



-
油松毛虫(Dendrolimus punctatus tabulaeformis Tsai et Liu)作为我国重要的森林害虫,具有周期性爆发的特点,对生态安全以及林业可持续发展造成了严重威胁[1]。昆虫行为学的充分认知是制定害虫监测和防治策略的基础,现有油松毛虫监测方法就是利用幼虫上、下树行为和成虫召唤行为进行的。由于绝大多数昆虫种类的初孵幼虫活动能力较弱,其存活有赖于雌成虫对产卵寄主植物的明智选择[2],而幼虫的取食偏嗜行为又是评价昆虫对某种寄主接受程度的一个重要指标[3-5]。因此,在昆虫行为学研究过程中,研究其取食和产卵偏嗜行为,可为探讨昆虫对寄主嗜好的程度、适应机制及其综合防治提供理论依据和参考[2, 6]。但是油松毛虫初孵幼虫取食和成虫产卵的偏嗜行为却研究较少。
明确昆虫寄主选择机理是制定行为调控的重要理论基础。昆虫可以通过味觉感受器感知寄主植物中的内含物,从而进行寄主植物的选择和适应。寄主叶片含水量是阻止昆虫进化的主要因素,它可以直接或间接影响昆虫的生长[7]。昆虫取食不同的植物或是植物因为一些因素发生的营养物质的质与量的变化都能在昆虫的生长发育过程中表现出来,而在众多的营养物质中,可溶性糖和可溶性蛋白是昆虫的基本营养成分[8-9]。酚类等化学防御物质能够对昆虫的味觉产生刺激,使昆虫不能完成某一发育阶段,对昆虫有毒害或阻止其进食的作用。李兆麟等采用定株、定期取样的方式指出油松针叶内氨基酸、总糖和酚类物质的含量随季节波动明显,是影响油松毛虫生长发育的主要因素[10]。陈晨研究不同林分类型对油松诱导抗虫性影响时,指出油松受害植株还原糖、蛋白质含量均低于未受害油松植株。不同时间和不同林分的油松针叶内含物存在显著差异,并且能够影响油松毛虫正常的生长发育[11]。但不同部位油松针叶内含物是否有差异及其对油松毛虫寄主选择的影响尚不清楚。
本研究通过室内模拟试验,明确油松毛虫的产卵和初孵幼虫取食偏好,结合寄主针叶中内含物含量的测定,分析影响油松毛虫成虫产卵和初孵幼虫取食选择的影响因子。以期弥补油松毛虫行为学研究的空缺,明确油松毛虫行为选择的诱因,为了解油松毛虫的发生动态、危害程度和制定害虫监测防治策略提供依据。
-
在辽宁省凌源市上樱桃沟村采集油松毛虫上树期幼虫,带回实验室利用油松针叶和油松盆栽幼苗饲养至化蛹。将刚羽化的成虫作为本研究中成虫产卵选择试验的供试虫源;成虫经过羽化、交尾和产卵后,将卵收集起来于室温下自然孵化,初孵的油松毛虫幼虫作为本研究中幼虫取食部分的供试虫源。
-
将4株生长状况良好、针叶茂盛、5年生的盆栽油松幼苗(株高1.2 m)均匀放置于边长1.5 m的正方体养虫笼(120目尼龙网)中,将刚羽化的雌虫和雄虫各5头,置于养虫笼中让其自由交尾、产卵。在成虫死亡后,统计油松一年生针叶、二年生针叶和三年生针叶上的卵块数,尼龙网上的卵块数忽略不计。试验重复10次。
-
在直径150 mm的培养皿底部铺设一层滤纸,在培养皿中间画一个直径5 cm的同心圆作为幼虫取食的未选择区,将环形区平均分为3部分作为幼虫取食的选择区(如图 1所示)。将3种不同的油松针叶(一年生针叶、二年生针叶和三年生针叶)随机置于选择区内,选取活性一致的初孵幼虫(20头)置于未选择区。在全黑暗条件下,让初孵幼虫自由爬行选择。爬出“未选择区”,即认定初孵幼虫进行了取食选择,24 h后统计3种油松针叶选择区内油松毛虫数量。试验重复6次。
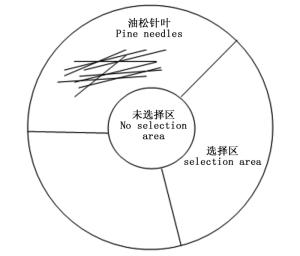
图 1 幼虫取食选择试验培养皿底面示意图
Figure 1. The bottom view of culture dish used in larvae feeding choice experiment
利用一年生油松针叶、二年生油松针叶和三年生油松针叶饲喂油松毛虫初孵幼虫。在直径150 mm的培养皿底部铺设一层滤纸,将油松针叶均匀的平铺在培养皿中,每个培养皿接虫10头,每2天更换一次油松针叶。5天后统计油松毛虫幼虫体长,30天后(油松毛虫进入三龄期)统计油松毛虫幼虫死亡率。试验重复5次。
-
试剂:40% NaOH、20% NaOH、4% H3BO3储备液、4% H3BO3吸收液、重铬酸钾标准溶液、0.2 mol·L-1硫酸亚铁溶液、N-苯基邻胺基苯甲酸、浓硫酸、硫酸银、没食子酸、Folin试剂、20% Na2CO3、乙醇、催化剂片(硫酸钾:硫酸铜=10:1)等。
仪器:烘箱、粉碎机、油浴锅、凯氏消化炉(DK20 Heating Digester意大利Velp公司)、全自动凯氏定氮仪(型号:UK152 Distillation & Titration Unit意大利Velp公司)等。
