


-
土壤动物是陆地生态系统中物质循环和能量流动的主要参与者,在凋落物分解、土壤有机质形成和结构塑造、土壤微生物活性及地下生物网结构与功能的调控等方面具有十分重要的作用,是联系地上和地下生态系统健康平衡发展的重要组成部分[1-4]。跳虫是弹尾纲(Collembola)动物的俗称,是分布范围极广、类群和数量最丰富、群落多样性最高的中小型土壤节肢动物[5-6]。它们生活周期短,具有稳定的地理分布和年变化规律,并且活动范围小、迁移能力弱,对土壤微环境变化敏感,其群落结构和多样性与地表植被组成、土壤理化性质等关系密切,可直接反应生境的土壤健康状况和植被演替阶段,通常被看作是评价土壤质量和森林生态环境变化的重要指示生物[7-11]。
八大公山国家级自然保护区(以下简称八大公山)被誉为“天然博物馆”和“物种基因库”,是亚热带地区保存最完整、面积最大的常绿落叶阔叶混交林原生性林区之一。向昌国等于2000年曾对八大公山的森林土壤动物群落多样性做过初步的调查,结果表明,螨类、弹尾类、线虫类和线蚓类为优势类群,是该区土壤动物的主体[12]。此后十多年来,未见对该区土壤动物分布特征的详细研究报道。为了弄清八大公山土壤跳虫群落结构和多样性特征,笔者选择八大公山不同海拔梯度的典型植被类型为研究对象,从海拔梯度、植物群落组成、季节变化及土壤理化性质等方面研究土壤跳虫的分布规律,探讨了不同环境因子对土壤跳虫群落结构和多样性的影响,以期进一步揭示八大公山土壤跳虫群落的分布特征,为筛选出理想的、能全面反映森林土壤生态系统健康的量化生物指标奠定基础,从而为建立高效、准确的森林健康评价和监测体系提供理论依据。
-
八大公山位于湖南省桑植县西北部,地处云贵高原古陆块东北边缘的武陵山脉北端(109°41′45″110°09′50″ E,29°39′18″29°49′48″ N)。属于亚热带山地湿润季风气候,年平均气温为11.5℃,年平均降水量为2 105.4 mm,全年相对湿度在90%以上。土壤类型主要有山地黄棕壤、石灰土和局部的山地沼泽土[13]。植被群落原生性较好,以常绿落叶阔叶混交林为主。
根据八大公山植被现状,笔者选择了落叶阔叶林、常绿落叶阔叶混交林和沟谷常绿阔叶3种生境的土壤跳虫群落为研究对象。3个生境根据海拔高度自上而下分别为:样地Ⅰ青冈林,优势树种为亮叶水青冈(Fagus lucida Rehd. et Wils.)、多脉青冈(Cyclobalanopsis multinervis W.C.Cheng et T.Hong),主要植被包括四川山矾(Symplocos setchuensis Brand)、杜鹃(Rhododendron simsii Planch.)、茶叶山矾(Symplocos theaefolia D. Don)、石灰花楸(Sorbus folgneri (Schneid.) Rehd.)、尾叶山茶(Camellia caudata Wall.)等。样地Ⅱ光皮桦林,优势树种为光皮桦(Betula luminifera H. Winkl.)、鹅耳枥(Carpinus turczaninowii Hance),主要植被包括青榨槭(Acer davidii Franch.)、灯台树(Bothrocaryum controversum (Hemsl.) Pojark.)、油茶(Camellia oleifera Abel.)、尖叶四照花(Dendrobenthamia angustata (Chun) Fang)、中华石楠(Photinia beauverdiana Schneid.)、水马桑(Weigela japonica Thunb. var. sinica (Rehd.)Bailey)。样地Ⅲ栲树林,优势树种为丝栗栲(Castanopsis fargesii Franch.),主要植被包括刨花润楠(Machilus pauhoi Kanehira)、锥栗(Castanea henryi (Skan) Rehd.et Wils.)、拟赤杨(Alniphyllum fortunei (Hemsl.) Makino)、木荷(Schima superba Gardn. et Champ.)、广东冬青(Ilex kwangtungensis Merr.)、鹅掌柴(Schefflera octophylla (Lour.) Harms)等。各生境基本情况见表 1。
表 1 样地基本情况
Table 1. The basic situation of plots
生境
Habitats海拔
Attitude/m坡度
Slope degree
/(°)森林类型
Forest type优势树种
Dominant species郁闭度
Canopy
density凋落物厚
The thickness of
litter layer/cmⅠ 1 475 22 落叶阔叶林
Deciduous Broad-leaved forest亮叶水青冈Fagus lucida
多脉青冈Cyclobalanopsis multinervis0.7 5~6 Ⅱ 1 100 30 常绿落叶混交林
Evergreen and deciduous mixed broad-leaved forest光皮桦Betula luminifera 0.8 3~5 Ⅲ 545 15 常绿阔叶林
Evergreen broad-leaved forest丝栗栲Castanopsis fargesii 0.7 3~4 -
于2016年4月、7月和10月分别在选定的3个样地中各设5个20 m×30 m的样方,每个样方按5点取样法设5个采样点,两采样点之间相距5 m以上。每个采样点按凋落物层和其下的真土层分别进行采样。凋落物层取样时,用面积为20 cm×20 cm的木制框放置在地表,收集框内凋落物和腐殖质;然后在其下的真土层用直径为5 cm的土钻钻取15 cm深的土样,每个点采集5个土样,4个混合用于分离土壤跳虫,1个混合均匀用于土壤理化性质检测。采集的凋落物层和土壤层样品分别装入PE自封袋带回实验室。
用于收集土壤跳虫的样品分别用改进的Tullgren干漏斗法分48 h,将标本收集到75%酒精中固定。将收集到的土壤跳虫标本在体视显微镜(Olympus SZX16)下进行分类鉴定和数量统计。分类检索方法参考尹文英[14-15]和Bellinger等[16]共同修改的分类系统,鉴定到属。
用于测试理化性质的土壤样品带回实验室后先风干,然后再分别采用重铬酸钾氧化外加热法、半微量凯氏法、碱熔-钼锑抗比色法、碱熔-火焰光度计法、电位法和原子吸收分光光度计法测定土壤有机质、全氮、全磷、全钾、全钙、全镁含量和pH值[17]。
-
土壤跳虫群落特征主要采用Shannon-Wiener多样性指数H′、Pielou均匀度指数Js、Margalef丰富度指数R和Simpson优势度指数D来计算[18]。计算公式如下:
Shannon-Wiener多样性指数${H^\prime } = - \sum\limits_{i = 1}^s {\left( {{P_i} ln {P_i}} \right)} $;
Pielou均匀度指数${J_s} = {H^\prime }/ln S$;
Margalef丰富度指数$R=(S-1) / ln N$;
