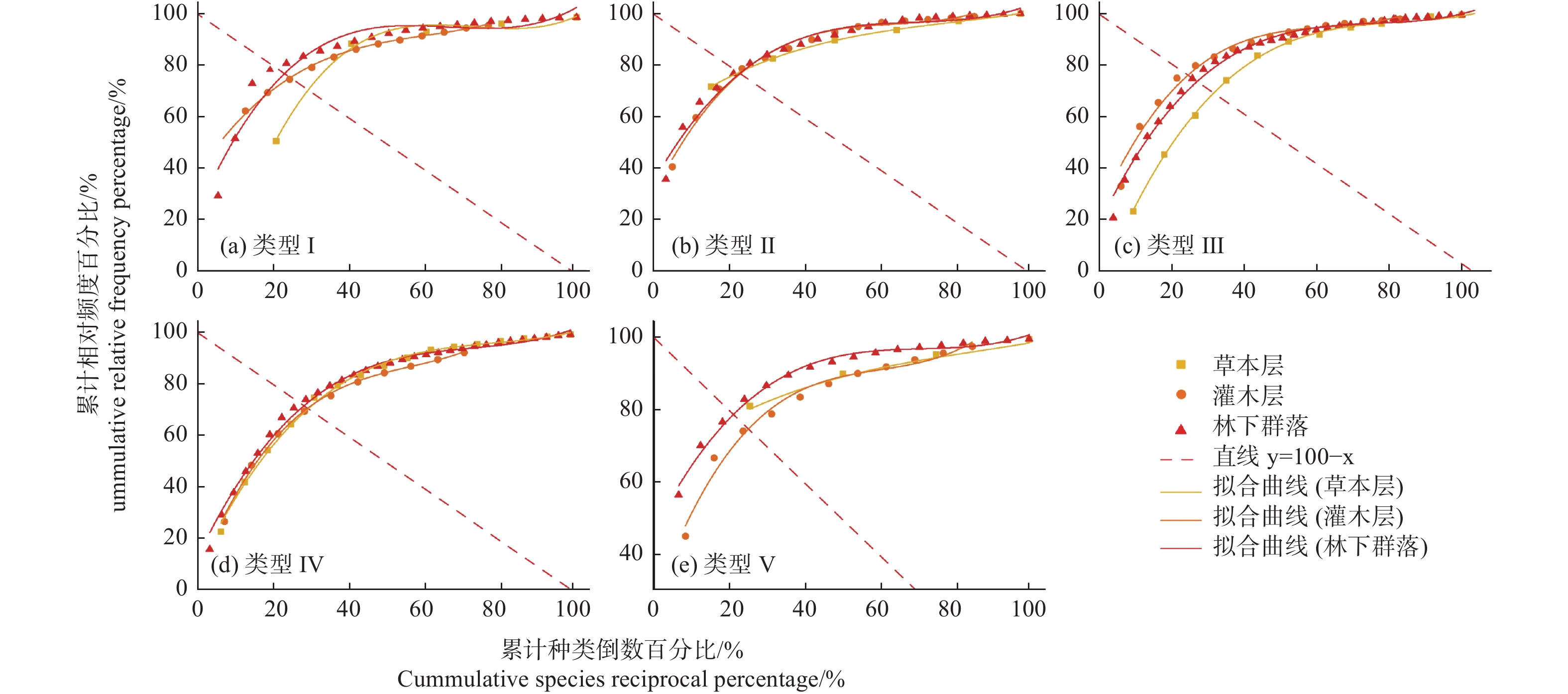
采用郑元润[14]改进的M.Godron方法计算林下不同层次的植物群落稳定性,绘制物种的累计种类倒数百分比(x)和累计相对频度百分比(y)散点图,进行曲线回归拟合,采用的拟和曲线和直线方程如下:
平滑曲线拟合方程:y = ax3 + bx2 + cx + d 或 y = ax2 + bx + c
直线方程:y = 100 − x
上述两方程的交点(x, y)即为稳定参考点,计算交点坐标与(20, 80)的之间的欧氏距离,该距离越小表示植物群落的稳定性越高。
-
使用单因素方差分析(ANOVA)和Tukey HSD检验,分析不同林分类型林下植被物种多样性指数的差异;分析物种多样性指数与土壤因子之间的Pearson相关性;采用冗余分析(RDA)研究不同林分类型林下不同层次物种组成与土壤因子的关系;使用方差分解(VP)方法分析各主要土壤因子对不同植物组成变化的解释率;采用一般线性回归,探究不同林分类型林下植物群落稳定性与物种多样性之间的关系。
采用Excel 2020对数据进行整理,统计分析过程使用R 4.0.2进行,其中ANOVA分析使用“car”程序包完成,Tukey HSD检验使用“multcomp”程序包完成,RDA、VP和ANOSIM分析使用“vegan”程序包完成,之后使用“ggplot2”程序包进行作图。
-
植物群落调查结果显示5种类型样地灌木层物种共40种,分属24科32属;草本层物种共26种,分属17科23属(表2)。其中III灌木层物种最多,IV最少;但IV草本层物种最多,V草本物种最少。灌木层方面,I以老鼠矢(Symplocos stellaris)为优势物种,II、III、V均以檵木为优势种;Ⅳ以木荷(Schima superba)、枫香为优势种。草本层方面,II、V均以芒萁为优势种;I以狗脊蕨(Woodwardia japonica (L. F.) Sm.)为优势种;III以苔草(Carex hirta L.)为优势种;IV以苔草、金毛耳草(Hedyotis chrysotricha (Palib.) Merr.)为优势种。
表 2 不同林分类型林下主要物种及其重要值
Table 2. Understory dominant species and their important values in different stand types
层次
Layer类型
Type科
Family属
Genus种
Species主要物种(重要值)
Dominant species (Important value)重要值累计/%
Accumulation灌木层
Shrub layerI 12 14 16 老鼠矢(0.39) + 青冈(0.10) + 黄栀(0.09) + 杉木(0.08) 66.36 II 12 13 16 檵木(0.29) + 油茶(0.22) + 槲栎(0.10) 61.16 III 14 17 20 檵木(0.26) + 苦槠(0.15) + 山莓(0.13) + 细齿叶柃(0.09) 63.13 IV 7 8 11 木荷(0.29) + 枫香(0.19) + 山莓(0.13) + 苦槠(0.10) 70.40 V 12 13 14 檵木(0.29) + 刺花悬钩子(0.24) + 油茶(0.13) 65.75 样地合计 Total 24 32 40 蔷薇科(Rosaceae)种类,金缕梅科
(Hamamelidaceae)数量最多草本层
Herb layerI 4 4 5 狗脊蕨(0.45) + 芒萁(0.33) 77.68 II 5 6 6 芒萁(0.55) + 金毛耳草(0.13) + 苔草(0.10) 78.80 III 10 12 12 苔草(0.26) + 海金沙(0.17) + 蕨(0.13) 55.58 IV 13 15 16 苔草(0.19) + 金毛耳草(0.16) + 霍香蓟(0.12) 46.77 V 3 3 4 芒萁(0.66) + 苔草(0.19) 85.36 样地合计 Total 17 23 26 菊科(Asteraceae)和莎草科(Cyperaceae)种类最多,
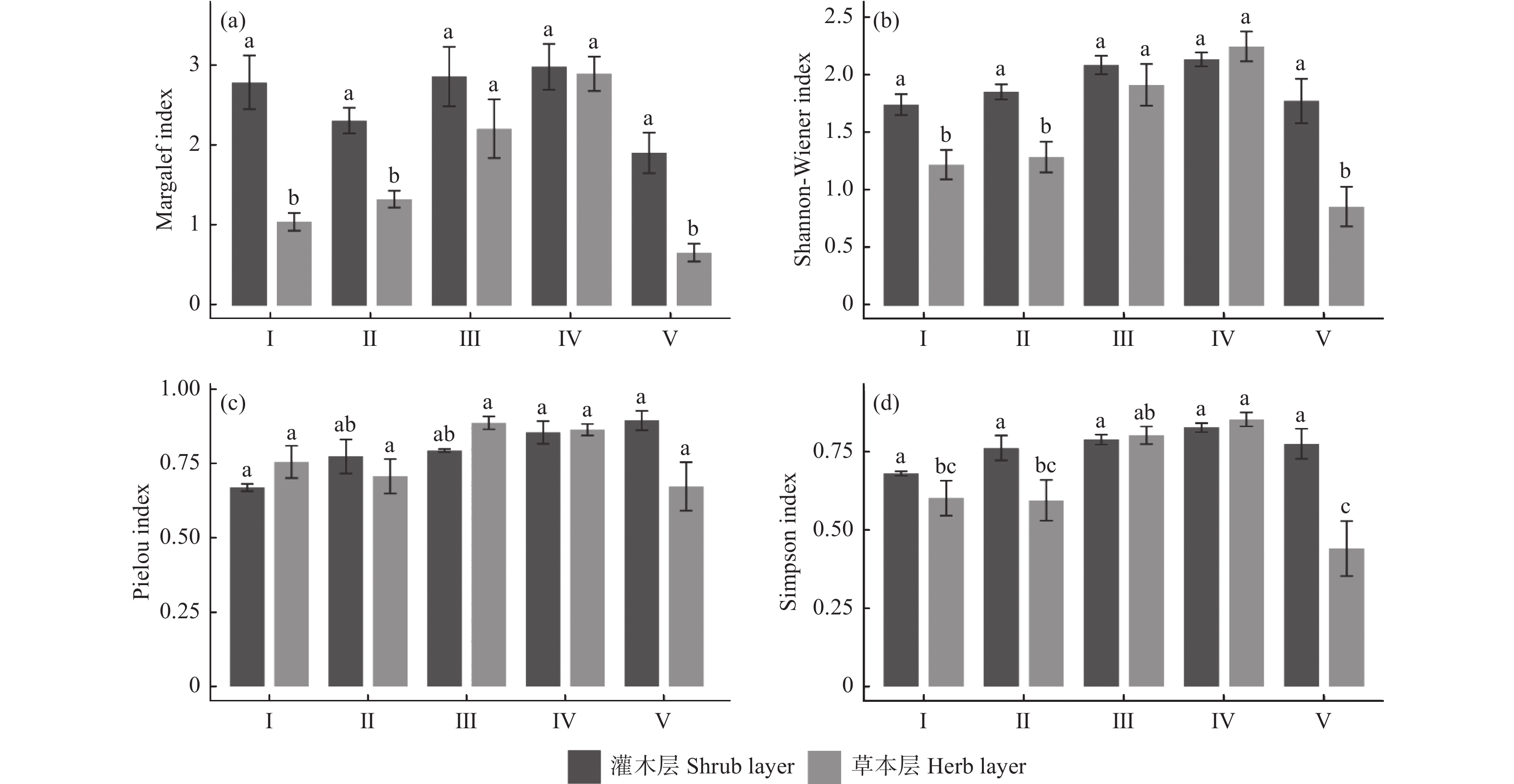
里百科(Gleicheniaceae)数量最多林下植被物种多样性方面,不同类型样地灌木层的Margalef指数、Shannon-Wiener指数和Simpson指数均无显著差异(图1 a、 b、 d),I的Pielou指数显著低于其他类型样地(图1 c)。而各类型样地林下草本层的多样性指数差异更大,其中III和IV的Margalef指数、Shannon-Wiener指数和Simpson指数显著高于其他样地,Pielou指数在不同林分类型间无显著差异。总体而言,III和IV的灌木层和草本层多样性较高,I和V草本层多样性较低。

图 1 不同林分类型的林下植被的物种多样性指数
Figure 1. The species diversity index of understory vegetation in different stand types
-
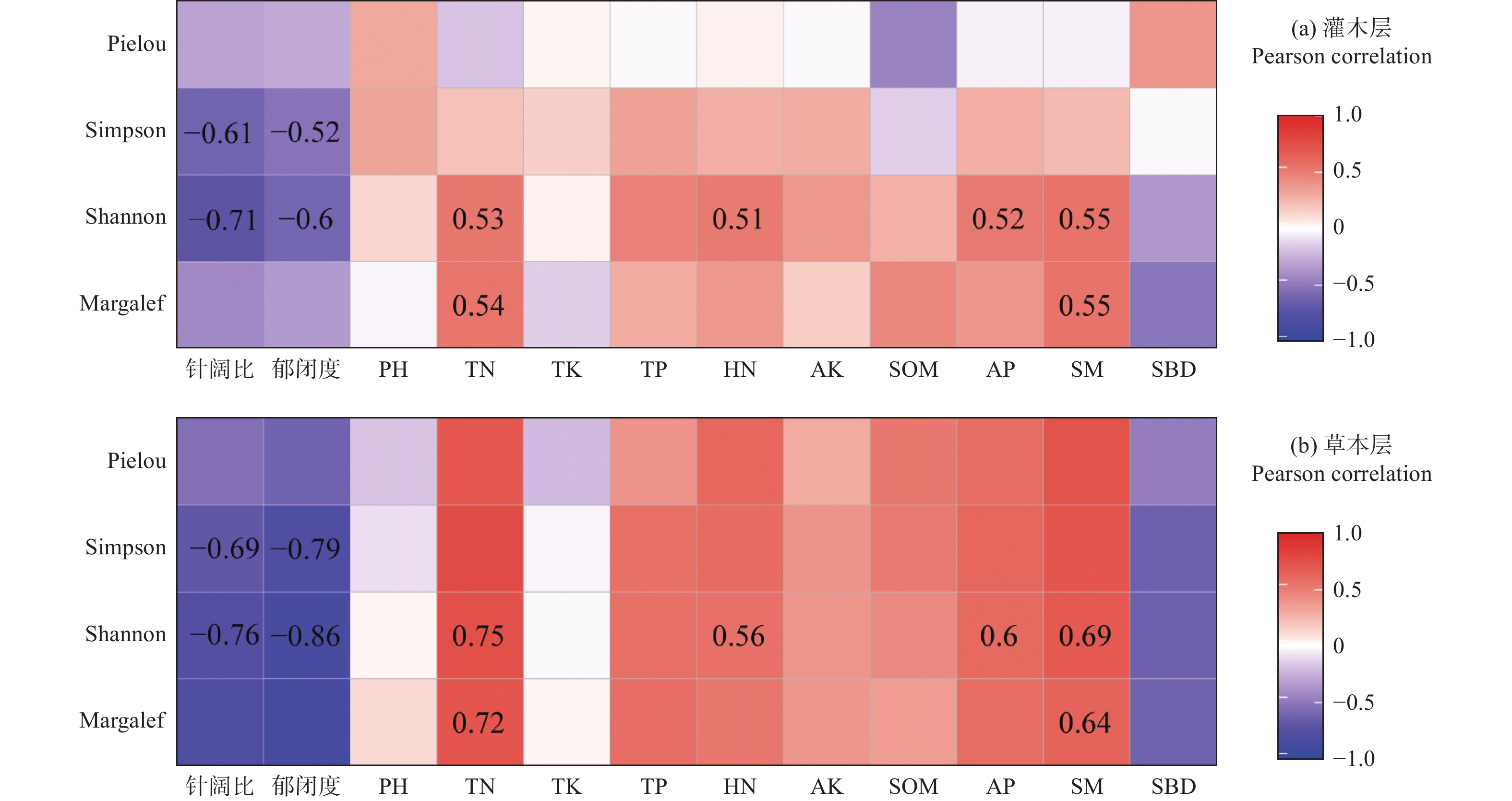
灌木层和草本层的结果类似(图2),Pielou指数与各林地特征因子均无显著相关性;Simpson指数仅与针阔比和郁闭度呈显著负相关,与土壤因子无显著相关性;Shannon-Wiener指数与针阔比、郁闭度呈显著负相关,与TN、HN、AP、SM呈显著正相关;Margalef指数仅与TN和SM呈显著正相关。以上相关性系数大小,草本层均大于灌木层。总体而言,除TN和SM外,林下植被物种多样性与其他土壤因子关系不密切。

图 2 林下植被物种多样性指数与林地特征的Pearson相关性
Figure 2. The Pearson correlation between species diversity of understory vegetation and plot characteristics
-
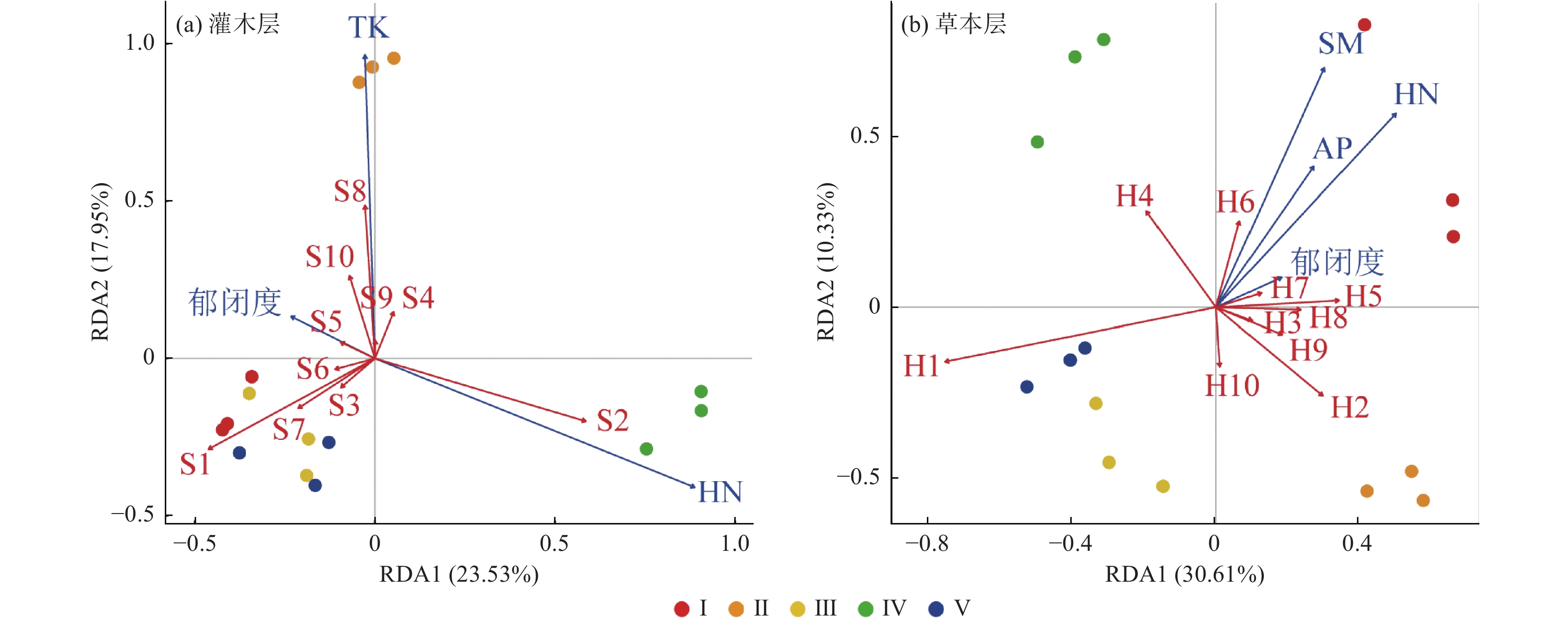
