


-
杜鹃花是世界上最著名的观赏花卉之一,因种类繁多、花美色艳而闻名于世。全世界超过1 000余种,起源于距今约6 700万年至13 700万年中生代的白垩纪;广泛分布于亚洲、欧洲和北美洲,但有2种进入北美阿拉斯加和格陵兰东部,形成分布区的北缘;有1种越过赤道达昆士兰,形成分布南界[1-3]。中国约581种,被划分为6个亚属,其中421种为特有种;主要集中于西南地区,为植物群落中常见的伴生种、优势种或建群种;云南、四川、西藏三省的高海拔山地几乎拥有中国杜鹃花种类的90%以上,有“世界杜鹃大花园”之称[3-4]。中国西南的高山、亚高山因有杜鹃花而绚丽多彩。
欧美至今流传着一句名言“没有中国的杜鹃花,就没有西方的园林”。近百年来,比利时、美国、英国、日本等通过从我国西南引进野生种,并与本地种进行杂交培育出了大量极具商品性的杜鹃新品种[5],可见,我国众多的杜鹃种质资源为世界杜鹃花育种做出了重大贡献;而我国杜鹃花育种工作进行得较晚,仅34个品种进行了登记[6]。时至今日,常规杂交育种仍是培育杜鹃花新品种的主要途径,且当前国际上杜鹃花的花色育种趋向于培育纯色花,尤其是明亮的黄色。同时,为迎合中国节日活动布置和年宵花供应所需,培育不同花期的杜鹃品种也是杜鹃花育种者的目标之一[7]。长梗杜鹃(Rhododendron longipedicellatum Lei Cai & Y.P. Ma)因其罕见的花期和纯黄色的花朵为上述育种目标提供了契机,若将其与其他优良种质进行杂交,使有益基因相互渗透,有望创制出更多市场所需的新品种,进而加快国内外杜鹃花育种产业的快速发展。
长梗杜鹃隶属类越桔杜鹃亚组,为1.0~3.3 m高的多分枝常绿灌木;其树形美观、花叶兼美,花梗长达3.0~5.5 cm;花冠鲜黄、无任何斑点。与野生杜鹃花期多集中于3~6月不同,长梗杜鹃的自然花期从11月下旬持续至翌年的2月上旬,历时3个月之久且正值春节和圣诞节[4,8],在市场上有巨大的产业发展前景。然而,其分布区极其狭窄,仅发现残存于麻栗坡县石灰岩生境的5个自然居群,植株数量不足2 000株。根据IUCN Red List Categories and Criteria (version 3.1) [9]属于极危状态[CR B1ab (ⅰ, ⅲ, ⅴ)]。
花粉是遗传信息的载体,其形态特征受核基因的控制,具有很强的遗传保守性,其固有的轮廓、外壁纹饰、萌发孔数目及位置等特征有助于解决某些植物在分类系统上的地位问题[10-11]。杂交亲本的花粉生活力高低、贮藏寿命的长短直接影响着育种的成效。为避免其盲目性,在开展杂交育种之前,对亲本材料的花粉生活力和贮藏特性进行检测尤为必要。测定杜鹃花花粉活力的染色法和液体培养基方法已有一些报道[12-14],而固体培养基法很少见。鉴于此,本研究利用光学显微镜和扫描电子显微镜对滇东南稀有濒危植物长梗杜鹃的花粉形态进行了观察;通过TTC染色法和花粉离体萌发法对长梗杜鹃的花粉生活力进行比较检测,同时研究了其在不同贮藏温度下花粉活力随贮藏时间的变化情况。本研究旨在对长梗杜鹃开花过程中的花粉活力进行快速检测,明确采粉的最佳时期,找出适合测定长梗杜鹃花粉萌发力的最佳固体培养基配方,并了解其花粉适宜的贮藏温度,为今后开展以长梗杜鹃为父本材料的杂交育种积累有用的资料;同时,也为探讨该种的起源演化和亲缘关系提供孢粉学理论依据。
-
采自残存于云南省麻栗坡县的最大野生居群(文保居群, 海拔1 312 m, 104°94′ E, 23°15′ N),于2016年12月分别从15个生长良好的单株上剪下带花蕾或刚开放花朵的3~5个枝条,竖插于放有适量水的桶中带回实验室。置于室温下,每天更换3次蒸馏水使花朵自然开放,直至试验结束。本试验中每个处理所用花粉均分别从15个单株的枝条上收集并充分混匀。
-
取3个不同发育时期(花蕾膨大期、花瓣初开期和花瓣盛开期)的新鲜花粉,分别用蒸馏水清洗2~3次,使花粉粒充分散开。水洗后,部分花粉粒用甘油胶封片,在蔡司Axio Scope.A1显微镜下用油镜(放大1 000X)进行观察和拍照;部分花粉粒经30%至95%酒精逐级脱水,晾干后用双面导电胶带将其固定在样品台上,经喷金镀膜后,在日立高新TM-3000扫描电镜下观察、拍照并记录,并在花瓣盛开期随机选取30粒花粉测量其直径大小、萌发孔长宽及萌发孔相对长度。相对长度 = 萌发孔长度/直径。
-
从水培培养的瓶插花不同枝条上,选取当天开放的10个花朵,从9:00至21:00,每隔2 h取1次花粉,每次从各朵花中选1个雄蕊取花粉,混合后置于凹型载玻片上;滴加1~2滴0.5%TTC的5%蔗糖溶液,迅速盖上盖玻片,置入内有湿润滤纸的培养皿中,连同培养皿放置在室温黑暗条件下2 h左右。在光学显微镜下分别统计载玻片中央3~5个视野里全部观测到的花粉(每个视野不少于50粒)中红色花粉所占的比例,重复3次。
-
在水培培养的瓶插花不同枝条上,用标签纸标记合瓣期(主要指开花前1~2 d)以及开花后不同时间(1~9 d)的花朵45朵。每天9:00选取5朵花,每朵花选3~5个雄蕊,混匀后测定其花粉活力,具体方法同上。
花粉活力 = 每个视野被染红的花粉数/观察到的花粉总数×100%
-
