

-
截至目前,氮沉降对森林树木生物量的影响并没有统一结论,Braun等[1]发现氮添加能显著提高森林生物量,适量氮可增加光合作用中酶的活性,进而影响植物固碳速率[2];氮沉降还会通过改变凋落物产量来影响森林树木生物量,同纬度的兴安落叶松(Larix gmelinii (Rupr.) Kuzen.)凋落物产量随施氮浓度的增加先减少再增加[3],而胸径增长量则随氮沉降量增加而增加[4]。在Harvard森林施氮9年的针叶树生物量随氮素输入的增加而减少,阔叶树生物量随氮素输入的增加而增加[5]。Gough等[6]对火炬松(Pinus taeda Linn.)林的研究发现,土壤氮含量的增加会使针叶树根的生长量减少,同时会增加根呼吸的消耗[7]。McNulty等[11]提出施氮处理会抑制针叶树种的生长。当植物体内氮的积累量达到一定量时,植物叶片的光合速率就会下降[8-9];我国兴安落叶松在生长季内的树干呼吸会随着施氮量的增加而增加[10]。不同氮处理对树木的影响表现并不一致,可能是由于不同的气候条件[11]、植物种类[5, 12]和土壤养分背景值[13]等原因造成的。有研究表明,兴安落叶松林在大兴安岭各林型中,碳储量随林龄的增加量表现为最大[14-15],其对氮沉降如何响应,很大程度上可影响我国北方森林的碳库。
本研究主要探讨兴安落叶松生物量增长率对氮沉降是否存在响应,年际间是否存在差异;不同径级是否会影响兴安落叶松生物量增长率对氮沉降的响应,径级间是否存在差异。
-
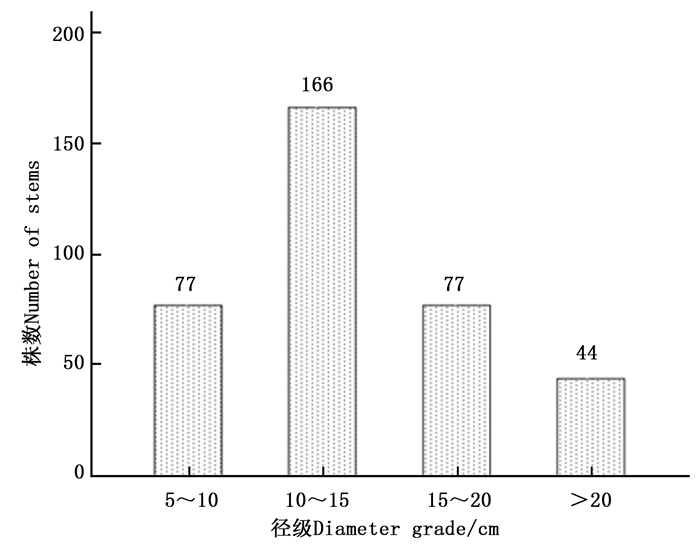
本试验氮添加控制样地位于黑龙江省大兴安岭松岭区南翁河国家级自然保护区(125°07′55″125°50′05″ E,51°05′07″51°39′24″ N)。该区冬季长达9个月,年平均气温-3℃,最低气温-48℃。夏季最长不超过1个月,最高气温36℃,年有效积温为1 4001 600℃,无霜期90100 d。初霜始于9月中旬,晚霜到翌年5月中旬,植物生长期为110 d左右,年降雨量500 mm左右。样地内土壤类型为暗棕色砂砾土[16],pH值为4.44.7,优势树种为兴安落叶松,所占比例近95%,林分密度(2 852 ± 99)株·hm-2,伴有零星白桦(Betula platyphylla Suk.)分布,兴安落叶松径级分布情况如图 1所示。
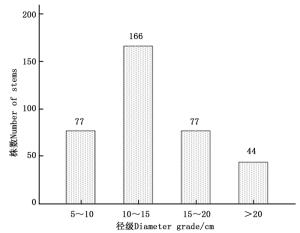
图 1 2013年4 800 m2样地中兴安落叶松径级分布
Figure 1. The diameter-class distribution of Larix gmelinii in 4 800 m2 sample plot in 2013
-
2011年10月在南翁河国家级自然保护区选择立地条件均一、坡向相同、坡度小于3°的兴安落叶松林,建立样地。共设立3块样地,每块样地均包括4块20 m×20 m的样方,各样方间有10 m的缓冲带阻隔。2012年5月开始,在各样地的4块样方中分别进行对照(CK,0 g·m-2·a-1)、低氮(TL,2.5 g·m-2·a-1)、中氮(TM,5 g·m-2·a-1)和高氮(TH,7.5 g·m-2·a-1)的施氮处理。样地划分主要参考了欧洲的NITREX项目[17]以及杜恩在等[18]在兴安落叶松林的试验设计。我国大兴安岭地区年氮沉降量约为2.5 g·m-2·a-1 [19-20],同时也是欧洲氮饱和临界值[21],5 g·m-2·a-1为净初级生产力达到峰值的最大值[22]。每年的生长季(5—10月)按月将NH4NO3溶于32 L蒸馏水,利用背喷式喷雾器均匀喷洒于样方内,对照样地喷洒等量纯净水。
-
于2011年10月在样地中所有胸径大于5 cm的树木上安装号牌及胸径测树器[4],并记录初始胸径及窗口大小,并在胸径测树器的下沿用红油漆标记。测树器稳定一年后,从2012开始每年10月测量胸径测树器的窗口大小并记录。为减小金属热胀冷缩带来的误差,测量工作必须在温度达到-5℃之前完成。每月施氮时需检查胸径测树器的工作情况,如有损坏及时更换。
-
本研究中所采用的生物量计算模型是Wang等[23]在研究中国温带森林不同树种的生物量模型中有关兴安落叶松的部分,公式为:
$ {\log _{10}}B = 2.021 + 2.474 \times \left( {{{\log }_{10}}DBH} \right) $
(1) 其中, B是生物量,DBH是胸径。
树木生物量年增长率如下式所示:
$ 生物量年增长率 = (当年生物量 - 上年生物量)/上年生物量 \times 100\% $
(2) 将兴安落叶松按照胸径划分为510 cm,1015 cm,1520 cm以及大于20 cm等4个径级。采用协方差分析胸径大小对生物量增长率的影响,用单因素方差分析得到其在不同氮处理间的差异。采用多因素方差分析计算各径级在不同氮添加水平和不同年份之间的差异。所有数据均采用EXCEL及SPSS(24.0)处理。
-
