

-
树木根系在森林生态系统物质和能量循环过程中具有十分重要作用,养分和有机质通过根系输入到土壤中可以维持土壤肥力和固定碳,生态系统中根系生物量占有很大的比例[1]。细根通过较快的周转率为土壤提供丰富的养分和有机质[2],细根通常是指直径小于2 mm的根系,具有巨大的吸收表面积,是树木吸收水分和养分的主要器官[3-4],多数具有菌根侵染,是植物生长最活跃和最敏感的部分[5],由于细根较快的周转率而成为土壤碳输入的主要来源[6],在森林生态系统碳平衡和养分循环中有重要作用。研究发现, 净初级生产力分配到地下的常常高于地上部分,每年通过细根输入到土壤中的碳和养分高于叶片,林龄是森林净初级生产力、碳储量和碳固定的重要影响因素,随着林龄的增长,净初级生产力随之增长,达到顶峰后下降[7];同时,由于不同林龄发展阶段群落物种组成及养分条件有较大的不同,影响到细根的生产和周转[1-2],因而, 林龄对森林细根生物量有很大影响[6]。研究表明,随着林龄的增大,阔叶林细根生物量在增加,而针叶林细根生物量在减少,通常在林冠闭合前达到顶点,然后逐渐减少[8]。
根系的垂直分布很难被衡量,普遍认为,在林分的所有林龄阶段中根系生物量随土壤深度的增加而减少,但在不同林分发展过程中,根系在不同土层中的变化依然十分重要,中幼林具有更快的生物量积累的能力,因而,比老龄林可以更快的积累养分,这是因为中幼林的根可以从较深的矿质土壤中更好吸收养分,老龄林的根相对更多分布在土壤表层[4, 9]。粗根生物量一般随着树干生长而增加,而细根以树干为中心可以扩散到更远的距离,因而,森林细根生物量的分布可以很好的反映出土壤养分的分布[10]。
思茅松(Pinus kesiya Royle ex Gord. var. langbianensis (A. Chev.) Gaussen)作为喜阳速生树种,是云南省南亚热带重要的造林树种之一,广泛用于该区域的退耕还林工程中。近年来,思茅松人工林的从1987年的2.86%已发展到2007年的10.57%,成为重要的森林碳储量的来源[11-12]。据估计,思茅松林每年的细根生长量为734~1 054 g·m-2,远高于温带针叶林[1]。思茅松林龄的增大不但改变地上部分与地下部分生物量分布,同时也影响地下部分不同根系的性状与生物量分配[13],而目前对于云南省南亚热带思茅松细根生物量的研究还相对较少。本研究以云南省南亚热带区域的思茅松人工林为研究对象,利用根钻法研究思茅松根系的大小分布,探讨分析思茅松人工林中不同物种的根系在不同林龄阶段的大小分布格局,分析林龄和土壤深度对根系生物量的交互作用,为了解南亚热带地区思茅松人工林的地下碳分配提供基础数据,同时也为进一步分析思茅松人工林的细根在森林生态系统碳循环过程中的作用奠定基础。
-
研究区位于云南省普洱市思茅区(100°53′~101°2′ E,22°41′~ 22°49′ N),海拔为1 270~1 485 m,处于滇南热带与南亚热带的过渡位置,属南亚热带高原季风气候,受到较明显的印度洋西南季风暖湿气流的影响,形成干湿季分明的气候特征,雨季多集中在5月至10月,年降水量为1 547.6 mm,年平均蒸发量1 590 mm,相对湿度为82 %[11]。赤红壤为当地森林的地带性土壤。该区域自然植被主要有暖热性针叶林和季风常绿阔叶林两种类型,其中,暖热性针叶林主要以思茅松林为主。思茅松作为云南主要的用材和采脂优良乡土树种之一,有着悠久的采伐利用历史,目前在思茅区分布有大面积的思茅松人工林,大部分在思茅松天然林的采伐迹地上人工造林而成,其中,从1998年分2个阶段,前期主要使用的是采伐迹地撒播造林,后期主要采用营养袋人工育苗造林[12]。思茅松人工林森林内组成物种除思茅松外,其它物种还有红木荷(Schima wallichii(DC.) Choisy)、短刺栲(Castanopsis echidnocarpa J. D. Hooker et Thomson ex Miquel)、刺栲(C. hystrix Miq.)、小果栲(C. fleuryi Hick. et A. Camus)、茶梨(Anneslea fragrans Wall.)、野漆(Toxicodendron succedaneum(L.) O. Kuntze)、红皮水锦树(Wendlandia tinctoria(Roxb.) DC. subsp. intermedia(How) W. C. Chen)和毛银柴(Aporusa villosa(Lindl.) Baill.)等乔木物种以及多花野牡丹(Melastoma affine D. Don)和黑面神(Breynia fruticosa (Linn.) Hook. f.)等灌木树种。此外,草本植物在群落内有较大的盖度,常见物种有紫茎泽兰(Eupatorium adenophora Spreng)、毛果珍珠茅(Scleria herbecarpa Nees)、多毛姜(Zingiber densissimum S. Q. Tong et Y. M. Xia)、云南草蔻(Alpinia blepharocalyx K. Schum.)和狗脊(Woodwardia japonica(L. f.) Sm.)等物种。
-
在普洱市思茅区万掌山林场、清水河林区、大寨及梅子湖水库周边设置试验林,选择5、8、15、25、36年生等5个不同林龄阶段的思茅松人工林,其中,5、8、15年生初始造林密度为2 500株·hm-2,而后2个林龄则采取撒播更新。2014年10月选择海拔较一致,生长况状及经营管理水平相似的地段进行样地选择及取样调查。每个林龄选取3个样地,共设置15个样地。样地大小为20 m×30 m,每个样地之间至少有500 m的过渡带,对每个样地胸径≥1 cm的乔木进行每木检尺,调查其胸径及树高等林分因子。其林分特征见表 1。
表 1 思茅松人工林林分特征
Table 1. Stand traits in the Pinus kesiya var. langbianensis plantations
