




-
土壤N、P养分状况影响植物的养分代谢和循环,使其叶、根等主要功能器官具有不同的N、P含量和特定的化学计量特征[1-3],进而影响植物的光合作用及养分吸收利用效率,调控植物的生长发育[4-5]。植物体内N、P元素含量、分布及化学计量特征体现了其对环境的响应和适应性,对植物的生长发育及生产力有显著的影响[6]。顾大形等[3]研究表明,四季竹(Oligostachyum lubricum (Wen) Keng f.)叶片N/P是影响叶片叶绿素含量的主要因素,进而影响和调控四季竹叶片光合作用。Liu等[7]对我国热带森林内不同个体植物研究发现,植物个体大小与其叶片N、P养分特征相关。张蕾蕾等[8]研究发现,刨花楠(Machilus pauhoi Kanchira)不同个体叶片N、P含量及其化学计量特征与胸径呈显著相关。
我国南方森林立地普遍为酸性红壤,土壤有效P严重匮乏成为限制森林生产力的主要因素之一[9-11]。近年来,人类活动加剧了全球大气N沉降的持续增加,导致森林土壤有效N含量增加及N/P发生改变,影响了植物的生长发育和生产力[12-13]。植物体功能器官N、P元素含量及其化学计量特征是在一定生境条件下植物与环境之间相互作用的结果,能够控制植物的生长状况和生长速率[14-15]。因此,研究叶片N、P含量及其化学计量特征对了解森林生态系统N、P需求及森林生产力具有重要的理论意义。
马尾松(Pinus massoniana Lamb.)是我国南方重要的用材和采脂树种,也是荒山造林的先锋树种,分布广泛,速生丰产,经济价值高。由于南方亚热带多为贫瘠的酸性红壤,缺P成为马尾松生长的主要限制因子。当前研究已揭示马尾松主要通过根构型的适应性变化、根系有机酸和APase分泌的增加等方式来适应低P胁迫[16-18]。然而,已有研究较少考虑马尾松响应、适应土壤养分状况下,功能器官养分含量及化学计量特征,以及其对植物生长发育的影响。鉴于此,本文选用30个马尾松二代无性系,对其树高和胸径等生长性状、成熟和当年生针叶N、P含量及其化学计量比进行调查和分析,以揭示马尾松二代无性系生长与针叶N/ P化学计量比的相关性,为深入研究马尾松响应和适应低P立地及大气N沉降,调节体内养分的含量、分布和利用机制,预测南方低P森林立地人工林的生产力提供重要参考。
-
本试验地点为浙江省淳安县姥山林场国家马尾松种质资源库(119。03 E,29。37 N),海拔200~250 m,年平均气温17℃,≥10℃的年积温为5 410℃,年降水量1 430 mm,年日照时数1 951 h。用以研究的30个马尾松二代无性系(表 1)来自于2002年嫁接建立的二代育种群体,每个无性系嫁接保存4~6株,株行距为6 m × 8 m。无性系所处立地条件一致,东南坡向,坡度25°,土层厚度60 cm左右,土壤有效P含量约为(10.36±3.35)mg·kg-1,马尾松二代无性系植株生长良好。
表 1 马尾松二代无性系的遗传背景
Table 1. The genetic background of P. massoniana second generation clones
无性系编号
Clone number遗传背景
Genetic background无性系编号
Clone number遗传背景
Genetic background无性系编号
Clone number遗传背景
Genetic backgroundC2 5476×1127 C14 3201×1123 C57 1217×1121 C3 5476×5906 C15 6101×6101 C64 开化1 C4 5131×5906 C43 种源1107 C65 开化5 C5 5910×1134 C44 种源1123 C69 9002×6623 C7 5158×1134 C45 种源1133 C71 开化小油1 C8 6617×1134 C46 种源1145 C75 开化茹坞12 C10 6101×5907 C48 种源1321 C81 三门1 C11 6313×1130 C49 种源6613 C82 东阳3 C12 8101×1128 C55 5907×1003 C97 缙云4 C13 7750×1126 C56 3412×5907 C101 丽水3 马尾松二代无性系11年生时,于当年8月中旬(马尾松生长旺季),在每个二代无性系中随机选择生长正常的3株,在其树冠的东、西、南、北4个方向,选取上、中、下不同部位的枝条,分别采摘当年生(颜色为嫩绿色)和上一年成熟的针叶(颜色为深绿色)各约15 g,4个方向的针叶分别混合后,采用四分法取样,并装入自封袋编号备用。同时,调查各无性系分株的树高、胸径和冠幅等指标。
-