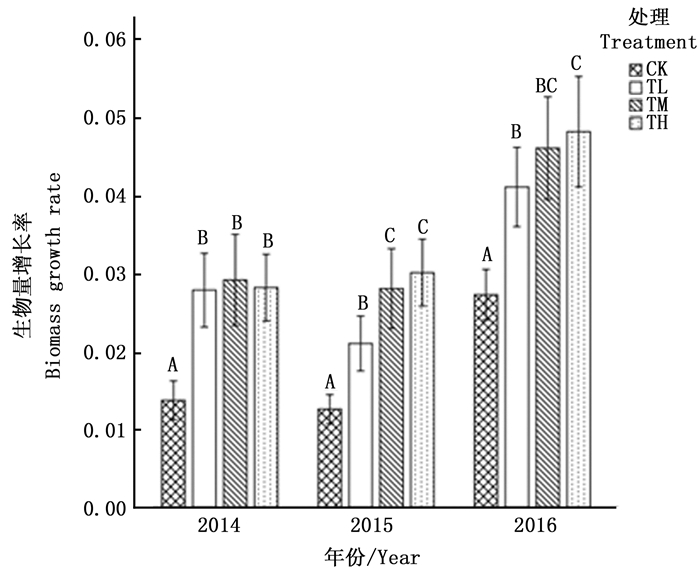
通过对4种处理以及不同年份进行的双因素方差分析可知,3年间各处理、各年份生物量的增长率均存在显著差异(P<0.001)(见表 1),而且2016年的增长率明显高于前两年(见图 2)。总体来说,在3年中不同氮处理对生物量的增长均存在显著的促进作用,但不同处理间存在差异。其中,2014年,TL、TM、TH三组间没有出现显著差异(P<0.05),其促进作用都高达100%;2015年,TL与TM、TH均存在显著差异,但TM和TH之间没有显著差异;2016年,TL与TH间存在显著差异,但TM与TL、TH两组均无差异。年际间各组对兴安落叶松生物量增长率的影响也存在差异。其中,3年间TL的促进作用逐渐减小,分别为102.9%、67.2%和50.3%;TM的促进作用呈现先增加后减小的趋势,分别为112.4%、124.0%和68.8%;TH的促进作用趋势与TM相同,分别为109.4%、138.4%和84.6%。
表 1 兴安落叶松生物量增长率与施氮浓度、年份的双因素方差分析
Table 1. Two-way ANOVA between Larix gmelinii biomass growth rate and nitrogen concentration and years
方差源
Sources of variationdf 均方
Mean squareF P 年份Years 2 333.421 75.471 <0.001 处理Treatments 3 188.029 42.561 <0.001 年份*处理Years*Treatments 6 4.343 0.983 0.435 
图 2 2014—2016年各处理生物量平均增长率
Figure 2. Percentage of average biomass growth rate of every treatment during 2014—2016
-
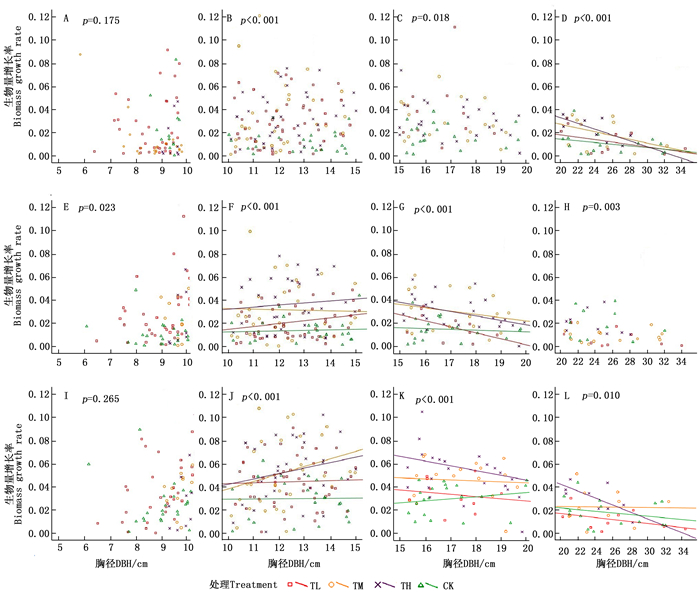
5~10 cm径级的兴安落叶松生物量增长率与胸径在3年中均无显著相关关系(图 3)。生物量的相对增长率在这一径级对氮沉降的响应无明显规律,其中2014和2016年响应不显著,2015年则表现为显著正相关(P=0.175,图 3-A;P=0.023,图 3-E;P=0.265,图 3-I)。TL处理的增长率在2015和2016年与CK差异显著,比CK分别增加了154%和42.2%;TM处理的增长率在2015年与CK差异显著,比CK增长了179%(表 2)。
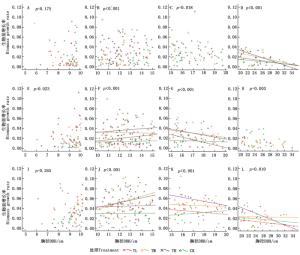
图 3 2014—2016年各处理生物量增长率随胸径的变化
Figure 3. The increase rate of biomass and change with DBH during 2014—2016
表 2 2014—2016各处理生物量平均增长率(平均值±标准误差)
Table 2. Average biomass growth rate of every treatment during 2014—2016(Mean±SE)
% 处理
Treatment年份
Year径级diameter-class/cm 5~10 10~15 15~20 20 CK 2014 1.24±0.40Aa 1.45±0.17Aa 1.63±0.22Aa 0.92±0.25Aa 2015 1.00±0.23 Aa 1.34±0.15Aa 1.53±0.20Aa 0.92±0.14Aa 2016 2.63±0.44 Ab 2.80±0.25Ab 3.16±0.27Ab 1.92±0.35Ab TL 2014 2.59±0.43 Aa 3.03±0.35Ba 3.48±0.74Ba 1.03±0.34Aa 2015 2.54±0.36 Aa 2.07±0.21Ab 1.68±0.28Ab 0.71±0.26Aa 2016 3.74±0.56 Aa 4.89±0.34Bc 3.60±0.34ABb 1.11±0.31ABa TM 2014 2.43±0.67 Aa 3.39±0.54Ba 2.99±0.41Ba 1.88±0.40ABa 2015 2.79±0.69 Aa 2.98±0.43Ba 3.03±0.43Ba 1.88±0.40Ba 2016 3.21±0.62 Aa 5.38±0.54Bb 4.80±0.43BCb 2.75±0.33ABa TH 2014 1.57±0.84 Aa 3.22±0.37Ba 3.00±0.29Ba 2.23±0.33Ba 2015 1.75±0.84 Aa 3.44±0.37Ba 3.05±0.28Ba 1.99±0.31Ba 2016 2.31±0.87 Aa 5.38±0.44Bb 5.74±0.60Cb 3.30±0.47Ba A、B、C为相同年份中各处理与生物量平均增长率的多重比较显著性水平(P<0.05);a、b为相同处理在不同年份与生物量平均增长率的多重比较显著性水平(P<0.05)。