-
从油松树上随机采集一年生针叶、二年生针叶和三年生针叶。将样品称质量后放入105℃的烘箱内杀青30 min,然后转移至70℃烘箱内烘烤至恒质量。计算针叶含水量。烘干至恒质量的油松针叶利用粉碎机粉碎,过60 mm筛,筛样用于测定针叶内总酚、有机氮和有机碳含量。
-
油松针叶总酚含量测定采用Folin-Cioealteu比色法[12]。
样品溶液制备:称取1.000 g油松针叶粉,置于250 mL锥形瓶中,加入100 mL无水乙醇,微波震荡提取2 h。将样液转移到100 mL离心管中,3 500转离心5 min。
总酚标准曲线:精密称取没食子酸标准品0.010 0g,用蒸馏水溶解并定容至100 mL,得到浓度为0.10 mg·mL-1的标准液。准确吸取0、0.4、0.6、0.8、1.0、1.2、1.4、1.6 mL置于25 mL的容量瓶中,加蒸馏水至6.0 mL,然后分别加入FC试剂(福林试剂)0.5 mL,混匀,在0.58 min内分别加入1.5 mL20%的Na2CO3溶液,充分混合后定容,30℃避光放置0.5 h,以不加标准液的6.0 mL蒸馏水为空白对照,760 nm下测定吸光值,每个样品平行测定4次。以没食子酸在反应体系中的质量浓度为横坐标,吸光度值为纵坐标,绘制标准曲线。
总酚含量的测定:准确量取制备好的乙醇粗提液0.5 mL于50 mL容量瓶中,加入9.5 mL蒸馏水,摇匀,再加入0.5 mL的FC试剂,混匀,在0.58 min内加入1.5 mL 20%的Na2CO3溶液,充分混合后定容,30℃避光放置0.5 h,以没食子酸标准液为空白对照,760 nm下测定吸光值,每个样品平行测定4次。
-
油松针叶有机氮含量测定采用凯氏定氮法测定。
准确称取样品0.50 g(70℃烘干,过筛60目,称量范围0.500 00.505 0 g)于消解管底部,加入2粒混合催化剂片(硫酸钾:硫酸铜=10:1),再慢慢加入15 mL H2SO4,用少量水冲洗消化管壁后,放置在专用消化管架上,静置过夜于通风柜中,第二天在DK20凯氏消化炉中消解,消解结束冷却后,使用UK152全自动凯氏定氮仪测定并记录滴定体积和全N含量。
-
油松针叶有机碳含量测定采用重铬酸钾反滴定法测定。
准确称取样品0.50 g于硬质大试管中,加粉末状硫酸银0.1 g,用吸管加入5 mL的0.800 0 mol·L-1重铬酸钾标准溶液,然后用注射器注入5 mL浓硫酸,并小心旋转摇匀。将大试管于油浴锅中170180℃沸腾5 min。加入N-苯基邻胺基苯甲酸指示剂,样品颜色由棕红色经紫至蓝色为滴定终点。,记录硫酸亚铁用量。每批分析时,做3个空白标定,空白标定加入0.5 g石英砂,其他步骤与测定样品是完全相同,记录硫酸亚铁用量。
-
采用EXCEL和SPSS17.0对数据进行单因素方差分析和相关性分析。
-
油松毛虫对不同的油松针叶表现出明显的产卵选择偏嗜现象(图 2)。一年生油松针叶上的卵块数最多(3.10±0.28),占全部卵块数的70%以上,明显高于产在二年生针叶(1.30±0.30)和三年生针叶(0.20±0.13)上的卵块数。由此可知,油松毛虫偏向于将卵产在一年生油松针叶上。
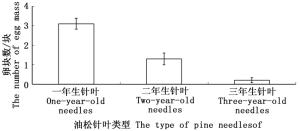
图 2 油松毛虫在不同油松针叶上的产卵量
Figure 2. The number of egg mass laid on different years of pine needles
-
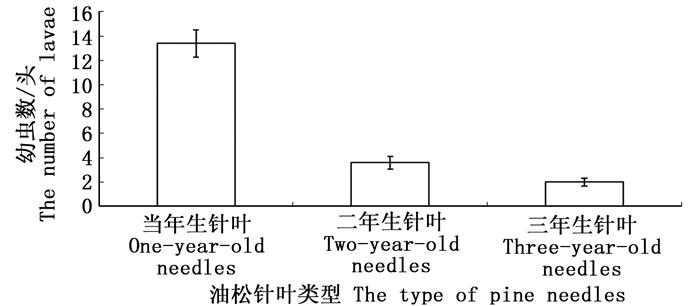
初孵的油松毛虫幼虫对不同油松针叶表现出明显的取食偏嗜行为,油松毛虫初孵幼虫偏嗜于取食一年生油松针叶(图 3)。幼虫取食选择24 h后,选择在一年生油松针叶上取食的初孵幼虫数达到13.40±0.91头,显著高于取食二年生油松针叶的3.60±0.42头和取食三年生油松针叶的2.00±0.26头初孵幼虫数量。
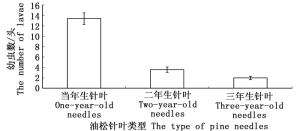
图 3 在不同油松针叶上取食的幼虫数量
Figure 3. The number of larvae feeding on different types of pine needles
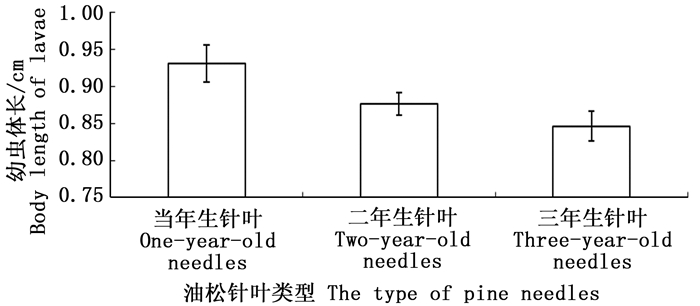