参照张超仪等[12]的方法,利用L25(53)正交试验探讨10 g·L-1琼脂、不同浓度蔗糖(0、50、100、150、300 g·L-1)、H3BO3 (0、50、100、200、300 mg·L-1)及CaCl2 (0、50、150、250、350 mg·L-1)等体积混合组成的培养基配方对长梗杜鹃花粉萌发的影响,找出适宜的固体培养基。具体方法为:于早上9:00收集不同植株当天开放花朵的新鲜花粉充分混匀,用毛笔将其均匀的撒播在含有0.1 mL以上培养基的双孔凹型载玻片上,盖上盖玻片,放入铺有3层湿润滤纸的培养皿内,并置于(25±1)℃的光照培养箱中。每隔4 h在低倍显微镜下观察1次,并适当补充相应的培养基成分,发现花粉萌发后,每隔1 h观察1次,最终结束时间以2次观察到的萌发率基本相同为准。每次观察从载玻片中央选取3~5个清晰视野(每个视野不少于50粒花粉)统计萌发率,3次重复。以花粉管长度等于或超过花粉粒直径、四合体只要有1个单花粉粒萌发即视为已萌发。
萌发率 = 某视野中已萌发的花粉粒数/该视野中花粉粒总数×100%
-
于花朵开放当天9:00 am收集15个不同植株的新鲜花粉充分混匀,装于1 mL冻存管中,放入适量干燥剂,并贴好标签,分别置于室温(20~25℃)、4℃和-18℃低温冰箱中保存。室温保存的花粉每隔2 d检测1次萌发力变化情况,4℃和-18℃贮藏的花粉每隔6 d检测1次,直至萌发力低于5%,基本不能再用于杂交授粉为止。检测方法用1.4节中筛选出的最适培养基培养17 h,并统计花粉的萌发情况,4个重复。
-
通过光学显微镜和扫描电镜观察,长梗杜鹃的花粉为复合型四合体,呈正四面体(即极面观:1粒在上3粒在下或3粒在上1粒在下)或十字形(即赤道面观:2粒在上2粒在下)排列。四合体直径43.0~65.4 μm,平均直径51.3 μm; 花粉表面具粘丝,单粒花粉近球形,均具3个子午向的孔沟,沟长9.03~23.9 μm,相对长为0.15~0.51,平均沟长和平均相对长分别为13.9 μm和0.27;沟宽3.6~8.6 μm,平均5.9 μm。相邻花粉分体上的孔沟相连,并且相通,沟通常与相邻花粉粒紧靠的一端平,向着花粉粒极面的一端尖。孔沟边缘呈加厚,沟内内含物外露,花粉分体间区为均匀的颗粒状同外壁,孔沟周围颗粒细密呈波状突起,与外壁纹饰区别明显。
花朵不同发育时期在扫描电镜下的花粉结构特征存在一定差异,而光镜下无明显不同。花蕾膨大期:四合花粉初具形状,花粉分体中部凹陷,三萌发沟可见雏形;四合体间缝浅,花粉分体间区为粗糙的聚颗粒状,有清晰裂纹;沟周围纹饰不明显(图 1: Ⅰ-1、2、3)。花瓣初开期:花粉形状趋于清晰,呈正四面体排列时为锐角三角形,单花粉粒椭圆形,四合体间缝较浅,萌发沟内出现疏松的内含物;花粉分体间区为均匀的粗颗粒状,沟周围开始加厚(图 1: Ⅱ-1、2、3)。花瓣盛开期:花粉成熟,形状饱满,呈正四面体排列时为圆角三角形;单花粉粒近球形,四合体间缝深,萌发沟内充满内含物。花粉分体间区为均匀的细颗粒状,沟周围加厚且呈波状突起(图 1: Ⅲ-1、2、3)。

图 1 长梗杜鹃花朵不同发育时期的花粉形态
Figure 1. Pollen grains morphology of R. longipedicellatum at different development stages
光学显微镜下观察到长梗杜鹃花粉外壁为内外2层,厚度约相等为0.8~1.2 μm, 表面可见模糊的细网状雕纹(图 1: Ⅳ-1、2、3)。
-
通过TTC(氯化三苯基四氮唑)染色法能使长梗杜鹃新鲜花粉在短时间内快速着色,且染色效果明显,容易区分。花粉活力检测结果见图 2。从图 2A可看出:长梗杜鹃开花当天从9:00—17:00花粉活力随着开花时间的延长逐渐递减,以9:00最高,达92.18%;17:00最低,为42.85%;17:00之后有缓慢上升的趋势。从图 2B可看出:长梗杜鹃合瓣期(开花前1~2 d)的花粉活力相对较高,为78.20%,但显著低于开花当天(Student’s t-test, t = 22.54; P<0.001);之后,随着花粉从花粉囊散出其活力逐渐下降,直至花末期(花后第9天)仍保持着较高的花粉活力(48.5%),约为最高花粉活力的一半。由此可见,整个单花花期长梗杜鹃能维持较高的花粉活力,有利于后期相关杂交育种工作的开展。

图 2 长梗杜鹃花朵开放期间的花粉活力变化情况
Figure 2. Change of pollen viability at anthesis of R. longipedicellatum
-
试验观察发现:大部分培养基上的花粉培养4 h后已开始萌发,培养9 h后,2号和6号培养基上的花粉萌发量达最大,而22号和25号培养基直到26 h后花粉萌发数才达到稳定。L25(53)正交试验各培养基组合的花粉萌发率差异较大(表 1),以100 g·L-1蔗糖+ 200 mg·L-1 H3BO3+ 0 mg·L-1 CaCl2培养基配方培养17 h的萌发率最高(达90.26%)。对表 1中的数据进行极差分析,表明3因素的主次顺序为:蔗糖→ CaCl2→ H3BO3,极差分别为2.36, 0.92和0.88,相对应的最佳蔗糖因子为A3 (k3 = 3.72), H3BO3为B4 (k4 = 3.35), CaCl2为C1 (k1 = 2.93),即适宜的蔗糖和H3BO3浓度组合最有利于长梗杜鹃的花粉萌发,而CaCl2的添加显著抑制了其萌发。蔗糖作为影响长梗杜鹃花粉萌发率高低的决定因素,随其浓度的增加,萌发率呈现出先增后减的趋势,且以100 g·L-1为最适浓度,当浓度达到300 g·L-1时,花粉萌发率下降,萌发所需时间延长,部分培养基花粉管先端出现破裂现象;H3BO3的促萌效果虽不如蔗糖显著,但在一定程度上也促进了其花粉萌发。此外,蔗糖、H3BO3以及CaCl2都并非是长梗杜鹃花粉萌发的必要条件,在仅含10 g·L-1的琼脂培养基中仍有22.54%的萌发率。
表 1 长梗杜鹃花粉在L25(53)正交试验中的萌发率
