




-
被子植物花朵的结构特征是其有性生殖与环境相适应的表现[1],因此,植物花朵的形态结构特征、开花进程及花期物候等与其生态进化之间紧密联系在一起[2],观测花期物候是研究植物生殖生态学的重要内容。有关植物开花动态的研究已有许多报道,其中,在生态异质生境中及种群内的变化动态是研究热点之一[3-5]。高等植物的花、花序、果实和种子及着生这些器官的生殖枝被称作生殖构件[6],而每种植物生殖构件的分布格局和数量变化具有一定的规律性[7],主要由植物自身遗传及植物所处环境因子决定[8]。
灰木莲(Manglietia glauca Blume)为木兰科木莲属常绿阔叶大乔木,其干形通直、树形优美、花大洁白有芳香,是一种较好的速生用材及园林绿化树种。灰木莲原产越南及印度尼西亚,1960年,我国从越南引种在中国林科院夏石树木园试种[9],生长表现良好[10],具有较好的发展前景。在广东、云南、福建等地相继成功开展了灰木莲的引种栽培[11]。在原产地越南,灰木莲可以正常结实,引种到中国后开花结实率极低[12],严重制约其大规模发展。因此,本文从灰木莲在广西凭祥地区的开花进程、花期物候特征及生殖构件分布等方面开展研究,探索其生殖生态特征,以期为灰木莲在引种地败育机制研究积累资料。
-
试验地设置于广西凭祥市(21°57′47″~22°19′27″ N;106°39′50″~106°59′30″ E)中国林业科学研究院热带林业实验中心树木园、白云实验场和伏波实验场,属南亚热带季风气候区,年均气温21.5℃,≥10℃积温7 500 ℃,年降水量1 220~1 400 mm,降水集中期为4月中旬至8月末,年均日照约1 260 h。
选择中国林科院热林中心树木园(ZX)、白云实验场(BY)、伏波实验场(FB)灰木莲人工林作为观测对象。造林所用苗木均是用越南老街省宝安县收集种子培育的1年生苗。各林分无病虫侵害,健康状况良好,基本情况见表 1。
表 1 观测林分基本情况
Table 1. General information of the observation stands
林分Stand 造林时间(年)Planting time(year) 树高Tree height/m 胸径DBH/cm 保留密度Conserved density/(tree·hm-2) 海拔Altitude/m ZX 2002 16.69±1.13 26.13±1.46 800 240 BY 1998 18.45±3.75 28.70±5.59 1 200 540 FB 2003 24.65±2.85 32.40±4.19 1 000 630 -
参照Dafni[13]方法观测灰木莲的开花物候。2014年01—05、10—12月,2015年01—05、10—12月,2016年01—05月,从林分和单株水平,对中国林科院热林中心树木园、白云实验场、伏波实验场3个生境下的灰木莲人工林的开花物候进行定期观测。
-
开花前,在中国林科院热林中心树木园、白云实验场、伏波实验场分别选择5、30、30棵单株进行标记,观察记录始花日期(单株开花数达5%的开花日期为始花期)及当日花数,开花盛花期(单株开花数≥50%)及当日花数、持续时间,终花期(95%的花已开放)及当日花数、平均开花振幅、相对开花强度和开花同步性(同步指数)。
-
5%的个体开花为种群始花期,50%的个体达到开花高峰时为种群开花高峰期,95%的植株开花结束时为种群花期结束。
用“花数·株-1·d-1”来表示平均开花振幅[14-15],描述单位时间开花数。
单株相对开花强度指单株开花高峰日开放的花朵数量与该种群中植株在其开花高峰日产生的单株最大花数之比[16]。
根据文献[16]用同步指数(Si)检测开花同步性高低。
$ S_{i}=\frac{1}{n-1}\left(\frac{1}{f_{i}}\right) \sum\limits_{j=1}^{n} e_{j} \neq i $
式中:n为样地中个体总数,fi为个体i开花的总时间(d),ej为个体i和j花期重叠时间(d);Si的变异范围为0~1,“0”表示种群内个体花期无重叠,“1”则表示完全重叠。
-
分别于2014年及2015年灰木莲盛花期,在中国林科院热林中心树木园固定选择5株灰木莲定时观察开花动态。记录花蕾期花蕾发育的形态变化、花被片张开到花被片全部脱落的时间及特征、雌蕊群和雄蕊群的形态特征及动态变化等。取新鲜花样,带回室内置于Olympus SZX7体视镜下观察、拍照,测量与统计花部组件大小、数量及位置等指标。
-
在3个海拔高度的灰木莲人工林中,分别选择10棵标准木,林内、林缘各5棵。每棵间距20 m以上,分上、中、下3个冠层选标准枝,在花期记录生殖枝、花、花芽数量及花、花芽在生殖枝上的位置。
-
采用Excel 2013进行数据整理作图,用DPS V14.50进行单因素方差分析,采用LSD法进行多重比较。
-
灰木莲花属两性单花,生于枝顶。花蕾期,花蕾外部有2层苞片,厚纸质,多呈绿色,在展开前期逐渐变成褐色。花开后,直径15~25 cm,具香味,无花盘或蜜腺,花被片乳白或绿白色,厚肉质,汤勺形,长6~10 cm,宽5~7 cm,共6片;雌蕊群由45~55枚单生心皮组成,心皮绿色短纺锤形,长0.4~0.6 cm,呈锥形紧密组合在一起,柱头微向外卷曲;雄蕊长0.7~1.1 cm,宽0.08~0.15 cm,乳白色,花药内向开裂。雄蕊群着生于雌蕊群的下部,雄蕊短小,其高度仅达雌蕊群的下缘(图 1)。