取食不同油松针叶的油松毛虫初孵幼虫生长发育速度表现出明显差异(图 4)。取食一年生油松针叶的初孵幼虫5天后体长达到0.93±0.025 cm;高于取食二年生针叶的油松毛虫体长0.88±0.015 cm;显著高于取食三年生针叶的油松毛虫体长0.85±0.020 cm。取食不同油松针叶30天后(油松毛虫生长到三龄幼虫,即幼虫下树时的龄期),油松毛虫幼虫死亡率表现出显著不同(图 5)。取食一年生油松针叶的初孵幼虫死亡率最低为13.60%±4.87%;取食二年生油松针叶的初孵幼虫死亡率最高为40.00%±9.49%;其次为取食三年生油松针叶的初孵幼虫,死亡率为30.00%±5.48%。
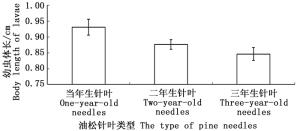
图 4 取食不同油松针叶5天后幼虫体长
Figure 4. The body length of larvae feeding on different types of pine needles after 5 days

图 5 取食不同油松针叶的幼虫死亡率
Figure 5. The death rate of larvae feeding on different types of pine needles
-
测定不同油松针叶的内含物,结果如表 1所示:不同的油松针叶中各内含物含量不同,并且表现出显著差异。一年生油松针叶内含水量最高,总酚含量最低,有机碳含量最低。一年生油松针叶中含水量达到62.43%±0.38%,明显高于二年生针叶(55.95%±0.73%)和三年生针叶(54.07%±0.80%);一年生油松针叶内总酚含量仅有9.11±0.61 g·100 g-1,低于二年生油松针叶的12.35±1.18 g·100 g-1,明显低于三年生油松针叶的16.20±1.98 g·100 g-1;一年生针叶内有机碳含量最低55.42±0.32 g·100 g-1,显著低于二年生针叶和三年生针叶;但不同油松针叶内有机氮含量和C/N比并没有显著差异。
表 1 油松针叶内含物含量
Table 1. The content of different inclusions in pine needles
项目
Items含水量
Water content/%总酚
Total phenols/(g·100 g-1)有机碳
C/(g·100 g-1)有机氮
N/(g·100 g-1)碳氮比
C/N一年生针叶One-year-old needles 62.43±0.38 a 9.11±0.61 a 55.42±0.32 a 1.36±0.02 a 40.81±0.97 a 二年生针叶Two-years-old needles 55.95±0.73 b 12.35±1.18 ab 58.30±0.37 b 1.57±0.12 a 37.90±3.06 a 三年生针叶Three-years-old needles 54.07±0.80 b 16.20±1.98 b 59.78±0.33 c 1.41±0.10 a 42.93±2.99 a 注:表中所有数值均为平均值±标准误。同列不同字母表示差异显著(含水量:F = 44.023; df = 2, 9; p = 0.000, ANOVA;总酚:F = 6.651; df = 2, 9; p = 0.017, ANOVA;有机C:F = 42.571; df = 2, 9; p = 0.000, ANOVA;有机N:F = 1.410; df = 2, 9; p = 0.291, ANOVA;C/N:F = 0.994; df = 2, 9; p = 0.407, ANOVA)。
Note: Data in the table are the means ± SE. Different letters in column indicate significant differences (Water content:F = 44.023; df = 2, 9; p = 0.000, ANOVA; Total phenols:F = 6.651; df = 2, 9; p = 0.017, ANOVA; C:F = 42.571; df = 2, 9; p = 0.000, ANOVA; N:F = 1.410; df = 2, 9; p = 0.291, ANOVA; C/N:F = 0.994; df = 2, 9; p = 0.407, ANOVA). -