林龄
Stand age/a地点
Location纬度(N)
Latitude纬度(E)
Longitude海拔
Altitude/m胸径
DBH/cm树高
Height/m胸高断面积
Basal area at DBH/cm2林分密度
Stand density/(trees·hm-2)5 万掌山林场 22°49′ 100°53′ 1 410 6.46±0.36c 4.63±0.37d 5.72±0.99d 1 600±150a 8 万掌山林场 22°49′ 100°54′ 1 485 12.50±0.53b 9.36±0.82c 15.3±1.24c 1 200±156b 15 清水河林区 22°41′ 100°57′ 1 270 13.33±1.38b 10.94±0.66b 23.56±4.50b 1 130±5b 25 大寨 22°45′ 101°2′ 1 415 19.84±1.51a 16.77±0.21a 26.21±3.42a 817±226b 36 梅子湖水库 22°44′ 100°58′ 1 370 12.04±1.73b 8.63±0.72c 30.56±9.60a 1 989±377a 注:同行不同字母表示差异显著P < 0.05。
Notes:the different letters with in the same row mean significant difference at the 0.05 level. -
根系生物量采用根钻进行野外取样,根钻规格为内直径8.5 cm、高10 cm以及有锋利边缘,以充分进行根系生物量取样。参考长白落叶松(Larix olgensis Henry)人工林细根生物量的研究方法[14],于2014年10月以思茅松人工林株行距的中间位置为起点,呈Z字形在每个林分内随机取样,每个样地林分内取10个土样。每个土样取样点按0~10、10~20、20~30 cm等3个垂直土层进行取样,用于根系垂直分布特征调查。野外所取土壤样品挑出石砾后装入保鲜袋内,做好标记储存在4℃的冷藏箱中,带回实验室放入-3℃的冰箱中,对不同林龄类型的不同土层深度的样品进行统一处理与分析。
将野外采集的土样在实验室内用清水充分浸泡5 h,用筛孔为20目的筛网用水反复淘洗,进行根的挑拣,将洗净后的根放入白色的盘中,注入少量水分,根据思茅松根系特征(思茅松根系呈暗紫红色,根表皮有皲裂,不易折断),用镊子将思茅松和其它植物的根系进行归类整理。将根系分为以下3个类型:(1)细根,根径≤2 mm;(2)粗根,根径>2 mm;(3)死根,根据细根的颜色、外形、弹性、根皮与中柱分离的难易程度并结合漂浮法进行活根和死根的区分;同时根据林分物种的组成,将根系分为思茅松和其它物种2个类型。将区分好的细根样品放置烘箱65℃下烘干至恒重,用电子天平称质量(精确到0.001 g),根据以下公式计算细根、粗根和死根的生物量[4-5, 7]:
Broot =m×100/ [π(r/2)2]
式中,Broot为林分的细根、粗根及死根的生物量(t·hm-2),m为平均每个土芯的根干质量(g),r为根钻的直径(cm),本文为8.5 cm。
-
将根系生物的数据中不符合正态分布的数据进行对数转化,使其服从正态分布,对不同林龄思茅松人工林的林分密度、胸径、树高、胸高断面积、根系生物量、细根生物量、粗根生物量以及死根生物量及其垂直分布进行单因素方差分析(ANOVA),并用最小显著法(LSD)进行多重比较。利用有交叉作用的双因子方差分析,将思茅松、其它物种及全部细根生物量、思茅松粗根、其它物种粗根及全部粗根生物量、死根生物量以及根系生物量作为因变量,林龄和土层深度作为固定因素,分析所有因素对思茅松根系生物量分布的影响效应。所有数据分析在R3.4.0中完成,同时利用SigmaPlot 10.0软件进行作图。
-
不同林龄思茅松人工林根系生物量分布见表 2。将思茅松人工林中的根系分为细根、粗根和死根等3个类型,细根生物量的大小顺序分别是:36年生>5年生>15年生>8年生>25年生,其中,思茅松细根生物量最大的是8年生思茅松人工林,其次是5年生;其它物种的细根生物量中最大的是36年生,显著高于8、15、25年生的思茅松人工林。随着林龄的增长,思茅松人工林的粗根生物量也随之增加,其中,思茅松粗根生物量在不同林龄之间无显著差异,其它物种的粗根生物量随林龄的增长而增加。15年生思茅松人工林的死根生物量最大,是36年生思茅松林的3.16倍。36年生思茅松林的根系生物量显著高于8年生和25年生思茅松人工林。活/死细根比是细根现存量与细根的发生与分解中十分重要的根系参数,土壤水分、养分、树木年龄等会对活/死根比产生重要影响[4],思茅松人工林的活/死根比随林龄的增长而增大,反映了林龄较大的思茅松根系处于较快的生长、死亡和周转过程中。
表 2 不同林龄思茅松人工林根系生物量分布特征
Table 2. Distribution traits of root biomass in different stand age of Pinus kesiya var. langbianensis plantation
类型Trait 林龄stand age/a 5 8 15 25 36 细根生物量
Fine root biomass/(t·hm-2)思茅松Pinus kesiya 2.42±0.50a 2.72±0.62a 2.35±0.55a 1.23±0.09b 1.2±0.31b 其它物种Other species 3.56±0.69ab 2.27±1.02b 3.28±1.11b 3.28±1.41b 6.2±0.59a 合计Total 5.98±1.01ab 5.00±0.43b 5.63±0.63ab 4.51±1.43b 7.4±0.76a 粗根生物量