Table 1. The germination rate of R. longipedicellatum pollen grains in L25(53) orthogonal experiment
试验号
No.蔗糖
Sucrose/(g·L-1)硼酸
H3BO3/ (mg·L-1)二氯化钙
CaCl2 /(mg·L-1)萌发率
Germination rate/%1 0 0 0 22.54±1.59c 2 0 50 50 71.06±1.72mn 3 0 100 150 58.73±1.20j 4 0 200 250 74.32±1.58n 5 0 300 350 43.21±1.72g 6 50 0 50 78.44±1.40o 7 50 50 150 69.38±3.16lm 8 50 100 250 63.82±1.27k 9 50 200 350 68.27±1.65lm 10 50 300 0 74.05±1.52n 11 100 0 150 78.22±1.13o 12 100 50 250 85.90±1.41p 13 100 100 350 46.72±1.47h 14 100 200 0 90.26±1.92q 15 100 300 50 70.48±1.75lm 16 150 0 250 53.20±0.96i 17 150 50 350 67.58±0.94l 18 150 100 0 87.52±1.43pq 19 150 200 50 87.94±0.95pq 20 150 300 150 32.71±1.62e 21 300 0 350 40.36±1.68g 22 300 50 0 18.57±1.37b 23 300 100 50 36.65±1.51f 24 300 200 150 13.86±0.77a 25 300 300 250 25.90±1.53d 注:同列数据不同小写字母表示差异显著(P<0.05)。
Note: Different small letters in each column indicate significant difference at P<0.05 level. -
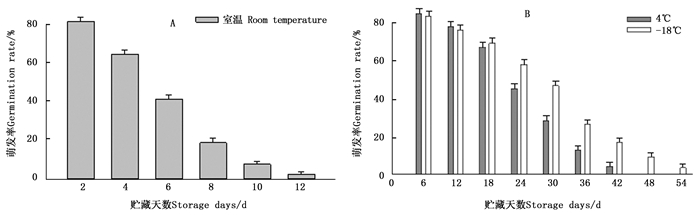
试验对长梗杜鹃在室温(20~25℃)、4℃和-18℃ 3个贮藏温度下不同贮藏时间的花粉萌发率进行了测定,结果(图 3)表明:随着贮藏天数的增加,3个温度下的花粉萌发率均呈逐步下降的趋势。在室温条件下,长梗杜鹃花粉萌发率下降最迅速,在第8天,已降至较低水平(18.86%),至第12天,其萌发率已低于5%,基本不能再用作父本进行杂交授粉(图 3A)。4℃和-18℃低温条件下其萌发率下降较缓慢,在贮藏的前12天,4℃的贮藏效果稍好一些,但在第24天萌发率出现较大降幅,至第36天仍有13.21%的萌发率;而-18℃保存的花粉直到第36天才出现较大降幅,在第48天仍有9.65%的萌发率(图 3B)。因此,适宜长梗杜鹃花粉贮藏的温度为-18℃,其次是4℃,即一定的低温有利于延长其花粉寿命。

图 3 长梗杜鹃花粉在不同贮藏温度下的萌发率变化情况
Figure 3. Change of germination rate of R.longipedicellatum pollen grains at different storage temperatures
-
对植物进行分类是为了更好的认识、保护和利用植物资源[11]。杜鹃属植物由于种类繁多,分布广、变异大,且在自然条件下存在非常明显的杂交现象,使得分类问题十分复杂,在系统划分和某些种的归属上长期众说纷纭。到目前为止,仍没有一个具有公认性的权威分类系统。花粉特征是植物分类和探讨植物起源演化及亲缘关系的重要依据之一[11,15],如山矾科植物中一些明显的花粉特征为该科的系统发育提供了重要证据[16]。杜鹃属植物孢粉学研究始于20世纪中叶,随着光学显微镜和扫描电子显微镜的应用得到了很快的发展。毛子军等[17]对产于我国东北的杜鹃属6种、1变种、1变型的花粉观察表明,其形态具有种间差别,可用于分类学研究;高连明等[18-19]对杜鹃属广义马银花亚属和杜鹃属4个亚属的25种和仿杜鹃属1种的花粉进行分析,认为花粉形态在杜鹃属中不能做为区分亚属的主要特征,而对组或亚组的划分具有重要的分类学意义;花粉粒大小、外壁纹饰和孔沟长短等微小差异可为某些种的分类学处理提供一些有用的孢粉学证据。同样,周兰英等[10]对四川26种野生杜鹃属植物的花粉观察表明,结合组或亚组的划分,孢粉学差异对种的确定具有一定实际意义。因此,对该属植物花粉形态特征的深入研究可为其准确分类提供重要依据。
Cai等[4]因长梗杜鹃罕见的花期、鲜黄的花朵及花、叶各部形态特征的特殊性,明显区别于相似种滇越杜鹃(R. rushforthii Argent & D.F. Chamberlain)、R. trancongii Argent & Rushforth和大田顶杜鹃(R. datiandingense Z.J. Feng),将其鉴定为新种,并划分系统位置为杜鹃亚属(subg. Rhododendron)、越桔杜鹃组(sect. Vireya)、类越桔杜鹃亚组(subsect. Pseudovireya)。本文对长梗杜鹃花粉形态的观察结果表明,从花蕾膨大期到花瓣盛开期,花粉逐渐成熟,且为复合体形成的四合花粉,表面具粘丝,使得各花粉粒在花药中粘连在一起,但经过蒸馏水和不同浓度酒精充分洗脱后的制片上仅有少数残留(图 1:Ⅱ-1、2,Ⅲ-1、2)。四合体直径介于高连明等[18]所报道的26.38~66.40 μm,萌发沟的长宽比已知的杜鹃种稍大一些。花粉分体间区为均匀的颗粒状同外壁,孔沟周围颗粒细密呈波状突起,这与高连明[18-19]和王玉国等[20]认为绝大多数杜鹃属植物花粉孔沟周围与极面区域光滑、近光滑或纹饰不明显的结论不符。Cullen[21]认为,越桔组的花粉形态在杜鹃属中最特殊,三孔花粉,孔为圆形,花粉表面纹饰为均匀的颗粒状,孔沟周围呈波状突起;胎座成熟时从中轴分离,种子两端具长尾状附属物等,是杜鹃亚属中一个特化的类群。长梗杜鹃具有越桔组杜鹃属植物花粉的典型特征,这间接从孢粉学层面支持了Cai等[4]对其系统位置划分的合理性。此外,根据周凌瑜等[22]对不同传粉方式的花粉形态比较研究,长梗杜鹃无论是花粉直径大小、还是外壁纹饰特征均与虫媒传粉的花粉特征相似,这为今后进一步开展长梗杜鹃的传粉生物学研究奠定了孢粉学基础。