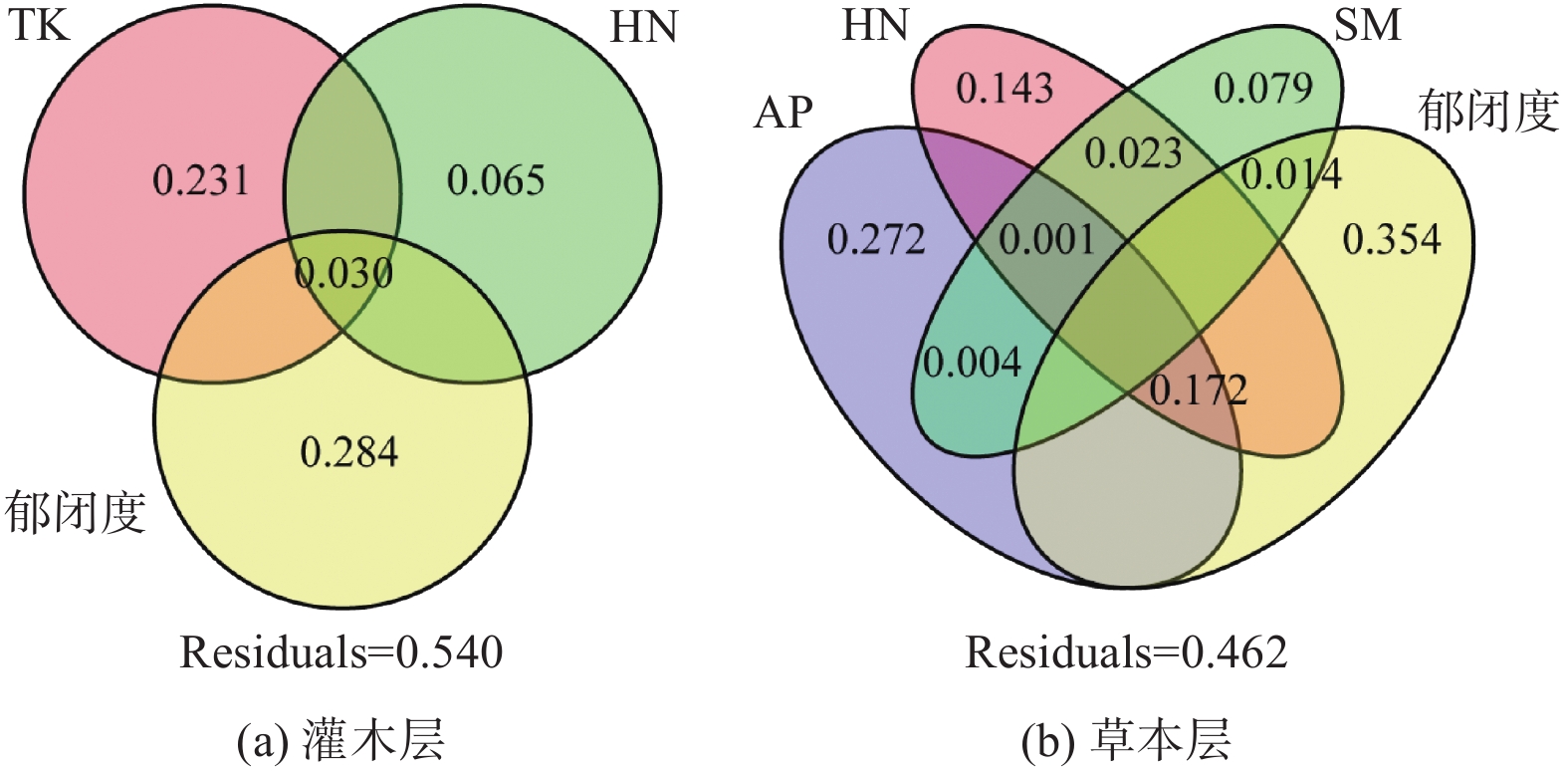
灌木层植被与林地因子RDA前两轴共同解释了物种组成变化的41.48%(图3 a),其中郁闭度、TK、HN为主要影响因素,分别单独解释了28.4%、23.1%和6.5%的方差(图4 a)。I、III、V的灌木层物种组成与土壤TK均呈负相关,与郁闭度呈正相关,Ⅱ的灌木层物种组成与土壤TK呈正相关,与HN呈负相关,而IV的情形与II恰好相反(图3 a)。从多度前10的物种来看,灌木层中优势种檵木主要与郁闭度呈显著正相关,老鼠矢主要与HN呈显著正相关(图3 a)。
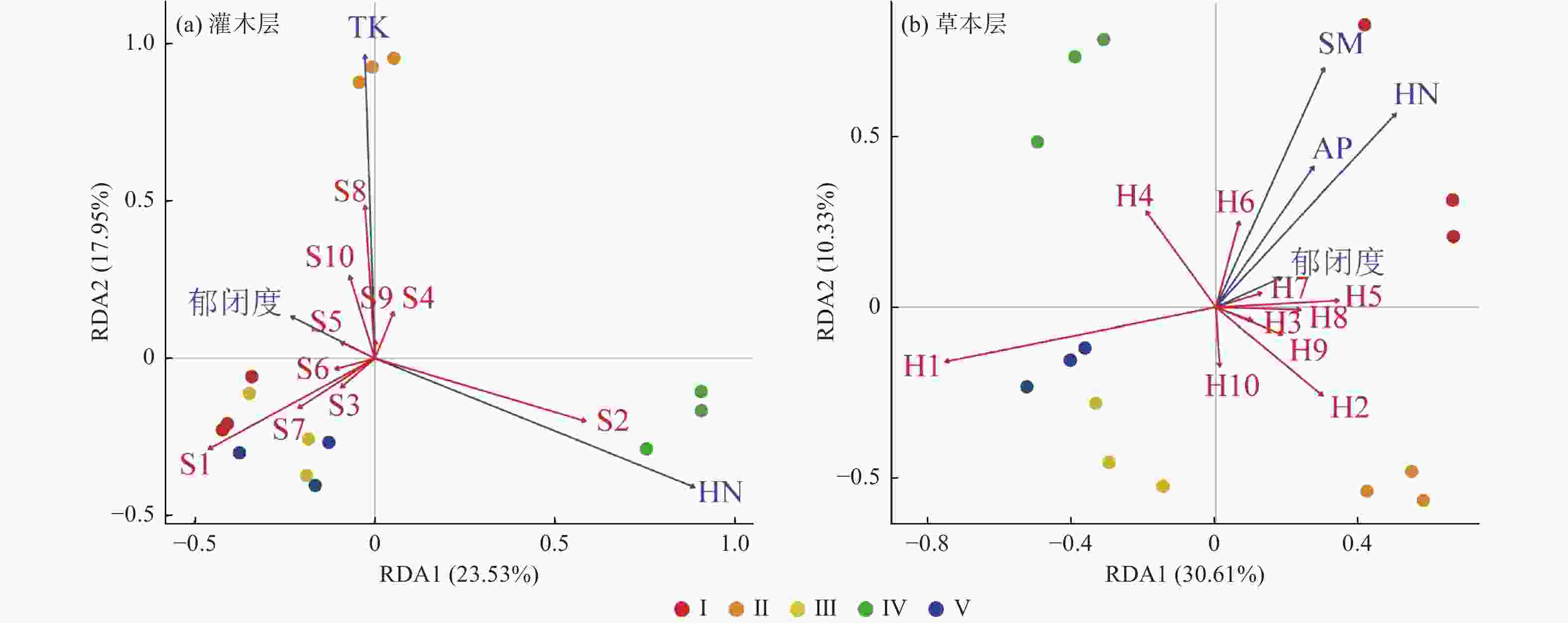
图 3 不同林分类型的林下主要植物种(多度排序前10的物种)和林地特征因子的RDA排序
Figure 3. The RDA ordination for understory main species (the top 10 species in abundance) and plot characteristics variables in different stand types

图 4 林下植物群落物种组成的林地解释因子方差分解
Figure 4. The variation partitioning of plot characteristics variables for the understory species composition
草本层植被与林地因子RDA前两轴共同解释物种组成变化的40.94%(图3 b),其中郁闭度、AP、HN、SM为主要影响因素,分别单独解释了35.4%、27.2%、14.3%和7.9%的方差,郁闭度、AP、HN共同解释了17.2%的方差,其他解释率极少(图4 b)。III和V的草本层物种组成与上述林地因子均呈负相关,I的草本层物种组成与上述林地因子呈正相关,II和IV与上述林地因子呈无显著相关性(图3 b)。从多度前10物种来看,草本层优势种芒萁与郁闭度、AP、HN、SM均呈负相关,金毛耳草与上述因子基本无显著相关性(图3 b)。
-
植物群落结构稳定性分析结果表明(图5和表3),不同类型样地的林下群落稳定性排序为:V(1.01)> I(2.86)> II(5.67)> III(7.80)> IV(11.32),其中灌木层群落稳定性排序为:III(3.85)> I(4.80)> II(5.78)> V(6.81)> IV(12.55),草本层群落稳定性排序为:V(1.54)> II(4.94)> I(11.84)> IV(13.16)> III(14.45)。总体来看,对照和疏伐促新样地的林下植物群落稳定性较高,而皆伐补阔样地的林下植物群落暂时仍为较不稳定状态。分不同层次来看,皆伐补植林分灌木层的群落稳定性远低于其他林分,暂时处于不稳定状态;对照林分草本层的群落稳定性远高于其他林分,处于较稳定状态,而皆伐补植和促新林分的草本层群落稳定性较低,暂时处于不稳定状态。
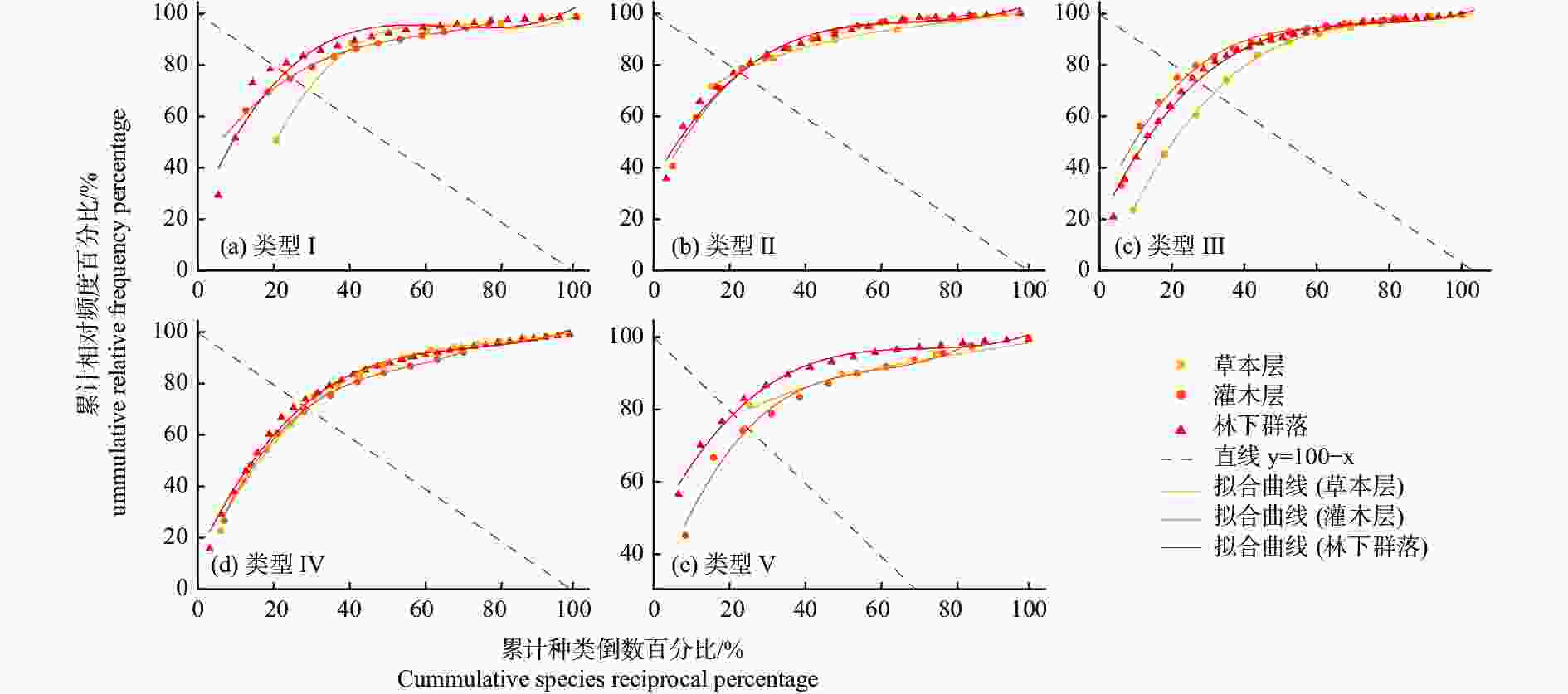
图 5 不同林分类型的林下植物群落稳定性的曲线拟合
Figure 5. The curve fitting of understory community stability in different stand types
表 3 不同林分类型的林下植物群落稳定性计算结果
Table 3. The results of understory community stability in different stand types
类型
Type层次
Layer拟合曲线
Fitted curveR2 交点坐标
Coordinate of intersection欧氏距离
Euclidean distanceI 灌木层 Shrub layer y=0.00017x3−0.031x2 + 2.13x + 41.54 0.997 (23.35,76.56) 4.80 草本层 Herb layer y=0.00038x3−0.074x2 + 5.26x-26.00 0.965 (28.42,71.68) 11.84 林下群落 Understory y=0.00026x3−0.051x2 + 3.31x + 27.22 0.925 (22.08,78.04) 2.86 II 灌木层 Shrub layer y=0.00023x3−0.044x2 + 3.02x + 25.72 0.988 (24.11,75.93) 5.78 草本层 Herb layer y=0.00005x3−0.012x2 + 1.17x + 54.97 0.999 (23.54,76.56) 4.94 林下群落 Understory y=0.00017x3−0.036x2 + 2.65x + 31.03 0.974 (24.07,76.05) 5.67 III 灌木层 Shrub layer y=0.00037x3−0.064x2 + 3.74x + 20.97 0.980 (22.75,77.30) 3.85 草本层 Herb layer y=0.00018x3−0.043x2 + 3.56x-3.25 0.999 (30.21,69.78) 14.45 林下群落 Understory y=0.00018x3−0.04x2 + 2.98x + 20.87 0.999 (25.57,74.54) 7.80 IV 灌木层 Shrub layer y=0.00039x3−0.064x2 + 3.87x + 3.92 0.996 (28.99,71.24) 12.55 草本层 Herb layer y=0.00017x3−0.041x2 + 3.27x + 6.08 0.997 (29.28,70.67) 13.16 林下群落 Understory y=0.00018x3−0.039x2 + 3.05x + 13.35 0.999 (28.08,72.07) 11.32 V 灌木层 Shrub layer y=0.00024x3−0.044x2 + 2.81x + 28.93 0.978 (24.95,75.32) 6.81 草本层 Herb layer y=-0.0019x2 + 0.492x + 70.45 0.995 (21.07,78.89) 1.54 林下群落 Understory y=0.000130x3−0.028x2 + 2.01x + 48.36 0.988 (20.80,79.38) 1.01 -