Coarse root biomass/(t·hm-2)思茅松Pinus kesiya 0.62±0.41a 0.99±0.27a 0.65±0.24a 0.88±0.42a 0.63±0.16a 其它物种Other species 0.70±0.19c 0.65±0.04c 1.23±0.43bc 2.22±1.09ab 2.86±0.66a 合计Total 1.32±0.29b 1.64±0.28b 1.88±0.37b 3.10±0.93a 3.5±0.83a 死根生物量Dead /(t·hm-2) 0.92±0.48a 1.03±0.14a 1.17±0.42a 0.52±0.29ab 0.37±0.29b 根系生物量Root biomass/(t·hm-2) 8.22±1.68ab 7.67±0.12b 8.68±0.87ab 8.14±2.66b 11.26±1.81a 注:同行不同字母表在P < 0.05水平下差异显著。
Notes:the different letters with in the same row mean significant difference at the 0.05 level. -
不同林龄思茅松人工林根系生物量垂直分布分析结果(图 1)表明:在细根生物量的垂直分布上,25年生思茅松人工林中细根生物量在不同土壤深度之间无显著差异,其它林龄的思茅松人工林细根生物量总量随土壤深度增加细根生物量呈减少趋势。细根生物量主要分布在土壤表层中,其中,土壤深度0~10 cm的细根生物量占所有土层细根生物量的41.37%~54.34 %,在8年生思茅松人工林中,思茅松细根生物量占所有思茅松细根生物量的56.10 %;土壤深度10~20 cm中细根生物量占所有土层细根生物量的21.60%~36.24%;而土壤深度20~30 cm中的细根生物量占所有土层细根生物量的17.97%~25.42%。
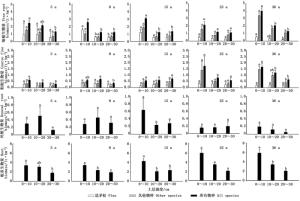
图 1 不同林龄思茅松人工林根系生物量的垂直分布(图中字体调大1号)
Figure 1. Vertical distribution of root biomass in different stand age of Pinus kesiya var. langbianensis plantations
不同林龄思茅松人工林中粗根生物量差异较大,5、15、和25年生人工林中粗根生物量之间无显著差异,而8年生思茅松人工林中,土壤深度10~20 cm的中粗根生物量最大,主要是其它物种粗根生物量的贡献;36年生思茅松人工林中,粗根生物量随土层增加而减少,其中,思茅松粗根之间无显著差异,主要是其它物种的粗根生物量之间差异显著。不同林龄思茅松人工林的死根生物量之间无显著差异。不同林龄思茅松人工林的根系生物量在垂直方向上的大小差异显著,随着土壤深度的增加根系生物量减少。
-
思茅松人工林中不同土层根系生物量与土壤深度和林龄的方差分析(表 3)显示:林龄、林龄与土壤深度的交叉作用对思茅松人工林细根生物量无显著影响,而土壤深度对细根生物量有显著影响,其中,思茅松细根受土壤深度和林龄影响较大,而林龄与土壤深度的交叉作用则无显著影响,其它物种的细根生物量则受林龄、土壤深度以及林龄与土壤深度的交叉作用影响显著或极显著;林龄、土壤深度及林龄与土壤深度的交叉作用对思茅松人工林的粗根生物量之间具有显著或极显著的影响,其中,土壤深度及林龄与土壤深度的交叉作用对思茅松粗根生物量无显著影响,而对其他物种粗根生物量有显著或极显著影响。土壤深度及林龄与土壤深度的交叉作用对思茅松人工林死根生物量无显著影响,而对思茅松人工林所有根系生物量都有显著或极显著影响。
表 3 土壤深度和林龄对思茅松人工林细根、粗根和死根生物量交互影响的方差分析
Table 3. The effect of different soil depth and stand age on the fine root, coarse root and dead root biomass of Pinus kesiya var. langbianensis plantation
类型Types 变异来源
Source of variation自由度
Degree of freedomⅢ平方和Ⅲ
Type sum of square均方
Mean squareF 差异显著性
Significance细根生物量
Fine root biomass/(t·hm-2)思茅松
Pinus kesiya林龄Stand age 4 1.735 1.735 18.975 P < 0.001 土壤深度Soil depth 2 1.915 1.915 20.951 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 0.11 0.11 1.207 0.278 其它物种
Other species林龄Stand age 4 5.448 5.448 19.714 P < 0.001 土壤深度Soil depth 2 8.569 8.569 31.008 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 2.156 2.156 7.803 P < 0.01 合计Total 林龄Stand age 4 1.034 1.034 2.768 0.104 土壤深度Soil depth 2 18.586 18.586 49.744 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 1.291 1.291 3.456 0.07 粗根生物量
Coarse root biomass/(t·hm-2)思茅松
Pinus kesiya林龄Stand age 4 0.004 0.004 0.19 0.665 土壤深度Soil depth 2 0.016 0.016 0.769 0.386 林龄×土壤深度
Stand age×Soil depth8 0.013 0.013 0.319 0.436 其它物种