将采集的当年生针叶和成熟针叶,于烘箱中105℃杀青30 min,60℃烘干至恒质量,以获得其干物质量。用H2SO4-H2O2消煮后测定当年生针叶和成熟针叶的含N量和含P量,含N量利用凯氏定N法测定,含P量用钼锑抗比色法测定。成熟针叶或当年生针叶N/P=针叶N含量/针叶P含量。无性系重复力估算方法参照文献[19-20]。利用SAS软件6.2的ANOVA程序对二代无性系各生长性状、针叶N和P含量及N/P等进行方差分析,CORR程序对马尾松二代无性系生长性状与当年生针叶和成熟针叶N、P含量及N/P进行相关分析,CLUSTER程序对马尾松二代无性系主要生长性状与针叶养分含量进行Q型聚类分析。
-
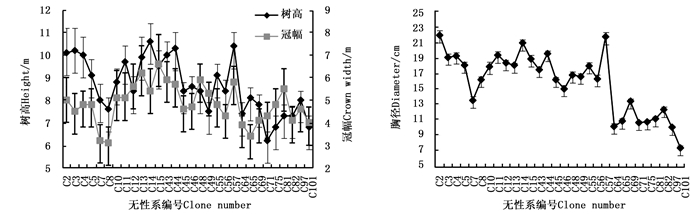
由表 2可知: 树高、胸径和冠幅等生长性状在30个马尾松二代无性系间存在显著差异。二代无性系树高变化范围是6.2~10.6 m,平均树高为8.60 m,其中,树高生长量较大的马尾松无性系有C14、C57、C44、C3、C2、C4、C43、C13、C11和C15;胸径变化范围是7.4~22.1 m,平均胸径为16.22 cm,其中,胸径较大的无性系有C2、C57、C14、C44、C11、C4、C3、C13、C12和C5;冠幅变化范围是3.1~6.6 m,平均冠幅为4.81 m,其中,冠幅较大的马尾松无性系有C15、C13、C48、C43、C57、C44、C12、C81、C14和C49(图 1)。比较分析表明:大部分树高生长量较大的二代无性系,其胸径也较大,其中,无性系C13、C14、C57和C44冠幅也较大。另外,马尾松二代无性系树高、胸径和冠幅等重复力很高,均达到0.95以上。
表 2 马尾松二代无性系主要生长性状的差异
Table 2. The difference of main growth traits of P. massoniana second generation clones
性状
Trait树高
Height/m胸径
Diameter/cm冠幅
Crown width /m平均值Average 8.60±0.31 16.22±0.53 4.81±0.20 显著性水平Probability <0.0001 <0.0001 <0.0001 无性系重复力
Clone repeatability0.96 0.98 0.97 
图 1 马尾松二代无性系树高、胸径和冠幅
Figure 1. Tree height, diameters and crown width of P. massoniana second generation clones
-
方差分析表明:成熟针叶和当年生针叶的N、P含量及N/P在马尾松二代无性系间差异均达极显著(表 3)。二代无性系成熟针叶P含量变化范围是0.62~1.35 mg·g-1,平均值为1.11 mg·g-1,含量较高的无性系包括C46、C7、C2、C8、C64、C5、C55、C65、C45和C11(图 2);当年生针叶P含量变化范围是0.58~0.85 mg·g-1,平均值为0.70 mg·g-1,含量较高的无性系有C65、C69、C13、C14、C81、C64、C82、C45、C49和C55(图 3)。与马尾松二代无性系当年生针叶相比,其成熟针叶P含量较高,是当年生针叶的1.62倍,但N含量差异不明显。成熟针叶N含量变化范围是9.25~16.66 mg·g-1,平均值为14.03 mg·g-1,含量较高的无性系包括C4、C13、C11、C65、C43、C56、C3、C57、C12和C14(图 2);当年生针叶N含量变化范围是10.11~17.82 mg·g-1,平均值为14.42 mg·g-1,含量较高的无性系有C43、C65、C56、C69、C2、C45、C4、C44、C49和C15(图 3)。可见,马尾松当年生针叶和成熟针叶N含量相差不大,说明植株生长可能并不缺乏N素。由于P素较多地分配于成熟针叶内,而当年针叶中的P含量较低,导致当年生针叶的N/P较大。马尾松二代无性系当年生针叶的N/P是成熟针叶的1.6倍(表 3)。成熟针叶N/P的变化范围是8.30~22.43,平均值为13.04;当年生针叶N/P的变化范围是13.70~26.61,平均值为20.91。
表 3 马尾松二代无性系成熟针叶和当年生针叶N、P含量和N/P
Table 3. The N, P content and N/P ratios of mature needle and current year needle of P. massoniana second generation clones
处理Treatment N含量N content /(mg·g-1) P含量P content /(mg·g-1) N/P ratios 成熟针叶Mature needle 14.03±2.15 1.11±0.17 13.04±3.22 显著性水平Probability <0.000 1 <0.000 1 <0.000 1 当年生针叶Current year needle 14.42±2.04 0.70±0.07 20.91±3.41 显著性水平Probability <0.000 1 <0.000 1 <0.000 1 成熟针叶/当年生针叶