Simpson优势度指数$D=\sum\limits_{i=1}^{s} P_{i}^{2}$。
式中,Pi=ni/N,ni为第i类群个体数,N为群落总个体数,S为群落类群数。
数据分析前,对土壤跳虫数据进行log(x+1)对数转换,对于服从正态分布的土壤跳虫数据,采用SPSS 19.0软件进行相关分析。用单因素方差分析(One-way ANOVA)和最小显著差异法(Least significant difference)分析不同生境土壤跳虫群落结构和多样性的季节变化及土壤理化性质的差异;用双因素方差分析(Two-way ANOVA)分析生境和季节对土壤跳虫群落结构和多样性的影响;用Pearson相关系数分析土壤跳虫群落与土壤理化性质相关性。同时通过CANOCO 4.5软件进行冗余分析(Redundancy analysis,RDA),分析土壤跳虫主要群落与土壤理化性质的相关性,并用蒙特卡罗置换检验(Monte Carlo permutation test)筛选显著影响土壤跳虫主要类群的因子(P<0.05)。
-
3个样地的凋落物层和土壤层共分离出土壤跳虫3 785头,隶属于4目15科29属(表 2)。其中优势类群包括符䖴属Folsomia、长䖴属Entomobrya、球角䖴属Hypogastrura和棘䖴属Onychiurus,4属共占总个体数的71.08%。常见类群有小圆䖴属Smithurinus、土䖴属Tullbergia、疣䖴属Neanura、等节䖴属Isotoma、德䖴属Desoria、原等䖴属Proisotoma、圆䖴属Sminthurus、刺齿䖴属Homidia和齿棘䖴圆属Arrhopalites,9属共占总个体数的23.91%。稀有类群包括长角长䖴属Orchesellides、类符䖴属Folsomina、鳞䖴属Tomocerus和原䖴属Protanura等16属,共占个体总数的5.01%。
表 2 八大公山不同生境土壤跳虫群落组成
Table 2. Composition of soil Collembolan communities in different habitats of Badagong Mountains
属
GenusⅠ(n=5) Ⅱ(n=5) Ⅲ(n=5) 小计
Subtotal优势度/%
Dominance土壤层
Layer of soil凋落物层
Layer of
plant litter土壤层
Layer of soil凋落物层
Layer of
plant litter土壤层
Layer of soil凋落物层
Layer of
plant litter符䖴属 Folsomia sp. 126 359 57 190 87 274 1 093 28.88 长䖴属 Entomobrya sp. 11 204 45 170 14 188 632 16.70 球角䖴属 Hypogastrura sp. 55 214 49 111 35 103 567 14.98 棘䖴属 Onychiurus sp. 62 147 24 51 32 82 398 10.52 小圆䖴属 Smithurinus sp. 7 81 15 175 6 94 378 9.99 土䖴属 Tullbergia sp. 17 30 1 21 6 30 105 2.77 疣䖴属 Neanura sp. 9 23 6 7 8 36 89 2.35 等节䖴属 Isotoma sp. 19 21 10 9 10 9 78 2.06 德䖴属 Desoria sp. 11 15 8 11 4 11 60 1.59 原等䖴属 Proisotoma sp. 14 9 11 4 3 16 57 1.51 圆䖴属 Sminthurus sp. 1 26 1 9 15 52 1.37 刺齿䖴属 Homidia sp. 1 21 5 21 48 1.27 齿棘圆䖴属 Arrhopalites sp. 12 8 18 38 1.00 长角长䖴属 Orchesellides sp. 13 6 8 27 0.71 类符䖴属 Folsomina sp. 1 8 1 1 7 5 23 0.61 鳞䖴属 Tomocerus sp. 6 1 13 20 0.53 原䖴属 Protanura sp. 10 1 1 7 19 0.50 钩圆䖴属 Bourletiella sp. 12 2 4 18 0.48 短角䖴属 Neelus sp. 2 11 13 0.34 鳞长䖴属Lepidocyrtus sp. 4 2 5 11 0.29 拟裸长角䖴属 Pseudosinella sp. 6 1 3 10 0.26 刺䖴属 Acanthocyrtus sp. 6 1 1 2 10 0.26 裔符䖴属Folsomides sp. 7 2 9 0.24 羽圆䖴属 Dicyrtoma sp. 5 1 1 1 8 0.21 爪䖴属Paronella sp. 4 3 7 0.18 铲圆䖴属Papirinus sp. 5 1 6 0.16 奇䖴属 Xenylla sp. 1 3 1 5 0.13 裸长角䖴属 Sinella sp. 3 3 0.08 柳䖴属 Willowsia sp. 1 1 0.03 个体数合计Total of Individual Number 335 1 252 230 807 215 946 3 785 类群数合计Total of Genus Number 14 28 14 26 13 23 29 注Note:Ⅰ:青冈林Forest of Fagus lucida,Ⅱ:光皮桦林Forest of Betula luminifera,Ⅲ:栲树林Forest of Castanopsis fargesii。 3个样地跳虫的个体数量分布情况为:青冈林(41.93%)>光皮桦林(30.67%)>栲树林(27.40%)。从属这一分类等级上来看,符䖴属、长䖴属、球角属在3个样地均为优势类群,棘䖴属在青冈林为优势类群,在光皮桦林和栲树林都为常见类群,小圆䖴属只在光皮桦林为优势类群,在青冈林和栲树林为常见类群。
-
凋落物层和土壤层提供不同的栖息环境和食物源,造成土壤跳虫在两层中的差异分布。
从类群组成和个体数量来看,凋落物层土壤跳虫的类群数和个体数均高于土壤层。在凋落物层共分离土壤跳虫3 005头,占总个体数的79.39%,隶属于4目15科29属。土壤层共分离土壤跳虫780头,仅占总个体数的20.61%,隶属于3目11科18属。凋落物层和土壤层中的优势类群也并不相同。在土壤层中,符䖴属占绝对优势,所占比例达到34.61%,其次是球角䖴属(17.82%)和棘䖴属(15.13%);在凋落物层优势类群按数量由高到低依次为符䖴属(27.39%)、长䖴属(18.7%)、球角䖴属(14.24%)、小圆䖴属(11.65%)。此外,齿棘圆䖴属、长角长䖴属、鳞䖴属、钩圆䖴属Bourletiella、短角䖴属Neelus、鳞长䖴Lepidocyrtus、拟裸长角䖴属Pseudosinella、爪䖴属Paronella、铲圆䖴属Papirinus、裸长角䖴属Sinella、柳䖴属Willowsia都只在凋落物层分布(表 2)。
-