对3种油松针叶内含物与油松毛虫成虫产卵选择、初孵幼虫取食选择和幼虫体长进行了相关性分析(表 2)。结果表明:油松毛虫初孵幼虫取食选择、成虫产卵选择和幼虫体长与油松针叶内含水量成极显著正相关(P < 0.01),Pearson相关性系数均在0.80以上;油松毛虫初孵幼虫取食选择、成虫产卵选择和幼虫体长与针叶内总酚含量成极显著负相关(P < 0.01),Pearson相关性系数均在-0.80以下;初孵幼虫取食选择和幼虫体长与碳氮比成极显著负相关(P < 0.01),成虫产卵选择与碳氮比成显著负相关(P < 0.05)。油松毛虫成虫产卵选择与初孵幼虫取食选择和幼虫体长之间极显著性相关(P < 0.01),并且Pearson相关系数均在0.80以上。
表 2 油松针叶内含物含量与油松毛虫行为选择相关系数
Table 2. Correlation coefficient for the content of inclusions and the behavior choice
项目Items 含水量Water content 总酚Total phenols 有机碳C 有机氮N 碳氮比C/N 幼虫取食选择Feeding choice 0.950** -0.939** -0.159 -0.153 -0.734** 成虫产卵选择Oviposition selection 0.827** -0.884** -0.094 -0.214 -0.692* 幼虫体长Larvae length 0.813** -0.827** -0.159 -0.132 -0.715** 注:*表示在0.05水平上显著相关;**表示在0.01水平上显著相关。
Note: *significant at 0.05 level; ** significant at 0.01 level.对油松针叶内含水量、总酚含量、有机碳含量、有机氮含量和碳氮比进行相关性分析(表 3)。结果表明:油松针叶内含水量与总酚含量、碳氮比成极显著负相关(P < 0.01),Pearson相关性系数分别为-0.920和-0.851,即油松针叶内含水量越高,总酚含量越低,C/N越低;油松针叶内总酚含量与C/N成显著正相关,相关系数为0.724,即总酚含量越高油松针叶内C/N越高。
表 3 油松针叶各内含物之间的相关系数
Table 3. Correlation coefficient for the different inclusions in pine needles
项目Items 含水量Water content 总酚Total phenols 有机碳C 有机氮N 碳氮比C/N 含水量Water content 1 总酚Totol phenols -0.920** 1 有机碳C -0.007 0.112 1 有机氮N -0.298 0.208 -0.943** 1 碳氮比C/N -0.851** 0.724** -0.401 0.648* 1 注:*表示在0.05水平上显著相关;**表示在0.01水平上显著相关。
Note: *significant at 0.05 level; ** significant at 0.01 level. -
本研究对油松毛虫的初孵幼虫取食和成虫产卵行为进了初步研究。结果表明:油松毛虫成虫产卵和初孵幼虫取食均偏嗜一年生油松针叶。这与《中国松毛虫》一书中提到的油松毛虫一般取食两年生针叶的理论相悖[13],也与张雄帅利用二年生油松针叶饲喂油松毛虫相矛盾[14]。这可能是由于一年的不同时期造成的,上树期(4月)油松毛虫主要取食二年生针叶,初孵幼虫期(8月初)油松毛虫主要取食一年生针叶,这与陈晨利用一年生针叶饲喂初孵幼虫而利用二年生针叶饲喂上树期幼虫的结论一致[11]。因此,我们推断油松毛虫成虫产卵后(8月初)到下树前(10月初)主要活动范围是在油松一年生针叶上。现有监测方法监测油松毛虫时间为3—4月(上树期)、6—8月(成虫羽化期)、和10—2月(下树期和越冬期)[15-18],而4—6月和8—10月(幼虫活动期)并没有很好的监测手段。根据本实验研究结果(油松毛虫幼虫在4龄前主要在一年生针叶上活动),在以后的油松毛虫监测过程中,在8—10月可以重点对油松一年生针叶进行了调查,根据调查结果适时进行防治。