Mature needle/Current year needle0.97 1.62 0.63 平均值Average 14.23±2.01 0.91±0.10 15.72±2.84 
图 2 马尾松二代无性系成熟针叶N、P含量和N/P
Figure 2. The N, P content and N/P ratios of mature needle and current year needle of P. massoniana second generation clones

图 3 马尾松二代无性系当年生针叶N、P含量和N/P
Figure 3. The N, P content and N/P ratios of current year needle of P. massoniana second generation clones
-
分析30个马尾松二代无性系生长性状与针叶N、P含量间的相关性发现:无性系的树高和胸径不管是与成熟针叶,还是与当年生针叶的N含量、N/P均呈显著或极显著正相关;无性系树高和胸径与成熟针叶N含量的相关系数分别为0.541 1和0.607 5,与N/P的相关系数分别为0.608 6和0.542 8,与当年生针叶N含量的相关系数分别为0.477 4和0.528 7,与N/P的相关系数分别为0.515 9和0.679 2(表 4)。此外,无性系冠幅与成熟针叶N/P呈显著正相关,相关系数达0.541 9。表明成熟针叶和当年生针叶N含量及N/P比能够影响马尾松植株树高和胸径的生长,而冠幅与成熟针叶N/P相关性较大; 然而,马尾松二代无性系生长与针叶P含量相关性较小。
表 4 马尾松二代无性系成熟针叶和当年生针叶养分含量与主要生长性状的相关系数
Table 4. Correlation coefficients between nutrient content of mature needle and current year needle and main growth traits of P. massoniana second generation clones
性状
Trait成熟针叶Mature needle 当年生针叶Current year needle N含量
N contentP含量
P contentN/P
ratiosN含量
N contentP含量
P contentN/P
ratios树高Height 0.541 1* -0.273 9 0.608 6** 0.477 4* -0.166 4 0.515 9* 胸径Diameter 0.607 5** -0.132 5 0.542 8* 0.528 7* -0.342 7 0.679 2** 冠幅Crown width 0.162 1 -0.540 7* 0.541 9* 0.139 7 -0.207 2 0.280 0 注:*P<0.05; **P<0.01。Note: *P<0.05; **P<0.01. -
为充分了解马尾松二代无性系主要生长性状与针叶养分含量的关系,以马尾松树高、胸径和成熟针叶及当年生针叶N含量、N/P为指标,对30个马尾松二代无性系进行Q型聚类分析。图 4表明:在欧氏距离为0.173处,可将30个无性系划分为4大类。第1类包括C13和C14两个无性系,其树高和胸径等生长表现优良,成熟针叶N含量、N/P较高,而当年生针叶N含量、N/P相对较低;2个无性系成熟针叶的N/P分别为22.43和20.24,当年生针叶的N/P分别为18.38和20.23;成熟针叶N/P远高于30个无性系平均值,而当年生针叶N/P低于其平均值。第2类包括C2、C3、C4、C10、C11、C44、C55和C57共8个无性系,其树高、胸径等生长表现较好,成熟针叶和当年生针叶N含量、N/P在30个无性系中相对居中;除C2成熟针叶和当年生针叶N/P分别为10.68和24.28,相差较大外,其它7个无性系成熟针叶和当年生针叶N/P相差较小,其变化范围分别为11.98~16.53和19.77~23.61。第3类包括C5、C7、C8、C12、C15、C43、C45、C46、C48、C49、C56、C64、C65、C69和C71共计15个无性系,占试验无性系总数的一半;这些无性系树高、胸径等生长表现一般,成熟针叶和当年生针叶N/P相差较大;大部分无性系成熟针叶N/P较低,而当年生针叶N/P相对较高。第4类包括C75、C81、C82、C97和C101共5个无性系,其树高、胸径等生长表现较差,成熟针叶和当年生针叶N/P均较低。

图 4 基于主要生长性状、针叶N含量和N/P的马尾松二代无性系聚类分析
Figure 4. Cluster analysis of P. massoniana second generation clones based on main growth traits, N content and N/P ratio of needles