图 1 灰木莲花部形态
Figure 1. The flower morphology of M. glauca Blume
-
灰木莲花芽到开花,过程可分为6个阶段(图 2):(1)混合芽阶段:该阶段芽以混合芽形式存在,花被片外具有2层内层苞片,之外还存在4~5层外苞片,每层外苞片内包有一对新叶,此阶段可持续45~50 d。(2)花芽阶段:待混合芽外层苞片脱落完毕,花被片外仅剩一层内苞片,此时的芽称为花芽,此阶段持续25~30 d。(3)花蕾露白阶段:花芽个体发育已足够大,内苞片停止生长,花被片生长舒展将内苞片顶开,花蕾顶部露出一点白色的花被片,此阶段可持续1~2 d。(4)白蕾阶段:外苞片脱落,外层绿色花被片完全展开,露出6片白色花被片并抱合组成的花蕾,称为白蕾,此时花蕾内柱头向外卷曲,带有透明液体,至花被片完全展开成花瓣为止,此阶段需1~3 d,在此阶段花被片展开经历第1次开放与长闭合。(5)展花阶段:在形成白蕾第2天,花瓣出现松动,其中1~2片花瓣微开(持续3~5 h),此时雄蕊群开始松动,柱头粘液量达到最多;至下午5:00微开的花瓣再次闭合(短闭合),之后1~2 h花瓣完全展开,同时花粉从花药中散出,雄蕊群完全脱落,至花瓣脱落为止;自花瓣完全展开柱头粘液颜色逐渐加深,变成深褐色胶体,该阶段持续4~6 h;(6)花瓣脱落阶段:花瓣颜色由白色逐渐变成褐色脱落。如遇阴雨天气,温度降低,开花进程将推迟1~2 d。

图 2 灰木莲开花动态
Figure 2. Floral dynamic of M. glauca Blume
-
表 2表明:热林中心树木园(ZX)(海拔240 m)的灰木莲花期主要从3月上旬持续至5月上旬,少数从1月下旬开始,偶尔出现一年中2次花期,第2次花期从10月中旬持续到12月上旬;第1次花期3月上、中旬为始花期,3月下旬到4月中旬为盛花期,4月下旬到5月上旬为终花期。白云实验场白云山(BY)(海拔540 m)的灰木莲林分的开花物侯始花期比热林中心树木园相应推迟15~20 d,盛花期推迟8~10 d,终花期推迟6~7 d;伏波实验场(FB)(海拔630 m)的灰木莲林分的开花物侯始花期比热林中心树木园相应推迟18~25 d,盛花期推迟12~15 d,终花期推迟8~10 d。灰木莲开花相对集中,同步指数较高。
表 2 灰木莲林分水平开花物侯
Table 2. Phenology of M. glauca Blume at stand level
林分Stand 始花期First-flowering stage 花期持续时间Duration /d 盛花期Full-bloom Stage 平均开花振幅Mean flowering amplitude/ (flower·株-1·d-1) 终花期Final flowering stage 平均开花同步性指数Synchrony index 时间(月-日)Time(month-day) 平均花数Average number of flower /flower 时间(月-日)Time(month-day) 平均花数Average number of flower /flower 时间(月-日)Time(month-day) 平均花数Average number of flower /flower ZX 03-10 11 53 04-05 102 21.65 05-05 5 0.87±0.03a BY 03-25 8 46 04-12 63 14.31 05-10 3 0.68±0.07b FB 03-28 7 51 04-25 80 16.87 05-15 2 0.71±0.04c 注:表中字母表示各处理间的LSD多重比较结果。Note: The letters in the figure refer to the results of multiple comparison of LSD. 灰木莲在广西凭祥地区个体相对开花强度(图 3)表明:在该地区有2个主要分布频度范围,分别在30%~50%、70%~80%,开花强度中等。
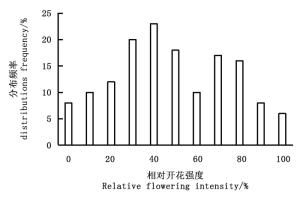
图 3 灰木莲个体相对开花强度分布
Figure 3. Relative flowering intensity frequency distributions of individual plants of M. glauca Blume
由图 4可知:热林中心树木园(ZX)、白云实验场(BY)、伏波实验场(FB)灰木莲林分的开花进程基本相似,均呈上升至高峰期后下降趋势。与热林中心树木园林分相比,白云实验场、伏波实验场灰木莲林分达到开花高峰期比较缓慢。4月是广西凭祥灰木莲开花高峰期。
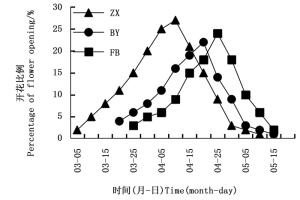
图 4 不同灰木莲林分的开花物候曲线
Figure 4. Flowering phenology curves for different stands of M. glauca Blume
-