不同时间和不同林分的油松针叶内含物含量存在显著变化。李兆麟等研究结果表明油松针叶内氨基酸、总糖和酚类物质的含量随季节波动明显,是影响油松毛虫生长发育的主要因素[10];陈晨研究不同林分类型对油松诱导抗虫性影响时,指出不同林分油松受害植株还原糖、蛋白质含量差异显著[11]。不同时间和不同林分的油松针叶内含物存在显著差异,并且能够影响油松毛虫正常的生长发育。本研究首次对不同部位油松针叶内含物进行了测定并与油松毛虫成虫产卵选择、幼虫取食选择和幼虫体长进行了相关性分析。结果表明:油松毛虫幼虫取食选择、成虫产卵选择和幼虫体长与油松针叶内含水量、总酚含量和碳氮比具有显著相关性。油松毛虫成虫产卵和幼虫取食均偏嗜于一年生针叶,可能是由于一年生油松针叶内含水量高、总酚含量低、有机碳含量低造成的。由于油松毛虫缺少人工饲料,所以只能推测油松毛虫产生偏嗜行为的原因可能是含水量、总酚和碳氮比。在油松毛虫人工饲料问世后,可以人为的控制这几种物质饲喂油松毛虫,直接证明其对油松毛虫产卵取食选择的影响。
湿度是影响油松毛虫种群数量的主要气象因子。种群动态预测模型指出湿度是影响油松毛虫种群动态的主要气象因子。例如:段君博等研制的油松毛虫实时动态预测预报系统中每月的平均温度和湿度是发生量预测预报的基础[19]。屠泉洪等研究表明影响越冬幼虫种群数量变动的关键因子主要是越冬幼虫生境的温度和相对湿度[20]。周广学等和宋雄刚等均指出夏季均湿、10月均湿与油松毛虫的种群发生相关[21-23]。本研究结果表明:含水量与总酚含量、碳氮比成极显著负相关,而含水量、总酚含量和C/N与油松毛虫的生长发育成显著相关。谢冬梅和董文宇[24]研究证明油松针叶含水量与土壤含水量之间也存在显著相关。因此,降雨量越大、土壤含水量越高、油松针叶含水量越高越有利于油松毛虫生长发育。从试验角度直接证明了宋雄刚等人的研究结果。
害虫行为调节中包括三个主要因素:害虫的行为、调节其行为的手段和实际应用方法。本研究结果证明油松毛虫具有偏嗜行为,但并没有提出具体的调节行为的手段和实际应用方法,这可能是以后研究的方向。本研究只针对初孵幼虫取食期不同部位油松针叶内含物含量差异进行了测定,对于一年中不同时期、不同林分油松上不同类型的油松针叶内含物有无差异并未研究。
-
油松毛虫对一年生油松针叶具有明显的产卵、取食偏嗜行为。油松毛虫在一年生油松针叶上平均产卵块数最多,显著高于二年生针叶和三年生针叶;取食一年生油松针叶的油松毛虫初孵幼虫数量最多、死亡率最低,显著优于取食二年生油松针叶和三年生针叶的初孵幼虫。油松针叶内含物含量与油松毛虫行为选择相关性分析结果表明初孵幼虫取食选择、成虫产卵选择和幼虫体长与油松针叶内含水量成显著正相关;与寄主总酚含量和碳氮比成显著负相关。因此,推断一年生油松针叶内含水量高、总酚含量和有机碳含量低可能是造成寄主偏嗜行为的重要原因。
油松毛虫产卵、取食偏嗜行为及油松针叶内含物测定
Feeding and Oviposition Preferences of Pine Caterpillar and Inclusions of Chinese Pine Needles
-
摘要:
目的 为明确油松毛虫在油松上的产卵和取食部位以及可能机理, 为未来害虫发生预测模拟提供基础信息数据。 方法 通过室内模拟试验, 统计油松毛虫在盆栽油松幼苗不同部位上的产卵量和取食不同油松针叶的偏嗜情况及死亡率; 测定油松针叶内含物含量并与油松毛虫偏嗜行为进行了相关性分析。 结果 在一年生油松针叶上平均产卵块数最多(3.10±0.28), 显著高于二年生针叶和三年生针叶; 取食一年生油松针叶的油松毛虫数量最多(13.40±0.91)、死亡率最低(13.60%±4.87%), 显著优于取食二年生油松针叶和三年生油松针叶的幼虫。油松毛虫幼虫取食选择、成虫产卵选择和幼虫体长与油松针叶内含水量成显著正相关(P < 0.01);与寄主总酚含量(P < 0.01)和碳氮比(P < 0.05)成显著负相关。 结论 油松毛虫对一年生油松针叶具有明显的产卵和取食偏嗜行为。一年生油松针叶内含水量最高、总酚含量最低、有机碳含量最低可能是造成寄主偏嗜行为的原因。 Abstract:Objective The aim of this study was to know the different preferences on different parts of Chinese pine by the pine caterpillar and the possible mechanisms which provides the basis information data for the future pest forecast simulation. Methods According to the indoor simulated experiments, the author analyzed the