A, B and C represent the significant level of multiple comparisons of average biomass growth rate under different N treatments in each year(P<0.05); a and b represent the significant level of multiple comparisons of mean average biomass growth rate in different years of each N treatments (P<0.05).10~15 cm径级的兴安落叶松,生物量增长率与胸径在2014年无显著相关,但在2015和2016年表现为显著正相关(P<0.05,图 3-F;P<0.05,图 3-J),各处理间也表现为极显著差异(P<0.001,图 3-B;P<0.001,图 3-F;P<0.001,图 3-J)。除TL处理的增长率在2015年与CK没有显著差异外,3年中TL、TM和TH处理对增长率均表现出了显著的促进作用(P<0.05)(表 2)。
15~20 cm径级的兴安落叶松,生物量增长率与胸径在2014年无显著相关,但在2015和2016年均表现为显著负相关(P<0.05,图 3-G;P<0.05,图 3-K),各处理间的增长率均存在显著的差异(P=0.018,图 3-C;P<0.001,图 3-G;P<0.001,图 3-K)。TM和TH处理3年增长率均显著促进兴安落叶松生长(P<0.05);2014和2015年TL与TM处理的增长率差异显著(P<0.05),但在2016年没有显著差异(表 2)。
大于20 cm径级的兴安落叶松,生物量增长率与胸径在2015年无显著相关,但在2014和2016年表现为显著负相关(P<0.05,图 3-D;P<0.05,图 3-L),3年各处理间增长率均存在显著差异(P<0.001,图 3-D;P=0.003,图 3-H;P=0.010,图 3-L)。TH处理3年增长率均与CK存在显著差异(P<0.05);TL处理3年增长率均无显著响应;TM处理的增长率在2015年与CK有显著差异(P<0.05),但在2014和2016年差异不显著(表 2)。
-
2016年各处理的生物量增长率都高于前两年,这有可能是当年的降水和温度等气象因素所致。有研究表明树木径向生长对温度存在“滞后效应”。如,刘欣[24]在研究大兴安岭不同区域兴安落叶松树木生长时发现,与本试验样地最接近的奥克里堆山树木的生长与上一年秋季的温度成正相关[21]。本试验样地在2013—2015年中8—10月的平均气温分别为6.3℃、10℃和11℃,这可能是因为温暖的秋季可以使树木能够更多的积累光合作用产物用于过冬;同时秋季也是兴安落叶松产生休眠芽的时期,较高的温度有利于树木产生休眠芽,从而影响第二年树木的生长[25]。也有研究指出上一年冬季直至当年生长季之前的温度也会对树木的生长有影响,试验样地所在地区上一年12月到当年4月的平均气温分别为-15.4℃、-13.7℃和-14.5℃,这一时期的温度较低会减慢树木的呼吸速率,从而使得生长季初期能有更多的养分参与到下一年的固碳过程中。受二者共同影响,导致2016年各处理生物量增长率均高于前两年。
整体上看,3种水平的氮添加相对于对照组均促进了兴安落叶松生物量的相对增长率的增长。在我国小兴安岭的兴安落叶松林模拟氮沉降的研究结果,也出现了树木胸径生长量随氮添加的增加而增加的现象[4]。
3种施氮处理在2014年无显著差异;2015年TM、TH显著高于TL;2016年TH显著高于TL,这可能与我国兴安落叶松的氮耐受性有关。Vries等[22]在研究森林生态系统固碳速率对氮沉降的响应时指出,森林生态系统净初级生产力在添加2~5 g·m-2·a-1时达到峰值,在达到10~15 g·m-2·a-1时净初级生产力消失为0。本试验中设置的3个施氮梯度中TH处理的氮添加量要高于Vries等[22]所提出的初级净生产力达到峰值时的氮添加量,但其生物量的增长率仍然要比更接近峰值的TL、TM两个处理高。刘修元[4]在研究兴安落叶松原始林胸径随氮沉降变化时出现的情况与本试验相似,兴安落叶松胸径增长率在输入10 g·m-2·a-1时比输入2和5 g·m-2·a-1高。这说明在我国东北地区北方森林生态系统的各组分中,兴安落叶松是氮限制型,其最适的氮添加浓度有可能远高于2~5 g·m-2·a-1。2014年3个水平氮处理均显著增加树木生物量的增长率,但3种处理间没有显著差异,这可能是施氮早期森林群落对其响应机制较为明显所导致[26]。值得注意的是,2015年TH处理的平均生物量增长率显著高于TM处理,2016年TH处理的平均生物量增长率仍然高于TM处理,但没有达到显著差异水平,根据Aber[27]提出的氮素积累假说可以推测,随着施氮年份的增加氮素逐渐在植物体中累积,从而导致TL、TM、TH三种施氮处理均无显著的差异。更进一步,甚至同小兴安岭云冷杉红松林中红松(Pinus koraiensis Sieb. et Zucc.)和臭冷杉(Abies nephrolepis (Trautv.) Maxim.)一样,出现TH处理显著抑制植物生长的结果[28]。