-
本研究结果表明:30个马尾松二代无性系11年生时树高、胸径和冠幅等存在显著差异,且各生长性状无性系重复力较高,均达0.95以上。说明对马尾松二代无性系生长开展进一步的评价和选择是必须的,也是可行的。
在南方亚热带贫P立地条件下,马尾松二代无性系成熟针叶和当年生针叶N、P含量及N/P在不同无性系间差异均显著。马尾松二代无性系成熟针叶N/P平均值为13.04,当年生针叶N/P平均值为20.91,整株针叶N/P的平均值为15.72。与中国不同温度带753种陆生植物叶片N/P的几何平均值l4.4[21]相比,本研究结果偏高。这可能与叶片N、P含量在一定程度上可以反映立地的养分有效性及供应水平有关[22]。我国南方森林土壤有效P严重匮乏,N素含量一般较为充足,导致马尾松二代无性系针叶N/P偏高。与马尾松二代无性系当年生针叶相比,其成熟针叶P含量较高,而N含量差异不明显,因此,当年生针叶的N/P较大。说明P素对马尾松二代无性系当年生针叶的限制性更强,其更容易因缺乏P素而发育不良,这与以往研究结果相似[23]。
-
有研究表明,叶片N、P元素组成的差异与叶片主要生理生态功能相联系,能够影响叶片叶绿素的含量[3],这在一定程度上反映植物体N、P养分的动态平衡,影响植物的生长量和生产力[24-25]。在自然状况下,植物的成熟功能叶和当年生新叶的生长速度和发育程度不同,从而对营养元素的需求也不同。二者N、P含量及其化学计量特征可以作为判断植物生长状况的指标[23]。本研究表明,马尾松二代无性系树高和胸径与成熟针叶和当年生针叶N含量、N/P均呈显著或极显著正相关,冠幅仅与成熟针叶N/P呈显著正相关。这表明成熟针叶和当年生针叶N含量及N/P可能通过调控叶的生长发育、光合作用及养分代谢等,显著影响马尾松植株树高和胸径的生长。与当年生针叶相比,功能叶成熟针叶的生长发育对冠幅影响较大。张蕾蕾等[8]研究表明,刨花楠不同个体叶片N、P含量及其化学计量特征与胸径呈显著线性相关,与本研究结果一致。
为充分了解马尾松二代无性系主要生长性状与针叶养分含量的关系,利用聚类分析将30个11年生马尾松二代无性系分为4类。与第3类无性系相比,第1类和第2类无性系树高和胸径等生长表现优良,成熟针叶N/P较高,而当年生针叶N/P相对较低,尤其是第1类无性系。以往的研究表明,N/P在一定阈值范围内,N、P同时限制或都不限制生长,当N/P低于这一阈值时,植物生长受N限制,植物对N素敏感,增加N素可促进生长,而当N/P高于阈值时,生长受P限制,施加P素可促进生长[28]。本研究结果可能的原因是在缺乏P素环境下,第1类无性系优先将P素分配给生长中心,满足新叶的生长需要[26]; 而当年生针叶相对较低的N/P,增加了马尾松对N素的敏感性,促进了N素的吸收,从而增加了无性系生长量。这与以往的研究一致[27]。与第1类和第2类无性系相比,第4类无性系树高和胸径等生长量较小,成熟针叶和当年生针叶N/P均较低。
-
研究表明,在养分相对贫瘠的立地上,植物能够以其独特的养分再吸收、体内养分运转和再分配等策略来提高其养分利用效率,改善其生长状况进而适应贫瘠环境。在贫P立地下,马尾松二代亲本无性系C13(7750×1126)和C14(3201×1123)各生长指标均较高,可能是通过调整当年生针叶和成熟针叶的养分分配,增加了当年生针叶对限制其生长的P素的再利用和再吸收,从而降低了当年生针叶的N/P,促进了N素的吸收。植物体内P的有效循环利用被普遍认为是植物P高效的生理机制之一[29],养分分配对策体现了植物对不同环境压力作出的响应[30]; 然而,植物体内养分有效利用及分配对环境的响应是一个漫长且缓慢的过程,要确切了解马尾松针叶养分含量及其化学计量特征与无性系生长的关系,还有待于开展长期的定位研究。
马尾松二代无性系生长和针叶N/P化学计量特征
Plant Growth and Needles N and P Stoichiometric Characteristics of Second-generation Pinus massoniana Clones
-
摘要:
目的 在土壤有效P匮乏的立地条件下,揭示针叶N、P含量及其化学计量比与马尾松生长性状的相关性。 方法 以马尾松二代育种群体内30个无性系为试验对象,野外调查11年生时马尾松二代无性系树高、胸径等生长性状与针叶N、P含量及其化学计量比的相关性。 结果 结果表明:(1)各生长性状在马尾松二代无性系间差异显著,且无性系重复力较高,均达到0.95以上。马尾松成熟针叶和当年生针叶N、P含量及N/P在二代无性系间差异均达显著水平;(2)相关分析表明,马尾松二代无性系树高、胸径与成熟针叶和当年生针叶N含量、N/P均呈显著或极显著正相关,与成熟针叶N/P的相关系数分别为0.608 6和0.542 8,与当年生针叶N/P的相关系数分别为0.515 9和0.679 2;(3)基于主要生长性状、针叶N和P含量及其化学计量比,可将30个马尾松二代无性系聚为4类:类群1无性系树高、胸径等生长表现优良,成熟针叶N/P较高,而当年生针叶N/P相对较低;类群2无性系树高、胸径等生长表现较好,成熟针叶和当年生针叶N/P值在30个无性系中居中间水平;类群3占试验无性系总数的一半,树高、胸径等生长表现一般,成熟针叶与当年生针叶N/P相差较大,大部分无性系成熟针叶N/P较低,而当年生针叶N/P相对较高;类群4树高、胸径等生长表现较差,成熟针叶和当年生针叶N/P均较低。 