花粉活力强弱是植物进行有性繁殖的基础,且直接影响着育种的效率。花粉在自然状态下可存活的时间不同植物间差异较大,锦带花(Weigela florida Bunge A. DC.)的花粉活力只能维持约3天[23],而海枣(Phoenix dactylifera L.)的花粉可达数月至1年[24]。TTC染色法因具有简单、快捷和有效等优点,越来越多被用于杜鹃属植物花粉活力的快速测定[25-26]。本研究采用TTC染色法对长梗杜鹃的花粉生活力进行了测定,结果表明,整个单花花期花粉活力均较高,于开花当天9:00 am达最大,为92.18%,低于大白花杜鹃(R. decorum Franch.)(开花72 h达到96.84%)和迷人杜鹃(R. agastum Balf. f. et W. W. Smith)(开花96 h达95.84%)[25],但高于大喇叭杜鹃(R. excellens Hemsl. et Wils.)(开花当天达84.43%)[27]和锈叶杜鹃(R. siderophyllum Franch.)(开花当天9:00 am达83.95%)[26]。可能由于花药中未成熟的花粉逐渐成熟累积的结果,使得长梗杜鹃花粉在末花期(花后第9天)仍保持着相对较高的活力水平[28],这与大喇叭杜鹃[27]和常绿杜鹃品种‘XXL’ [29]表现相似,但锈叶杜鹃花粉至开花第7天已无活力[26]。可见,不同杜鹃属植物花粉生活力差异较大,在开展杂交育种工作之前需要进行具体测定。长梗杜鹃因其稀有的花期、纯黄色的花朵、美观的树形,加之单花花粉活力持续时间长,很适宜作为父本材料开展杜鹃花期和花色育种工作,且采集开花当天9:00 am的花粉进行人工授粉更有利于杂交结实。
杜鹃属植物种类繁多,花粉离体培养过程中对蔗糖、硼离子和钙离子的需求以及最优培养组合存在较大差异[12]。本研究表明, 长梗杜鹃新鲜花粉萌发的适宜固体培养基为10 g·L-1琼脂+ 100 g·L-1蔗糖+ 200 mg·L-1 H3BO3+ 0 mg·L-1 CaCl2。同锦绣杜鹃(R. pulchrum Sweet)[13]、睫毛杜鹃(R. ciliatum Hook. f.)和比利时杜鹃(R. hybridum Hot)[12]一样,蔗糖对长梗杜鹃花粉萌发具有主导作用,适宜的蔗糖浓度均能显著促进其花粉萌发,浓度过低或过高会导致萌发率下降或花粉管先端破裂,这可能是因为适宜浓度的蔗糖为花粉提供了萌发所需的碳素营养,并能维持花粉与培养基之间的渗透平衡[30];而浓度过高会使外部环境渗透压过高产生质壁分离,进而使花粉萌发和生长受到抑制[31]。硼元素是花粉萌发的必需元素,能与蔗糖形成络合物,易于在组织中运输和参与花粉管果胶物质的合成[32]。一般情况下,花粉细胞中H3BO3含量较少,其正常萌发需要吸收柱头和花柱中的硼,故在离体萌发试验中需要添加适量的H3BO3来促进花粉的有效萌发[33]。本次正交试验表明,H3BO3对长梗杜鹃花粉萌发的影响水平皆不及蔗糖和CaCl2,但添加一定量的H3BO3起到了很好的促萌效果。姚成义等[34]对蓝猪耳花粉及张超仪等[12]对马银花(R. ovatum (Lindl.) Planch. ex Maxim)花粉特性的研究表明,其萌发不需要外源Ca2+的存在。本研究中,CaCl2的添加也显著抑制了长梗杜鹃花粉的萌发,这或许是蔗糖、H3BO3的添加降低了长梗杜鹃花粉萌发对Ca2+的需求,花粉细胞内已有相对较高的游离钙,能满足花粉萌发和花粉管伸长所需,这一点在今后的研究中还需要进一步的验证。此外,长梗杜鹃花粉在上述最适培养基中的萌发率稍低于TTC染色法对其活力的检测结果,可能是由于培养基配方的组成、培养条件、TTC的浓度、染色时间或是染色标准的主观判断误差等造成,但差值并不显著。这进一步验证了TTC染色法适合用于野外条件下长梗杜鹃花粉活力的快速测定。
花粉的贮藏寿命对种质资源保存和解决远缘杂交问题至关重要,而贮藏温度直接影响各花粉贮藏期的长短[14]。本研究初步探索了不同贮藏温度对长梗杜鹃花粉萌发力的影响,结果表明,在室温下可保存的时间较短,而低温大大延长了花粉的贮藏天数。这可能是由于低温干燥的贮藏环境降低了花粉细胞的呼吸强度和酶活性,进而减少了可溶性糖类和有机酸类物质的消耗[35]。长梗杜鹃花粉生命力在室温下只能保存10天左右,且整个贮藏期的活力均显著小于相对应开花后不同天数的测定结果,原因可能是花朵开放期间,各花药内花粉不断成熟累加及花丝、花柄等花器官为花粉提供了一定的营养,从而延缓了花粉的衰亡;而在4℃和-18℃低温条件下分别能保存36和48天左右,其萌发力水平高于映山红(R. simsii Planch.)、马银花和锦绣杜鹃[13],但低于黄杯杜鹃(R. wardii W. W. Smith),其在20℃、4℃和-16℃可分别贮藏30、60、120 d[14]。由此可见,长梗杜鹃花粉在-18℃低温条件下能贮藏较长时间,有利于解决杂交育种中与另一亲本花期不一致和远距离杂交等问题。