amount of egg mass laid on different types of pine needles by the pine caterpillar adult, the feeding preference and death rate of larvae feeding on different types of pine needles. We tested the content of inclusions in pine needles and analyzed the relationship between the preference and the content of pine needles. Results The results showed that the number of egg mass laid on One-year-old pine needles was 3.10±0.28 which was significantly higher than Two-years-old pine needles and Three-years-old pine needles. The amount of larvae feeding on one-year-old pine needles was the biggest (13.40±0.91) and the death rate was the lowest (13.60%±4.87%), which were significantly better than feeding on Two-years-old pine needles and three-years-old pine needles. The paper results indicated that the pine caterpillar had feeding and oviposition preferences. The preference behaviors were significant positive correlation on water content (P < 0.01), and negative correlation on the contents of total phenol (P < 0.01) and C/N (P < 0.05) of these three kinds of pine needles. Conclusion The pine caterpillar preferred to feed and oviposition on one-year-old pine needles. The reason of the preference behaviors maybe was the higher water content, the lower total phenol and carbon content of One-year-old pine needles. -
Key words:
- Dendrolimus punctatus tabulaeformis
- / oviposition
- / feeding
- / behavior preference
- / inclusions
-
图 1 幼虫取食选择试验培养皿底面示意图
Figure 1. The bottom view of culture dish used in larvae feeding choice experiment
图 2 油松毛虫在不同油松针叶上的产卵量
Figure 2. The number of egg mass laid on different years of pine needles
图 3 在不同油松针叶上取食的幼虫数量
Figure 3. The number of larvae feeding on different types of pine needles
图 4 取食不同油松针叶5天后幼虫体长
Figure 4. The body length of larvae feeding on different types of pine needles after 5 days
图 5 取食不同油松针叶的幼虫死亡率
Figure 5. The death rate of larvae feeding on different types of pine needles