结论 在贫P立地下,马尾松二代亲本无性系C13(7750×1126)和C14(3201×1123)各生长指标均较高,可能是通过调整当年生针叶和成熟针叶的养分分配,增加了当年生针叶对限制其生长的P素的再利用和再吸收,促进了N素的吸收,调整了当年生针叶和成熟针叶的N/P,以形成特定的养分分配机制。 Abstract:Objective The purpose of this study is to reveal the correlation between stoichiometric characteristics of N, P and the growth traits of Pinus massoniana on P deficiency site. Method Taking 11-year-old P. massoniana clones in second-generation breeding population as test materials, the major growth traits, including height, DBH, and its correlation with N, P stoichiometric characteristics of needles, were studied through field investigation. Result (1) Significant differences were observed in traits among clones, and high clonal repeatability was detected. There were significant differences among clones in N, P content and N/P of mature and current year needle; (2) The major growth traits of clones had positive correlations with N content and N/P of mature and current year needle; (3) Based on major growth traits and N, P stoichiometric characteristics of needles, the 30 clones tested were clustered into 4 groups. Group 1 exhibited superior growth traits, with higher N/P of mature needle, but relatively lower N/P of current year needle. Group 2 exhibited good growth traits, with medium N/P of mature and current year needle. Group 3 had the ordinary growth traits, but this group accounted for half of the tested clones. The N/P of mature needle had larger difference with current year needle. Group 4 exhibited poor growth traits, while N/P of mature and current year needle were lower. Conclusion Two clones belonging to group 1 exhibited perfect growth traits, which might by adjusting N, P content and N/P of mature and current year needle to form a particular nutrient allocation mechanism. -
Key words:
- Pinus massoniana
- / second-generation clones
- / growth
- / needles
- / N/P
-
图 1 马尾松二代无性系树高、胸径和冠幅