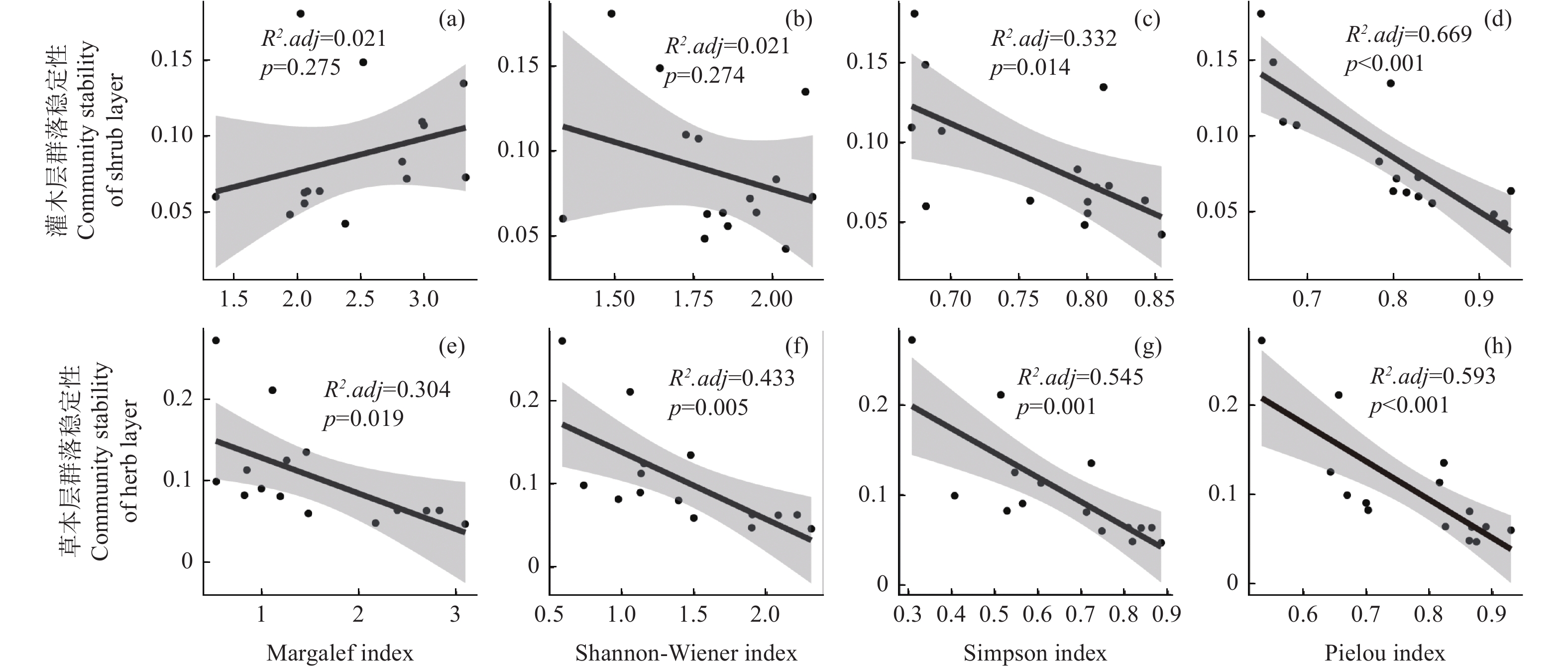
林下不同层次植物群落稳定性(此处取欧氏距离的倒数)与物种多样性的线性回归结果表明,灌木层和草本层群落稳定性与物种多样性指数呈现不同的关系。具体而言,林下灌木层群落稳定性与Margalef丰富度指数(图6 a)和Shannon-Wiener多样性指数(图6 b)无显著相关性(p > 0.05),与Simpson优势度指数(图6 c)和Pielou均匀度指数(图6 d)呈显著负相关性(p < 0.05)。林下草本层群落稳定性与四种指数(图6 e~h)均呈显著负相关(p < 0.05),其中Pielou指数(R2.adj = 0.593)和Simpson指数(R2.adj = 0.545)对草本层群落稳定性的解释率要高于Shannon-Wiener指数(R2.adj = 0.433)和Margalef指数(R2.adj = 0.304)。
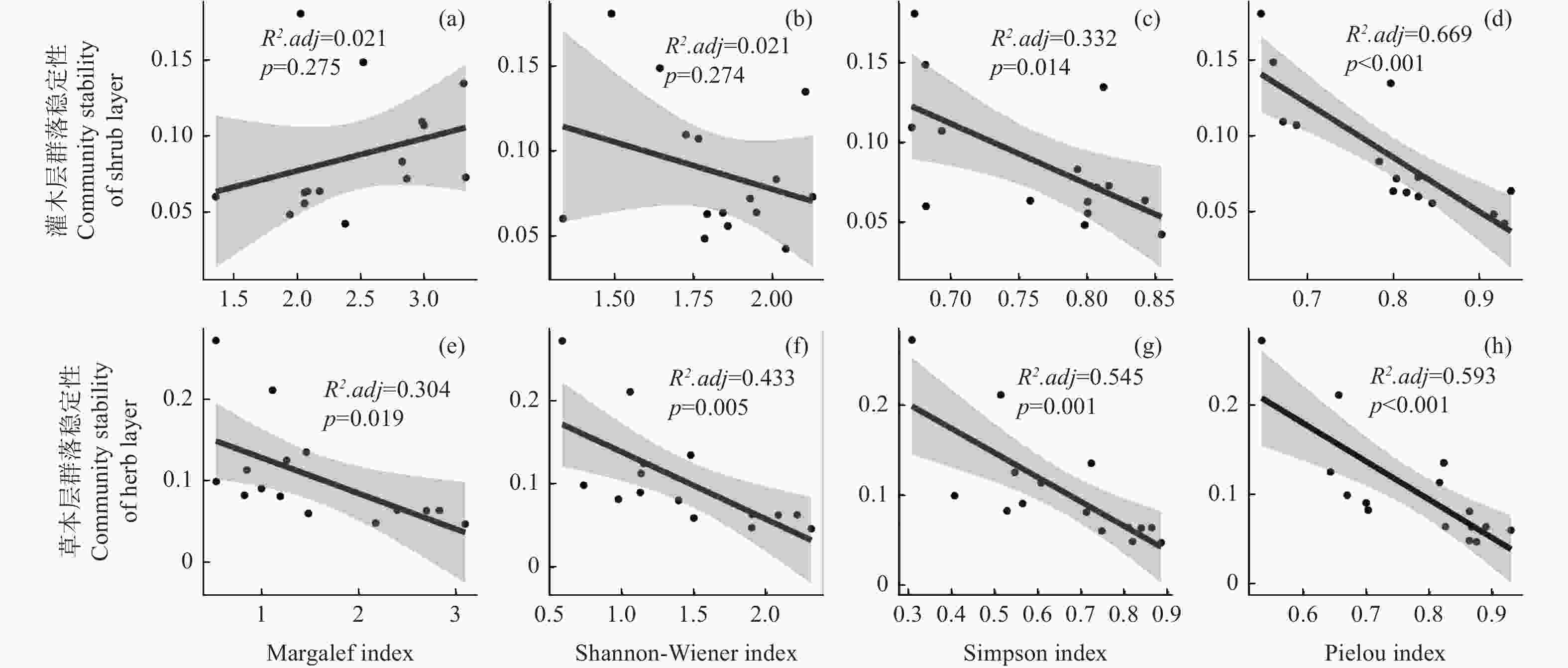
图 6 林下不同层次植物群落稳定性与物种多样性的线性回归
Figure 6. Linear regression analysis of community stability and species diversity in different understory vegetation layers
-
马尾松林阔叶化改造后不同林分灌木层优势种差异不大,除去补植的树种外,疏伐林分(I、II)檵木和老鼠矢幼树为主要优势种,而苦槠幼树在大部分样地也均有分布。这主要是由于疏伐后林分郁闭度降低,檵木和老鼠矢等喜光耐阴物种在疏伐初期更易占据优势地位。林分改造前后的草本层优势种差异较大,马尾松次生纯林(V)郁闭度较高,林下光照有限,虽然会限制林下木本植物生长,但却有利于芒萁、狗脊蕨等耐荫蕨类的生长[15]。随着采伐强度增大,皆伐林分(III、IV)林下蕨类植物显著减少,草本层以苔草、金毛耳草为优势种,这与前人研究类似[16]。在物种多样性方面,改造措施对灌木层多样性无显著影响,但皆伐抚育对草本层多样性要高于疏伐无抚育,且均显著高于对照。说明木本物种对生长空间要求更高,其物种多样性变化相对于草本层有一定的滞后性[11]。但林分改造的效果具有长期性,林下植物多样性也具有动态变化过程,有研究表明一定时期后林下灌木会抑制草本的生长发育[17],因此还需长期观测分析。
从影响因素来看,本研究表明,郁闭度、针阔比、土壤理化性质等林地因子对林下植物组成的解释率在40%左右,即仍有其他生物(土壤微生物)或非生物(林分结构、地形等)因子发挥重要作用,其影响过程也较复杂[18]。另外,林地因子对草本层物种组成的影响程度要高于灌木层,这是由于III、IV灌木层有大量物种为栽植的幼苗(幼树),现阶段主要受人为作用的影响;而其他灌木幼苗自然更新时间较草本更长,因此灌木层物种组成暂时与林地因子的关系有限。具体而言,郁闭度是影响林下植被物种组成和多样性最主要的因素之一。随着采伐强度增大,林分郁闭度下降,林地光照和空间资源更充足,能为更多物种的侵入、定居和更新提供更多生态位空间,有利于林下草本的共存和生长[19]。当然,土壤氮、有效磷和水分也是影响林下植物组成和多样性的重要因素,针阔比也与林下植物多样性显著相关。一方面,针阔混交林同时具备了阔叶林和针叶林的林分特征,冠层结构更丰富,增加了生境异质性,提高了林地更新潜力,有利于幼树生长[20-21]。另一方面,针叶凋落物质地粗硬,纤维素含量高不易分解,导致土壤氮含量较低[22];而林分改造后阔叶凋落物拥有较高的N、P含量及较低的木质素含量,且N、P释放速度较快[23],还能有效增强土壤表层的保水能力[24];而且不同树种交错的根系可增强土壤养分归还和循环能力,改善林地土壤通气状况和水分条件,有助于林下草本生长[25-26]。
-
有研究表明,针叶林阔叶化改造形成的复层林冠结构提升了空间资源的利用率,且树种多样性的增加有利于增强林地抗逆性,进而提高生态系统的稳定性[27]。但本研究表明对照林分的林下植物群落稳定性最高,疏伐林分(I、II)的林下植被处于亚稳定状态,群落稳定性高于皆伐样地(III、IV),且IV林下植被的稳定性最低,暂时仍为不稳定状态。这是由于Godron稳定性测定遵循“优势度效应理论”,即植物种群维持稳定性机制的主要途径是繁殖更新,优势物种增加会提高群落稳定性[28]。I、II马尾松间伐比例不高,且未进行割灌、除草,其林下植被与对照类似,主要以芒萁或狗脊蕨占据了大量的生态位,抑制了其他草本植物的生长。而III、IV林下群落尚在竞争初级阶段,草本层优势种还未完成新生态位的侵占和定居,物种组成仍处于动态发展中,群落结构和功能尚不稳定[29];但随着演替的持续进行,林下植被物种组成和群落结构会逐渐趋于稳定,生态功能进一步增强[30-31]。
物种多样性是反映植物群落物种组成、更新演替以及稳定性评价等方面的重要指标[32],一般认为物种多样性越高,群落结构越复杂,其稳定性也越高[33]。但也有研究表明物种多样性与稳定性具有更复杂的关系[34-35]。主要原因是群落稳定性的驱动力并不仅限于物种多样性,可能还包含功能冗余和功能多样性[36]。从本研究结果来看,灌木层群落稳定性与物种多样性的相关性较低,而草本层群落稳定性与物种多样性呈显著负相关,这与汪永文等[37]在安徽马尾松混交林的研究结果相似。植物群落稳定性受种内和种间竞争、环境压力以及人为活动干扰等生物和非生物因素的共同影响[38],并不能仅以物种多样性来反映群落稳定性,尤其是处于群落构建的初期阶段。此外,本研究在改造样地内也发现了一定程度的人为干扰(如樵采等),这些都对林下群落稳定性产生不利影响,故需加强对林下群落的保护与管理,保证其顺利进行正向更新演替,提高植物群落的稳定性。
-
(1)马尾松林阔叶化改造措施对林下草本层的影响要高于灌木层。除疏伐促新外,其他改造措施的林下植被物种多样性指数均显著高于对照林地,其中皆伐抚育的林下物种多样性最高。林地特征因子对草本层物种组成的影响程度高于灌木层,其中郁闭度是最显著的影响因子。