-
美丽的株型、纯黄色的花朵及罕见的花期赋予了长梗杜鹃极高的观赏价值,也是国内外开展杜鹃花育种极为难能可贵的亲本材料,在市场上有着巨大的产业发展前景。本研究对其不同开花时期的花粉形态、开花当天和开花后不同天数的花粉活力、离体萌发最适固体培养基的筛选及不同贮藏温度对花粉萌发力的影响进行了系统研究,据此了解该稀有濒危种的花粉特性。结果表明,长梗杜鹃花粉活力较强,适宜作为父本开展杂交育种研究,并确定了最佳采粉期以及为解决今后以长梗杜鹃为杂交父本的育种中可能遇到的花期不遇及远距离杂交等问题奠定了理论基础。同时,长梗杜鹃特殊的花粉形态特征从孢粉学层面支持了前人对其系统位置划分的合理性。
滇东南濒危植物长梗杜鹃的花粉形态及其特性研究
Pollen Morphology and Characteristics of Rhododendron longipedicellatum(Ericaceae), an Endangered Species Endemic to Southeastern Yunnan, China
-
摘要:
目的 对滇东南特有濒危植物长梗杜鹃花粉形态及其特性进行系统研究,为长梗杜鹃进一步的遗传育种、花粉种质保存和传粉生物学等相关保护利用提供一些科学依据,并为系统分类研究作旁证。 方法 利用光学显微镜和扫描电子显微镜对长梗杜鹃花朵不同发育时期(花蕾膨大期、花瓣初开期和花瓣盛开期)的花粉形态进行了观察;通过TTC染色法对开花当天不同时段及开花后不同天数的花粉活力进行了快速检测;做L25(53)正交试验探讨10 g·L-1琼脂和不同浓度的蔗糖、H3BO3、CaCl2组成的培养基配方对长梗杜鹃花粉萌发的影响。同时,研究了其花粉在不同贮藏温度下活力随贮藏时间的变化情况。 结果 表明:从花蕾膨大期到花瓣盛开期,花粉逐渐成熟,为四合体花粉,呈正四面体排列,四合体直径为43.0 65.4 μm,平均51.3 μm;单粒花粉近球形且表面有粘丝,具三孔沟。在扫描电镜下,花粉粒外壁为均匀的颗粒状,孔沟周围颗粒细密呈波状突起;在光镜下,花粉粒外壁为内外2层,厚度约相等为0.8 1.2 μm。长梗杜鹃整个单花花期花粉活力较强,于开花当天9:00达最高,为92.18%,至花末期(花后第9天)仍保持着48.5%的活力,适宜做父本开展相关杂交育种工作。长梗杜鹃开花当天9:00的花粉在10 g·L-1琼脂+100 g·L-1蔗糖+200 mg·L-1 H3BO3+ 0 mg·L-1 CaCl2的培养基中萌发率最高,达90.26%;极差分析表明,影响长梗杜鹃花粉萌发率高低的主次顺序为蔗糖→CaCl2→H3BO3,一定浓度的蔗糖和H3BO3均起到了很好的促萌效果,而CaCl2的添加则显著抑制了其花粉萌发。此外,适宜的低温有利于长梗杜鹃花粉的贮藏,-18℃条件下贮藏48 d仍有一定的萌发力。 结论 长梗杜鹃特殊的花粉外壁纹饰支持了前人对其系统位置划分的合理性。其较高的花粉活力、适宜的固体萌发培养基及贮藏温度的研究为今后以长梗杜鹃为父本的杂交育种提供了可靠的理论依据,对其他同属植物花粉形态及特性研究具有重要的指导意义。 Abstract:Objective To provide some references for the studies on genetic breeding, pollen germplasm preservation and pollination biology of Rhododendron longipedicellatum by investigating the pollen grains morphology and its characteristics. Method The pollen grains of R. longipedicellatum at different development stages including alabastrum intumescence stage, petal loose stage and petal full bloom stage were observed by light microscope (LM) and scanning electron microscope (SEM). The pollen viability in the process of flowering was detected by TTC staining method. The effects of 10 g·L-1 agar and different concentrations of sucrose, H3BO3 and CaCl2 on R. longipedicellatum pollen grains germination were studied by L25(53) orthogonal test. Simultaneously, the authors also studied the changes of pollen germination rate under different storage temperature. Result The pollen grains mature gradually from the alabastrum intumescence to the petal full bloom stage, and are arranged in tetrahedral