Other species林龄Stand age 4 3.743 3.743 30.514 P < 0.001 土壤深度Soil depth 2 1.652 1.652 13.649 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 1.215 1.215 9.905 P < 0.01 合计Total 林龄Stand age 4 3.501 3.501 22.384 P < 0.001 土壤深度Soil depth 2 1.998 1.998 12.773 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 0.975 0.975 6.234 P < 0.05 死根生物量Dead root biomass/(t·hm-2) 林龄Stand age 4 0.336 0.336 6.252 P < 0.05 土壤深度Soil depth 2 0.106 0.106 1.981 0.167 林龄×土壤深度
Stand age×Soil depth8 0.001 0.001 0.009 0.926 根系生物量
Root biomass/(t·hm-2)林龄Stand age 4 5.33 5.33 7.189 P < 0.05 土壤深度
Soil depth2 36.61 36.61 49.385 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 4.42 4.42 5.96 P < 0.05 -
不同物种、不同林龄林分地上和地下部分的生物量大小分布差异较大,思茅松人工林地下生物量比例随林龄的增长逐渐增加,并成为森林碳储量的主要来源[12]。本研究发现,思茅松人工林中不同物种组成之间的细根生物量随林龄增长呈现截然相反的变化趋势,思茅松细根生物量随林龄的增长而减少,而其它物种细根生物量则随林龄的增长而增加。林分发展过程中,群落组成、林分结构、土壤理化性质及养分含量变化会导致细根生物量组成和大小分布的变化[15]。研究表明,植物获取土壤资源的多寡以及植物个体间或种群间对土壤资源的竞争能力决定了根系在土壤中的大小结构和分布[4]。一方面,由于思茅松是阳性树种,是南亚热带季风常绿阔叶林群落演替的先锋树种,能适应多种生境,在演替早期,对养分、水分的需求高于演替后期的林分,生长旺盛,思茅松更倾向于分配更多的生物量给根系,因而细根较多,并通过细根迅速、广泛的占据土壤空间获取资源[2];同时,在思茅松人工林幼龄林阶段,有效的林分管理措施,如除草及择伐等抚育手段,使思茅松人工林内具有较小的种间竞争,尤其是根系竞争,个体单株有更多的生存空间、养分及水分可以用来维持根系的生长[16],根生长速率更快,因而思茅松的细根生物量更高;另一方面,随着林龄的增长,在思茅松中龄林和近熟林阶段,由于林分内乔木的自疏效应以及其它物种在群落中优势增大,物种丰富度增加且群落结构更加复杂,土壤有机质和养分含量更加丰富,种间竞争变得相对激烈[17],为提高种间和种内竞争力,其它物种对资源的利用势必影响到思茅松细根对养分的利用效率,导致思茅松细根生物量降低,而随着耐阴阔叶树种在群落中增加,群落分层开始出现,即思茅松占据乔木上层,其它阔叶树种占据乔木下层,林冠更加稠密,透光度更小,演替后期树种的细根相对其演替早期具有更强的养分获取能力,生长速度可能更快[2],而阔叶树种比针叶树种有更高的细根生物量[18],因而,其它物种的细根生物量在增加。
由于土壤资源有效性在空间上分布的差异及外界环境条件的影响,思茅松人工林细根生物量垂直分布随土层的加深而减少,且主要分布在土壤表层。研究表明,幼龄或早期演替阶段的腐质层薄、土壤贫瘠,随着林分的发展,大量凋落物在表层积累,使细根趋于土壤表层,尤其是在同一树种年龄较大的林分[4]。思茅松的细根主要分布在土壤深度0~30 cm内,77 %的细根出现在土壤表层20 cm内[1],马尾松人工林中有79.2 %的细根生长量也分布土壤表层30 cm内[2],思茅松根系一般主根较明显,根系生物量更倾向于侧根,以保持和土壤之间巨大的抓合力,而侧根主要分布在土壤表层[13, 19]。随着林龄的发展,思茅松人工林土壤表层养分含量渐渐增高,且与较深的土层相比,水分充足,温度适宜,同时土壤表层具有更高的沙子比例、较低的土壤密度以及相对较高的有机质、氮和磷含量[11-12],从而使表层土壤养分丰富,且结构较疏松,具有更好的通气状况,渗透速率较快,对根系穿透的机械阻力就越小[3],细根则主要集结在土壤表层内以获取水分和养分,而土壤下层由于养分及其它生物状况的恶化,根系生物量则显著下降[20]。
林龄和土壤深度对思茅松人工林内不同物种组成的细根、粗根及根系有显著影响,其中,随着林龄的增大,人工林内细根生物量随物种多样性增加而增加,这是由于群落物种对资源利用的生态位分化,形成对资源利用的互补,同时减少了细根感染死亡的几率[21]。细根生物量在思茅松人工林中由于物种组成的不同而差异较大,在5年生人工林阶段,由于思茅松处于幼龄林阶段,人工林郁闭度较低,光照充分,适应草本植物的生长,其主要分布在土壤表层中[14, 22],因而草本植物细根生物量在垂直分布上超过相应的思茅松细根生物量;到8年生人工林阶段,思茅松林冠增大,形成郁闭,林内草本植物减少,各土层思茅松细根生物量的比例高于其它植物。此外,随着林龄的继续发展,思茅松林内出现较多的耐阴乔木与灌木植物,改变了群落的物种组成,因而,其它物种的细根生物量又逐渐占据优势。由于粗根的主要功能不在于养分吸收,而在于为乔木个体提供机械支撑,因而主要分布在养分贫瘠的土壤亚层中[1],随着林龄增长,更多的细根转化成粗根为乔木提供更好的结构支持,在近熟林阶段,其它耐阴植物贡献了大部分的粗根生物量。
-