3种生境土壤跳虫群落分布的季节变化规律并不一致(图 1)。青冈林土壤跳虫类群数大小顺序为春季>秋季>夏季,且春季极显著地高于夏季(P<0.01);个体数为秋季>春季>夏季,且秋季显著高于夏季(P<0.05)。光皮桦林和栲树林土壤跳虫类群数均为春季>夏季>秋季,春季显著高于秋季(P<0.05),但二者个体数季节变化规律不同,光皮桦林为秋季>春季>夏季,秋季显著高于夏季(P<0.05),而栲树林为春季>秋季>夏季,并没有显著的季节差异(P>0.05)。与光皮桦林和栲树林相比,青冈林土壤跳虫群落分布随季节变化波动更显著。

图 1 不同生境土壤跳虫分布的季节变化
Figure 1. The seasonal distribution of soil Collembolan in different habitats of Badagong Mountains
-
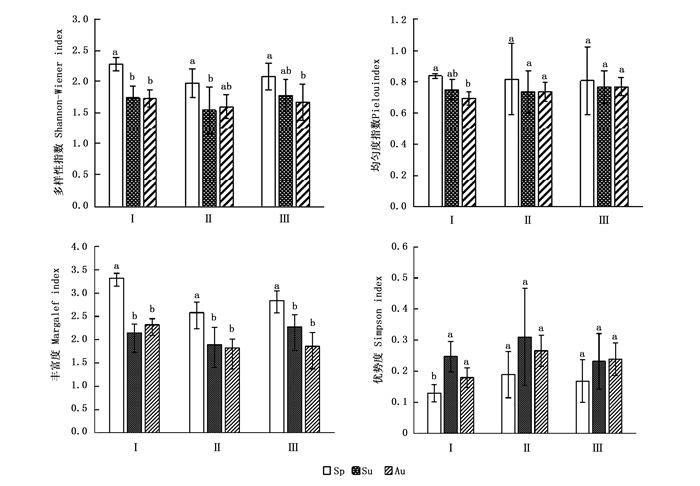
八大公山3种生境土壤跳虫的Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数变化趋势基本一致,Simpson优势度指数季节变化趋势则截然不同(图 2)。就Shannon-Wiener多样性指数来看,3种生境均是在春季最高并达到显著差异水平(P<0.05),青冈林和栲树林在秋季最低,光皮桦林在夏季最低;青冈林春季均匀度指数也显著高于秋季(P<0.05),而光皮桦林和栲树林的均匀度指数的季节差异不显著(P>0.05);丰富度指数在青冈林季节变化顺序为春季>秋季>夏季,光皮桦林和栲树林均为春季>夏季>秋季;3种生境土壤跳虫的优势度指数都在春季最低,青冈林和光皮桦林夏季最高,且青冈林达到显著差异水平(P<0.05),而光皮桦林和栲树林差异不显著(P>0.05)。

图 2 不同生境土壤跳虫多样性指数的季节变化
Figure 2. The seasonal diversity index of soil Collembolan in different habitats from Badagong Mountains
-
生境和季节变化对土壤跳虫群落结构具有显著影响(P<0.05)(表 3)。就不同生境比较来看,青冈林的土壤跳虫类群数显著高于光皮桦林(P<0.01)和栲树林(P<0.05),并且青冈林土壤跳虫个体数(P<0.01)、Shannon-Wiener多样性指数(P<0.05)和Margalef丰富度指数(P<0.05)都显著高于光皮桦林,与栲树林没有显著差异(P>0.05)。栲树林土壤跳虫类群数、个体数、Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数都高于光皮桦林,但未达到显著差异水平(P>0.05)。表明在3种生境间,青冈林生境条件更为优越,更适合土壤跳虫分布,栲树林次之,光皮桦林最差。从季节变化来看,土壤跳虫类群数、Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数都表现为春季显著地高于夏季和秋季(P<0.05);而个体数则是秋季显著高于夏季,与春季差异不显著;Simpson优势度指数夏季显著高于春季,与秋季差异不显著(P>0.05)。
表 3 不同生境和季节的土壤跳虫群落多样性
Table 3. Diversity of soil Collembolan community in different habitats and seasons
生境
Habitats季节
Seasons类群数
Group number个体数
Individual number多样性(H′)
Shannon-Wiener index均匀度(Js)
Pielou index丰富度(R)
Margalef index优势度(D)
Simpson indexⅠ 13.13±3.16 a 105.80±42.59 a 1.92±0.32 a 0.76±0.09 ab 2.60±0.65 a 0.19±0.07 b Ⅱ 9.93±2.46 b 69.13±31.6 b 1.71±0.33 a 0.76±0.10 ab 2.10±0.56 b 0.25±0.11 a Ⅲ 10.87±2.77 b 77.40±26.98 ab 1.85±0.27 a 0.78±0.05 ab 2.32±0.56 ab 0.21±0.05 ab Sp 13.67±3.09 A 81.07±33.05 AB 2.12±0.19 A 0.82±0.05 A 2.91±0.51 A 0.16±0.04 B Su 10.07±2.25 B 65.93±23.91 B 1.70±0.28 B 0.75±0.08 B 2.11±0.43 B 0.26±0.10 A Au 10.20±2.46 B 105.33±42.69 A 1.67±0.25 B 0.73±0.09 B 2.00±0.46 B 0.23±0.08 A 注:表中数据为平均值±标准误,n=15.同一栏相同小写字母表示生境间差异不显著,不同小写字母表示生境间差异显著(P<0.05)。同一栏相同大写字母表示季节间差异不显著,不同大写字母表示季节间差异显著(P<0.05)。下同。
Note: Data in the figures are mean±SE, n=15.The same small letters in the column indicate no significant difference between diffrent habitats, while different small letters indicate significant differences (P<0.05). The same capital letters in the column indicate no significant difference between diffrent seasons, while different capital letters indicate significant differences(P<0.05). The same below.对生境和季节变化与土壤跳虫群落多样性的双因素方差分析结果表明(表 4),生境对土壤跳虫类群数、个体数具有极显著的影响(P<0.01),对Margalef丰富度和Simpson优势度影响显著(P<0.05),对Shannon-Wiener多样性指数和Pielou均匀度指数影响不显著(P>0.05)。季节对土壤跳虫的类群数、个体数、Shannon-Wiener多样性指数、Margalef丰富度指数和Simpson优势度指数均有极显著的影响(P<0.01),并对Pielou均匀度指数影响显著(P<0.05)。同时,生境与季节交互作用对土壤跳虫群落多样性影响不显著(P>0.05)。以上结果表明,季节变化对八大公山土壤跳虫群落类群数和个体数的影响与生境具有同等重要的作用,对多样性、均匀度、丰富度和优势度的影响则比生境更强,且生境和季节的交互作用抵消了单个因子对土壤跳虫群落多样性的影响。
表 4 不同生境和季节变化对土壤跳虫群落多样性的影响
Table 4. Effect of different habitats and seasons on diversity of soil Collembolan community