(2)对照林地的林下植物群落稳定性最高,疏伐林地次之,皆伐林地最低;林下灌木层稳定性与物种多样性相关程度较低,而草本层稳定性与物种多样性呈显著负相关。
(3)从林下植物群落稳定性及可持续经营角度,若非必要,不宜对马尾松林进行皆伐,抚育措施应适度渐进式开展,促使针阔混交林逐渐过渡到阔叶混交林;在林分改造初期,尽量减少林下人为扰动,促进林下植被更新和土壤养分积累。
千岛湖区马尾松林改造后的林下植物多样性和群落稳定性
Diversity and Community Stability of Understory Vegetation in the Pinus massoniana Forest after Transformation in Thousand-island Lake Area
-
摘要:
目的 马尾松针叶林阔叶化改造有利于提升森林生态系统结构和功能,对林下植被的更新和演替也有着重要影响。分析林分改造后的林下植物多样性和稳定性特征,可为森林的可持续经营提供参考。 方法 本研究以淳安县千岛湖林区马尾松不同改造林分(I 疏伐促新、II 疏伐补阔、III 皆伐促新、IV 皆伐补阔)为对象,周边无改造措施林分(V)为对照,进行植被调查、土壤采样与测定,分析了林下植被物种组成和多样性与林地特征因子的关系,并探究了林下群落稳定性与物种多样性的关系。 结果 样地间的林下灌木层优势种的差异低于草本层,V的林下物种多样性显著低于改造后林地(p < 0.05),IV的草本层物种多样性显著高于其它林地(p< 0.05)。林地特征因子能解释林下植物种变化的40%,且对草本层的影响程度高于灌木层,其中郁闭度是最主要的影响因素。在马尾松林分改造初期,林下植物群落稳定性V > I > II > III > IV,林下灌木层稳定性与物种多样性的相关程度较低,而草本层稳定性与物种多样性呈显著负相关(p< 0.05)。 结论 千岛湖区马尾松林阔叶化改造初期,主要通过降低郁闭度显著提高了林下草本层多样性,但皆伐措施的林下植物群落稳定性较低。因此,综合考虑植物多样性和稳定性维持,对马尾松林采取适当间伐等抚育措施能促进其林下植被的发展。 Abstract:Objective Transforming Pinus massoniana coniferous forest into broadleaved forest is beneficial to improving the structure and function of forest ecosystem and has significant influence on the regeneration and succession of understory vegetation. The analysis of the diversity and community stability of understory vegetation after forest transformation can provide reference for sustainable forest management. Method In this study, based on the four types of stand transformation (i.e., I thinning and promoting regeneration, II thinning and planting broadleaf trees, III clear cutting and promoting regeneration, clear cutting and IV clear cutting and planting broadleaf trees) of Pinus massoniana forests in Thousand-island Lake Forest area, Chun’an county, and control treatment (V no-transformation forest )), the species composition, diversity and community stability of understory vegetation were investigated. The relationship between understory species composition and stand factors was analyzed, and the relationship between species diversity and community stability was also explored. Result The difference of understory dominant species among different stands was lower in shrub layer than in herb layer. The understory species diversity in control group was significantly lower than that in transformed stand (P < 0.05) and the species diversity of herb layer in type IV was significantly higher than that in other types (P < 0.05). The stand factors could explain about 40% of understory species variation and had higher effect on the species composition of herb layer than that of shrub layer. Among these, the canopy density was the most important factor. In