-
5~10 cm径级,由于属径级较小的树木,株高比较矮,其生长速率受光照的影响较大,从而使得仅有TL处理在2015年对兴安落叶松生物量的增长率表现为显著影响。
10~15 cm和15~20 cm径级,在3年中TM、TH处理对生物量增长率均出现了显著的促进作用,这说明此径级兴安落叶松是氮限制类型,外源无机氮的增加使其生物量的增长率加大。能出现如此显著的差异可能与大兴安岭地区频繁遭遇火烧有一定关系,有研究表明,大兴安岭地区不同程度的火烧均会导致全氮的质量分数下降,这是因为火烧后有一部分氮素直接气化挥发出去;同时,火烧时的高温会导致土壤中微生物数量和种类的减少,火烧后微生物生境的改变会进一步使其数量和种类减少,从而使得微生物的固氮能力和氮矿化能力降低[29]。也有研究表明,北方森林普遍受到氮的限制[30],其原因可能是低温导致土壤中微生物固氮能力较弱。
在胸径大于20 cm的径级中,仅有TH处理对生物量增长率在3年中均有显著的促进作用,2015年开始TH与TM处理均没有显著的差异,到2016年TH与TL处理的差异也表现为不显著,这也印证了Aber[27]提出的氮素积累假说,长期的外源氮添加会在土壤和植物体中累积,最终对植物的生长产生影响。此径级中TL、TM处理的生物量增长率的平均值都要高于CK,但差异不显著,可能是因为大径级的树木生长需要更多的氮,而这两个处理所能提供的氮并不能解除大径级兴安落叶松的氮限制。3年中无外源氮添加的对照组生物量增长率表现为大径级低于较小径级,这说明大径级的树木趋于成熟,生长速度已经开始下降,外源氮添加有可能使植物各器官的呼吸量加大[6, 9],二者共同作用将进一步导致碳积累速率下降。
-
我国大兴安岭地区的兴安落叶松大径级树木的生长对短期氮添加表现为较高浓度氮处理(TH)的响应明显且表现为显著促进,中、低浓度的氮添加(TL、TM)在较小径级的树木中显著促进,而在较大径级的树木中无显著影响。氮素会在植物体中积累,随着氮添加年份的增加,中、低浓度的氮添加(TL、TM)也可能在大径级的树木中表现为促进生长。
2016年最大径级的树木出现了中、高浓度施氮(TM、TH)与低浓度施氮(TL)差异不显著现象,这有可能是施氮的浓度超过了植物的承受极限,植物各器官的呼吸速率随氮素增加而增加,而光合速率没有明显增加,使得碳积累率减少。同时,也有可能是植物各器官的呼吸速率随氮素增加而增加导致了植物的提前衰亡。因此,后续研究应将重心放在植物各生理过程对外源氮添加的响应机制上。
大兴安岭兴安落叶松林树木生物量对氮沉降的响应
Response of Tree Biomass to Nitrogen Deposition in Larch Forest in Greater Khingan Mountains
-
摘要:
目的 研究兴安落叶松生物量增长率对氮沉降是否存在响应,年际间是否存在差异;不同径级是否会影响兴安落叶松生物量增长率对氮沉降的响应。 方法 通过人工氮添加的方法模拟大气氮沉降变化,探究兴安落叶松林连续3年(2014—2016)树木生物量的变化对氮沉降的响应。从2012年开始,共设置4个氮添加水平,分别为对照(CK,0 g·m-2·a-1)、低氮(TL,2.5 g·m-2·a-1)、中氮(TM,5 g·m-2·a-1)和高氮(TH,7.5 g·m-2·a-1),每年的生长季(5—10月)按月将NH4NO3溶于32 L蒸馏水,利用背喷式喷雾器均匀喷洒于样方内,对照样地喷洒等量纯净水。 结果 (1)3年间各施氮处理生物量增长率均显著高于对照组(P < 0.05)、各年份生物量增长率均存在显著差异(P < 0.05),而且2016年生物量增长率明显高于前两年(P < 0.05)。(2)2014年各处理间无显著差异;2015年TL处理的生物量增长率显著高于TM和TH处理(P < 0.05),2016年TL处理的生物量增长率显著高于TH处理(P < 0.05)。(3)不同处理对年际间不同径级生物量增长率也产生不同影响。TL处理对2014和2016年的10 15 cm径级的兴安落叶松生物量增长率有显著促进作用(P < 0.05);TM处理对所有年份10 15 cm和15 20 cm径级兴安落叶松生物量增长率均有显著促进作用(P < 0.05);TH处理对所有年份胸径大于10 cm的3个径级(10 15、15 20、> 20 cm)兴安落叶松生物量增长率均有显著促进作用(P < 0.05)。 结论 (1)长期氮添加有可能导致树木生长率的增量逐渐减少甚至出现抑制,将改变现有北方森林生态系统碳源汇动态;(2)由于不同径级树木对氮沉降的响应不同,为提高氮沉降对北方森林碳库影响的模型预测精度,应按不同径级分别模拟。 