热林中心树木园、白云实验场、伏波实验场不同海拔灰木莲林分的花枝/枝、花/花枝、果和果/花统计见表 3。表 3表明:热林中心树木园(ZX)(海拔240 m)与伏波实验场(FB)(海拔630 m)灰木莲林分的花枝/枝差异不显著,白云实验场(BY)(海拔540 m)与热林中心树木园和伏波实验场灰木莲林分的花枝/枝差异显著;3个林分的花/花枝间差异不显著。从林分密度分析,随林分密度的升高,灰木莲林分的花枝/枝和花/花枝呈下降趋势。灰木莲生殖构件的败育率极高,调查观察发现,只有伏波实验场灰木莲有零星结实,其他林分均未发现结实。
表 3 不同灰木莲林分生殖构件分布
Table 3. Distribution of reproductive modules in different stands
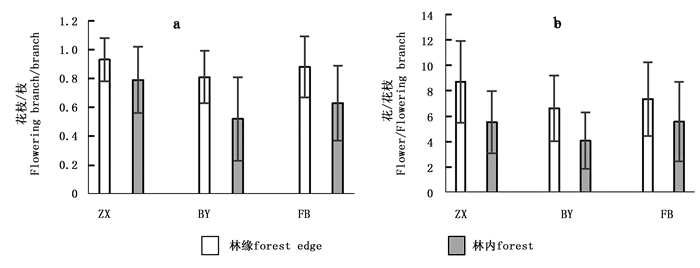
林分Stand 花枝/枝Flowering branch/ branch 花/花枝Flower/Flowering branch 果Fruit 果/花Fruit/Flower ZX 0.81±0.23a 6±2.13 0 0 BY 0.65±0.26b 5±1.56 0 0 FB 0.67±0.13a 6±2.51 3±1.33 0.01±0.008 由图 5可知:在林缘和林内,灰木莲树冠内花枝数量差异显著(P < 0.05),3个立地条件下,林缘植株生殖个体上的花枝均比林内的高,且林缘和林内花枝上的单花数差异显著(P < 0.05),林缘花枝单花数量明显比林内的多。

图 5 不同生境中灰木莲生殖构件的变化
Figure 5. The reproductive modules of M. glauca Blume in the different habitats
-
花部结构特点、展花方式及时间是影响植物有性生殖成功与否的重要因素[17]。传粉是种子植物受精的必经阶段,传粉系统由花粉、柱头、传粉媒介三部分组成[18]。灰木莲为子房上位的两性花,雄蕊群着生于雌蕊群的下部,雄蕊短小,其高度仅达雌蕊群的下缘,花粉无法自然散落到雌蕊柱头上,即使到达柱头,其花粉数量不足,使胚珠难以受精,不利于自然结实。灰木莲花部结构特点决定了虫媒传粉是其有效传粉途径。
植物的开花物候不仅与其遗传特性及其类群的系统发生有关[19],还与环境密切联系[20, 21]。灰木莲引种到广西凭祥,海拔越高,花期开始越晚,表明环境因子对灰木莲花期物侯影响较大。灰木莲单花开放时间相对固定,表明其单花开花节律受遗传因子的影响较大。灰木莲开花强度中等,但开花相对集中,同步指数较高,这与招礼军等[12]对广西南宁地区引种灰木莲的研究结果一致。单花寿命与开花期间的环境条件有关,在始花期,气温较低,且阴雨天气较多,单花寿命较长;在盛花期,气温较高,阴雨天气减少,单花寿命缩短。
根据不同海拔灰木莲林分的生殖构件分布结果分析表明:在适生范围内,海拔高度不是影响灰木莲生殖构件分布的关键因子。从林分密度分析,随着林分密度的升高,灰木莲林分的花枝/枝和花/花枝呈下降趋势,且林缘单株生殖构件数量显著高于林内植株,这表明林分密度影响单株的生长空间,导致树体接受光照不同[22-24]。植物开花数量超过最终成熟果实的数量是普遍现象[25],但灰木莲引种到广西凭祥地区生殖构件的败育率极高,花果转化率近乎为零。影响果实败育的因子有内部因素[26],也有外部因素[27]。因此,为探明灰木莲在引种地种实败育机制,将进一步开展灰木莲雌雄配子体发育、传粉媒介等方面的研究。
-
(1) 灰木莲在引种地区广西凭祥热林中心树木园(ZX)(海拔240 m)的花期从3月上旬持续至5月上旬,偶尔出现一年2次花期,第2次花期从10月中旬持续到12月上旬;第1次花期的始花期为3月上中旬,3月下旬到4月中旬为盛花期,4月下旬到5月上旬为终花期。白云实验场白云山(BY)(海拔540 m)始花期的开花物侯比热林中心树木园的相应推迟15~20 d,盛花期推迟8~10 d,终花期推迟6~7 d;伏波实验场(FB)(海拔630 m)的开花物侯始花期比热林中心树木园相应推迟18~25 d,盛花期推迟12~15 d,终花期推迟8~10 d。
(2) 灰木莲单花开花时间较稳定,均为下午5:00—6:00。灰木莲开花强度中等,分别出现在30% ~50%、70%~80%,但开花相对集中,同步指数较高。单花寿命与开花期间的环境条件有关,在始花期,气温较低,且阴雨天气较多,单花寿命一般2~3 d;在盛花期,气温较高,阴雨天气减少,单花寿命缩短为1~2 d。
(3) 灰木莲的生殖构件分布主要受林分密度的影响,海拔高度不是影响灰木莲生殖构件分布的关键因子。随着林分密度的升高,灰木莲林分的花枝/枝和花/花枝呈下降趋势,林缘单株生殖构件数量显着高于林内植株,这表明林分密度影响单株的生长空间,导致树体接受光照不同。
灰木莲花期物侯观测及生殖构件分布
Flowering Phenology and Reproductive Modules Distribution of Manglietia glanca Blume
-
摘要:
目的 为了解灰木莲引种到广西凭祥地区花期物侯及生殖构件分布状况, 为其在引种地败育机制的研究奠定基础。 方法 选择中国林科院热林中心树木园、白云实验场、伏波实验场的灰木莲人工林为观测对象, 参考Dafni方法, 从林分到单株水平调查研究灰木莲在广西凭祥地区的花期物候、开花特征及生殖构件分布, 跟踪观察单花开花动态进程, 统计花部组件大小、数量及位置等特征。 结果 表明:灰木莲花为子房上位的两性花, 雄蕊群着生于雌蕊群下部, 雄蕊短小, 其高度仅达到雌蕊群下缘; 灰木莲花芽到开花过程可分为混合芽阶段、花芽阶段、花蕾露白阶段、白蕾阶段、展花阶段、花瓣脱落阶段6个阶段, 花被片展开存在二次开合现象; 在适生区, 灰木莲开花特征稳定, 开花强度中等, 不同海拔高度灰木莲林分的花期物侯存在差异, 林分的生殖构件在林内、林缘分布差异显著, 且生殖构件的败育率极高。 结论 灰木莲引种到广西凭祥地区花期物侯稳定, 开花同步性较高, 但是花果转化率极低。 Abstract:Objective The flowering phenology and reproductive modules distribution of Manglietia glanca Blume was studied to provide a basis for the research of its abortion mechanism in introduction area. Method The M. glanca stands in Arboretum, Baiyun Experimental Farm and Fubo Experimental Farm of the Experimental Center of Tropical Forestry in Pingxiang were chosen as observation object. Taking Dafni's method as reference, the flowering phenology, flowering characteristics and reproductive modules distribution of M. glanca were investigated at both stand and individual plant level, the flowering dynamic was observed, and the size, quantity and location of floral components were calculated. Result The results showed that the flower was bisexual flower. The androecium bore in the bottom of the gynoecium, its height reached only the lower edge of the gynoecium. The flowering process could be divided into six stages, i.e. mixed bud, flower bud, white alabastrum appearing, white alabastrum, blooming and falling. Twice opening/closure flowering was observed during the blossoming process. In suitable region, the flowering characteristics of M. glanca were relatively stable, with moderate intensity blossoming. The effect of altitude on flowering phenology was obvious. The distribution of reproductive modules was also significantly different between forest edge and interior, the fruit setting rate was low. Conclusion According to the result of observation, the flowering phase of M. glanca is tranquil, flowering synchronization index is higher, but conversion rate of blossom into bear fruit is extremely low in Pingxiang, Guangxi. -
图 3 灰木莲个体相对开花强度分布
Figure 3. Relative flowering intensity frequency distributions of individual plants of M. glauca Blume
图 4 不同灰木莲林分的开花物候曲线
Figure 4. Flowering phenology curves for different stands of M. glauca Blume
图 5 不同生境中灰木莲生殖构件的变化
Figure 5. The reproductive modules of M. glauca Blume in the different habitats
表 1 观测林分基本情况
Table 1. General information of the observation stands