the early stage of stand transformation, the understory community stability in mode V was the best, followed by I, II, III, and IV. The community stability of shrub layer had low correlation with species diversity, while the stability of herb layer was negatively correlated with species diversity (P < 0.05). Conclusion In the early stage of broad-leaf transformation of P. massoniana forest in Thousand-island Lake area, the diversity of herbaceous layer was significantly improved mainly by reducing canopy density, but the stability of understory vegetation community was low under clear cutting treatments. Therefore, considering vegetation diversity and community stability, appropriate thinning and tending measures can promote the development of understory vegetation in P. massoniana forest. -
图 1 不同林分类型的林下植被的物种多样性指数
Figure 1. The species diversity index of understory vegetation in different stand types
图 2 林下植被物种多样性指数与林地特征的Pearson相关性
Figure 2. The Pearson correlation between species diversity of understory vegetation and plot characteristics
图 3 不同林分类型的林下主要植物种(多度排序前10的物种)和林地特征因子的RDA排序
Figure 3. The RDA ordination for understory main species (the top 10 species in abundance) and plot characteristics variables in different stand types
图 4 林下植物群落物种组成的林地解释因子方差分解
Figure 4. The variation partitioning of plot characteristics variables for the understory species composition
图 5 不同林分类型的林下植物群落稳定性的曲线拟合
Figure 5. The curve fitting of understory community stability in different stand types
图 6 林下不同层次植物群落稳定性与物种多样性的线性回归
Figure 6. Linear regression analysis of community stability and species diversity in different understory vegetation layers
表 1 研究区不同类型林分特征
Table 1. Stand characteristics of different types in the study area
林型(编号)
Type (Number)郁闭度
Canopy density主要树种
Main species树种占比/%
Proportion重要值
Important value胸径
DBH/cm树高
Height/m冠幅
Crown breadth/m疏伐促新(I)
Thinning and
promoting
regeneration0.70 杉木
Cunninghamia lanceolata32.4 0.225 11.83 ± 4.53 b 8.71 ± 1.92 b 3.17 ± 0.96 b 马尾松
Pinus massoniana4.5 0.151 36.40 ± 2.17 a 21.17 ± 2.04 a 6.27 ± 0.98 a 老鼠矢
Symplocos stellaris Brand15.5 0.068 4.00 ± 1.76 c 4.28 ± 1.18 c 1.93 ± 0.69 c 疏伐补阔(II)
Thinning and
supplementing
broadleaf0.65 马尾松
Pinus massoniana31.3 0.445 12.11 ± 5.30 a 8.83 ± 2.71 a 3.08 ± 1.00 a 油茶
Camellia oleifera35.6 0.247 4.64 ± 2.16 b 3.39 ± 0.29 b 1.90 ± 0.60 b 槲栎
Quercus aliena9.8 0.083 4.13 ± 0.90 b 3.37 ± 0.27 b 1.74 ± 0.33 b 皆伐促新(III)
Clear cutting and
promoting
regeneration0.55 苦槠
Castanopsis sclerophylla27.4 0.293 11.25 ± 4.25 b 7.20 ± 2.19 b 2.91 ± 1.25 b 杉木
Cunninghamia lanceolata11.6 0.188 16.78 ± 2.72 a 9.95 ± 0.96 a 3.56 ± 0.84 a 枫香