项目Items DF 类群数
Group number个体数
Individual number多样性(H′)
Shannon-Wiener index均匀度(Js)
Pielou index丰富度(R)
Margalef index优势度(D)
Simpson indexF p F p F p F p F p F p 生境Habitats 2 7.01 0.00 5.73 0.00 2.81 0.73 0.31 0.74 4.56 0.02 4.00 0.03 季节Seasons 2 10.22 0.00 5.67 0.01 13.54 0.00 4.55 0.02 16.76 0.00 7.70 0.00 交互作用Interaction 4 0.46 0.76 0.77 0.55 0.36 0.84 0.55 0.70 0.73 0.58 0.63 0.64 -
土样检测结果表明,3种生境土壤有机质、全氮、全钾、Ca、Mg含量及pH值均存在显著差异(表 5)。其中,青冈林土壤有机质、全氮和钙含量显著高于光皮桦林和栲树林(P<0.05),全钾和镁含量、pH值则显著低于光皮桦林和栲树林(P<0.05)。且光皮桦林的土壤有机质和镁含量显著低于青冈林和栲树林。表明3个样地中,青冈林土壤肥力最高,而光皮桦林最低。
表 5 不同生境土壤主要理化性质比较
Table 5. Chief soil properties in different habitats
生境Habitats SOM/(g·kg-1) TN/(g·kg-1) TP/(g·kg-1) TK/(g·kg-1) Ca/(g·kg-1) Mg/(g·kg-1) pH Ⅰ 127.06±17.98 a 6.29±2.14 a 6.49±2.02 a 14.94±2.01 a 1.19±0.93 a 0.15±0.03 a 3.89±0.45 a Ⅱ 63.27±7.80 b 3.38±0.41 b 5.42±0.92 a 25.31±0.82 b 0.22±0.19 b 0.23±0.07 b 4.89±0.15 b Ⅲ 96.65±28.47 c 4.37±0.88 b 6.44±0.39 a 23.78±2.27 b 0.47±0.51 ab 0.38±0.06 c 4.87±0.26 b 注:SOM:土壤有机质;TN:土壤全氮;TP:土壤全磷;TK:土壤全钾;Ca:土壤全钙;Mg:土壤全镁;pH:土壤pH值。
Note: SOM: Soil organic matter; TN: Soil total Nitrogen; TP: Soil total Phosphor; TK: Soil total Potassium; Ca:Soil total Calcium; Mg: Soil total Magnesium; pH: Soil pH.Pearson相关系数分析结果显示(表 6),土壤跳虫类群数与土壤有机质、全氮和全磷存在极显著的正相关关系(P<0.01),与全钾和pH值显著负相关(P<0.05);土壤跳虫个体数量与有机质、全氮和全磷极显著地正相关(P<0.01);多样性与土壤有机质和全氮极显著正相关(P<0.01),与全钾和pH值显著负相关(P<0.05);土壤跳虫丰富度也与土壤有机质、全氮和全磷存在极显著的正相关关系(P<0.01),与全钾和pH值显著负相关(P<0.05);优势度与土壤有机质显著负相关(P<0.05),与全钾显著正相关(P<0.05);而均匀度与各土壤理化指标相关性不显著(P>0.05)。
表 6 土壤跳虫群落与土壤主要理化性质相关系数
Table 6. Pearson correlation coefficients between Collembolan community and soil properties
项目 SOM/(g·kg-1) TN/(g·kg-1) TP/(g·kg-1) TK/(g·kg-1) Ca/(g·kg-1) Mg/(g·kg-1) pH 类群数Genus 0.75** 0.83** 0.78** -0.56* 0.39 -0.21 -0.6* 个体数Individuals 0.67** 0.70** 0.69** -0.42 0.34 -0.12 -0.51 多样性Diversity 0.68** 0.685** 0.49 -0.63* 0.41 -0.33 -0.56* 均匀度Evenness -0.05 -0.18 -0.4 -0.14 0.08 -0.21 0.3 丰富度Richness 0.67** 0.79** 0.71** -0.56* 0.36 -0.25 -0.57* 优势度Dominance -0.57* -0.46 -0.26 0.55* -0.46 0.33 0.41 注Note:**P<0.01,*P<0.05。 进一步对3种生境土壤跳虫主要类群与土壤理化性质的相关性进行冗余度(RDA)分析,结果显示,第一轴和第二轴分别解释了土壤跳虫主要类群总变量的52.0%和9.1%,与土壤因子的相关性系数分别为0.896和0.894。总体来看,第一轴主要反应了土壤pH、全钾、全镁对主要类群的影响,第二轴主要区分了长属、符属、棘属、球角属与小属的分布特征。蒙特卡罗置换检验显示,土壤有机质(F=4.549,p=0.018)、pH(F=5.323,p=0.012)、土壤全钾(F=6.49,p=0.002)和全钙(F=3.745,p=0.048)对土壤主要群落的影响显著(p<0.05)。且土壤有机质和全氮与第一排序轴呈显著正相关(r=0.61,r=0.43,p<0.05),土壤全钾、pH与第一排序轴呈显著负相关(r=-0.69,r=-0.64,p<0.05)。棘属、符属、球角属与土壤有机质、全氮呈较强的正相关性,而与土壤pH和全钾呈较强的负相关性;长属与土壤有机质、全氮、全磷呈一定程度的相关性;球角属与全钙、全磷含量有较强正相关性,与pH和全钾、全镁呈较强负相关;小圆属与土壤主要理化性质相关性不明显(图 3)。
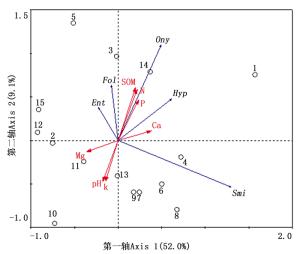
图 3 土壤跳虫主要类群的RDA分析
Figure 3. Results of redundancy analysis of dominant group of soil Collembolan
-
符䖴属、长䖴属、球角䖴属和棘䖴属是八大公山土壤跳虫的优势类群,对土壤跳虫群落结构和分布起主要作用。3个季节调查均有分布的属有20个,且符属、长属和球角属均为优势类群,表明八大公山土壤跳虫群落组成具有一定的稳定性,同时也反映出稀有类群对季节变化具有较高的敏感性。