Abstract:Objective To study whether there is a response of biomass growth rate of Larix gmelinii forest to nitrogen deposition, whether there is difference among years and whether different diameters can affect the response. Method This study simulated the change of atmospheric nitrogen deposition by artificial nitrogen addition method to investigate the response of biomass growth in three years to nitrogen deposition in L. gmelinii forests during the period of 2014-2016. The nitrogen was added from 2012, 4 nitrogen addition levels were set as control (CK, 0 g·m-2·a-1), low nitrogen (TL, 2.5 g·m-2·a-1), middle nitrogen (TM, 5 g·m-2·a-1) and high nitrogen (TH, 7.5 g·m-2·a-1). The growth season (May-October) will dissolve NH4NO3 in 32 L distilled water monthly, spray evenly on the sample plot with back atomize, and spray the same amount of pure water in the same way on the CK. Result (1) The biomass growth rate of every treatment was significantly higher in three years (P < 0.05). The annual biomass growth rate was significantly different (P < 0.05), and the growth rate of 2016 was significantly higher than that of the previous two years (P < 0.05). (2) There was no significant difference between the three treatments in 2014 (P < 0.05). The growth rate of TL in 2015 was significantly higher than that of TM and TH(P < 0.05). The growth rate of TL in 2016 was significantly higher than that of TH (P < 0.05). (3) Different treatments had different effects on different diameter class. Between 2014 and 2015, TL significant increased the biomass growth rate of L. gmelinii (P < 0.05) in 10 15 cm diameter. TM had a significant effect on the biomass growth rate of L. gmelinii of 10 15 cm and 15 20 cm diameter in all the 3 years (P < 0.05). In 2016, the biomass growth rate of L. gmelinii significantly increased (P < 0.05) in the 15 20 cm diameter. TH had more significant effect on the biomass growth rate of three diameters class (10 15, 15 20, and > 20 cm) than that < 10 cm (P < 0.05) in all the 3 years. Conclusion (1) Long-term nitrogen addition will lead to a growth rate gradually reduce or even suppress, which is likely to change the boreal forest carbon source & sink dynamic; (2) Since different diameter trees have different responses to the nitrogen deposition, it should be as the basal factor in simulation in order to increase the prediction accuracy about nitrogen deposition on the boreal forest carbon pool. -