Liquidambar formosana25.3 0.166 2.68 ± 0.28 c 3.17 ± 0.27 c 1.20 ± 0.30 c 皆伐补阔(IV)
Clear cutting and
supplementing
broadleaf0.30 枫香
Liquidambar formosana25.0 0.451 9.03 ± 1.88 a 3.00 ± 0.25 b 1.40 ± 0.35 b 野柿
Diospyros kaki var.
sylvestris
Makino50.0 0.285 1.08 ± 0.04 c 4.70 ± 0.28 a 2.68 ± 0.33 a 紫叶李
Prunus cerasifera
‘Atropurpurea’25.0 0.264 4.72 ± 1.03 b 3.04 ± 0.30 b 1.64 ± 0.45 b 对照(V)
Control group0.75 马尾松
Pinus massoniana40.5 0.555 21.30 ± 4.59 a 21.90 ± 4.29 a 4.13 ± 1.40 a 檵木
Loropetalum chinense30.0 0.167 4.55 ± 1.11 b 4.97 ± 1.33 b 1.74 ± 0.72 b 油茶
Camellia oleifera7.9 0.080 5.30 ± 1.99 b 3.56 ± 1.31 b 2.24 ± 148 b  下载: 导出CSV
下载: 导出CSV
表 2 不同林分类型林下主要物种及其重要值
Table 2. Understory dominant species and their important values in different stand types
层次
Layer类型
Type科
Family属
Genus种
Species主要物种(重要值)
Dominant species (Important value)重要值累计/%
Accumulation灌木层
Shrub layerI 12 14 16 老鼠矢(0.39) + 青冈(0.10) + 黄栀(0.09) + 杉木(0.08) 66.36 II 12 13 16 檵木(0.29) + 油茶(0.22) + 槲栎(0.10) 61.16 III 14 17 20 檵木(0.26) + 苦槠(0.15) + 山莓(0.13) + 细齿叶柃(0.09) 63.13 IV 7 8 11 木荷(0.29) + 枫香(0.19) + 山莓(0.13) + 苦槠(0.10) 70.40 V 12 13 14 檵木(0.29) + 刺花悬钩子(0.24) + 油茶(0.13) 65.75 样地合计 Total 24 32 40 蔷薇科(Rosaceae)种类,金缕梅科
(Hamamelidaceae)数量最多草本层
Herb layerI 4 4 5 狗脊蕨(0.45) + 芒萁(0.33) 77.68 II 5 6 6 芒萁(0.55) + 金毛耳草(0.13) + 苔草(0.10) 78.80 III 10 12 12 苔草(0.26) + 海金沙(0.17) + 蕨(0.13) 55.58 IV 13 15 16 苔草(0.19) + 金毛耳草(0.16) + 霍香蓟(0.12) 46.77 V 3 3 4 芒萁(0.66) + 苔草(0.19) 85.36 样地合计 Total 17 23 26 菊科(Asteraceae)和莎草科(Cyperaceae)种类最多,
里百科(Gleicheniaceae)数量最多
下载: 导出CSV
表 3 不同林分类型的林下植物群落稳定性计算结果
Table 3. The results of understory community stability in different stand types
类型
Type层次
Layer拟合曲线
Fitted curveR2 交点坐标
Coordinate of intersection欧氏距离
Euclidean distanceI 灌木层 Shrub layer y=0.00017x3−0.031x2 + 2.13x + 41.54 0.997 (23.35,76.56) 4.80 草本层 Herb layer y=0.00038x3−0.074x2 + 5.26x-26.00 0.965 (28.42,71.68) 11.84 林下群落 Understory y=0.00026x3−0.051x2 + 3.31x + 27.22 0.925 (22.08,78.04) 2.86 II 灌木层 Shrub layer y=0.00023x3−0.044x2 + 3.02x + 25.72 0.988 (24.11,75.93) 5.78 草本层 Herb layer y=0.00005x3−0.012x2 + 1.17x + 54.97 0.999 (23.54,76.56) 4.94 林下群落 Understory y=0.00017x3−0.036x2 + 2.65x + 31.03 0.974 (24.07,76.05) 5.67 III 灌木层 Shrub layer y=0.00037x3−0.064x2 + 3.74x + 20.97 0.980 (22.75,77.30) 3.85 草本层 Herb layer y=0.00018x3−0.043x2 + 3.56x-3.25 0.999 (30.21,69.78) 14.45 林下群落 Understory y=0.00018x3−0.04x2 + 2.98x + 20.87 0.999 (25.57,74.54) 7.80 IV 灌木层 Shrub layer y=0.00039x3−0.064x2 + 3.87x + 3.92 0.996 (28.99,71.24) 12.55 草本层 Herb layer y=0.00017x3−0.041x2 + 3.27x + 6.08 0.997 (29.28,70.67) 13.16 林下群落 Understory y=0.00018x3−0.039x2 + 3.05x + 13.35 0.999 (28.08,72.07) 11.32 V 灌木层 Shrub layer y=0.00024x3−0.044x2 + 2.81x + 28.93 0.978 (24.95,75.32) 6.81 草本层 Herb layer y=-0.0019x2 + 0.492x + 70.45 0.995 (21.07,78.89) 1.54 林下群落 Understory y=0.000130x3−0.028x2 + 2.01x + 48.36 0.988 (20.80,79.38) 1.01