土壤跳虫在3种生境的类群和个体分布不均匀,青冈林的土壤跳虫群落结构和多样性显著优于光皮桦林和栲树林(P<0.05)。多项研究表明,影响土壤动物群落的环境因素复杂多样,包括土壤(土壤温度、土壤湿度、pH值、有机质、凋落物数量、土壤矿质元素以及污染物质含量等)、植被、气候和地形等,它们构成土壤动物群落的栖息环境,对土壤动物群落的组成与数量、水平结构和垂直结构等具有重要影响[19-21]。八大公山的3种生境因林型和立地条件不同,凋落物层数量和质量,以及土壤理化性质均有差异。其中青冈林为落叶阔叶林,林内优势树种亮叶水青冈与多脉青冈树形高大,冠层稠密,郁闭度高,形成了良好的林内小气候,林下凋落物层丰厚。而凋落物是森林生态系统有机物质的主要来源,能为土壤动物的生存提供丰富的食物源和多样的微环境[22],因此,落叶阔叶青冈林丰厚的凋落物层是土壤跳虫生存和繁殖理想场所,其土壤跳虫类群数和个体数量多,多样性、均匀度、丰富度等指数比其他两种生境高,群落稳定性更好。Sayer等[23]和袁志忠等[24]的研究也证实,森林凋落物的数量和质量是影响土壤节肢动物群落结构和多样性的主要因素。而且,地上植物凋落物数量和质量的差异不仅直接影响以其为栖息生境和物质能量来源的土壤生物群落结构和功能,还将通过凋落物分解和系统养分循环影响土壤养分含量,并将反馈至植物的生长和群落演替过程[25-26]。青冈林土壤有机质、全氮含量均显著高于光皮桦林和栲树林,通过对3种生境土壤性质与主要跳虫群落相关性的分析也表明,土壤跳虫主要群落变化与土壤有机质和全氮密切正相关,并与土壤pH值、全钾负相关。表明青冈林较高的土壤有机质、全氮含量和较低的土壤pH值、全钾含量是导致土壤跳虫种类和数量显著高于光皮桦林和栲树林的主要原因。
土壤跳虫群落的季节变化主要表现在类群数、个体数以及不同生境多样性指数的变化,这种变化体现了不同类群的土壤跳虫在生态学习性上的差异及其对环境的适应[27]。3种生境土壤跳虫类群数、个体数和多样性指数等均随季节变化而波动。但总体来说,青冈林和光皮桦林受季节变化影响大,类群数、个体数和多样性指数均存在显著的季节差异。而栲树林土壤跳虫群落季节差异相对较小。这种差异主要应归因于环境温湿度与凋落物层质量和数量两个方面的季节差异。一方面,位于较高海拔的青冈林和光皮桦林水热条件受季节变化影响较位于低海拔的栲树林更显著;另一方面,落叶阔叶林与常绿落叶阔叶混交林凋落物层的数量和质量也会随季节更替而明显不同。在秋季,高山地带温度下降迅速,大量的新鲜凋落物伴随低温累积,为土壤跳虫生存提供了较好的栖息环境,部分凋落物中生存的类群迅速繁殖,种群数量大幅增加,优势集中现象突出。而在春季,地处亚热带的八大公山气温已逐渐回升,凋落物经过秋冬两季大型土壤动物的咀嚼、撕碎,变得易被土壤跳虫分解,适宜的环境条件和丰富的食物源使得土壤跳虫类群数和均匀度增高,Shannon-Wiener指数也随之上升。调查结果还表明,土壤跳虫主要分布在凋落物及腐殖土中,而真土层数量较少,且随着季节变化,部分类群在凋落物层和真土层中呈现上下移动,导致凋落物层和真土层跳虫优势类群存在季节性变化。在凋落物层生存的小圆属在春、秋季为优势类群,在夏季则为常见类群;半土生的棘属在春季为优势类群,夏、秋两季则为常见类群。土壤跳虫类群的这种季节变化与其生活史策略紧密相关,许多类群主要在落叶分解中后期起作用[28-29]。同时,不同土壤跳虫类群对环境变化的适应程度和响应方式不同。随着季节变化,各生境的凋落物数量、质地以及土壤理化性质均会发生变化,为跳虫群落提供了不同的微环境,从而形成了不同季节主要类群的变化[30-31]。
-
八大公山3种典型植被类型的土壤跳虫主要集中分布于凋落物层。生境的凋落物数量、土壤理化性质对土壤跳虫群落结构与多样性具有显著影响。土壤跳虫个体数、类群数、多样性指数和丰富度指数大小顺序为青冈林>栲树林>光皮桦林,且主要类群与生境的土壤有机质、全氮呈显著正相关,与土壤全钾、pH呈显著负相关。季节对土壤跳虫群落结构与多样性同样具有显著影响,春季(4月)土壤跳虫类群数最多,多样性指数、均匀度指数和丰富度指数均最高,群落结构最复杂;秋季个体数量最多,但均匀度和丰富度最低。因此,八大公山土壤跳虫群落结构与多样性受生境和季节变化影响显著,生境的植被类型与土壤理化性质共同决定土壤跳虫分布。
本研究结果反映出土壤跳虫群落结构随气候、植被类型、土壤性质和海拔高度等重要环境因素变化而存在显著差异,这些差异表明土壤跳虫对自然环境的适应有其独特的生理生态学机制。同时也提示,保持地表植被多样性和保留地表凋落物对促进土壤动物多样性、提高土壤生态系统健康平衡发展具有重要意义。而进一步探索土壤跳虫对环境的适应机制和生物学习性,将有助于我们了解其生态功能,并为自然保护区生物多样性资源保护和整个森林生态系统健康持续发展提供科学的理论依据。
八大公山土壤跳虫群落结构与多样性的季节动态
Seasonal Dynamics of Community Structure and Diversity of Soil Collembolan in Badagong Mountains
-
摘要:
目的 探索八大公山国家级自然保护区生境和季节变化对土壤跳虫群落分布的影响,为自然保护区森林健康的评价和监测提供科学的理论依据。 方法 于2016年4月、7月和11月对该区青冈林、光皮桦林和栲树林的凋落物和土壤进行取样调查,采用Tullgren干漏斗法分离土壤跳虫,分析土壤跳虫群落结构和多样性与生境和季节变化的相关性。 结果 共捕获土壤跳虫3 785头,隶属于4目15科29属,其中优势类群为符䖴属、长䖴属、球角䖴属和棘䖴属,4属共占总个体数的71.08%。分析数据表明,土壤跳虫主要集中分布于凋落物层,其个体数、类群数、Shannon-Wiener多样性指数和Margalef丰富度指数大小顺序为青冈林>栲树林>光皮桦林,且主要类群与生境的土壤有机质、全氮呈显著正相关,与土壤全钾、pH呈显著负相关。春季(4月)土壤跳虫类群数最多,Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数均最高,群落结构最复杂;秋季个体数量最多,但丰富度最低。 结论 八大公山土壤跳虫群落结构与多样性受生境和季节变化影响显著,生境的植被类型与土壤理化性质共同决定土壤跳虫分布,保持地表植被多样性和保留凋落物对促进土壤跳虫多样性具有积极作用。 Abstract:Objective In order to analysis the effects of habitat and seasonal variation on the structure and diversity of soil Collembolan community in Badagong Mountains National Nature Reserve, and provides basis for further evaluating and monitoring forest health in nature reserves. Method The soil Collembolan from litter and soil layer of Fagus lucida forest, Betula luminifera forest and Castanopsis fargesii forest was collected by Tullgren dry funnel method in April, July and November 2016. Result A total of 3 785 soil Collembolans were captured, belonging to 4 orders, 15 families and 29 