Figure 1. Tree height, diameters and crown width of P. massoniana second generation clones
图 2 马尾松二代无性系成熟针叶N、P含量和N/P
Figure 2. The N, P content and N/P ratios of mature needle and current year needle of P. massoniana second generation clones
图 3 马尾松二代无性系当年生针叶N、P含量和N/P
Figure 3. The N, P content and N/P ratios of current year needle of P. massoniana second generation clones
图 4 基于主要生长性状、针叶N含量和N/P的马尾松二代无性系聚类分析
Figure 4. Cluster analysis of P. massoniana second generation clones based on main growth traits, N content and N/P ratio of needles
表 1 马尾松二代无性系的遗传背景
Table 1. The genetic background of P. massoniana second generation clones
无性系编号
Clone number遗传背景
Genetic background无性系编号
Clone number遗传背景
Genetic background无性系编号
Clone number遗传背景
Genetic backgroundC2 5476×1127 C14 3201×1123 C57 1217×1121 C3 5476×5906 C15 6101×6101 C64 开化1 C4 5131×5906 C43 种源1107 C65 开化5 C5 5910×1134 C44 种源1123 C69 9002×6623 C7 5158×1134 C45 种源1133 C71 开化小油1 C8 6617×1134 C46 种源1145 C75 开化茹坞12 C10 6101×5907 C48 种源1321 C81 三门1 C11 6313×1130 C49 种源6613 C82 东阳3 C12 8101×1128 C55 5907×1003 C97 缙云4 C13 7750×1126 C56 3412×5907 C101 丽水3  下载: 导出CSV
下载: 导出CSV
表 2 马尾松二代无性系主要生长性状的差异
Table 2. The difference of main growth traits of P. massoniana second generation clones
性状
Trait树高
Height/m胸径
Diameter/cm冠幅
Crown width /m平均值Average 8.60±0.31 16.22±0.53 4.81±0.20 显著性水平Probability <0.0001 <0.0001 <0.0001 无性系重复力
Clone repeatability0.96 0.98 0.97
下载: 导出CSV
表 3 马尾松二代无性系成熟针叶和当年生针叶N、P含量和N/P
Table 3. The N, P content and N/P ratios of mature needle and current year needle of P. massoniana second generation clones
处理Treatment N含量N content /(mg·g-1) P含量P content /(mg·g-1) N/P ratios 成熟针叶Mature needle 14.03±2.15 1.11±0.17 13.04±3.22 显著性水平Probability <0.000 1 <0.000 1 <0.000 1 当年生针叶Current year needle 14.42±2.04 0.70±0.07 20.91±3.41 显著性水平Probability <0.000 1 <0.000 1 <0.000 1 成熟针叶/当年生针叶
Mature needle/Current year needle0.97 1.62 0.63 平均值Average 14.23±2.01 0.91±0.10 15.72±2.84
下载: 导出CSV
表 4 马尾松二代无性系成熟针叶和当年生针叶养分含量与主要生长性状的相关系数
Table 4. Correlation coefficients between nutrient content of mature needle and current year needle and main growth traits of P. massoniana second generation clones