Key words:
- Larix gmelinii
- / biomass
- / diameter class
- / nitrogen deposition
- / response
-
图 1 2013年4 800 m2样地中兴安落叶松径级分布
Figure 1. The diameter-class distribution of Larix gmelinii in 4 800 m2 sample plot in 2013
图 2 2014—2016年各处理生物量平均增长率
Figure 2. Percentage of average biomass growth rate of every treatment during 2014—2016
图 3 2014—2016年各处理生物量增长率随胸径的变化
Figure 3. The increase rate of biomass and change with DBH during 2014—2016
表 1 兴安落叶松生物量增长率与施氮浓度、年份的双因素方差分析
Table 1. Two-way ANOVA between Larix gmelinii biomass growth rate and nitrogen concentration and years
方差源
Sources of variationdf 均方
Mean squareF P 年份Years 2 333.421 75.471 <0.001 处理Treatments 3 188.029 42.561 <0.001 年份*处理Years*Treatments 6 4.343 0.983 0.435  下载: 导出CSV
下载: 导出CSV
表 2 2014—2016各处理生物量平均增长率(平均值±标准误差)
Table 2. Average biomass growth rate of every treatment during 2014—2016(Mean±SE)
% 处理
Treatment年份
Year径级diameter-class/cm 5~10 10~15 15~20 20 CK 2014 1.24±0.40Aa 1.45±0.17Aa 1.63±0.22Aa 0.92±0.25Aa 2015 1.00±0.23 Aa 1.34±0.15Aa 1.53±0.20Aa 0.92±0.14Aa 2016 2.63±0.44 Ab 2.80±0.25Ab 3.16±0.27Ab 1.92±0.35Ab TL 2014 2.59±0.43 Aa 3.03±0.35Ba 3.48±0.74Ba 1.03±0.34Aa 2015 2.54±0.36 Aa 2.07±0.21Ab 1.68±0.28Ab 0.71±0.26Aa 2016 3.74±0.56 Aa 4.89±0.34Bc 3.60±0.34ABb 1.11±0.31ABa TM 2014 2.43±0.67 Aa 3.39±0.54Ba 2.99±0.41Ba 1.88±0.40ABa 2015 2.79±0.69 Aa 2.98±0.43Ba 3.03±0.43Ba 1.88±0.40Ba 2016 3.21±0.62 Aa 5.38±0.54Bb 4.80±0.43BCb 2.75±0.33ABa TH 2014 1.57±0.84 Aa 3.22±0.37Ba 3.00±0.29Ba 2.23±0.33Ba 2015 1.75±0.84 Aa 3.44±0.37Ba 3.05±0.28Ba 1.99±0.31Ba 2016 2.31±0.87 Aa 5.38±0.44Bb 5.74±0.60Cb 3.30±0.47Ba A、B、C为相同年份中各处理与生物量平均增长率的多重比较显著性水平(P<0.05);a、b为相同处理在不同年份与生物量平均增长率的多重比较显著性水平(P<0.05)。