下载: 导出CSV
-
[1] De LOMBAERDE E, BAETEN L, VERHEYEN K, et al. Understorey removal effects on tree regeneration in temperate forests: A meta‐analysis[J]. Journal of Applied Ecology, 2021, 58(1): 9-20. doi: 10.1111/1365-2664.13792 [2] WEI L P, ARCHAUX F, HULIN F, et al. Stand attributes or soil micro-environment exert greater influence than management type on understory plant diversity in even-aged oak high forests[J]. Forest Ecology and Management, 2020, 460: 117897. doi: 10.1016/j.foreco.2020.117897 [3] ECHIVERRI L, MACDONALD S E. A topographic moisture index explains understory vegetation response to retention harvesting[J]. Forest Ecology and Management, 2020, 474: 118358. doi: 10.1016/j.foreco.2020.118358 [4] SU X P, LI S J, WAN X H, et al. Understory vegetation dynamics of Chinese fir plantations and natural secondary forests in subtropical China[J]. Forest Ecology and Management, 2021, 483: 118750. doi: 10.1016/j.foreco.2020.118750 [5] 吴 文, 李月辉, 胡远满. 不同营林措施对林下层多样性和群落结构的影响[J]. 生态环境学报, 2018, 27(7):1369-1376. [6] 王佳佳, 贺 涛, 张 沂, 等. 间伐强度对阿尔泰山天然林下植被的影响[J]. 生态学报, 2022, 42(23):9761-9768. [7] 薛世玉, 李小飞, 徐大平, 等. 抚育措施对檀香 + 降香黄檀混交林林下植被多样性的影响[J]. 热带作物学报, 2021, 42(8):2422-2429. [8] LI X K, LI Y N, ZHANG J, et al. The effects of forest thinning on understory diversity in China: A meta-analysis[J]. Land Degradation & Development, 2020, 31(10): 1225-1240. [9] 戎福仁, 王祖华, 吴初平, 等. 生境片段化对千岛湖次生马尾松林土壤理化性质的影响[J]. 浙江大学学报(理学版), 2018, 45(6):748-755 + 764. [10] 王秀云, 朱汤军, 陆 超, 等. 千岛湖马尾松林抚育改造后主要种群生态位研究[J]. 浙江林业科技, 2014, 34(3):1-7. [11] 刘思泽, 尹海锋, 沈 逸, 等. 间伐强度对马尾松人工林间伐初期林下植被群落植物群落物种组成和多样性的影响[J]. 应用生态学报, 2020, 31(9):2866-2874. [12] 孙千惠, 吴 霞, 王媚臻, 等. 林分密度对马尾松林林下物种多样性和土壤理化性质的影响[J]. 应用生态学报, 2018, 29(3):732-738. [13] 张甘霖, 龚子同. 土壤调查实验室分析方法[M]. 北京: 科学出版社, 2012. [14] 郑元润. 森林群落稳定性研究方法初探[J]. 林业科学, 2000, 36(5):28-32. [15] 姚 珊, 郭其强, 盘金文, 等. 马尾松人工林发育过程中林下灌草群落优势种生态位动态分析[J]. 西北植物学报, 2021, 41(8):1417-1424. [16] 李 萌, 陈永康, 徐浩成, 等. 不同间伐强度对南亚热带杉木人工林林下植物功能群的影响[J]. 生态学报, 2020, 40(14):4985-4993. [17] 崔佳佳, 铁 牛. 大兴安岭北部森林群落结构及植物多样性特征研究[J]. 西北林学院学报, 2021, 36(2):24-30. [18] DENG J J, FANG S, FANG X M, et al. Forest understory vegetation study: current status and future trends[J]. Forestry Research, 2023, 3: 6. [19] STEIN A, GERSTNER K, KREFT H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales[J]. Ecology letters, 2014, 17(7): 866-880. doi: 10.1111/ele.12277 [20] WILSON D S, PUETTMANN K J. Density management and biodiversity in young Douglas-fir forests: Challenges of managing across scales[J]. Forest Ecology and Management, 2007, 246(1): 123-134. doi: 10.1016/j.foreco.2007.03.052 [21] 孙冬婧, 温远光, 罗应华, 等. 