随着思茅松人工林林龄增大根系生物量呈增加趋势,由于群落结构与物种组成发生较大变化,不同物种之间根系生物量大小分布也产生较大差异,其中,思茅松细根生物量随林龄增大而减少,而其它植物细根随林龄增大而增加;同时,粗根生物量随林龄增大的变化趋势主要是由于其它物种粗根生物量贡献量的增加。思茅松人工林根系生物量主要分布在土壤表层,随土壤深度增加而减少,受林龄与土层深度影响较显著。本文仅从林龄角度分析思茅松人工林细根生物量的大小分布特征,而未进行土壤因子等环境因子对细根生长综合影响的分析,要准确评价思茅松人工林根系生物量的动态特征,需深入研究思茅松人工林细根生物量在不同生长季的变化。
林龄对思茅松人工林根系生物量的影响
Effect of Stand Age on Root Biomass in Pinus kesiya var. langbianensis Plantation
-
摘要:
目的 以云南省普洱市主要植被思茅松人工林为研究对象, 探讨不同林龄思茅松人工林根系生物量的大小分布及变化特征。 方法 分别在5、8、15、25、36年生思茅松人工林内, 利用内径为8.5 cm的根钻分3层(010、1020、2030 cm)获取思茅松与其它物种的细根、粗根及死根生物量数据。 结果 表明:随着思茅松人工林林龄的增长, 思茅松细根生物量呈减少的趋势, 而其它物种细根生物量呈增加趋势, 细根生物量最大出现在36年生思茅松人工林。不同林龄思茅松人工林的思茅松粗根和死根生物量之间无显著差异, 而其它物种及林分的粗根生物量和根系生物量则随林龄增长而增加。思茅松人工林的细根生物量主要分布在土壤深度010 cm内, 其中, 思茅松、其它物种、林分细根生物量以及根系生物量随土层深度的增加呈减少趋势。林龄和土壤深度对思茅松与其它物种的细根生物量有显著影响, 林龄与土壤深度的交叉作用对思茅松细根生物量有显著影响, 林龄对死根生物量有显著影响, 林龄、土壤深度及林龄与土壤深度的交叉作用对粗根与根系的生物量有显著影响。 结论 思茅松人工林随着林龄增长, 群落结构与树种组成随之发生变化, 从而对根系生物量产生较大影响。 Abstract:Objective To analyze the size distribution and determine the effects of stand age on root biomass in Pinus kesiya var. langbianensis plantation based on the data of fine root, coarse root and dead root biomass among different species groups. Method This study was conducted in 5-year-old, 8-year-old, 15-year-old, 25-year-old and 36-year-old P. kesiya" var.langbianensis plantations located in Pu'er city, Yunnan province, China. The soil cores with diameter of 8.5 cm were collected in October, 2014 at different stand age. Each soil core was separated into three sections at the depths of 010, 1020, and 2030 cm. Result The fine root biomass of P. kesiya var. langbianensis decreased with the stand age, while the fine root biomass of other species groups increased with the stand age. The maximum of fine root biomass was found in the 36-year-old P. kesiya var. langbianensis plantation. There were no significant difference in the coarse root of P. kesiya var. langbianensis and dead root of different stand age in P. kesiya var. langbianensis plantations. On the contrary, the coarse root biomass of other species group and stand, and root biomass increased with the stand age. The fine root biomass mainly distributed in the soil with the depth of 010 cm. The fine root biomass of P. kesiya var. langbianensis, the other species group and all stand as well as root biomass decreased with the increase of soil depth. The stand age and soil depth had significant effect on the fine root biomass of P. kesiya var. langbianensis and other species group, respectively. The cross effect of stand age and soil depth had no significant influences on the fine root biomass of P. kesiya var. langbianensis. The stand age had a significant effect on the dead root biomass. The stand age, soil depth and the cross of stand age and soil depth had significant effect on the coarse root and all root biomass. Conclusion With the growth of P. kesiya var. langbianensis plantations, the community structures and species composition will change and has a significant effect on the root biomass. -