genera. The dominant genera are Folsomia, Entomobrya, Hypogastrura and Onychiurus, accounting for 71.08% of the total individuals. Most of the soil Collembolan distributed in the litter layer. The quantity of genus, quantity of individual, the Shannon-Wiener diversity index and the Margalef richness index were in order of F. lucida forest > C. fargesii forest > B. luminifera forest. The dominant genus of soil Collembolan was significant and positively correlated with total soil organic matter and soil nitrogen, but negatively correlated with soil total potassium and pH value. The quantity of genus, the Shannon-Wiener diversity index, the Pielou evenness index and the Margalef richness index were the highest in spring (April), while the largest quantity of individuals but the lowest Margalef richness index in autumn (November). Conclusion The structure and diversity of soil Collembolan community in Badagong Mountains National Nature Reserve are significantly affected by the seasonal variation and habitats. The vegetation types and soil physical and chemical properties of habitat shared the common distribution of soil Collembolan. Therefore, maintaining diversity of vegetation and preserving plant litter have a positive effect on promoting soil Collembolan diversity. -
图 1 不同生境土壤跳虫分布的季节变化
Figure 1. The seasonal distribution of soil Collembolan in different habitats of Badagong Mountains
图 2 不同生境土壤跳虫多样性指数的季节变化
Figure 2. The seasonal diversity index of soil Collembolan in different habitats from Badagong Mountains
图 3 土壤跳虫主要类群的RDA分析
Figure 3. Results of redundancy analysis of dominant group of soil Collembolan
表 1 样地基本情况
Table 1. The basic situation of plots
生境
Habitats海拔
Attitude/m坡度
Slope degree
/(°)森林类型
Forest type优势树种
Dominant species郁闭度
Canopy
density凋落物厚
The thickness of
litter layer/cmⅠ 1 475 22 落叶阔叶林
Deciduous Broad-leaved forest亮叶水青冈Fagus lucida
多脉青冈Cyclobalanopsis multinervis0.7 5~6 Ⅱ 1 100 30 常绿落叶混交林
Evergreen and deciduous mixed broad-leaved forest光皮桦Betula luminifera 0.8 3~5 Ⅲ 545 15 常绿阔叶林
Evergreen broad-leaved forest丝栗栲Castanopsis fargesii 0.7 3~4  下载: 导出CSV
下载: 导出CSV
表 2 八大公山不同生境土壤跳虫群落组成
Table 2. Composition of soil Collembolan communities in different habitats of Badagong Mountains
属
GenusⅠ(n=5) Ⅱ(n=5) Ⅲ(n=5) 小计
Subtotal优势度/%
Dominance土壤层
Layer of soil凋落物层
Layer of
plant litter土壤层
Layer of soil凋落物层
Layer of
plant litter土壤层
Layer of soil凋落物层
Layer of
plant litter符䖴属 Folsomia sp. 126 359 57 190 87 274 1 093 28.88 长䖴属 Entomobrya sp. 11 204 45 170 14 188 632 16.70 球角䖴属 Hypogastrura sp. 55 214 49 111 35 103 567 14.98 棘䖴属 Onychiurus sp. 62 147 24 51 32 82 398 10.52 小圆䖴属 Smithurinus sp. 7 81 15 175 6 94 378 9.99 土䖴属 Tullbergia sp. 17 30 1 21 6 30 105 2.77 疣䖴属 Neanura sp. 9 23 6 7 8 36 89 2.35 等节䖴属 Isotoma sp. 19 21 10 9 10 9 78 2.06 德䖴属 Desoria sp. 11 15 8 11 4 11 60 1.59 原等䖴属 Proisotoma sp. 14 9 11 4 3 16 57 1.51 圆䖴属 Sminthurus sp. 1 26 1 9 15 52 1.37 刺齿䖴属 Homidia sp. 1 21 5 21 48 1.27 齿棘圆䖴属 Arrhopalites sp. 12 8 18 38 1.00 长角长䖴属 Orchesellides sp. 13 6 8 27 0.71 类符䖴属 Folsomina sp. 1 8 1 1 7 5 23 0.61 鳞䖴属 Tomocerus sp. 6 