tetrad. The tetrad diameter ranges from 43.0 μm to 65.4 μm with an average 51.3 μm. A single pollen is subspheroidal with tricolporate aperture, and viscin threads were seen on the pollen grains surface. The pollen grains exine sculpture is uniform granular in SEM, and the granulate around the colpus are more compact. Exine is composed with two layers in LM. The pollen viability maintained a high level over single flowering period, which was the highest (92.18%) at 9:00 on the first day of anthesis, and still remained relatively high (48.5%) until the end of flowering time (on the 9 days after flowering). The highest germination rate of R. longipedicellatum pollen grains at 9:00 on the first day of anthesis was 90.26% when cultured in the medium for 10 g·L-1 agar + 100 g·L-1 sucrose + 200 mg·L-1 H3BO3 + 0 mg·L-1 CaCl2. Range analysis showed that the important order of influence on pollen germination rate was sucrose, CaCl2 and H3BO3, a certain concentration of sucrose and H3BO3 had a good promote sprouting effect, while the addition of CaCl2 significantly inhibited the pollen grains germination. Furthermore, the appropriate low temperature was helpful to storage of R. longipedicellatum pollen, the germination rate maintained a certain level after 48 days storage with -18℃. Conclusion The special pollen grain exine sculpture of R. longipedicellatum supports the rationality of the division of its systematic position by predecessors. These findings of high pollen viability, suitable solid germination medium and storage temperature provide a reliable theoretical basis for hybridization breeding of R. longipedicellatum as the male parent. -
图 1 长梗杜鹃花朵不同发育时期的花粉形态
Figure 1. Pollen grains morphology of R. longipedicellatum at different development stages
图 2 长梗杜鹃花朵开放期间的花粉活力变化情况
Figure 2. Change of pollen viability at anthesis of R. longipedicellatum
图 3 长梗杜鹃花粉在不同贮藏温度下的萌发率变化情况
Figure 3. Change of germination rate of R.longipedicellatum pollen grains at different storage temperatures
表 1 长梗杜鹃花粉在L25(53)正交试验中的萌发率
Table 1. The germination rate of R. longipedicellatum pollen grains in L25(53) orthogonal experiment
试验号
No.蔗糖
Sucrose/(g·L-1)硼酸
H3BO3/ (mg·L-1)二氯化钙
CaCl2 /(mg·L-1)萌发率