性状
Trait成熟针叶Mature needle 当年生针叶Current year needle N含量
N contentP含量
P contentN/P
ratiosN含量
N contentP含量
P contentN/P
ratios树高Height 0.541 1* -0.273 9 0.608 6** 0.477 4* -0.166 4 0.515 9* 胸径Diameter 0.607 5** -0.132 5 0.542 8* 0.528 7* -0.342 7 0.679 2** 冠幅Crown width 0.162 1 -0.540 7* 0.541 9* 0.139 7 -0.207 2 0.280 0 注:*P<0.05; **P<0.01。Note: *P<0.05; **P<0.01.
下载: 导出CSV
-
[1] He J S, Fang J Y, Wang Z H, et al. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 2006, 149(1): 115-122. doi: 10.1007/s00442-006-0425-0 [2] 杨阔, 黄建辉, 董丹, 等. 青藏高原草地植物群落冠层叶片氮磷化学计量学分析[J]. 植物生态学报, 2010, 34(1): 17-22. doi: 10.3773/j.issn.1005-264x.2010.01.004 [3] 顾大形, 陈双林, 黄玉清, 等. 土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响[J]. 植物生态学报, 2011, 35 (12): 1219-1225. [4] 邢雪荣, 韩兴国, 陈灵芝. 植物养分利用效率研究综述[J]. 应用生态学报, 2000, 11(5): 785-790. doi: 10.3321/j.issn:1001-9332.2000.05.033 [5] 陈伏生, 胡小飞, 葛刚. 城市地被植物麦冬叶片氮磷化学计量比和养分再吸收效率[J]. 草业学报, 2007, 16(4): 47-54. doi: 10.3321/j.issn:1004-5759.2007.04.008 [6] 王振南, 杨惠敏. 植物碳氮磷生态化学计量对非生物因子的响应[J]. 草业科学, 2013, 30(6): 927-934. [7] Liu F D, Yang W J, Wang Z S, et al. Plant size effects on the relationships among specific leaf area, leaf nutrient content, and photosynthetic capacity in tropical woody species[J]. Acta Oecologica, 2010, 36(2): 149-159. doi: 10.1016/j.actao.2009.11.004 [8] 张蕾蕾, 钟全林, 程栋梁, 等. 刨花楠叶片碳氮磷化学计量比与个体大小的关系[J]. 应用生态学报, 2015, 26(7): 1928-1934. [9] Vance C P. Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resources[J]. Plant Physiology, 2001, 127(2): 390-397. doi: 10.1104/pp.010331 [10] 严小龙, 廖红. 根系生物学: 原理与应用[M]. 北京: 科学出版社, 2007: 128. [11] 张福锁, 崔振岭, 王激清, 等. 中国土壤和植物养分管理现状与改进策略[J]. 植物学通报, 2007, 24(6): 687-694. doi: 10.3969/j.issn.1674-3466.2007.06.001 [12] 阎恩荣, 王希华, 郭明, 等. 浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C∶N∶P化学计量特征[J]. 植物生态学报, 2010, 34(1): 48-57. doi: 10.3773/j.issn.1005-264x.2010.01.008 [13] 宾振钧, 王静静, 张文鹏, 等. 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响[J]. 