1 13 20 0.53 原䖴属 Protanura sp. 10 1 1 7 19 0.50 钩圆䖴属 Bourletiella sp. 12 2 4 18 0.48 短角䖴属 Neelus sp. 2 11 13 0.34 鳞长䖴属Lepidocyrtus sp. 4 2 5 11 0.29 拟裸长角䖴属 Pseudosinella sp. 6 1 3 10 0.26 刺䖴属 Acanthocyrtus sp. 6 1 1 2 10 0.26 裔符䖴属Folsomides sp. 7 2 9 0.24 羽圆䖴属 Dicyrtoma sp. 5 1 1 1 8 0.21 爪䖴属Paronella sp. 4 3 7 0.18 铲圆䖴属Papirinus sp. 5 1 6 0.16 奇䖴属 Xenylla sp. 1 3 1 5 0.13 裸长角䖴属 Sinella sp. 3 3 0.08 柳䖴属 Willowsia sp. 1 1 0.03 个体数合计Total of Individual Number 335 1 252 230 807 215 946 3 785 类群数合计Total of Genus Number 14 28 14 26 13 23 29 注Note:Ⅰ:青冈林Forest of Fagus lucida,Ⅱ:光皮桦林Forest of Betula luminifera,Ⅲ:栲树林Forest of Castanopsis fargesii。
下载: 导出CSV
表 3 不同生境和季节的土壤跳虫群落多样性
Table 3. Diversity of soil Collembolan community in different habitats and seasons
生境
Habitats季节
Seasons类群数
Group number个体数
Individual number多样性(H′)
Shannon-Wiener index均匀度(Js)
Pielou index丰富度(R)
Margalef index优势度(D)
Simpson indexⅠ 13.13±3.16 a 105.80±42.59 a 1.92±0.32 a 0.76±0.09 ab 2.60±0.65 a 0.19±0.07 b Ⅱ 9.93±2.46 b 69.13±31.6 b 1.71±0.33 a 0.76±0.10 ab 2.10±0.56 b 0.25±0.11 a Ⅲ 10.87±2.77 b 77.40±26.98 ab 1.85±0.27 a 0.78±0.05 ab 2.32±0.56 ab 0.21±0.05 ab Sp 13.67±3.09 A 81.07±33.05 AB 2.12±0.19 A 0.82±0.05 A 2.91±0.51 A 0.16±0.04 B Su 10.07±2.25 B 65.93±23.91 B 1.70±0.28 B 0.75±0.08 B 2.11±0.43 B 0.26±0.10 A Au 10.20±2.46 B 105.33±42.69 A 1.67±0.25 B 0.73±0.09 B 2.00±0.46 B 0.23±0.08 A 注:表中数据为平均值±标准误,n=15.同一栏相同小写字母表示生境间差异不显著,不同小写字母表示生境间差异显著(P<0.05)。同一栏相同大写字母表示季节间差异不显著,不同大写字母表示季节间差异显著(P<0.05)。下同。
Note: Data in the figures are mean±SE, n=15.The same small letters in the column indicate no significant difference between diffrent habitats, while different small letters indicate significant differences (P<0.05). The same capital letters in the column indicate no significant difference between diffrent seasons, while different capital letters indicate significant differences(P<0.05). The same below.
下载: 导出CSV
表 4 不同生境和季节变化对土壤跳虫群落多样性的影响
Table 4. Effect of different habitats and seasons on diversity of soil Collembolan community
项目Items DF 类群数
Group number个体数
Individual number多样性(H′)
Shannon-Wiener index均匀度(Js)
Pielou index丰富度(R)
Margalef index优势度(D)
Simpson indexF p F p F p F p F p F p 生境Habitats 2 7.01 0.00 5.73 0.00 2.81 0.73 0.31 0.74 4.56 0.02 4.00 0.03 季节Seasons 2 10.22 0.00 5.67 0.01 13.54 0.00 4.55 0.02 16.76 0.00 7.70 0.00 交互作用Interaction 4 0.46 0.76 0.77 0.55 0.36 0.84 0.55 0.70 0.73 0.58 0.63 0.64
下载: 导出CSV
表 5 不同生境土壤主要理化性质比较
Table 5. Chief soil properties in different habitats
生境Habitats SOM/(g·kg-1) TN/(g·kg-1) TP/(g·kg-1) TK/(g·kg-1) Ca/(g·kg-1) Mg/(g·kg-1) pH Ⅰ 127.06±17.98 a 6.29±2.14 a 6.49±2.02 a 14.94±2.01 a 1.19±0.93 a 0.15±0.03 a 3.89±0.45 a Ⅱ 63.27±7.80 b 3.38±0.41 b 5.42±0.92 a 25.31±0.82 b 0.22±0.19 b 0.23±0.07 b 4.89±0.15 b Ⅲ 96.65±28.47 c 4.37±0.88 b 6.44±0.39 a 23.78±2.27 b 0.47±0.51 ab 0.38±0.06 c 4.87±0.26 b 注:SOM:土壤有机质;TN:土壤全氮;TP:土壤全磷;TK:土壤全钾;Ca:土壤全钙;Mg:土壤全镁;pH:土壤pH值。
Note: SOM: Soil organic matter; TN: Soil total Nitrogen; TP: Soil total Phosphor; TK: Soil total Potassium; Ca:Soil total Calcium; Mg: Soil total Magnesium; pH: Soil pH.