Key words:
- Pinus kesiya var. langbianensis
- / fine root
- / coarse root
- / biomass
- / stand age
- / vertical distribution
-
图 1 不同林龄思茅松人工林根系生物量的垂直分布(图中字体调大1号)
Figure 1. Vertical distribution of root biomass in different stand age of Pinus kesiya var. langbianensis plantations
表 1 思茅松人工林林分特征
Table 1. Stand traits in the Pinus kesiya var. langbianensis plantations
林龄
Stand age/a地点
Location纬度(N)
Latitude纬度(E)
Longitude海拔
Altitude/m胸径
DBH/cm树高
Height/m胸高断面积
Basal area at DBH/cm2林分密度
Stand density/(trees·hm-2)5 万掌山林场 22°49′ 100°53′ 1 410 6.46±0.36c 4.63±0.37d 5.72±0.99d 1 600±150a 8 万掌山林场 22°49′ 100°54′ 1 485 12.50±0.53b 9.36±0.82c 15.3±1.24c 1 200±156b 15 清水河林区 22°41′ 100°57′ 1 270 13.33±1.38b 10.94±0.66b 23.56±4.50b 1 130±5b 25 大寨 22°45′ 101°2′ 1 415 19.84±1.51a 16.77±0.21a 26.21±3.42a 817±226b 36 梅子湖水库 22°44′ 100°58′ 1 370 12.04±1.73b 8.63±0.72c 30.56±9.60a 1 989±377a 注:同行不同字母表示差异显著P < 0.05。
Notes:the different letters with in the same row mean significant difference at the 0.05 level. 下载: 导出CSV
下载: 导出CSV
表 2 不同林龄思茅松人工林根系生物量分布特征
Table 2. Distribution traits of root biomass in different stand age of Pinus kesiya var. langbianensis plantation
类型Trait 林龄stand age/a 5 8 15 25 36 细根生物量
Fine root biomass/(t·hm-2)思茅松Pinus kesiya 2.42±0.50a 2.72±0.62a 2.35±0.55a 1.23±0.09b 1.2±0.31b 其它物种Other species 3.56±0.69ab 2.27±1.02b 3.28±1.11b 3.28±1.41b 6.2±0.59a 合计Total 5.98±1.01ab 5.00±0.43b 5.63±0.63ab 4.51±1.43b 7.4±0.76a 粗根生物量
Coarse root biomass/(t·hm-2)思茅松Pinus kesiya 0.62±0.41a 0.99±0.27a 0.65±0.24a 0.88±0.42a 0.63±0.16a 其它物种Other species 0.70±0.19c 0.65±0.04c 1.23±0.43bc 2.22±1.09ab 2.86±0.66a 合计Total 1.32±0.29b 1.64±0.28b 1.88±0.37b 3.10±0.93a 3.5±0.83a 死根生物量Dead /(t·hm-2) 0.92±0.48a 1.03±0.14a 1.17±0.42a 0.52±0.29ab 0.37±0.29b 根系生物量Root biomass/(t·hm-2) 8.22±1.68ab 7.67±0.12b 8.68±0.87ab 8.14±2.66b 11.26±1.81a 注:同行不同字母表在P < 0.05水平下差异显著。
Notes:the different letters with in the same row mean significant difference at the 0.05 level.
下载: 导出CSV
表 3 土壤深度和林龄对思茅松人工林细根、粗根和死根生物量交互影响的方差分析
Table 3. The effect of different soil depth and stand age on the fine root, coarse root and dead root biomass of Pinus kesiya var. langbianensis plantation
类型Types 变异来源
Source of variation自由度
Degree of freedomⅢ平方和Ⅲ