A, B and C represent the significant level of multiple comparisons of average biomass growth rate under different N treatments in each year(P<0.05); a and b represent the significant level of multiple comparisons of mean average biomass growth rate in different years of each N treatments (P<0.05).
下载: 导出CSV
-
[1] Braun S, Thomas V F D, Quiring R, et al. Does nitrogen deposition increase forest production? The role of phosphorus[J]. Environmental Pollution, 2010, 158(6):2043-2052. doi: 10.1016/j.envpol.2009.11.030 [2] 李德军, 莫江明, 方运霆, 等.氮沉降对森林植物的影响[J].生态学报, 2003, 23(9):1891-1900. doi: 10.3321/j.issn:1000-0933.2003.09.022 [3] 赵鹏武, 舒洋, 张波, 等.兴安落叶松林凋落物量动态对模拟氮沉降的响应[J].内蒙古农业大学学报:自然科学版, 2016, 37(2):34-39. [4] 刘修元, 杜恩在, 徐龙超, 等.落叶松原始林树木生长对氮添加的响应[J].植物生态学报, 2015, 39(5):433-441. [5] Magill A H, Aber J D, Berntson G M, et al. Long-term nitrogen additions and nitrogen saturation in two temperate forests[J]. Ecosystems, 2000, 3(3):238-253. doi: 10.1007/s100210000023 [6] Gough C M, Seiler J R. Belowground carbon dynamics in loblolly pine (Pinus taeda) immediately following diammonium phosphate fertilization[J]. Tree Physiology, 2004, 24(7):845-851. doi: 10.1093/treephys/24.7.845 [7] Janssens I A, Dieleman W, Luyssaert S, et al. Reduction of forest soil respiration in response to nitrogen deposition[J]. Nature Geoscience, 2010, 3(5):315-322. doi: 10.1038/ngeo844 [8] McNulty S G, Aber J D, Newman S D. Nitrogen saturation in a high elevation New England spruce-fir stand[J]. Forest Ecology and Management, 1996, 84(1-3):109-121. doi: 10.1016/0378-1127(96)03742-5 [9] Nakaji T, Fukami M, Dokiya Y, et al. Effects of high nitrogen load on growth, photosynthesis and nutrient status of Cryptomeria japonica and Pinus densiflora seedlings[J]. Trees-Structure and Function, 2001, 15(8):453-461. [10] 孙涛, 刘瑞鹏, 李兴欢, 等.模拟氮沉降对东北地区兴安落叶松树干呼吸的影响[J].生态学报, 2015, 35(11):3684-3691 [11] Martinelli L A, Piccolo M C, Townsend A R, et al. Nitrogen stable isotopic composition of leaves and soil:tropical versus temperate forests[J]. Biogeochemistry, 1999, 46(1-3):45-65. doi: 10.1007/BF01007573 [12] Thomas R Q, Canham C D, Weathers K C, et al. Increased tree carbon storage in response to nitrogen deposition in the US[J]. Nature Geoscience, 2010, 3(1):13. [13] Vitousek P M, Howarth R W. Nitrogen limitation on land and in the sea:how can it occur?