林分Stand 造林时间(年)Planting time(year) 树高Tree height/m 胸径DBH/cm 保留密度Conserved density/(tree·hm-2) 海拔Altitude/m ZX 2002 16.69±1.13 26.13±1.46 800 240 BY 1998 18.45±3.75 28.70±5.59 1 200 540 FB 2003 24.65±2.85 32.40±4.19 1 000 630  下载: 导出CSV
下载: 导出CSV
表 2 灰木莲林分水平开花物侯
Table 2. Phenology of M. glauca Blume at stand level
林分Stand 始花期First-flowering stage 花期持续时间Duration /d 盛花期Full-bloom Stage 平均开花振幅Mean flowering amplitude/ (flower·株-1·d-1) 终花期Final flowering stage 平均开花同步性指数Synchrony index 时间(月-日)Time(month-day) 平均花数Average number of flower /flower 时间(月-日)Time(month-day) 平均花数Average number of flower /flower 时间(月-日)Time(month-day) 平均花数Average number of flower /flower ZX 03-10 11 53 04-05 102 21.65 05-05 5 0.87±0.03a BY 03-25 8 46 04-12 63 14.31 05-10 3 0.68±0.07b FB 03-28 7 51 04-25 80 16.87 05-15 2 0.71±0.04c 注:表中字母表示各处理间的LSD多重比较结果。Note: The letters in the figure refer to the results of multiple comparison of LSD.
下载: 导出CSV
表 3 不同灰木莲林分生殖构件分布
Table 3. Distribution of reproductive modules in different stands
林分Stand 花枝/枝Flowering branch/ branch 花/花枝Flower/Flowering branch 果Fruit 果/花Fruit/Flower ZX 0.81±0.23a 6±2.13 0 0 BY 0.65±0.26b 5±1.56 0 0 FB 0.67±0.13a 6±2.51 3±1.33 0.01±0.008
下载: 导出CSV
-
[1] 刘林德, 祝宁, 申家恒.刺五加, 短梗五加的开花动态及繁育系统的比较研究[J].生态学报, 2002, 22(7):1041-1048. doi: 10.3321/j.issn:1000-0933.2002.07.010 [2] Dafni A. Pollination ecology[M]. IRL Press Ltd, 1992:1-57. [3] 陈波, 达良俊, 宋永昌.常绿阔叶树种栲树开花物候动态及花的空间配置[J].植物生态学报, 2003, 27(2):249-255. doi: 10.3321/j.issn:1005-264X.2003.02.016 [4] 肖宜安, 何平, 李晓红.濒危植物长柄双花木开花物候与生殖特性[J].生态学报, 2004, 24(1):14-21. [5] Bosch J, Retana J, Cerdá X. Flowering phenology, floral traits and pollinator composition in a herbaceous Mediterranean plant community[J]. Oecologia, 1997, 109(4):583-591. doi: 10.1007/s004420050120 [6] 方炎明, 樊汝汶.植物生殖生态学[M].济南:山东大学出版社, 1996:70. [7] Primack R B, Lloyd D G. Andromonoecy in the New Zealand Montane Shrub Manuka, Leptospermum scoparium(Myrtaceae)[J]. American Journal of Botany, 1980, 67(3):361-368. doi: 10.1002/j.1537-2197.1980.tb07661.x [8] Hendry A P, Day T. Population structure attributable to reproductive time:isolation by time and adaptation by time[J]. Molecular ecology, 2005, 14(4):901-916. doi: 10.1111/j.1365-294X.2005.02480.x [9] 王克建, 蔡子良.热带树种栽培技术[M].广西:广西科学技术出版社, 2008:71-72. [10] 卢立华, 何日明, 农瑞红, 等.坡位对灰木莲生长的影响[J].林业科学研究, 2012, 25(6):789-794. doi: 10.3969/j.issn.1001-1498.2012.06.019 [11] 刘玉壶.