表 1 油松针叶内含物含量
Table 1. The content of different inclusions in pine needles
项目
Items含水量
Water content/%总酚
Total phenols/(g·100 g-1)有机碳
C/(g·100 g-1)有机氮
N/(g·100 g-1)碳氮比
C/N一年生针叶One-year-old needles 62.43±0.38 a 9.11±0.61 a 55.42±0.32 a 1.36±0.02 a 40.81±0.97 a 二年生针叶Two-years-old needles 55.95±0.73 b 12.35±1.18 ab 58.30±0.37 b 1.57±0.12 a 37.90±3.06 a 三年生针叶Three-years-old needles 54.07±0.80 b 16.20±1.98 b 59.78±0.33 c 1.41±0.10 a 42.93±2.99 a 注:表中所有数值均为平均值±标准误。同列不同字母表示差异显著(含水量:F = 44.023; df = 2, 9; p = 0.000, ANOVA;总酚:F = 6.651; df = 2, 9; p = 0.017, ANOVA;有机C:F = 42.571; df = 2, 9; p = 0.000, ANOVA;有机N:F = 1.410; df = 2, 9; p = 0.291, ANOVA;C/N:F = 0.994; df = 2, 9; p = 0.407, ANOVA)。
Note: Data in the table are the means ± SE. Different letters in column indicate significant differences (Water content:F = 44.023; df = 2, 9; p = 0.000, ANOVA; Total phenols:F = 6.651; df = 2, 9; p = 0.017, ANOVA; C:F = 42.571; df = 2, 9; p = 0.000, ANOVA; N:F = 1.410; df = 2, 9; p = 0.291, ANOVA; C/N:F = 0.994; df = 2, 9; p = 0.407, ANOVA). 下载: 导出CSV
下载: 导出CSV
表 2 油松针叶内含物含量与油松毛虫行为选择相关系数
Table 2. Correlation coefficient for the content of inclusions and the behavior choice
项目Items 含水量Water content 总酚Total phenols 有机碳C 有机氮N 碳氮比C/N 幼虫取食选择Feeding choice 0.950** -0.939** -0.159 -0.153 -0.734** 成虫产卵选择Oviposition selection 0.827** -0.884** -0.094 -0.214 -0.692* 幼虫体长Larvae length 0.813** -0.827** -0.159 -0.132 -0.715** 注:*表示在0.05水平上显著相关;**表示在0.01水平上显著相关。
Note: *significant at 0.05 level; ** significant at 0.01 level.
下载: 导出CSV
表 3 油松针叶各内含物之间的相关系数
Table 3. Correlation coefficient for the different inclusions in pine needles
项目Items 含水量Water content 总酚Total phenols 有机碳C 有机氮N 碳氮比C/N 含水量Water content 1 总酚Totol phenols -0.920** 1 有机碳C -0.007 0.112 1 有机氮N -0.298 0.208 -0.943** 1 碳氮比C/N -0.851** 0.724** -0.401 0.648* 1 注:*表示在0.05水平上显著相关;**表示在0.01水平上显著相关。
Note: *significant at 0.05 level; ** significant at 0.01 level.