下载: 导出CSV
表 6 土壤跳虫群落与土壤主要理化性质相关系数
Table 6. Pearson correlation coefficients between Collembolan community and soil properties
项目 SOM/(g·kg-1) TN/(g·kg-1) TP/(g·kg-1) TK/(g·kg-1) Ca/(g·kg-1) Mg/(g·kg-1) pH 类群数Genus 0.75** 0.83** 0.78** -0.56* 0.39 -0.21 -0.6* 个体数Individuals 0.67** 0.70** 0.69** -0.42 0.34 -0.12 -0.51 多样性Diversity 0.68** 0.685** 0.49 -0.63* 0.41 -0.33 -0.56* 均匀度Evenness -0.05 -0.18 -0.4 -0.14 0.08 -0.21 0.3 丰富度Richness 0.67** 0.79** 0.71** -0.56* 0.36 -0.25 -0.57* 优势度Dominance -0.57* -0.46 -0.26 0.55* -0.46 0.33 0.41 注Note:**P<0.01,*P<0.05。
下载: 导出CSV
-
[1] Wardle D A, Bardgett R D, Klironomos J N, et al. Ecological linkages between aboveground and belowground biota[J]. Science, 2004, 304(5677):1629-1633. doi: 10.1126/science.1094875 [2] Lavelle P, Decaens T, Aubert M, et al. Soil invertebrates and ecosystem services[J]. European Journal of Soil Biology, 2006, 42:S3-S15. doi: 10.1016/j.ejsobi.2006.10.002 [3] 夏磊, 吴福忠, 杨万勤, 等.川西亚高山森林凋落物分解初期土壤动物对红桦凋落叶质量损失的贡献[J].应用生态学报, 2012, 23(2):301-306. [4] Maaβ S, Caruso T, Rillig M C. Functional role of microarthropods in soil aggregation[J]. Pedobiologia, 2015, 58(2-3):59-63. doi: 10.1016/j.pedobi.2015.03.001 [5] 陈建秀, 麻智春, 严海娟, 等.跳虫在土壤生态系统中的作用[J].生物多样性, 2007, 15(2):154-61. doi: 10.3321/j.issn:1005-0094.2007.02.006 [6] 邵元虎, 张卫信, 刘胜杰, 等.土壤动物多样性及其生态功能[J].生态学报, 2015, 35(20):6614-6625. [7] Chang L, Wu H, Wu D, et al. Effect of tillage and farming management on Collembola in marsh soils[J]. Applied Soil Ecology, 2013, 64(64):112-117. [8] Muscolo A, Panuccio M R, Mallamaci C, et al. Biological indicators to assess short-term soil quality changes in forest ecosystems[J]. Ecological Indicators, 2014, 45(5):416-423. [9] Turnbull M S, Lindo Z. Combined effects of abiotic factors on Collembola communities reveal precipitation may act as a disturbance[J]. Soil Biology & Biochemistry, 2015, 82(82):36-43. [10] Filho L C I D O, Filho O K, Baretta D, et al. Collembola community structure as a tool to assess land use effects on soil quality[J]. Rev Bras Ciênc Solo, 2016, 40:e0150432. [11] Rieff G G, Natal-Da-Luz T, Sousa J P, et al. Collembolans and mites communities as a tool for assessing soil quality:effect of eucalyptus plantations on soil mesofauna biodiversity[J]. Current Science, 2016, 110(4):713-719. doi: 10.18520/cs/v110/i4/713-719 [12] 向昌国, 李文芳, 于德珍.八大公山自然保护区森林土壤动物群落多样性的初步研究[J].生物多样性, 2000, 8(3):304-306. doi: 10.3321/j.issn:1005-0094.2000.03.010 [13] 刘博学.湘西八大公山的土壤[J].中南林学院学报, 1983, 3(2):141-159. [14] 尹文英.中国亚热带土壤动物[M].北京:科学出版社, 1992, 1-95. [15] 尹文英.中国土壤动物检索图鉴[M].北京:科学出版社, 1998, 51-89, 437-475. [16] Bellinger P, Christiansen K, Janssens F. Checklist of the Collembola of the world[EB/OL]. http://www.collembola.org, 1996-2016. [17] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社, 1999, 1-163. [18] 马克平, 刘玉明.生物群落多样性的测度方法[J].生物多样性, 1994, 2(4):231-233. doi: 10.3321/j.issn:1005-0094.1994.04.009 [19] Kardol P, Cregger M A, Campany C E, et al. Soil ecosystem functioning under climate change:plant species and community effects[J]. Ecology, 2010, 91(3):767-781. doi: 10.1890/09-0135.1 [20] 吴鹏飞, 杨大星.若尔盖高寒草甸退化对中小型土壤动物群落的影响[J].生态学报, 2011, 31(13):3745-3757. [21] 李伟, 崔丽娟, 赵欣胜, 等.太湖岸带湿地土壤动物群落结构与多样性[J].生态学报, 2015, 35(4):944-955. [22] Bardgett R D. Causes and consequences of biological diversity in soil[J]. Zoology, 2002, 105(4):367-375. doi: 10.1078/0944-2006-00072 [23] Sayer E J, Sutcliffe L M E, Ross R I C, et al. Arthropod abundance and diversity in a lowland tropical forest floor in Panama:the role of habitat space vs. nutrient concentrations[J]. Biotropica, 2010, 42(2):194-200. doi: 10.1111/j.1744-7429.2009.00576.x [24] 袁志忠, A.N.Singh, 胡颖圆.添加凋落物对土壤跳虫群落的影响[J].土壤通报, 2014, 45(4):841-846. [25] 杨赵, 杨效东.哀牢山不同类型亚热带森林地表凋落物及土壤节肢动物群落特征[J].应用生态学报, 2011, 22(11):3011-3020. [26] Caravaca F, Ruess L. Arbuscular mycorrhizal fungi and their associated microbial community modulated by Collembola grazers in host plant free substrate[J]. Soil Biology & Biochemistry, 2014, 69(1):25-33. [27] Rusek J. Biodiversity of collembola and their functional role in the ecosystem[J]. Biodiversity and Conservation, 1998, 7(9):1207-1279. doi: 10.1023/A:1008887817883 [28] 柯欣, 赵立军, 尹文英.三种乔木落叶分解过程中跳虫群落结构的演替[J].昆虫学报, 2001, 44(2):221-226. doi: 10.3321/j.issn:0454-6296.2001.02.015 [29] 靳娅丽, 由文辉, 易兰, 等.天童森林生态系统凋落物层跳虫群落的生态学研究[J].生态环境学报, 2011, 20(2):241-247. doi: 10.3969/j.issn.1674-5906.2011.02.006 [30] Bandyopadhyaya I, Choudhuri D K, Ponge J F. Effects of some physical factors and agricultural practices on Collembola in a multiple cropping programme in West Bengal (India)[J]. European Journal of Soil Biology, 2002, 38(1):111-117. doi: 10.1016/S1164-5563(01)01114-1 [31] Sechi V, D'Annibale A, Ambus P, et al. Collembola feeding habits and niche specialization in agricultural grasslands of different composition[J]. Soil Biology & Biochemistry, 2014, 74(3):31-38. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 4186
- HTML全文浏览量: 1693
- PDF下载量: 29
- 被引次数: 0