Type sum of square均方
Mean squareF 差异显著性
Significance细根生物量
Fine root biomass/(t·hm-2)思茅松
Pinus kesiya林龄Stand age 4 1.735 1.735 18.975 P < 0.001 土壤深度Soil depth 2 1.915 1.915 20.951 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 0.11 0.11 1.207 0.278 其它物种
Other species林龄Stand age 4 5.448 5.448 19.714 P < 0.001 土壤深度Soil depth 2 8.569 8.569 31.008 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 2.156 2.156 7.803 P < 0.01 合计Total 林龄Stand age 4 1.034 1.034 2.768 0.104 土壤深度Soil depth 2 18.586 18.586 49.744 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 1.291 1.291 3.456 0.07 粗根生物量
Coarse root biomass/(t·hm-2)思茅松
Pinus kesiya林龄Stand age 4 0.004 0.004 0.19 0.665 土壤深度Soil depth 2 0.016 0.016 0.769 0.386 林龄×土壤深度
Stand age×Soil depth8 0.013 0.013 0.319 0.436 其它物种
Other species林龄Stand age 4 3.743 3.743 30.514 P < 0.001 土壤深度Soil depth 2 1.652 1.652 13.649 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 1.215 1.215 9.905 P < 0.01 合计Total 林龄Stand age 4 3.501 3.501 22.384 P < 0.001 土壤深度Soil depth 2 1.998 1.998 12.773 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 0.975 0.975 6.234 P < 0.05 死根生物量Dead root biomass/(t·hm-2) 林龄Stand age 4 0.336 0.336 6.252 P < 0.05 土壤深度Soil depth 2 0.106 0.106 1.981 0.167 林龄×土壤深度
Stand age×Soil depth8 0.001 0.001 0.009 0.926 根系生物量
Root biomass/(t·hm-2)林龄Stand age 4 5.33 5.33 7.189 P < 0.05 土壤深度
Soil depth2 36.61 36.61 49.385 P < 0.001 林龄×土壤深度
Stand age×Soil depth8 4.42 4.42 5.96 P < 0.05
下载: 导出CSV
-
[1] John B, Pandey H N, Tripathi R S. Vertical distribution and seasonal changes of fine and coarse root mass in Pinus kesiya Royle ex. Gordon forest of three different ages[J]. Acta Oecologica, 2011, 22(5-6):293-300. [2] 陈云玉, 熊德成, 黄锦学, 等.中亚热带不同演替阶段的马尾松和米槠人工林细根生物量研究[J].植物生态学报, 2015, 39(11):1071-1081. doi: 10.17521/cjpe.2015.0104 [3] 张小全.环境因子对树木细根生物量、生产与周转的影响[J].林业科学研究, 2001, 14(5):566-573. doi: 10.3321/j.issn:1001-1498.2001.05.017 [4] 程瑞梅, 王瑞丽, 肖文发, 等.三峡库区马尾松根系生物量的空间分布[J].生态学报, 2012, 32(3):823-832. [5] 刘运科, 范川, 李贤伟, 等.间伐对川西亚高山粗枝云杉人工林细根生物量及碳储量的影响[J].植物生态学报, 2012, 36(7):645-654. [6] Børja I, Dewit H A, Steffenrem A, et al. Stand age and fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway[J]. Tree Physiology, 2008, 28(5):773-784. doi: 10.1093/treephys/28.5.773 [7] Yuan Z Y, Chen H Y H. Fine root dynamics with stand development in the boreal forest[J]. Functional Ecology, 2012, 26(4):991-998. doi: 10.1111/fec.2012.26.issue-4 [8] Jagodziński A M, Kalucka I. Fine roots biomass and morphology in a chronosequence of young Pinus silvestris stands growing on a reclaimed lignite mine