Germination rate/%1 0 0 0 22.54±1.59c 2 0 50 50 71.06±1.72mn 3 0 100 150 58.73±1.20j 4 0 200 250 74.32±1.58n 5 0 300 350 43.21±1.72g 6 50 0 50 78.44±1.40o 7 50 50 150 69.38±3.16lm 8 50 100 250 63.82±1.27k 9 50 200 350 68.27±1.65lm 10 50 300 0 74.05±1.52n 11 100 0 150 78.22±1.13o 12 100 50 250 85.90±1.41p 13 100 100 350 46.72±1.47h 14 100 200 0 90.26±1.92q 15 100 300 50 70.48±1.75lm 16 150 0 250 53.20±0.96i 17 150 50 350 67.58±0.94l 18 150 100 0 87.52±1.43pq 19 150 200 50 87.94±0.95pq 20 150 300 150 32.71±1.62e 21 300 0 350 40.36±1.68g 22 300 50 0 18.57±1.37b 23 300 100 50 36.65±1.51f 24 300 200 150 13.86±0.77a 25 300 300 250 25.90±1.53d 注:同列数据不同小写字母表示差异显著(P<0.05)。
Note: Different small letters in each column indicate significant difference at P<0.05 level. 下载: 导出CSV
下载: 导出CSV
-
[1] 方瑞征, 闵天禄.杜鹃属植物区系的研究[J].云南植物研究, 1995, 17(4):359-379. [2] Chamberlain D, Hyam R, Argent G, et al. The Genus Rhododendron:Its Classification and Synonymy[M]. Edinburgh:Royal Botanic Garden Edinburgh, 1996:181. [3] Fang M Y, Fang R Z, He M Y, et al. Ericaceae[M]//Wu Z Y, Raven P H, Hong D Y. Flora of China. Beijing: Science Press & St. louis: Missouri Botanical Garden, 2015, 14: 260-445. [4] Cai L, Neilsen J, Dao Z L, et al. Rhododendron longipedicellatum (Ericaceae), a new species from Southeastern Yunnan, China[J]. Phytotaxa, 2016,282(4):296-300. doi: 10.11646/phytotaxa.282.4 [5] Wasson E, Rodd T. Tree & shrubs[M]. Beijing:China Forestry Publishing House, 2004:612-627. [6] 张长芹, 高连明, 薛润光, 等.中国杜鹃花的保育现状和展望[J].广西科学, 2004, 11(4):354-359. doi: 10.3969/j.issn.1005-9164.2004.04.021 [7] 兰熙, 张乐华, 张金政, 等.杜鹃花属植物育种研究进展[J].园艺学报, 2012, 39(9):1829-1838. [8] 李太强, 刘雄芳, 万友名, 等.滇东南濒危植物长梗杜鹃转录组微卫星特征分析[J].林业科学研究, 2017, 30(4):533-541. [9] IUCN. IUCN Red List Categories and Criteria: version 3.1[M]. Gland, Switzerland and Cambridge, United Kingdom: IUCN Species Survival Commission, 2001: 30. [10] 周兰英, 王永清, 张丽. 26种杜鹃属植物花粉形态及分类学研究[J].林业科学, 2008, 44(2):55-63. doi: 10.3321/j.issn:1001-7488.2008.02.009 [11] 张艳红.中国杜鹃花属植物分类的研究进展[J].辽东学院学报:自然科学版, 2011, 18(3):198-202. doi: 10.3969/j.issn.1673-4939.2011.03.003 [12] 张超仪, 耿兴敏.六种杜鹃花属植物花粉活力测定方法的比较研究[J].植物科学学报, 2012, 30(1):92-99. [13] 李玉萍, 陈堃, 王燕青, 等.测定杜鹃花花粉萌发力的液体培养基和贮藏方法研究[J].金陵科技学院学报, 2012, 28(3):56-61. doi: 10.3969/j.issn.1672-755X.2012.03.013 [14] 陆琳, 彭绿春, 宋杰, 等.不同高山杜鹃品种花粉活力测定及贮藏方法研究[J].山西农业科学, 2016, 44(2):175-178. doi: 10.3969/j.issn.1002-2481.2016.02.11 [15] 郭红艳, 王森, 晏巢, 等.华中五味子的雄花特征与花粉特性[J].林业科学, 2016, 52(3):112-120. [16] Wang Y G, Fritsch P W, Shi S H, et al. Phylogeny and infrageneric classification of Symplocos (Symplocaceae) inferred from DNA sequence data[J]. American Journal of Botany, 2004, 91(11):1901-1914. doi: 10.3732/ajb.91.11.1901 [17] 毛子军, 杨永富, 侯丽君.