中国木兰[M].北京:科学技术出版社, 2003:142-143. [12] 招礼军, 韦善华, 朱栗琼, 等.灰木莲的开花特性及繁育系统的研究[J].西部林业科学, 2015(2):24-28. [13] Dafni A. Pollination ecology:a practical approach[M]. Oxford:Oxford University Press, 1992. [14] Mcintosh M E. Flowering phenology and reproductive output in two sister species of Ferocactus (Cactaceae)[J]. Plant Ecology, 2002, 159(1):1-13. doi: 10.1023/A:1015589002987 [15] Herrera J. Flowering and fruiting phenology in the coastal shrublands of Doaña, south Spain[J]. Vegetatio, 1986, 68(2):91-98. [16] 陈香, 胡雪华, 陆耀东, 等.中国特有植物血水草开花物候与生殖特性[J].生态学杂志, 2011, 30(9):1915-1920. [17] 何爽, 张爱勤, 夏荣, 等.新疆不同生态区域苜蓿花粉败育情况及影响因素的细胞学研究[J].草业学报, 2011(4):153-158. [18] 马玉心.兴安鹿蹄草的传粉生态学研究[D].哈尔滨: 东北林业大学, 2007. [19] Ollerton J, Diaz A. Evidence for stabilising selection acting on flowering time in Arum maculatum (Araceae):the influence of phylogeny on adaptation[J]. Oecologia, 1999, 119(3):340-348. doi: 10.1007/s004420050794 [20] Rathcke B, Lacey E P. Phenological patterns of terrestrial plants[J]. Annual Review of Ecology and Systematics, 1985, 16(ⅹ):179-214. [21] 李新蓉, 谭敦炎, 郭江.迁地保护条件下两种沙冬青的开花物候比较研究[J].生物多样性, 2006, 14(3):241-249. doi: 10.3321/j.issn:1005-0094.2006.03.007 [22] 郭连金, 李梅, 林盛.香果树种群开花物候、生殖构件特征及其影响因子分析[J].林业科学研究, 2015, 28(6):788-796. doi: 10.3969/j.issn.1001-1498.2015.06.005 [23] 赵志刚, 程伟, 郭俊杰, 等.西南桦花序生长和开花物候特征[J].林业科学研究, 2011, 24(3):385-389. [24] Chaves ó M, Avalos G. Is the inverse leafing phenology of the dry forest understory shrub Jacquinia nervosa(Theophrastaceae) a strategy to escape herbivory[J]. Revista de biología tropical, 2006, 54(3):951-963. [25] Stephenson A G. Flower and fruit abortion:proximate causes and ultimate functions[J]. Annual review of ecology and systematics, 1981, 12(1):253-279. doi: 10.1146/annurev.es.12.110181.001345 [26] Pías B, Guitián P. Breeding system and pollen limitation in the masting tree Sorbus aucuparia L.(Rosaceae) in the NW Iberian Peninsula[J]. Acta Oecologica, 2006, 29(1):97-103. doi: 10.1016/j.actao.2005.08.005 [27] Arnold A E, Mejía L C, Kyllo D, et al. Fungal endophytes limit pathogen damage in a tropical tree[J]. Proceedings of the National Academy of Sciences, 2003, 100(26):15649-15654. doi: 10.1073/pnas.2533483100 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4558
- HTML全文浏览量: 1823
- PDF下载量: 283
- 被引次数: 0