spoil heap[J]. Dendrobiology, 2010, 64:19-31. [9] Yanai R D, Park B B, Hamburg S P. The vertical and horizontal distribution of roots in northern hardwood stands of varying age[J]. Canadian Journal of Forest Research, 2006, 36(2):450-459. doi: 10.1139/x05-254 [10] Mou P, Jones R H, Mitchell R J, et al. Spatial distribution of roots in Sweetgum and Loblolly pine monocultures and relations with above-ground biomass and soil nutrients[J]. Functional Ecology, 1995, 9(4):689-698. doi: 10.2307/2390162 [11] 李帅锋, 苏建荣, 刘万德, 等.思茅松人工林土壤有机碳和氮储量变化[J].林业科学研究, 2015, 28(6):810-817. doi: 10.3969/j.issn.1001-1498.2015.06.008 [12] Li S F, Su J R, Liu W D, et al. Changes in biomass carbon and soil organic carbon stocks following the conversion from secondary coniferous forest to a pine plantation[J]. Plos One, 2015, 10(9):e0135946. doi: 10.1371/journal.pone.0135946 [13] 贾呈鑫卓, 李帅锋, 苏建荣, 等.思茅松人工林根系特征与生物量分配[J].生态学杂志, 2017, 36(1):21-28. [14] 郑金萍, 范春楠, 郭忠玲, 等.长白落叶松人工林生物量及其动态研究[J].南京林业大学学报:自然科学版, 2014, 28(2):175-179. [15] 杨秀云, 韩有志, 张芸香, 等.采伐干扰对华北落叶松细根生物量空间异质性的影响[J].生态学报, 2012, 32(1):64-73. [16] Makkonen K, Helmisaarl H S. Fine root biomass and production in Scots pine stands in relation to stand age[J]. Tree Physiology, 2001, 21(2-3):193-198. doi: 10.1093/treephys/21.2-3.193 [17] Brassard B W, Chen H Y H, Bergeron Y.Influence ofenvironmental variability on root dynamics in northernforests[J]. Critical Reviews in Plant Sciences, 2009, 28(3):179-197. doi: 10.1080/07352680902776572 [18] Akaburak S, Oral H V, Ozdemir E, et al. Temporal variations of biomass, carbon and nitrogen of roots under different tree species[J]. Scandinavian Journal of Forest Research, 2013, 28(1):8-16. doi: 10.1080/02827581.2012.679680 [19] 吴兆录.思茅松研究现状的探讨[J].林业科学, 1994, 30(2):151-157. [20] Bruton A J, Pregiter K S, Hendrick R L. Relationships between fine root dynamics and nitrogen availability in Miehigan nothern hardwood forests[J]. Oecologica, 2000, 125(3):389-399. doi: 10.1007/s004420000455 [21] Brassard B W, Chen H Y H, Cavard X, et al. Tree species diversity increases fine root productivity through increased soil volume filling[J]. Journal of Ecology, 2013, 101(1):210-219. doi: 10.1111/jec.2012.101.issue-1 [22] Olesinski J, Lavigne M B, Jr J A K, et al. Fine-root dynamics change during stand development and in response to thinning in balsam fir (Abies balsamea L.Mill.) forests[J]. Forest Ecology and Management, 2012, 286:48-58.) doi: 10.1016/j.foreco.2012.08.047 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3631
- HTML全文浏览量: 1366
- PDF下载量: 518
- 被引次数: 0