东北杜鹃花属植物花粉形态的研究[J].植物研究, 2000, 20(1):58-62,126-131. doi: 10.3969/j.issn.1673-5102.2000.01.009 [18] 高连明, 张长芹, 李德铢, 等.杜鹃花族花粉形态及其系统学意义[J].云南植物研究, 2002, 24(4):471-482. doi: 10.3969/j.issn.2095-0845.2002.04.008 [19] 高连明, 张长芹, 李德铢, 等.杜鹃属马银花亚属花粉形态的研究[J].武汉植物学研究, 2002, 20(3):177-181,245-247. doi: 10.3969/j.issn.2095-0837.2002.03.003 [20] 王玉国, 李光照, 漆小雪, 等.杜鹃花属植物花粉形态及其分类学意义[J].广西植物, 2006, 26(2):113-119. doi: 10.3969/j.issn.1000-3142.2006.02.001 [21] Cullen J. A revision of Rhododendron. I. Subgenus Rhododendron sections Rhododendron and Pogonanthum[J]. Notes from the Royal Botanic Garden Edinburgh, 1980, 39(1):207. [22] 周凌瑜, 刘群录, 邵邻相.虫媒花与风媒花花粉形态的比较[J].上海交通大学学报:农业科学版, 2008, 26(3):177-182. [23] 刘林德, 张萍, 张丽, 等.锦带花的花粉活力、柱头可授性及传粉者的观察[J].西北植物学报, 2004, 24(8):1431-1434. doi: 10.3321/j.issn:1000-4025.2004.08.013 [24] 胡适宜.被子植物胚胎学[M].北京:人民教育出版社, 1982:51-58. [25] 张敬丽.杜鹃花属迷人杜鹃的自然杂交起源研究[D].北京: 中科院研究生院, 2007. [26] 白天, 关文灵, 宋杰, 等.锈叶杜鹃的开花特性及繁育系统研究[J].西部林业科学, 2014, 43(1):47-53. doi: 10.3969/j.issn.1672-8246.2014.01.009 [27] 田晓玲.大喇叭杜鹃繁殖生物学研究[D].南京: 南京林业大学, 2011. [28] 张双双, 奚晓军, 张金凤.黑杨花粉活力和柱头可授性研究[J].安徽农业科学, 2010, 38(20):10550-10552. doi: 10.3969/j.issn.0517-6611.2010.20.024 [29] 林锐, 彭绿春, 李世峰, 等.常绿杜鹃品种'XXL'的花粉活力及柱头可授性观察[J].西部林业科学, 2016, 45(6):115-120. [30] Pierson E S, Li Y Q, Zhang H Q, et al. Pulsatory growth of pollen tubes:investigation of a possible relationship with the periodic distribution of cell wall components[J]. Acta Botanica Neerlandica, 1995, 44(2):121-128. doi: 10.1111/plb.1995.44.issue-2 [31] 卜志国, 杜绍华, 张晓曼.映山红开花习性与花粉生活力研究[J].安徽农业科学, 2011, 39(8):4562-4563, 4686. doi: 10.3969/j.issn.0517-6611.2011.08.061 [32] Fernando D D, Lazzaro M D, Owens J N. Growth and development of conifer pollen tubes[J]. Sexual Plant Reproduction, 2005, 18(4):149-162. doi: 10.1007/s00497-005-0008-y [33] 左丹丹, 明军, 刘春, 等.植物花粉生活力检测技术进展[J].安徽农业科学, 2007, 35(16):4742-4745. doi: 10.3969/j.issn.0517-6611.2007.16.015 [34] 姚成义, 赵洁.钙和硼对蓝猪耳花粉萌发及花粉管生长的影响[J].武汉植物学研究, 2004, 22(1):1-7. doi: 10.3969/j.issn.2095-0837.2004.01.001 [35] Nepi M, Franchi G G, Padni E. Pollen hydration status at dispersal:cytophysiological features and strategies[J]. Protoplasma, 2001,216(3-4):171-180. doi: 10.1007/BF02673869 -

 点击查看大图
点击查看大图

图(3) / 表(1)
计量
- 文章访问数: 4061
- HTML全文浏览量: 1996
- PDF下载量: 23
- 被引次数: 0