[J]. Biogeochemistry, 1991, 13(2):87-115. [14] 胡海清, 罗碧珍, 魏书精, 等.大兴安岭5种典型林型森林生物碳储量[J].生态学报, 2015, 35(17):5745-5760. [15] 石磊, 盛后财, 满秀玲, 等.兴安落叶松林降雨再分配及其穿透雨的空间异质性[J].南京林业大学学报:自然科学版, 2017, 41(2):90-96. [16] 闫国永, 王晓春, 邢亚娟, 等.兴安落叶松林细根解剖结构和化学组分对N沉降的响应[J].北京林业大学学报, 2016, 38(4):36-43. [17] Wright R F, Roelofs J G M, Bredemeier M, et al. NITREX:responses of coniferous forest ecosystems to experimentally changed deposition of nitrogen[J]. Forest Ecology and Management, 1995, 71(1-2):163-169. doi: 10.1016/0378-1127(94)06093-X [18] 杜恩在.氮添加对兴安落叶松原始林碳收支主要过程的影响[D].北京: 北京大学, 2013. [19] 顾峰雪, 黄玫, 张远东, 等. 1961-2010年中国区域氮沉降时空格局模拟研究[J].生态学报, 2016, 36(12):3591-3600. [20] Sheng W, Yu G, Jiang C, et al. Monitoring nitrogen deposition in typical forest ecosystems along a large transect in China[J]. Environmental Monitoring and Assessment, 2013, 185(1):833-844. doi: 10.1007/s10661-012-2594-0 [21] 江泽平.欧洲森林生态系统的氮循环研究近况[J].世界林业研究, 1997, 10(5):55-61. [22] De Vries W, Du E, Butterbach-Bahl K. Short and long-term impacts of nitrogen deposition on carbon sequestration by forest ecosystems[J]. Current Opinion in Environmental Sustainability, 2014, 9:90-104. [23] Wang C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J]. Forest Ecology and Management, 2006, 222(1):9-16. [24] 刘欣.不同坡向兴安落叶松径向生长对气候变化响应[D].哈尔滨: 东北林业大学, 2016. [25] 勾晓华, 陈发虎, 杨梅学, 等.青藏高原东北部树木年轮记录揭示的最高最低温的非对称变化[J].中国科学(D辑:地球科学), 2007, (11):1480-1492. [26] 刘博奇, 牟长城, 邢亚娟, 等.模拟氮沉降对云冷杉红松林土壤呼吸的影响[J].林业科学研究, 2012, 25(6):767-772. doi: 10.3969/j.issn.1001-1498.2012.06.015 [27] Aber J, McDowell W, Nadelhoffer K, et al. Nitrogen saturation in temperate forest ecosystems[J]. BioScience, 1998, 48(11):921-934. doi: 10.2307/1313296 [28] 王建宇, 王庆贵, 闫国永, 等.原始云冷杉、红松林树木生长对氮沉降的响应[J].北京林业大学学报, 2017, 39(4):21-28. [29] 郭爱雪, 郭亚芬, 崔晓阳.大兴安岭马尾松林下土壤在不同火烧强度下的养分变化[J].东北林业大学学报, 2011, 39(5):69-71. doi: 10.3969/j.issn.1000-5382.2011.05.022 [30] LeBauer D S, Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed[J]. Ecology, 2008, 89(2):371-379. doi: 10.1890/06-2057.1 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3787
- HTML全文浏览量: 1605
- PDF下载量: 29
- 被引次数: 0