近自然化改造对杉木人工林物种多样性的影响[J]. 林业科学研究, 2015, 28(2):202-208. [22] 张博文, 秦 娟, 任忠明, 等. 坡向对北亚热带区马尾松纯林及不同针阔混交林型林下植物多样性的影响[J]. 生态环境学报, 2022, 31(6):1091-1100. [23] 秦 娟, 孔海燕, 刘 华. 马尾松不同林型土壤C、N、P、K的化学计量特征[J]. 西北农林科技大学学报(自然科学版), 2016, 44(2):68-76 + 82. [24] 耿 琦, 王海燕, 张美娜, 等. 森林枯落物持水特性影响因素研究进展[J]. 生态科学, 2020, 39(5):220-226. [25] OYONARTE C, ARANDA V, DURANTE P. Soil surface properties in Mediterranean mountain ecosystems: Effects of environmental factors and implications of management[J]. Forest Ecology and Management, 2008, 254(2): 156-165. doi: 10.1016/j.foreco.2007.07.034 [26] WAN X H, HUANG Z Q, HE Z M, et al. Soil C: N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations[J]. Plant and soil, 2015, 387(1-2): 103-116. doi: 10.1007/s11104-014-2277-4 [27] JACTEL H, GRITTI E S, DROSSLER L, et al. Positive biodiversity-productivity relationships in forests: climate matters[J]. Biol Lett, 2018, 14(4): 20170747. doi: 10.1098/rsbl.2017.0747 [28] GRMAN E, LAU J A, SCHOOLMASTER JR D R, et al. Mechanisms contributing to stability in ecosystem function depend on the environmental context[J]. Ecology letters, 2010, 13(11): 1400-1410. doi: 10.1111/j.1461-0248.2010.01533.x [29] 杜忠毓, 王剑武, 邢文黎, 等. 喀斯特锑矿植被恢复区植物多样性及群落稳定性[J]. 环境科学研究, 2023, 36(1):188-197. [30] 吴 昊, 贾少奇, 朱亚星. 物种多样性及环境因子对入侵植物空心莲子草群落稳定性的影响[J]. 生态学杂志, 2022, 41(1):33-41. [31] 郑晓阳, 赵 冲, 刘青青, 等. 成熟杉木人工林林下草本层生态位特征[J]. 生态学杂志, 2018, 37(2):332-338. [32] GAO T, HEDBLOM M, EMILSSON T, et al. The role of forest stand structure as biodiversity indicator[J]. Forest Ecology and Management, 2014, 330: 82-93. doi: 10.1016/j.foreco.2014.07.007 [33] MOUGI A, KONDOH M. Diversity of interaction types and ecological community stability[J]. Science, 2012, 337(6092): 349-351. doi: 10.1126/science.1220529 [34] IVES A R, CARPENTER S R. Stability and diversity of ecosystems[J]. Science, 2007, 317(5834): 58-62. doi: 10.1126/science.1133258 [35] 田起隆, 张晓萍, 王妙倩, 等. 植被演替下植物组成多样性和群落稳定性与土壤团聚体的关系[J]. 中国环境科学, 2022, 42(12):5885-5895. [36] 冯 凌, 喻理飞, 王 阳, 等. 喀斯特地区植被不同恢复阶段功能冗余和功能多样性对群落稳定性的影响[J]. 生态环境学报, 2022, 31(4):670-678. [37] 汪永文, 王 力, 王丽丽, 等. 马尾松混交林林下植被结构及生物量特征研究[J]. 安徽农业大学学报, 2010, 37(2):312-316. [38] VALENCIA E, De BELLO F, GALLAND T, et al. Synchrony matters more than species richness in plant community stability at a global scale[J]. Proceedings of the National Academy of Sciences, 2020, 117(39): 24345-24351. doi: 10.1073/pnas.1920405117 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 618
- HTML全文浏览量: 179
- PDF下载量: 6
- 被引次数: 0