植物生态学报, 2014, 38(3): 231-237. [14] Aerts R, Chapin F S. The mineral nutrition of wild plants revisited: a re-evaluation of process and patterns[J]. Advances in Ecological Research, 2000, 30(1): 1-67. [15] Gusewell S. N∶P ratios in terrestrial plants: Variation and functional significance[J]. New Phytologist, 2004, 164(2): 243-266. doi: 10.1111/j.1469-8137.2004.01192.x [16] 周志春, 谢钰容, 金国庆, 等. 马尾松种源对磷肥的遗传反应及根际土壤营养差异[J]. 林业科学, 2003, 39(6): 62-67. doi: 10.3321/j.issn:1001-7488.2003.06.010 [17] 谢钰容, 周志春, 廖国华, 等. 低磷胁迫下马尾松种源酸性磷酸酶活性差异[J]. 林业科学, 2005, 41(3): 58-62. doi: 10.3321/j.issn:1001-7488.2005.03.010 [18] Zhang Y, Zhou Z C, Yang Q. Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana under heterogeneous and homogeneous low phosphorus conditions[J]. Plant and Soil, 2013, 364(1): 93-104. [19] 续九如. 林木数量遗传学[M]. 北京: 高等教育出版社, 2006: 26-33. [20] 孔繁玲. 植物数量遗传学[M]. 北京: 中国农业大学出版社, 2006: 224-249. [21] Han W X, Fang J Y, Guo D, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2): 377-385. doi: 10.1111/j.1469-8137.2005.01530.x [22] 程滨, 赵永军, 张文广, 等. 生态化学计量学研究进展[J]. 生态学报, 2010, 30(6): 1628-1637. [23] 高三平, 李俊祥, 徐明策, 等. 天童常绿阔叶林不同演替阶段常见种叶片N、P化学计量学特征[J]. 生态学报, 2007, 27(3): 947-952. doi: 10.3321/j.issn:1000-0933.2007.03.015 [24] Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle[J]. Biogeochemistry, 2000, 48(1): 7-20. doi: 10.1023/A:1006247623877 [25] 陈琳, 曾杰, 徐大平, 等. 氮素营养对西南桦幼苗生长及叶片养分状况的影响[J]. 林业科学, 2010, 46(5): 35-40. [26] Schreeg L A, Santiago L S, Wright S J, et al. Stem, root, and older leaf N∶P ratios are more responsive indicators of soil nutrient availability than new foliage[J]. Ecology, 2014, 95(8): 2062-2068. doi: 10.1890/13-1671.1 [27] 庞丽, 张一, 周志春, 等. 模拟氮沉降对低磷胁迫下马尾松不同家系根系分泌和磷效率的影响[J]. 植物生态学报, 2014, 38(1): 27-35. [28] Fujita Y, Robroek B J M, Ruiter P C D, et al. Increased N affects P uptake of eight grassland species: the role of root surface phosphatase activity[J]. Oikos, 2010, 119(10): 1665-1673. doi: 10.1111/j.1600-0706.2010.18427.x [29] 樊明寿, 陈刚, 孙国荣. 低磷胁迫下玉米根中磷的运转与再利用[J]. 作物学报, 2006, 32(6): 946-948. doi: 10.3321/j.issn:0496-3490.2006.06.028 [30] 何维明, 张新时. 沙地柏对毛乌素沙地3种生境中养分资源的反应[J]. 林业科学, 2002, 38(5): 1-6. doi: 10.3321/j.issn:1001-7488.2002.05.001 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3850
- HTML全文浏览量: 790
- PDF下载量: 498
- 被引次数: 0