下载: 导出CSV
-
[1] 宋雄刚.油松毛虫发生林分因子分析及气候情景模式下的灾害趋势预测[D].北京: 中国林业科学研究院, 2015 [2] Courtney S P, Chen G K, Gardner A. A general model for individual host selection[J]. Oikos, 1989, 55:55-56 doi: 10.2307/3565872 [3] 王争艳, 李心田, 鲁玉杰, 等.昆虫寄主选择行为的进化机制[J].应用昆虫学报, 2011(1):174-177 [4] Thompson J N, Pellmyr O. Evolution of oviposition behavior and host preference in Lepidoptera[J]. Annu Rev Entomol, 1991, 36:65-89 doi: 10.1146/annurev.en.36.010191.000433 [5] Thompson J N. Evolutionary ecology of the relationship between oviposition preference and performance of offspring in phytophagous insects[J]. Entomol Exp Appl, 1988, 47(1):3-14 doi: 10.1111/eea.1988.47.issue-1 [6] Singer M C, Lee J R. Discrimination within and between host species by a butterfly:implications for design of preference experiments[J]. Ecol Lett, 2000, 3:101-105 doi: 10.1046/j.1461-0248.2000.00121.x [7] 康乐.环境胁迫下的昆虫-植物相互关系[J].生态学杂志, 1995(5):51-57 [8] 钦俊德.昆虫与植物关系的研究进展和前景[J].动物学报, 1995(1):12-20 doi: 10.3321/j.issn:0001-7302.1995.01.008 [9] 钦俊德.植食性昆虫食性的生理基础[J].昆虫学报, 1980(1):106-122 [10] 李兆麟, 贾凤友, 何忠, 等.油松毛虫滞后发育与寄主的关系[J].林业科学研究, 1992, 5(6):671-674 [11] 陈晨.林分类型对油松诱导抗虫性的影响[D].河北保定: 河北农业大学, 2012 [12] 张毛莉, 罗仓学.石榴皮中总酚含量测定方法的比较[J].食品工业科技, 2011(5):383-384 [13] 侯陶谦.中国松毛虫[M].北京:科学出版社, 1989 [14] 张雄帅.油松毛虫体内酶系对油松不同诱导抗性的响应机制研究[D].河北保定: 河北农业大学, 2013 [15] 王小军.油松毛虫测报及无公害防治技术的应用[J].中国森林病虫, 2006(1):35-39 doi: 10.3969/j.issn.1671-0886.2006.01.013 [16] 何善勇, 骆有庆, 温俊宝, 等.气候变暖对油松毛虫幼虫越冬及上下树发生期的影响[J].应用昆虫学报, 2012(5):1231-1242 [17] 刘奎伟.油松毛虫性信息素的结构鉴定和诱芯载体中释放速率的研究[D].泰安: 山东农业大学, 2012 [18] 刘银菊, 付书英, 王冰.性信息素在油松毛虫防治中的应用[J].河北林业科技, 2009(5):41 doi: 10.3969/j.issn.1002-3356.2009.05.022 [19] 段君博, 赵殿一, 孙向文, 等.油松毛虫实时动态预测预报PDFS系统的研制[J].辽宁林业科技, 1997(1):32-35 [20] 屠泉洪, 夏乃斌, 邵海荣.油松毛虫发生的灰色预测模型[J].生态学报, 1990(3):261-265 [21] 宋雄刚, 王鸿斌, 李国宏, 等.区域尺度油松毛虫灾害发生相关气象因子筛选[J].东北林业大学学报, 2015, 43(7):127-132 doi: 10.3969/j.issn.1000-5382.2015.07.028 [22] 周广学, 张国林, 梁群.气象条件对油松毛虫的影响及其预测模型的构建[J].东北林业大学学报, 2012, 40(11):131-134 doi: 10.3969/j.issn.1000-5382.2012.11.032 [23] 宋雄刚, 王鸿斌, 张真, 等.应用最大熵模型模拟预测大尺度范围油松毛虫灾害[J].林业科学, 2016, 52(6):66-75 [24] 谢冬梅, 董文宇.油松春季含水量变化与N P K营养参数相关分析[J].吉林农业, 2012(3):53-55 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3819
- HTML全文浏览量: 1403
- PDF下载量: 301
- 被引次数: 0
